
Role of Glycated High Mobility Group Box-1 in Gastric Cancer

 ,
,
Abstract
:1. Introduction
2. Results
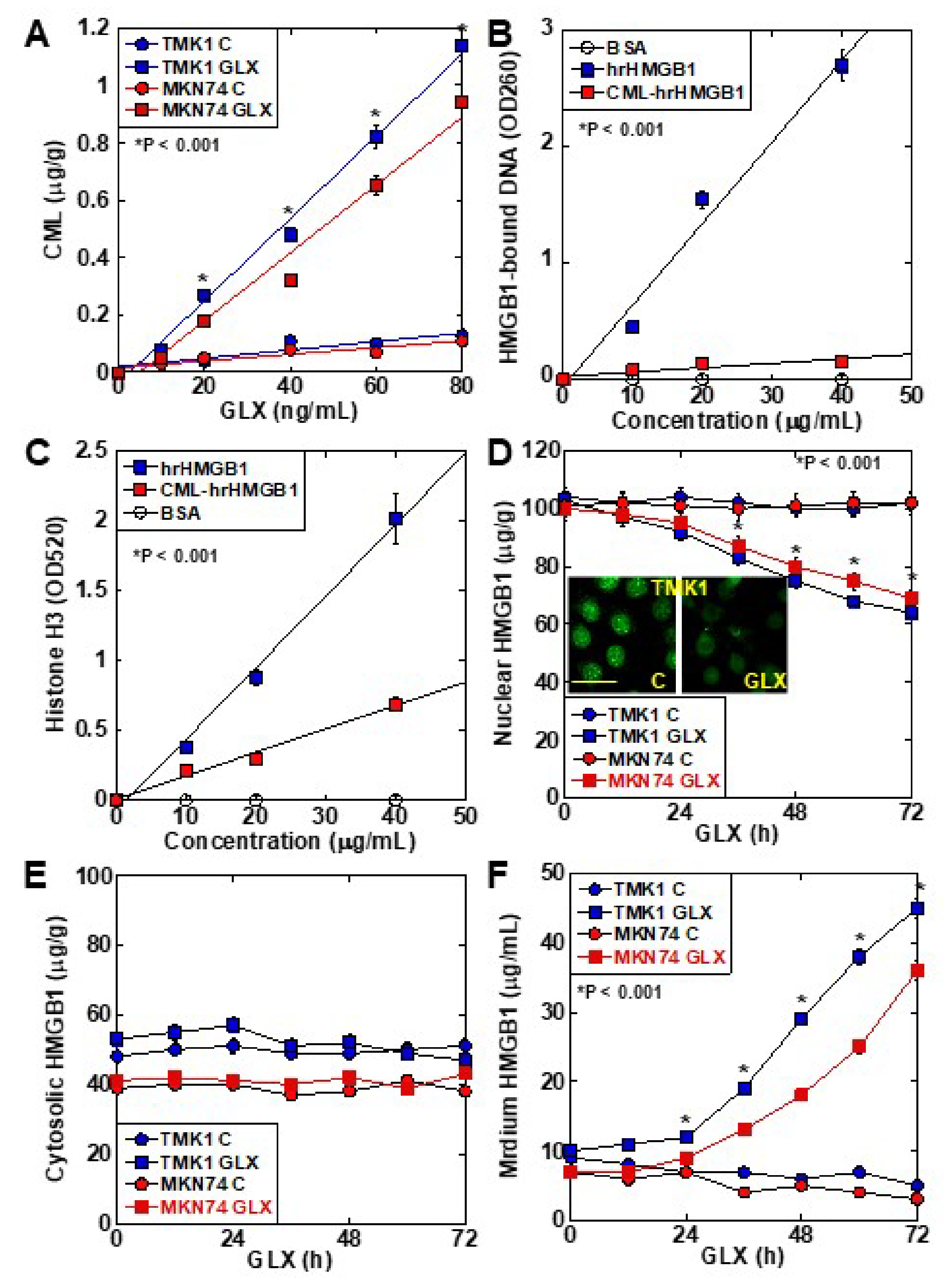
2.1. CML Modification of Cancer-Related Proteins in Human Gastric Cancer Cell Lines
2.2. CML Formation and HMGB1 Secretion in Human Gastric Cancer Cell Lines
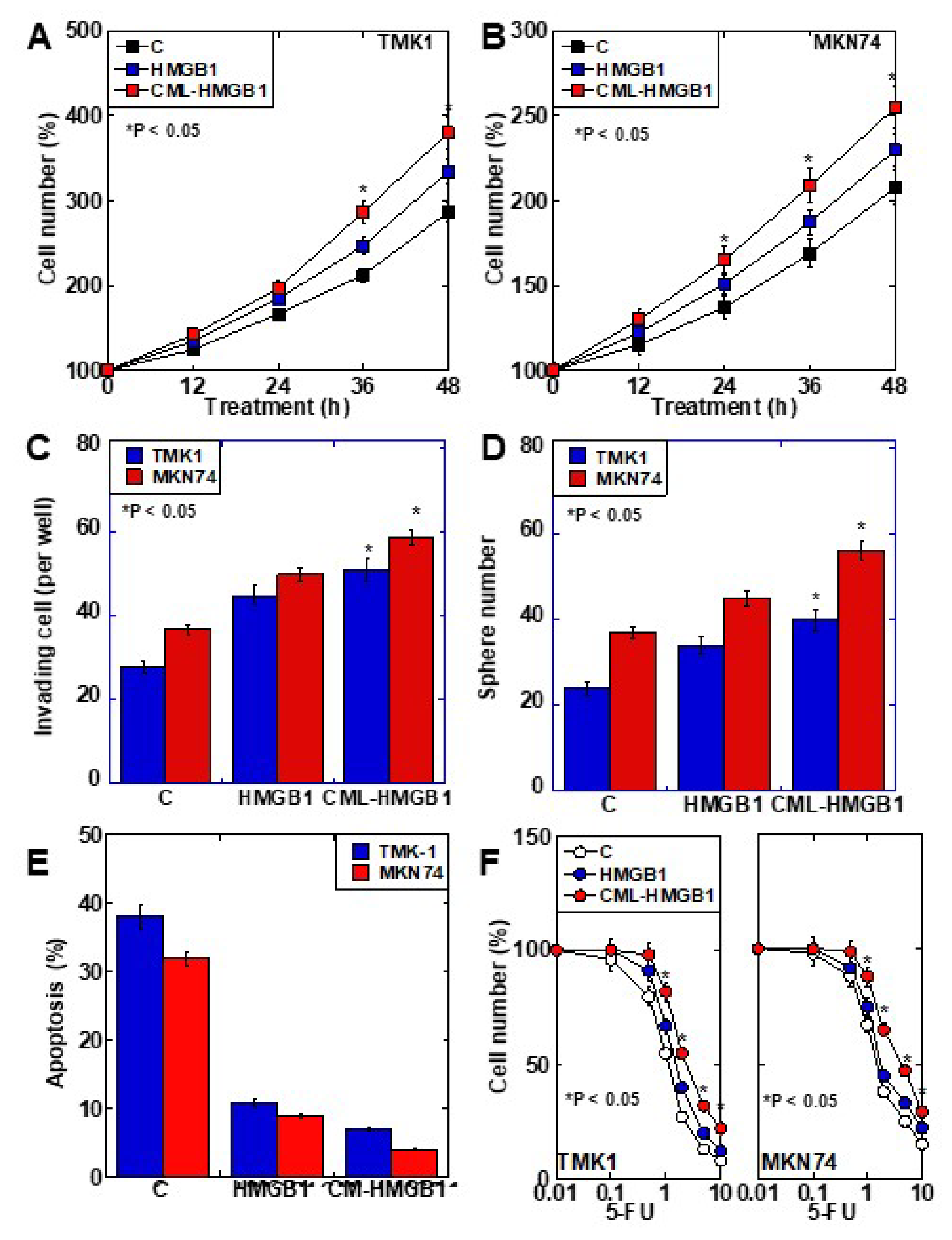
2.3. Role of CML-HMGB1 in Gastric Cancer Cells
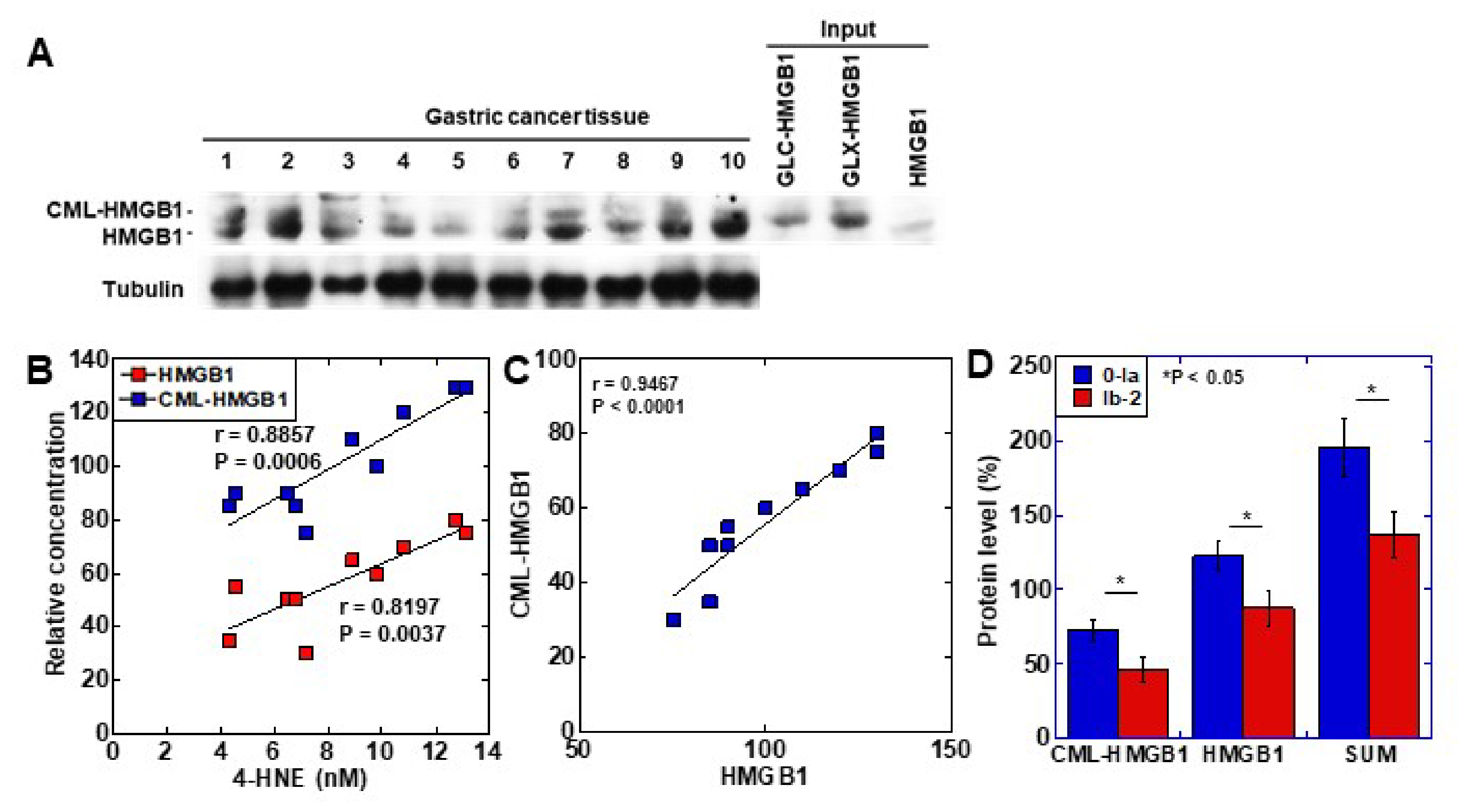
2.4. Clinical Significance of CML-HMGB1
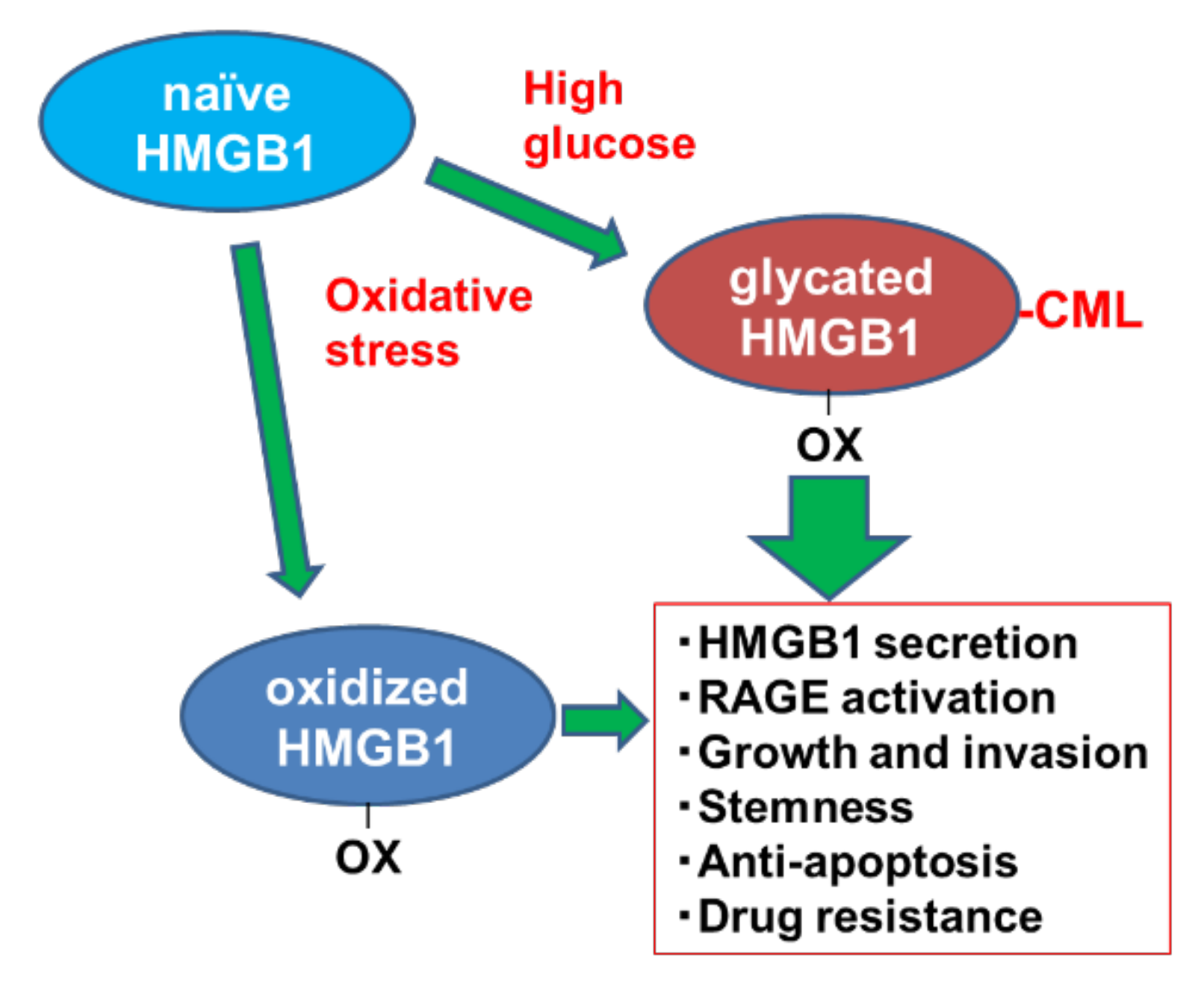
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Reagents
4.2. Patients
4.3. Glycation and Oxidization of HMGB1
4.4. Cell Growth and Apoptosis
4.5. Chamber Invasion Assay
4.6. Sphere Formation Assay
4.7. Protein Extraction
4.8. Immunoprecipitation
4.9. Immunoblot Analysis
4.10. Protein Antibody Array
4.11. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.12. Enzyme-Linked Immunosorbent Assay (ELISA) and Fluorometric Assay
4.13. HMGB1-Bound DNA
4.14. Histone Binding Ability of HMGB1
4.15. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AGE | Advanced glycation end products |
| CML | Nε-(Carboxymethyl)lysine |
| HMGB1 | high mobility group box protein-1 |
| RAGE | receptor for advanced glycation end products |
| NF | nuclear factor |
| GLX | glyoxal |
| GLC | glucose |
| SOD | superoxide dismutase |
References
- Statistics Bureau Ministry of Internal Affairs and Communications Japan. Statistical Handbook of Japan 2017; Statistics Bureau Ministry of Internal Affairs and Communications Japan: Tokyo, Japan, 2017.
- Mikami, H.; Nagase, H. Cancer Survival Rates at Japanese Association of Clinical Cancer Centers. Available online: https://kapweb.chiba-cancer-registry.org/ (accessed on 12 May 2021).
- Taguchi, A.; Blood, D.C.; del Toro, G.; Canet, A.; Lee, D.C.; Qu, W.; Tanji, N.; Lu, Y.; Lalla, E.; Fu, C.; et al. Blockade of RAGE-amphoterin signalling suppresses tumour growth and metastases. Nature 2000, 405, 354–360. [Google Scholar] [CrossRef]
- Kuniyasu, H.; Oue, N.; Wakikawa, A.; Shigeishi, H.; Matsutani, N.; Kuraoka, K.; Ito, R.; Yokozaki, H.; Yasui, W. Expression of receptors for advanced glycation end-products (RAGE) is closely associated with the invasive and metastatic activity of gastric cancer. J. Pathol. 2002, 196, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Kuniyasu, H.; Chihara, Y.; Kondo, H. Differential effects between amphoterin and advanced glycation end products on colon cancer cells. Int. J. Cancer 2003, 104, 722–727. [Google Scholar] [CrossRef] [PubMed]
- Kuniyasu, H.; Chihara, Y.; Kondo, H.; Ohmori, H.; Ukai, R. Amphoterin induction in prostatic stromal cells by androgen deprivation is associated with metastatic prostate cancer. Oncol. Rep. 2003, 10, 1863–1868. [Google Scholar] [CrossRef]
- Luo, Y.; Yoneda, J.; Ohmori, H.; Sasaki, T.; Shimbo, K.; Eto, S.; Kato, Y.; Miyano, H.; Kobayashi, T.; Sasahira, T.; et al. Cancer usurps skeletal muscle as an energy repository. Cancer Res. 2014, 74, 330–340. [Google Scholar] [CrossRef] [Green Version]
- Kuniyasu, H.; Chihara, Y.; Takahashi, T. Co-expression of receptor for advanced glycation end products and the ligand amphoterin associates closely with metastasis of colorectal cancer. Oncol. Rep. 2003, 10, 445–448. [Google Scholar] [CrossRef] [PubMed]
- Sasahira, T.; Kirita, T.; Oue, N.; Bhawal, U.K.; Yamamoto, K.; Fujii, K.; Ohmori, H.; Luo, Y.; Yasui, W.; Bosserhoff, A.K.; et al. High mobility group box-1-inducible melanoma inhibitory activity is associated with nodal metastasis and lymphangiogenesis in oral squamous cell carcinoma. Cancer Sci. 2008, 99, 1806–1812. [Google Scholar] [CrossRef]
- Ohmori, H.; Luo, Y.; Kuniyasu, H. Non-histone nuclear factor HMGB1 as a therapeutic target in colorectal cancer. Expert Opin. Ther. Targets 2011, 15, 183–193. [Google Scholar] [CrossRef]
- Schmidt, A.M.; Hori, O.; Cao, R.; Yan, S.D.; Brett, J.; Wautier, J.L.; Ogawa, S.; Kuwabara, K.; Matsumoto, M.; Stern, D. RAGE: A novel cellular receptor for advanced glycation end products. Diabetes 1996, 45, S77–S80. [Google Scholar] [CrossRef] [PubMed]
- Azizian-Farsani, F.; Abedpoor, N.; Hasan Sheikhha, M.; Gure, A.O.; Nasr-Esfahani, M.H.; Ghaedi, K. Receptor for Advanced Glycation End Products Acts as a Fuel to Colorectal Cancer Development. Front. Oncol. 2020, 10, 552283. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.M.; Stern, D.M. RAGE: A new target for the prevention and treatment of the vascular and inflammatory complications of diabetes. Trends Endocrinol. Metab. 2000, 11, 368–375. [Google Scholar] [CrossRef]
- Yamagishi, S.; Takeuchi, M.; Inagaki, Y.; Nakamura, K.; Imaizumi, T. Role of advanced glycation end products (AGEs) and their receptor (RAGE) in the pathogenesis of diabetic microangiopathy. Int. J. Clin. Pharmacol. Res. 2003, 23, 129–134. [Google Scholar]
- Wautier, J.L.; Guillausseau, P.J. Diabetes, advanced glycation endproducts and vascular disease. Vasc. Med. 1998, 3, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Shimomoto, T.; Luo, Y.; Ohmori, H.; Chihara, Y.; Fujii, K.; Sasahira, T.; Denda, A.; Kuniyasu, H. Advanced glycation end products (AGE) induce the receptor for AGE in the colonic mucosa of azoxymethane-injected Fischer 344 rats fed with a high-linoleic acid and high-glucose diet. J. Gastroenterol. 2012, 47, 1073–1083. [Google Scholar] [CrossRef]
- Shimomoto, T.; Ohmori, H.; Luo, Y.; Chihara, Y.; Denda, A.; Sasahira, T.; Tatsumoto, N.; Fujii, K.; Kuniyasu, H. Diabetes-associated angiotensin activation enhances liver metastasis of colon cancer. Clin. Exp. Metastasis 2012, 29, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Omofuma, O.O.; Turner, D.P.; Peterson, L.L.; Merchant, A.T.; Zhang, J.; Steck, S.E. Dietary Advanced Glycation End-products (AGE) and Risk of Breast Cancer in the Prostate, Lung, Colorectal and Ovarian Cancer Screening Trial (PLCO). Cancer Prev. Res. 2020, 13, 601–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado-Andrade, C. Carboxymethyl-lysine: Thirty years of investigation in the field of AGE formation. Food Funct. 2016, 7, 46–57. [Google Scholar] [CrossRef]
- Cho, S.J.; Roman, G.; Yeboah, F.; Konishi, Y. The road to advanced glycation end products: A mechanistic perspective. Curr. Med. Chem. 2007, 14, 1653–1671. [Google Scholar] [CrossRef]
- Ikeda, K.; Higashi, T.; Sano, H.; Jinnouchi, Y.; Yoshida, M.; Araki, T.; Ueda, S.; Horiuchi, S. Nε-(carboxymethyl)lysine protein adduct is a major immunological epitope in proteins modified with advanced glycation end products of the Maillard reaction. Biochemistry 1996, 35, 8075–8083. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.E.; Ponces Freire, A.M.; Voit, E.O. A quantitative model of the generation of Nε-(carboxymethyl)lysine in the Maillard reaction between collagen and glucose. Biochem. J. 2003, 376, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Bartling, B.; Desole, M.; Rohrbach, S.; Silber, R.E.; Simm, A. Age-associated changes of extracellular matrix collagen impair lung cancer cell migration. FASEB J. 2009, 23, 1510–1520. [Google Scholar] [CrossRef] [PubMed]
- Hsia, T.C.; Yin, M.C.; Mong, M.C. Advanced Glycation End-Products Enhance Lung Cancer Cell Invasion and Migration. Int. J. Mol. Sci. 2017, 18, 1642. [Google Scholar] [CrossRef] [Green Version]
- Menini, S.; Iacobini, C.; de Latouliere, L.; Manni, I.; Ionta, V.; Blasetti Fantauzzi, C.; Pesce, C.; Cappello, P.; Novelli, F.; Piaggio, G.; et al. The advanced glycation end-product Nε -carboxymethyllysine promotes progression of pancreatic cancer: Implications for diabetes-associated risk and its prevention. J. Pathol. 2018, 245, 197–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nass, N.; Ignatov, A.; Andreas, L.; Weißenborn, C.; Kalinski, T.; Sel, S. Accumulation of the advanced glycation end product carboxymethyl lysine in breast cancer is positively associated with estrogen receptor expression and unfavorable prognosis in estrogen receptor-negative cases. Histochem. Cell Biol. 2017, 147, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Pinney, S.M.; Mallick, P.; Ho, S.M.; Bracken, B.; Wu, T. Impact of Oxidative Stress Biomarkers and Carboxymethyllysine (an Advanced Glycation End Product) on Prostate Cancer: A Prospective Study. Clin. Genitourin. Cancer 2015, 13, e347–e351. [Google Scholar] [CrossRef] [Green Version]
- Wen, L.; Huang, J.K.; Johnson, B.H.; Reeck, G.R. A human placental cDNA clone that encodes nonhistone chromosomal protein HMG-1. Nucleic Acids Res. 1989, 17, 1197–1214. [Google Scholar] [CrossRef] [Green Version]
- Ohmori, H.; Luo, Y.; Fujii, K.; Sasahira, T.; Shimomoto, T.; Denda, A.; Kuniyasu, H. Dietary linoleic acid and glucose enhances azoxymethane-induced colon cancer and the metastasis through the expression of high mobility group box 1. Pathobiology 2010, 77, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Li, J.Y.; Xu, J.H.; Xia, Z.S.; Cheng, D.; Zhong, W.; Lai, Y.; Yu, T.; Chen, Q.K. Butyrate suppresses abnormal proliferation in colonic epithelial cells under diabetic state by targeting HMGB1. J. Pharmacol. Sci. 2019, 139, 266–274. [Google Scholar] [CrossRef]
- Huang, W.S.; Lin, C.T.; Chen, C.N.; Chang, S.F.; Chang, H.I.; Lee, K.C. Metformin increases the cytotoxicity of oxaliplatin in human DLD-1 colorectal cancer cells through down-regulating HMGB1 expression. J. Cell Biochem. 2018, 119, 6943–6952. [Google Scholar] [CrossRef]
- Japanese Gastric Cancer Association. Japanese classification of gastric carcinoma: 3rd English edition. Gastric Cancer 2011, 14, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Nishiguchi, Y.; Oue, N.; Fujiwara-Tani, R.; Sasaki, T.; Ohmori, H.; Kishi, S.; Mori, S.; Mori, T.; Ikeda, N.; Matsumoto, S.; et al. Role of Metastasis-Related Genes in Cisplatin Chemoresistance in Gastric Cancer. Int. J. Mol. Sci. 2020, 21, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piha, R.S.; Cuénod, M.; Waelsch, H. Metabolism of histones of brain and liver. J. Biol. Chem. 1966, 241, 2397–2404. [Google Scholar] [CrossRef]
- Commerford, S.L.; Carsten, A.L.; Cronkite, E.P. Histone turnover within nonproliferating cells. Proc. Natl. Acad. Sci. USA 1982, 79, 1163–1165. [Google Scholar] [CrossRef] [Green Version]
- Gautieri, A.; Passini, F.S.; Silván, U.; Guizar-Sicairos, M.; Carimati, G.; Volpi, P.; Moretti, M.; Schoenhuber, H.; Redaelli, A.; Berli, M.; et al. Advanced glycation end-products: Mechanics of aged collagen from molecule to tissue. Matrix Biol. 2017, 59, 95–108. [Google Scholar] [CrossRef] [Green Version]
- Anwar, S.; Khan, M.A.; Sadaf, A.; Younus, H. A structural study on the protection of glycation of superoxide dismutase by thymoquinone. Int. J. Biol. Macromol. 2014, 69, 476–481. [Google Scholar] [CrossRef]
- Lowe, S.W.; Bodis, S.; McClatchey, A.; Remington, L.; Ruley, H.E.; Fisher, D.E.; Housman, D.E.; Jacks, T. p53 status and the efficacy of cancer therapy in vivo. Science 1994, 266, 807–810. [Google Scholar] [CrossRef] [PubMed]
- Yokozaki, H. Molecular characteristics of eight gastric cancer cell lines established in Japan. Pathol. Int. 2000, 50, 767–777. [Google Scholar] [CrossRef]
- Kawahara, K.; Biswas, K.K.; Unoshima, M.; Ito, T.; Kikuchi, K.; Morimoto, Y.; Iwata, M.; Tancharoen, S.; Oyama, Y.; Takenouchi, K.; et al. C-reactive protein induces high-mobility group box-1 protein release through activation of p38MAPK in macrophage RAW264.7 cells. Cardiovasc Pathol. 2008, 17, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Wang, J.; Li, W.; Yuan, Y.Z.; Li, C.Y.; Qian, X.H.; Xu, W.X.; Zhan, Y.Q.; Yang, X.M. Proteomic screen defines the hepatocyte nuclear factor 1alpha-binding partners and identifies HMGB1 as a new cofactor of HNF1α. Nucleic Acids Res. 2008, 36, 1209–1219. [Google Scholar] [CrossRef] [Green Version]
- Stros, M.; Cherny, D.; Jovin, T.M. HMG1 protein stimulates DNA end joining by promoting association of DNA molecules via their ends. Eur. J. Biochem. 2000, 267, 4088–4097. [Google Scholar] [CrossRef]
- Ugrinova, I.; Mitkova, E.; Moskalenko, C.; Pashev, I.; Pasheva, E. DNA bending versus DNA end joining activity of HMGB1 protein is modulated in vitro by acetylation. Biochemistry 2007, 46, 2111–2117. [Google Scholar] [CrossRef]
- Sakaguchi, M.; Murata, H.; Yamamoto, K.; Ono, T.; Sakaguchi, Y.; Motoyama, A.; Hibino, T.; Kataoka, K.; Huh, N.H. TIRAP, an adaptor protein for TLR2/4, transduces a signal from RAGE phosphorylated upon ligand binding. PLoS ONE 2011, 6, e23132. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara-Tani, R.; Sasaki, T.; Fujii, K.; Luo, Y.; Mori, T.; Kishi, S.; Mori, S.; Matsushima-Otsuka, S.; Nishiguchi, Y.; Goto, K.; et al. Diabetes mellitus is associated with liver metastasis of colorectal cancer through production of biglycan-rich cancer stroma. Oncotarget 2020, 11, 2982–2994. [Google Scholar] [CrossRef] [PubMed]
- Kwak, M.S.; Kim, H.S.; Lee, B.; Kim, Y.H.; Son, M.; Shin, J.S. Immunological Significance of HMGB1 Post-Translational Modification and Redox Biology. Front. Immunol. 2020, 11, 1189. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Antoine, D.J.; Andersson, U.; Tracey, K.J. The many faces of HMGB1: Molecular structure-functional activity in inflammation, apoptosis, and chemotaxis. J. Leukoc. Biol. 2013, 93, 865–873. [Google Scholar] [CrossRef] [Green Version]
- Mori, S.; Kishi, S.; Honoki, K.; Fujiwara-Tani, R.; Moriguchi, T.; Sasaki, T.; Fujii, K.; Tsukamoto, S.; Fujii, H.; Kido, A.; et al. Anti-Stem Cell Property of Pterostilbene in Gastrointestinal Cancer Cells. Int. J. Mol. Sci. 2020, 21, 9347. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Huang, X.; Kuang, Y.; Xing, Z.; Deng, X.; Luo, Z. Thapsigargin induces apoptosis in adrenocortical carcinoma by activating endoplasmic reticulum stress and the JNK signaling pathway: An in vitro and in vivo study. Drug Des. Dev. Ther. 2019, 13, 2787–2798. [Google Scholar] [CrossRef] [Green Version]
- Kuniyasu, H.; Luo, Y.; Fujii, K.; Sasahira, T.; Moriwaka, Y.; Tatsumoto, N.; Sasaki, T.; Yamashita, Y.; Ohmori, H. CD10 enhances metastasis of colorectal cancer by abrogating the anti-tumoural effect of methionine-enkephalin in the liver. Gut 2010, 59, 348–356. [Google Scholar] [CrossRef]
- Matsushima-Otsuka, S.; Fujiwara-Tani, R.; Sasaki, T.; Ohmori, H.; Nakashima, C.; Kishi, S.; Nishiguchi, Y.; Fujii, K.; Luo, Y.; Kuniyasu, H. Significance of intranuclear angiotensin-II type 2 receptor in oral squamous cell carcinoma. Oncotarget 2018, 9, 36561–36574. [Google Scholar] [CrossRef] [Green Version]
- Sensi, M.; Bruno, M.R.; Valente, L.; Cioccia, G.P.; Chianelli, M.; Pozzilli, P. Retinol binding protein: A short half life determinant of protein non enzymatic glycation in diabetes. Diabetes Res. 1990, 13, 195–198. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter 1 | Level | Number | Protein Level | |||
|---|---|---|---|---|---|---|
| HMGB1 2 | p3 | CML-HMGB1 2 | p3 | |||
| Grade | 1 | 6 | 104 ± 21 | NS | 59 ± 17 | NS |
| 2 | 4 | 98 ± 21 | 54 ± 18 | |||
| pT | 2 | 3 | 8 ± 3 | 0.0015 | 43 ± 12 | 0.0245 |
| 3 | 4 | 96 ± 11 | 54 ± 13 | |||
| 4 | 3 | 127 ± 8 | 75 ± 2 | |||
| pN | 0 | 3 | 83 ± 8 | NS | 38 ± 10 | 0.0069 |
| 1 and 2 | 7 | 109 ± 10 | 65 ± 10 | |||
| pM | 0 | 8 | 94 ± 4 | 0.0123 | 52 ± 14 | 0.0377 |
| 1 | 2 | 130 ± 7 | 78 ± 4 | |||
| pStage | 1 and 2 | 3 | 83 ± 8 | NS | 38 ± 10 | 0.0069 |
| 3 and 4 | 7 | 109 ± 18 | 65 ± 11 | |||
| Variable | Coefficient | SE | 95% Confidence Interval | p |
|---|---|---|---|---|
| HMGB1 | −0.01398 | 0.01444 | −0.04812 to 0.02016 | 0.0014 |
| CML-HMGB1 | 0.03455 | 0.01189 | 0.006428 to 0.06267 | |
| SUM | −1.612 | 0.3951 | −2.523 to −0.7007 | 0.0008 |
| Protein | Company | Cat # |
|---|---|---|
| HMGB1 | Abcam | ab18256 |
| p16 | Santa Cruz | SC-377412 |
| mut p53 | Abcam | ab32509 |
| Ac Histone H4 | Rockland | 600-401-P06 |
| Histone H4 | Santa Cruz | SC-25260 |
| Rb | Santa Cruz | SC-74562 |
| Cox2 | Santa Cruz | SC-514489 |
| iNOS | Santa Cruz | SC-7271 |
| BCL-2 | Proteintech | 26593-1-AP |
| BAX | Proteintech | 50599-2-IG |
| Cyclin E | Proteintech | 11554-1-AP |
| Cyclin D1 | Proteintech | 26939-1-AP |
| β-catenin | Proteintech | 13665-1-AP |
| E-cadherin | Santa Cruz | SC-8426 |
| RAGE | Santa Cruz | SC-365154 |
| c-met | Life Span | LS-C826326-100 |
| EGFR | Santa Cruz | SC-373746 |
| RBP | Abcam | ab48624 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kishi, S.; Nishiguchi, Y.; Honoki, K.; Mori, S.; Fujiwara-Tani, R.; Sasaki, T.; Fujii, K.; Kawahara, I.; Goto, K.; Nakashima, C.; et al. Role of Glycated High Mobility Group Box-1 in Gastric Cancer. Int. J. Mol. Sci. 2021, 22, 5185. https://doi.org/10.3390/ijms22105185
Kishi S, Nishiguchi Y, Honoki K, Mori S, Fujiwara-Tani R, Sasaki T, Fujii K, Kawahara I, Goto K, Nakashima C, et al. Role of Glycated High Mobility Group Box-1 in Gastric Cancer. International Journal of Molecular Sciences. 2021; 22(10):5185. https://doi.org/10.3390/ijms22105185
Chicago/Turabian StyleKishi, Shingo, Yukiko Nishiguchi, Kanya Honoki, Shiori Mori, Rina Fujiwara-Tani, Takamitsu Sasaki, Kiyomu Fujii, Isao Kawahara, Kei Goto, Chie Nakashima, and et al. 2021. "Role of Glycated High Mobility Group Box-1 in Gastric Cancer" International Journal of Molecular Sciences 22, no. 10: 5185. https://doi.org/10.3390/ijms22105185