
Targeting of Deregulated Wnt/β-Catenin Signaling by PRI-724 and LGK974 Inhibitors in Germ Cell Tumor Cell Lines

 , , , , and
, , , , and 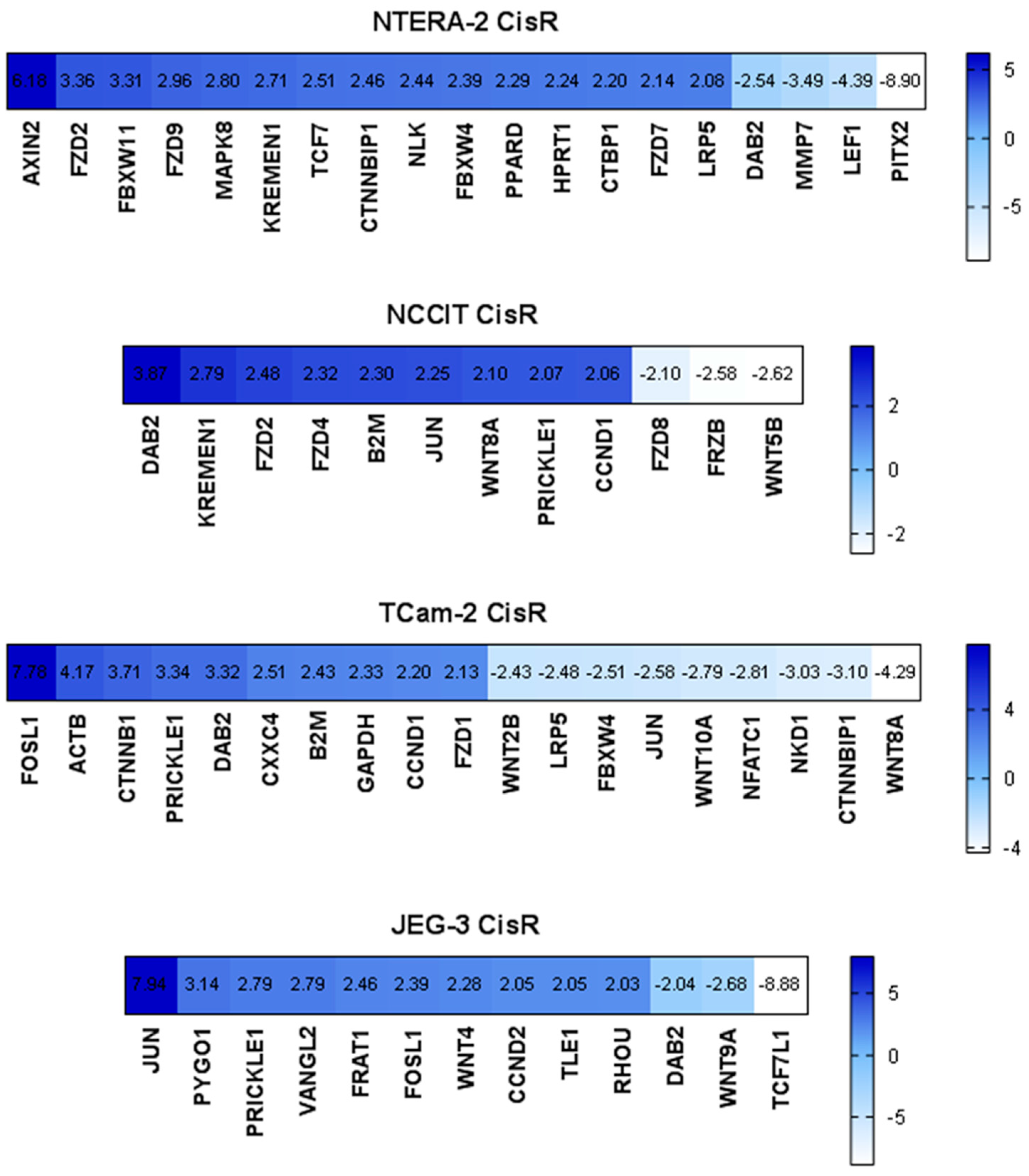{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Expression Analysis of the Wnt Signaling Pathway in Parental and Cisplatin-Resistant GCT Cell Lines
2.2. β-Catenin and Cyclin D1 Expression in Parental and Cisplatin-Resistant GCT Cell Lines
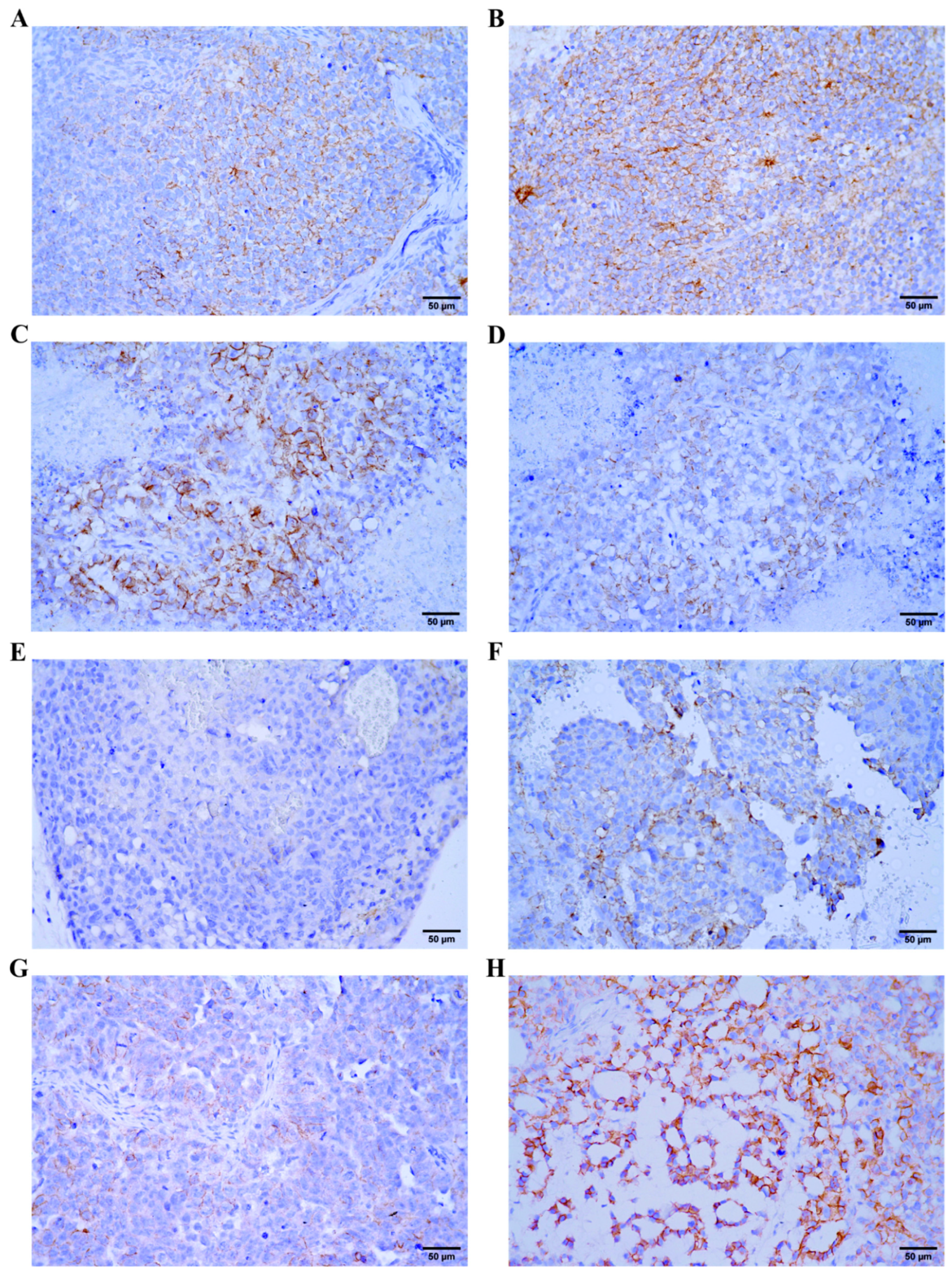
2.3. β-Catenin Expression in GCT Xenograft Models
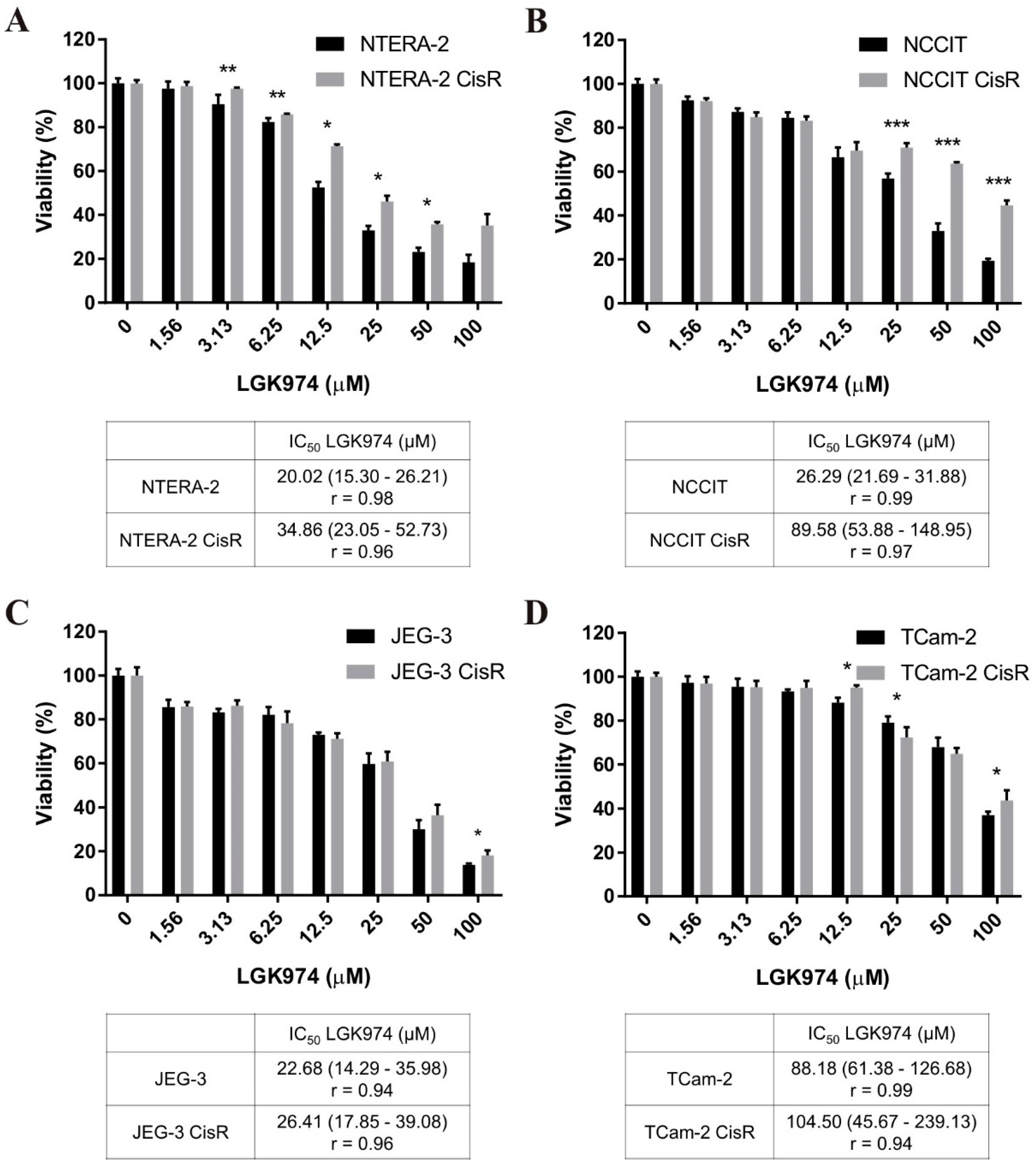
2.4. Therapeutic Targeting of Wnt/β-Catenin Signaling with LGK974 and PRI-724 in Parental and Cisplatin-Resistant GCT Cell Lines
2.5. Effect of PRI-724 Treatment on Caspase-3/7 Activity and Induction of Cell Death in Parental and Cisplatin-Resistant GCT Cell Lines
2.6. Effect of PRI-724 Treatment on Migration of Cisplatin-Resistant GCT Cell Lines
2.7. Effects of Combined Treatment with PRI-724 and Cisplatin in NTERA-2 CisR Cells
2.8. β-Catenin and Cyclin D1 Expression in PRI-724 Treated Cisplatin-Resistant GCT Cell Lines
2.9. In Vivo Efficacy of PRI-724 in NTERA-2 CisR Xenograft Model
3. Discussion
4. Materials and Methods
4.1. Cell Cultivation
4.2. Viability Assays
4.3. Caspase Assay
4.4. Annexin V Assay
4.5. 3D Migration Assay
4.6. Wound Healing Assay
4.7. Gene Expression Array
4.8. qPCR Expression Analysis
4.9. Western Blot
4.10. In Vivo Experiments
4.11. Immunohistochemistry
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, L.; Albers, P.; Berney, D.M.; Feldman, D.R.; Daugaard, G.; Gilligan, T.; Looijenga, L.H.J. Testicular cancer. Nat. Rev. Dis. Primers 2018, 4, 29. [Google Scholar] [CrossRef]
- Znaor, A.; Lortet-Tieulent, J.; Jemal, A.; Bray, F. International variations and trends in testicular cancer incidence and mortality. Eur. Urol. 2014, 65, 1095–1106. [Google Scholar] [CrossRef]
- Van de Geijn, G.J.; Hersmus, R.; Looijenga, L.H. Recent developments in testicular germ cell tumor research. Birth Defects Res. Part C Embryo Today Rev. 2009, 87, 96–113. [Google Scholar] [CrossRef]
- Oosterhuis, J.W.; Looijenga, L.H.J. Human germ cell tumours from a developmental perspective. Nat. Rev. Cancer 2019, 19, 522–537. [Google Scholar] [CrossRef] [PubMed]
- Moch, H.; Cubilla, A.L.; Humphrey, P.A.; Reuter, V.E.; Ulbright, T.M. The 2016 WHO classification of tumours of the urinary system and male genital organs—Part A: Renal, penile, and testicular tumours. Eur. Urol. 2016, 70, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Einhorn, L.H. Treatment of testicular cancer: A new and improved model. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 1990, 8, 1777–1781. [Google Scholar] [CrossRef]
- Allen, J.C.; Kirschner, A.; Scarpato, K.R.; Morgans, A.K. Current management of refractory germ cell tumors and future directions. Curr. Oncol. Rep. 2017, 19, 8. [Google Scholar] [CrossRef] [PubMed]
- Kondagunta, G.V.; Bacik, J.; Sheinfeld, J.; Bajorin, D.; Bains, M.; Reich, L.; Deluca, J.; Budnick, A.; Ishill, N.; Mazumdar, M.; et al. Paclitaxel plus Ifosfamide followed by high-dose carboplatin plus etoposide in previously treated germ cell tumors. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2007, 25, 85–90. [Google Scholar] [CrossRef]
- Motzer, R.J.; Sheinfeld, J.; Mazumdar, M.; Bains, M.; Mariani, T.; Bacik, J.; Bajorin, D.; Bosl, G.J. Paclitaxel, ifosfamide, and cisplatin second-line therapy for patients with relapsed testicular germ cell cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2000, 18, 2413–2418. [Google Scholar] [CrossRef] [PubMed]
- Mardiak, J.; Salek, T.; Sycova-Mila, Z.; Obertova, J.; Hlavata, Z.; Mego, M.; Reckova, M.; Koza, I. Paclitaxel plus ifosfamide and cisplatin in second-line treatment of germ cell tumors: A phase II study. Neoplasma 2005, 52, 497–501. [Google Scholar]
- Adra, N.; Abonour, R.; Althouse, S.K.; Albany, C.; Hanna, N.H.; Einhorn, L.H. High-dose chemotherapy and autologous peripheral-blood stem-cell transplantation for relapsed metastatic germ cell tumors: The Indiana University experience. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2017, 35, 1096–1102. [Google Scholar] [CrossRef] [PubMed]
- Feldman, D.R.; Sheinfeld, J.; Bajorin, D.F.; Fischer, P.; Turkula, S.; Ishill, N.; Patil, S.; Bains, M.; Reich, L.M.; Bosl, G.J.; et al. TI-CE high-dose chemotherapy for patients with previously treated germ cell tumors: Results and prognostic factor analysis. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2010, 28, 1706–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Prognostic Factors Study Group; Lorch, A.; Beyer, J.; Bascoul-Mollevi, C.; Kramar, A.; Einhorn, L.H.; Necchi, A.; Massard, C.; De Giorgi, U.; Flechon, A.; et al. Prognostic factors in patients with metastatic germ cell tumors who experienced treatment failure with cisplatin-based first-line chemotherapy. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2010, 28, 4906–4911. [Google Scholar] [CrossRef]
- Oing, C.; Alsdorf, W.H.; von Amsberg, G.; Oechsle, K.; Bokemeyer, C. Platinum-refractory germ cell tumors: An update on current treatment options and developments. World J. Urol. 2017, 35, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Fazal, Z.; Freemantle, S.J.; Spinella, M.J. Mechanisms of cisplatin sensitivity and resistance in testicular germ cell tumors. Cancer Drug Resist. 2019, 2, 580–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nusse, R.; Varmus, H. Three decades of Wnts: A personal perspective on how a scientific field developed. EMBO J. 2012, 31, 2670–2684. [Google Scholar] [CrossRef] [Green Version]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt signaling in cancer. Oncogene 2017, 36, 1461–1473. [Google Scholar] [CrossRef]
- Zhong, Z.; Virshup, D.M. Wnt signaling and drug resistance in cancer. Mol. Pharmacol. 2020, 97, 72–89. [Google Scholar] [CrossRef] [Green Version]
- Bugter, J.M.; Fenderico, N.; Maurice, M.M. Mutations and mechanisms of WNT pathway tumour suppressors in cancer. Nat. Rev. Cancer 2021, 21, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Honecker, F.; Kersemaekers, A.M.; Molier, M.; Van Weeren, P.C.; Stoop, H.; De Krijger, R.R.; Wolffenbuttel, K.P.; Oosterhuis, W.; Bokemeyer, C.; Looijenga, L.H. Involvement of E-cadherin and beta-catenin in germ cell tumours and in normal male fetal germ cell development. J. Pathol. 2004, 204, 167–174. [Google Scholar] [CrossRef]
- Vladusic, T.; Hrascan, R.; Vrhovac, I.; Kruslin, B.; Gamulin, M.; Grgic, M.; Pecina-Slaus, N.; Franekic Colic, J. Loss of heterozygosity of selected tumor suppressor genes in human testicular germ cell tumors. Pathol. Res. Pract. 2010, 206, 163–167. [Google Scholar] [CrossRef] [Green Version]
- Fabijanovic, D.; Zunic, I.; Martic, T.N.; Skenderi, F.; Serman, L.; Vranic, S. The expression of SFRP1, SFRP3, DVL1, and DVL2 proteins in testicular germ cell tumors. Apmis Acta Pathol. Microbiol. Immunol. Scand. 2016, 124, 942–949. [Google Scholar] [CrossRef] [PubMed]
- Lafin, J.T.; Bagrodia, A.; Woldu, S.; Amatruda, J.F. New insights into germ cell tumor genomics. Andrology 2019, 7, 507–515. [Google Scholar] [CrossRef] [Green Version]
- Young, J.C.; Kerr, G.; Micati, D.; Nielsen, J.E.; Rajpert-De Meyts, E.; Abud, H.E.; Loveland, K.L. WNT signalling in the normal human adult testis and in male germ cell neoplasms. Hum. Reprod. 2020, 35, 1991–2003. [Google Scholar] [CrossRef]
- Shen, H.; Shih, J.; Hollern, D.P.; Wang, L.; Bowlby, R.; Tickoo, S.K.; Thorsson, V.; Mungall, A.J.; Newton, Y.; Hegde, A.M.; et al. Integrated molecular characterization of testicular germ cell tumors. Cell Rep. 2018, 23, 3392–3406. [Google Scholar] [CrossRef] [PubMed]
- Bagrodia, A.; Lee, B.H.; Lee, W.; Cha, E.K.; Sfakianos, J.P.; Iyer, G.; Pietzak, E.J.; Gao, S.P.; Zabor, E.C.; Ostrovnaya, I.; et al. Genetic determinants of cisplatin resistance in patients with advanced germ cell tumors. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2016, 34, 4000–4007. [Google Scholar] [CrossRef] [PubMed]
- Chovanec, M.; Cierna, Z.; Miskovska, V.; Machalekova, K.; Kalavska, K.; Rejlekova, K.; Svetlovska, D.; Macak, D.; Spanik, S.; Kajo, K.; et al. betacatenin is a marker of poor clinical characteristics and suppressed immune infiltration in testicular germ cell tumors. BMC Cancer 2018, 18, 1062. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.S.; Park, J.I. Wnt signaling in cancer: Therapeutic targeting of Wnt signaling beyond beta-catenin and the destruction complex. Exp. Mol. Med. 2020, 52, 183–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harb, J.; Lin, P.J.; Hao, J. Recent development of Wnt signaling pathway inhibitors for cancer therapeutics. Curr. Oncol. Rep. 2019, 21, 12. [Google Scholar] [CrossRef]
- El-Khoueiry, A.B.; Ning, Y.; Yang, D.; Cole, S.; Kahn, M.; Zoghbi, M.; Berg, J.; Fujimori, M.; Inada, T.; Kouji, H. A phase I first-in-human study of PRI-724 in patients (pts) with advanced solid tumors. J. Clin. Oncol. Am. Soc. Clin. Oncol. J. 2013, 31, 2501. [Google Scholar] [CrossRef]
- Ko, A.H.; Chiorean, E.G.; Kwak, E.L.; Lenz, H.-J.; Nadler, P.I.; Wood, D.L.; Fujimori, M.; Inada, T.; Kouji, H.; McWilliams, R.R. Final results of a phase Ib dose-escalation study of PRI-724, a CBP/beta-catenin modulator, plus gemcitabine (GEM) in patients with advanced pancreatic adenocarcinoma (APC) as second-line therapy after FOLFIRINOX or FOLFOX. J. Clin. Oncol. Am. Soc. Clin. Oncol. J. 2016, 34, e15721. [Google Scholar] [CrossRef]
- McWilliams, R.R.; Ko, A.H.; Chiorean, E.G.; Kwak, E.L.; Lenz, H.-J.; Nadler, P.I.; Wood, D.L.; Fujimori, M.; Morita, K.; Inada, T. A phase Ib dose-escalation study of PRI-724, a CBP/beta-catenin modulator, plus gemcitabine (GEM) in patients with advanced pancreatic adenocarcinoma (APC) as second-line therapy after FOLFIRINOX or FOLFOX. J. Clin. Oncol. Am. Soc. Clin. Oncol. J. 2015, 33, e15270. [Google Scholar] [CrossRef]
- Emami, K.H.; Nguyen, C.; Ma, H.; Kim, D.H.; Jeong, K.W.; Eguchi, M.; Moon, R.T.; Teo, J.L.; Kim, H.Y.; Moon, S.H.; et al. A small molecule inhibitor of beta-catenin/CREB-binding protein transcription [corrected]. Proc. Natl. Acad. Sci. USA 2004, 101, 12682–12687. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Pan, S.; Hsieh, M.H.; Ng, N.; Sun, F.; Wang, T.; Kasibhatla, S.; Schuller, A.G.; Li, A.G.; Cheng, D.; et al. Targeting Wnt-driven cancer through the inhibition of Porcupine by LGK974. Proc. Natl. Acad. Sci. USA 2013, 110, 20224–20229. [Google Scholar] [CrossRef] [Green Version]
- Schmidtova, S.; Kalavska, K.; Gercakova, K.; Cierna, Z.; Miklikova, S.; Smolkova, B.; Buocikova, V.; Miskovska, V.; Durinikova, E.; Burikova, M.; et al. Disulfiram overcomes cisplatin resistance in human embryonal carcinoma cells. Cancers 2019, 11, 1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, T.C. Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharm. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef]
- Adra, N.; Einhorn, L.H. Testicular cancer update. Clin. Adv. Hematol. Oncol. HO 2017, 15, 386–396. [Google Scholar]
- Kozakova, K.; Mego, M.; Cheng, L.; Chovanec, M. Promising novel therapies for relapsed and refractory testicular germ cell tumors. Expert Rev. Anticancer Ther. 2021, 21, 53–69. [Google Scholar] [CrossRef] [PubMed]
- Oechsle, K.; Kollmannsberger, C.; Honecker, F.; Mayer, F.; Waller, C.F.; Hartmann, J.T.; Boehlke, I.; Bokemeyer, C.; German Testicular Cancer Study Group. Long-term survival after treatment with gemcitabine and oxaliplatin with and without paclitaxel plus secondary surgery in patients with cisplatin-refractory and/or multiply relapsed germ cell tumors. Eur. Urol. 2011, 60, 850–855. [Google Scholar] [CrossRef]
- Mego, M.; Svetlovska, D.; Miskovska, V.; Obertova, J.; Palacka, P.; Rajec, J.; Sycova-Mila, Z.; Chovanec, M.; Rejlekova, K.; Zuzak, P.; et al. Phase II study of everolimus in refractory testicular germ cell tumors. Urol. Oncol. 2016, 34, e117–e122. [Google Scholar] [CrossRef] [PubMed]
- Fenner, M.; Oing, C.; Dieing, A.; Gauler, T.; Oechsle, K.; Lorch, A.; Hentrich, M.; Kopp, H.G.; Bokemeyer, C.; Honecker, F. Everolimus in patients with multiply relapsed or cisplatin refractory germ cell tumors: Results of a phase II, single-arm, open-label multicenter trial (RADIT) of the German Testicular Cancer Study Group. J. Cancer Res. Clin. Oncol. 2019, 145, 717–723. [Google Scholar] [CrossRef]
- Jain, A.; Brames, M.J.; Vaughn, D.J.; Einhorn, L.H. Phase II clinical trial of oxaliplatin and bevacizumab in refractory germ cell tumors. Am. J. Clin. Oncol. 2014, 37, 450–453. [Google Scholar] [CrossRef]
- Oechsle, K.; Honecker, F.; Cheng, T.; Mayer, F.; Czaykowski, P.; Winquist, E.; Wood, L.; Fenner, M.; Glaesener, S.; Hartmann, J.T.; et al. Preclinical and clinical activity of sunitinib in patients with cisplatin-refractory or multiply relapsed germ cell tumors: A Canadian Urologic Oncology Group/German Testicular Cancer Study Group cooperative study. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2011, 22, 2654–2660. [Google Scholar] [CrossRef]
- Mego, M.; Svetlovska, D.; Chovanec, M.; Reckova, M.; Rejlekova, K.; Obertova, J.; Palacka, P.; Sycova-Mila, Z.; De Giorgi, U.; Mardiak, J. Phase II study of avelumab in multiple relapsed/refractory germ cell cancer. Investig. New Drugs 2019, 37, 748–754. [Google Scholar] [CrossRef]
- Adra, N.; Einhorn, L.H.; Althouse, S.K.; Ammakkanavar, N.R.; Musapatika, D.; Albany, C.; Vaughn, D.; Hanna, N.H. Phase II trial of pembrolizumab in patients with platinum refractory germ-cell tumors: A Hoosier Cancer Research Network Study GU14-206. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2018, 29, 209–214. [Google Scholar] [CrossRef]
- Schmidtova, S.; Kalavska, K.; Kucerova, L. Molecular mechanisms of cisplatin chemoresistance and its circumventing in testicular germ cell tumors. Curr. Oncol. Rep. 2018, 20, 88. [Google Scholar] [CrossRef]
- Kalavska, K.; Conteduca, V.; De Giorgi, U.; Mego, M. Molecular mechanisms of resistance in testicular germ cell tumors—Clinical implications. Curr. Cancer Drug Targets 2018, 18, 967–978. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.F.; Spoettl, G.; Maurer, J.; Nolting, S.; Auernhammer, C.J. Inhibition of Wnt/beta-catenin signaling in neuroendocrine tumors in vitro: Antitumoral effects. Cancers 2020, 12, 345. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wu, G.; Xu, Y.; Li, J.; Ruan, N.; Chen, Y.; Zhang, Q.; Xia, Q. Porcupine inhibitor LGK974 downregulates the Wnt signaling pathway and inhibits clear cell renal cell carcinoma. BioMed Res. Int. 2020, 2020, 2527643. [Google Scholar] [CrossRef] [PubMed]
- Suwala, A.K.; Koch, K.; Rios, D.H.; Aretz, P.; Uhlmann, C.; Ogorek, I.; Felsberg, J.; Reifenberger, G.; Kohrer, K.; Deenen, R.; et al. Inhibition of Wnt/beta-catenin signaling downregulates expression of aldehyde dehydrogenase isoform 3A1 (ALDH3A1) to reduce resistance against temozolomide in glioblastoma in vitro. Oncotarget 2018, 9, 22703–22716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boone, J.D.; Arend, R.C.; Johnston, B.E.; Cooper, S.J.; Gilchrist, S.A.; Oelschlager, D.K.; Grizzle, W.E.; McGwin, G., Jr.; Gangrade, A.; Straughn, J.M., Jr.; et al. Targeting the Wnt/beta-catenin pathway in primary ovarian cancer with the porcupine inhibitor WNT974. Lab. Investig. J. Tech. Methods Pathol. 2016, 96, 249–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, M.; Baker, A.; Goldstein, S.D.; Albert, C.M.; Jackson, K.W.; McCarty, G.; Kahlert, U.D.; Loeb, D.M. Inhibition of porcupine prolongs metastasis free survival in a mouse xenograft model of Ewing sarcoma. Oncotarget 2017, 8, 78265–78276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudy, S.F.; Brenner, J.C.; Harris, J.L.; Liu, J.; Che, J.; Scott, M.V.; Owen, J.H.; Komarck, C.M.; Graham, M.P.; Bellile, E.L.; et al. In vivo Wnt pathway inhibition of human squamous cell carcinoma growth and metastasis in the chick chorioallantoic model. J. Otolaryngol. Head Neck Surg. J. D’oto-Rhino-Laryngol. Chir. Cervico-Faciale 2016, 45, 26. [Google Scholar] [CrossRef] [Green Version]
- Fang, F.; VanCleave, A.; Helmuth, R.; Torres, H.; Rickel, K.; Wollenzien, H.; Sun, H.; Zeng, E.; Zhao, J.; Tao, J. Targeting the Wnt/beta-catenin pathway in human osteosarcoma cells. Oncotarget 2018, 9, 36780–36792. [Google Scholar] [CrossRef] [Green Version]
- Kleszcz, R.; Szymanska, A.; Krajka-Kuzniak, V.; Baer-Dubowska, W.; Paluszczak, J. Inhibition of CBP/beta-catenin and porcupine attenuates Wnt signaling and induces apoptosis in head and neck carcinoma cells. Cell. Oncol. 2019, 42, 505–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabata, R.; Harada, K.; Mizutani, Y.; Ouchi, H.; Yoshimura, K.; Sato, Y.; Kitao, A.; Kimura, K.; Kouji, H.; Miyashita, T.; et al. Anti-tumor activity of the small molecule inhibitor PRI-724 against beta-catenin-activated hepatocellular carcinoma. Anticancer Res. 2020, 40, 5211–5219. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Font, E.; Perez-Capo, M.; Ramos, R.; Felipe, I.; Garcias, C.; Luna, P.; Terrasa, J.; Martin-Broto, J.; Vogler, O.; Alemany, R.; et al. Impact of Wnt/beta-catenin inhibition on cell proliferation through CDC25A downregulation in soft tissue sarcomas. Cancers 2020, 12, 2556. [Google Scholar] [CrossRef]
- Andrews, P.W.; Damjanov, I.; Simon, D.; Banting, G.S.; Carlin, C.; Dracopoli, N.C.; Fogh, J. Pluripotent embryonal carcinoma clones derived from the human teratocarcinoma cell line Tera-2. Differentiation in vivo and in vitro. Lab. Investig. J. Tech. Methods Pathol. 1984, 50, 147–162. [Google Scholar]
- International Germ Cell Cancer Collaborative Group. International germ cell consensus classification: A prognostic factor-based staging system for metastatic germ cell cancers. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 1997, 15, 594–603. [Google Scholar] [CrossRef]
- Teshima, S.; Shimosato, Y.; Hirohashi, S.; Tome, Y.; Hayashi, I.; Kanazawa, H.; Kakizoe, T. Four new human germ cell tumor cell lines. Lab. Investig. J. Tech. Methods Pathol. 1988, 59, 328–336. [Google Scholar]
- Damjanov, I.; Horvat, B.; Gibas, Z. Retinoic acid-induced differentiation of the developmentally pluripotent human germ cell tumor-derived cell line, NCCIT. Lab. Investig. J. Tech. Methods Pathol. 1993, 68, 220–232. [Google Scholar]
- Albany, C.; Einhorn, L.H. Extragonadal germ cell tumors: Clinical presentation and management. Curr. Opin. Oncol. 2013, 25, 261–265. [Google Scholar] [CrossRef]
- Ganjoo, K.N.; Rieger, K.M.; Kesler, K.A.; Sharma, M.; Heilman, D.K.; Einhorn, L.H. Results of modern therapy for patients with mediastinal nonseminomatous germ cell tumors. Cancer 2000, 88, 1051–1056. [Google Scholar] [CrossRef]
- Gillessen, S.; Sauve, N.; Collette, L.; Daugaard, G.; de Wit, R.; Albany, C.; Tryakin, A.; Fizazi, K.; Stahl, O.; Gietema, J.A.; et al. Predicting outcomes in men with metastatic Nonseminomatous Germ Cell Tumors (NSGCT): Results from the IGCCCG update consortium. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2021. [Google Scholar] [CrossRef]
- Nagaraj, A.B.; Joseph, P.; Kovalenko, O.; Singh, S.; Armstrong, A.; Redline, R.; Resnick, K.; Zanotti, K.; Waggoner, S.; DiFeo, A. Critical role of Wnt/beta-catenin signaling in driving epithelial ovarian cancer platinum resistance. Oncotarget 2015, 6, 23720–23734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, K.; Ikoma, A.; Shibakawa, M.; Shimoda, S.; Harada, K.; Saio, M.; Imamura, J.; Osawa, Y.; Kimura, M.; Nishikawa, K.; et al. Safety, tolerability, and preliminary efficacy of the anti-fibrotic small molecule PRI-724, a CBP/beta-catenin inhibitor, in patients with hepatitis C virus-related cirrhosis: A single-center, open-label, dose escalation phase 1 trial. EBioMedicine 2017, 23, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Sundquist, T.; Moravec, R.; Niles, A.; O’Brien, M.; Riss, T. Timing your apoptosis assays. Cell Notes 2006, 16, 18–21. [Google Scholar]
- Zhang, J.; Liu, J.; Li, H.; Wang, J. beta-Catenin signaling pathway regulates cisplatin resistance in lung adenocarcinoma cells by upregulating Bcl-xl. Mol. Med. Rep. 2016, 13, 2543–2551. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Liu, H.C.; Wang, C.; Liu, X.; Hu, F.C.; Xie, N.; Lu, L.; Chen, X.; Huang, H.Z. Overexpression of beta-catenin induces cisplatin resistance in oral squamous cell carcinoma. BioMed Res. Int. 2016, 2016, 5378567. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Wei, W.; Sun, Y.; Gao, J.; Wang, Q.; Zheng, J. Interference with the expression of beta-catenin reverses cisplatin resistance in A2780/DDP cells and inhibits the progression of ovarian cancer in mouse model. DNA Cell Biol. 2015, 34, 55–62. [Google Scholar] [CrossRef]
- Barghout, S.H.; Zepeda, N.; Xu, Z.; Steed, H.; Lee, C.H.; Fu, Y. Elevated beta-catenin activity contributes to carboplatin resistance in A2780cp ovarian cancer cells. Biochem. Biophys. Res. Commun. 2015, 468, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Kar, M.; Roy, S.; Saha, A.; Padhi, S.; Banerjee, B. Role of beta-catenin in cisplatin resistance, relapse and prognosis of head and neck squamous cell carcinoma. Cell. Oncol. 2018, 41, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Lobo, J.; Gillis, A.J.M.; van den Berg, A.; Looijenga, L.H.J. Prediction of relapse in stage I testicular germ cell tumor patients on surveillance: Investigation of biomarkers. BMC Cancer 2020, 20, 728. [Google Scholar] [CrossRef]
- Wang, H.; Wang, H.; Makki, M.S.; Wen, J.; Dai, Y.; Shi, Q.; Liu, Q.; Zhou, X.; Wang, J. Overexpression of beta-catenin and cyclinD1 predicts a poor prognosis in ovarian serous carcinomas. Int. J. Clin. Exp. Pathol. 2014, 7, 264–271. [Google Scholar]
- Sewify, E.M.; Afifi, O.A.; Mosad, E.; Zaki, A.H.; El Gammal, S.A. Cyclin D1 amplification in multiple myeloma is associated with multidrug resistance expression. Clin. Lymphoma Myeloma Leuk. 2014, 14, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhang, Z.; Zhou, X.; Qiu, W.; Chen, F.; Chen, W. Identification of genes associated with cisplatin resistance in human oral squamous cell carcinoma cell line. BMC Cancer 2006, 6, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noel, E.E.; Yeste-Velasco, M.; Mao, X.; Perry, J.; Kudahetti, S.C.; Li, N.F.; Sharp, S.; Chaplin, T.; Xue, L.; McIntyre, A.; et al. The association of CCND1 overexpression and cisplatin resistance in testicular germ cell tumors and other cancers. Am. J. Pathol. 2010, 176, 2607–2615. [Google Scholar] [CrossRef]
- Cierna, Z.; Mego, M.; Miskovska, V.; Machalekova, K.; Chovanec, M.; Svetlovska, D.; Hainova, K.; Rejlekova, K.; Macak, D.; Spanik, S.; et al. Prognostic value of programmed-death-1 receptor (PD-1) and its ligand 1 (PD-L1) in testicular germ cell tumors. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2016, 27, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Huang, Y.; Gao, X.; Li, Y.; Lin, J.; Chen, L.; Chang, L.; Chen, G.; Guan, Y.; Pan, L.K.; et al. CCND1 Amplification contributes to immunosuppression and is associated with a poor prognosis to immune checkpoint inhibitors in solid tumors. Front. Immunol. 2020, 11, 1620. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Chi, Z.; Chen, Y.; Liu, X.; Wu, D.; Chen, J.; Song, X.; Wang, W.; Dong, L.; Song, H.; et al. Safety, efficacy, and biomarker analysis of toripalimab in previously treated advanced melanoma: Results of the POLARIS-01 multicenter phase II trial. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2020, 26, 4250–4259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Yan, J.; Guo, Q.; Chi, Z.; Tang, B.; Zheng, B.; Yu, J.; Yin, T.; Cheng, Z.; Wu, X.; et al. Genetic aberrations in the CDK4 pathway are associated with innate resistance to PD-1 blockade in Chinese patients with non-cutaneous melanoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2019, 25, 6511–6523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, D.; Wang, Y.; Singavi, A.K.; Mackinnon, A.C.; George, B.; You, M. Immunogenomic landscape contributes to hyperprogressive disease after anti-PD-1 immunotherapy for cancer. iScience 2018, 9, 258–277. [Google Scholar] [CrossRef] [Green Version]
- Singavi, A.; Menon, S.; Kilari, D.; Alqwasmi, A.; Ritch, P.; Thomas, J.; Martin, A.; Oxencis, C.; Ali, S.; George, B. 1140PDPredictive biomarkers for hyper-progression (HP) in response to immune checkpoint inhibitors (ICI)–analysis of somatic alterations (SAs). Ann. Oncol. 2017, 28, v405. [Google Scholar] [CrossRef]
- Lehn, S.; Tobin, N.P.; Berglund, P.; Nilsson, K.; Sims, A.H.; Jirstrom, K.; Harkonen, P.; Lamb, R.; Landberg, G. Down-regulation of the oncogene cyclin D1 increases migratory capacity in breast cancer and is linked to unfavorable prognostic features. Am. J. Pathol. 2010, 177, 2886–2897. [Google Scholar] [CrossRef] [PubMed]
- Lenz, H.J.; Kahn, M. Safely targeting cancer stem cells via selective catenin coactivator antagonism. Cancer Sci. 2014, 105, 1087–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidtova, S.; Dorssers, L.C.J.; Kalavska, K.; Gillis, A.J.M.; Oosterhuis, J.W.; Stoop, H.; Miklikova, S.; Kozovska, Z.; Burikova, M.; Gercakova, K.; et al. Napabucasin overcomes cisplatin resistance in ovarian germ cell tumor-derived cell line by inhibiting cancer stemness. Cancer Cell Int. 2020, 20, 364. [Google Scholar] [CrossRef]
- Stehlik, P.; Paulikova, H.; Hunakova, L. Synthetic isothiocyanate indole-3-ethyl isothiocyanate (homoITC) enhances sensitivity of human ovarian carcinoma cell lines A2780 and A2780/CP to cisplatin. Neoplasma 2010, 57, 473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmidtova, S.; Kalavska, K.; Liskova, V.; Plava, J.; Miklikova, S.; Kucerova, L.; Matuskova, M.; Rojikova, L.; Cierna, Z.; Rogozea, A.; et al. Targeting of Deregulated Wnt/β-Catenin Signaling by PRI-724 and LGK974 Inhibitors in Germ Cell Tumor Cell Lines. Int. J. Mol. Sci. 2021, 22, 4263. https://doi.org/10.3390/ijms22084263
Schmidtova S, Kalavska K, Liskova V, Plava J, Miklikova S, Kucerova L, Matuskova M, Rojikova L, Cierna Z, Rogozea A, et al. Targeting of Deregulated Wnt/β-Catenin Signaling by PRI-724 and LGK974 Inhibitors in Germ Cell Tumor Cell Lines. International Journal of Molecular Sciences. 2021; 22(8):4263. https://doi.org/10.3390/ijms22084263
Chicago/Turabian StyleSchmidtova, Silvia, Katarina Kalavska, Veronika Liskova, Jana Plava, Svetlana Miklikova, Lucia Kucerova, Miroslava Matuskova, Lucia Rojikova, Zuzana Cierna, Adriana Rogozea, and et al. 2021. "Targeting of Deregulated Wnt/β-Catenin Signaling by PRI-724 and LGK974 Inhibitors in Germ Cell Tumor Cell Lines" International Journal of Molecular Sciences 22, no. 8: 4263. https://doi.org/10.3390/ijms22084263