
Prophylaxis and Treatment against Klebsiella pneumoniae: Current Insights on This Emerging Anti-Microbial Resistant Global Threat

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Klebsiella pneumoniae Diversity
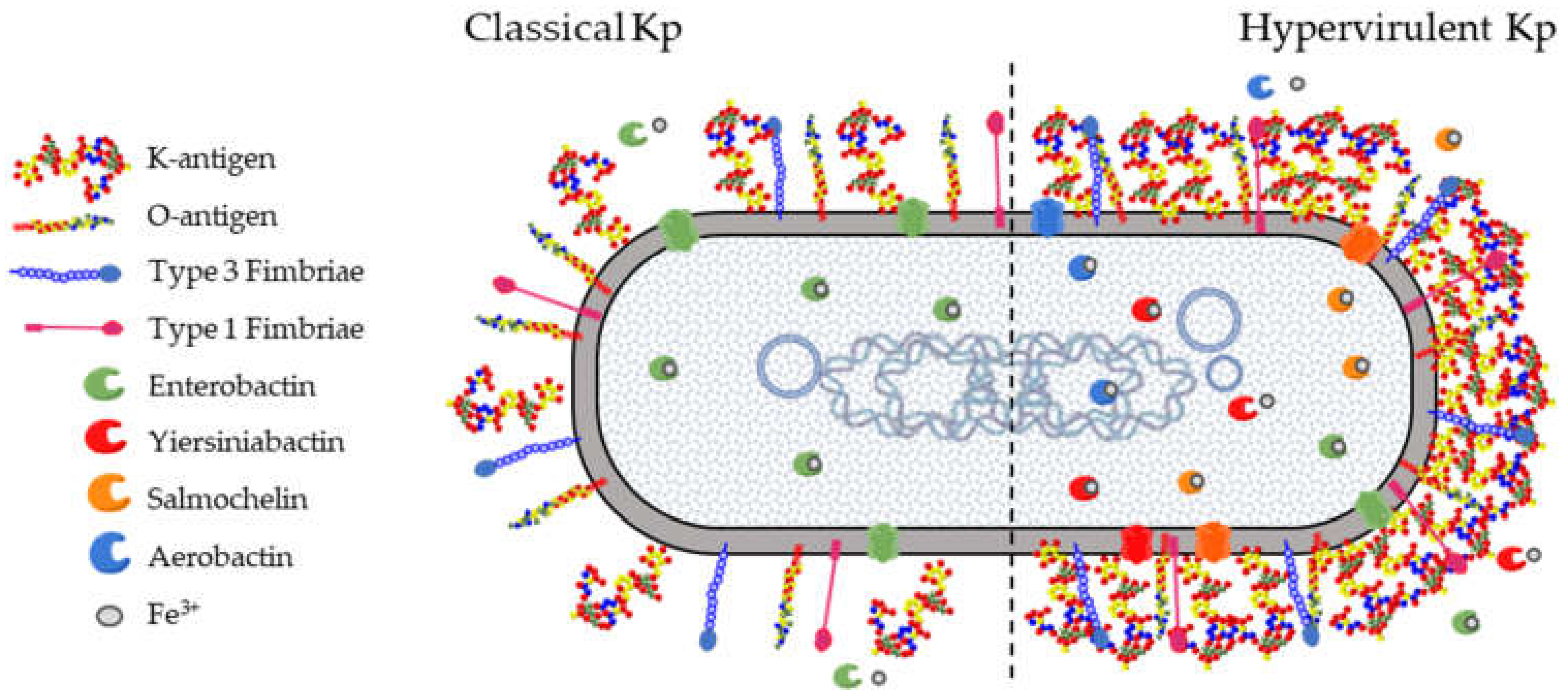
3. Klebsiella pneumoniae Virulence Factors and Key Target Antigens
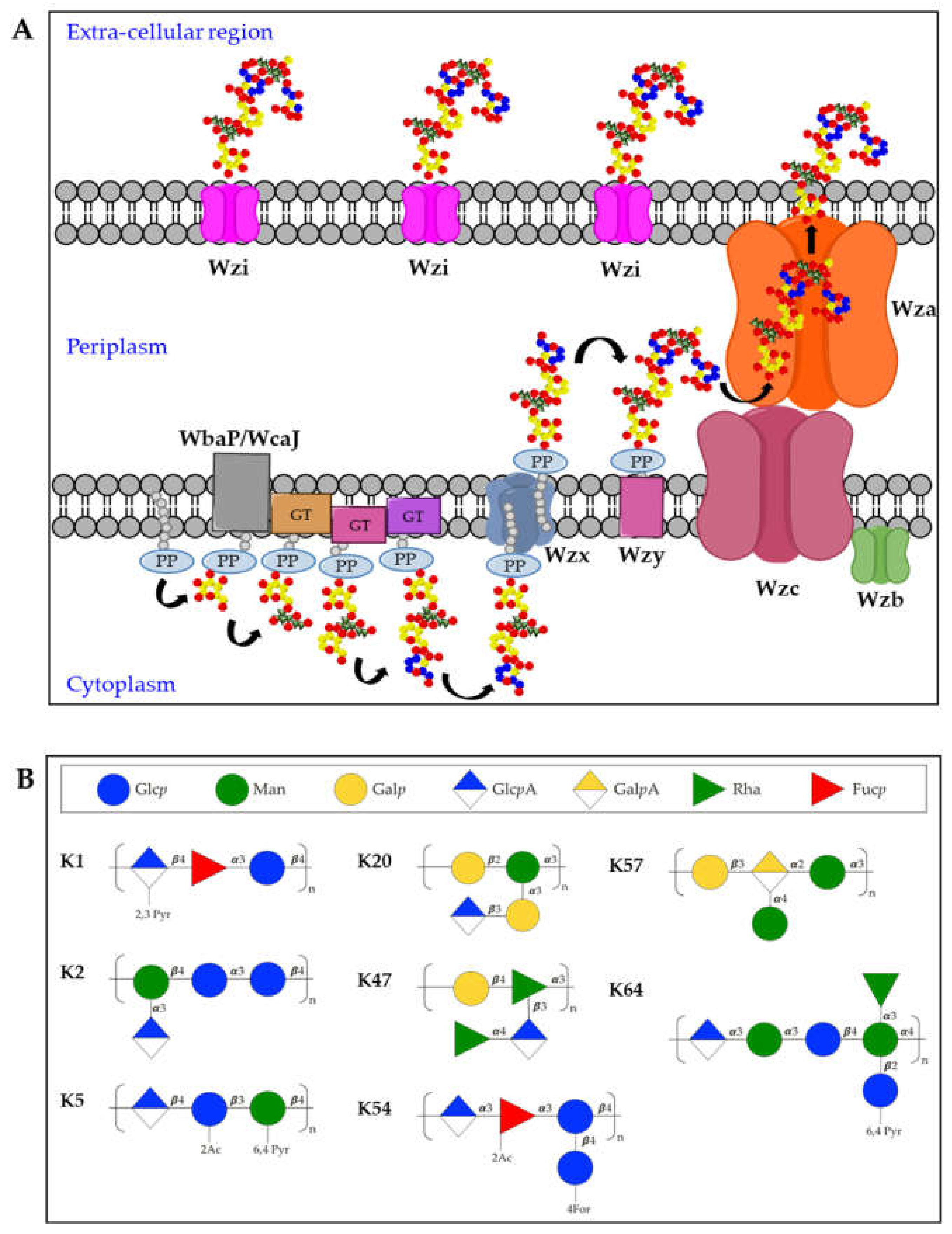
3.1. K-Antigens
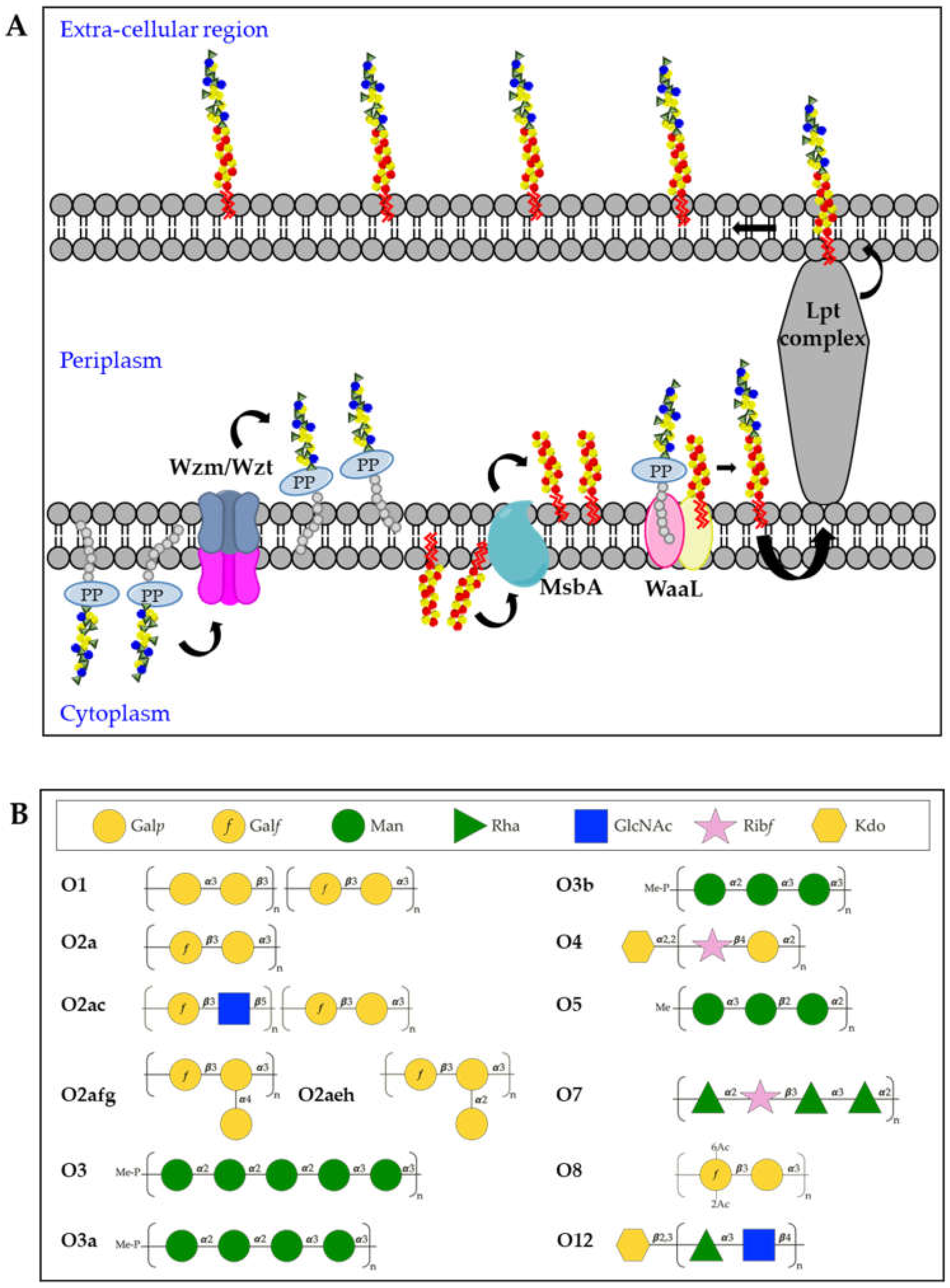
3.2. O-Antigens
3.3. Fimbriae
3.4. Siderophores
4. Vaccines and Monoclonal Antibodies (mAb) Strategies
4.1. K-Antigen Based Approaches
4.2. O-Antigen Based Approaches
4.3. Protein Based Approaches
5. Conclusions
6. Future Perspectives
Funding
Conflicts of Interest
References
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery and Development of New Antibiotics. Available online: https://www.who.int/medicines/publications/global-priority-list-antibiotic-resistant-bacteria/en/ (accessed on 13 April 2021).
- Rice, L.B. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- Paczosa, M.K.; Mecsas, J. Klebsiella pneumoniae: Going on the Offense with a Strong Defense. Microbiol. Mol. Biol. Rev. 2016, 80, 629–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol. Rev. 2017, 41, 252–275. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Holt, K.E. Klebsiella pneumoniae as a key trafficker of drug resistance genes from environmental to clinically important bacteria. Curr. Opin. Microbiol. 2018, 45, 131–139. [Google Scholar] [CrossRef]
- Lee, C.C.; Lee, C.H.; Hong, M.Y.; Hsieh, C.C.; Tang, H.J.; Ko, W.C. Propensity-matched analysis of the impact of extended-spectrum beta-lactamase production on adults with community-onset Escherichia coli, Klebsiella species, and Proteus mirabilis bacteremia. J. Microbiol. Immunol. Infect. 2018, 51, 519–526. [Google Scholar] [CrossRef]
- Shu, L.B.; Lu, Q.; Sun, R.H.; Lin, L.Q.; Sun, Q.L.; Hu, J.; Zhou, H.W.; Chan, E.W.; Chen, S.; Zhang, R. Prevalence and phenotypic characterization of carbapenem-resistant Klebsiella pneumoniae strains recovered from sputum and fecal samples of ICU patients in Zhejiang Province, China. Infect. Drug Resist. 2019, 12, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Han, J.H.; Goldstein, E.J.; Wise, J.; Bilker, W.B.; Tolomeo, P.; Lautenbach, E. Epidemiology of Carbapenem-Resistant Klebsiella pneumoniae in a Network of Long-Term Acute Care Hospitals. Clin. Infect. Dis. 2017, 64, 839–844. [Google Scholar] [CrossRef]
- Falagas, M.E.; Rafailidis, P.I.; Kofteridis, D.; Virtzili, S.; Chelvatzoglou, F.C.; Papaioannou, V.; Maraki, S.; Samonis, G.; Michalopoulos, A. Risk factors of carbapenem-resistant Klebsiella pneumoniae infections: A matched case control study. J. Antimicrob. Chemother. 2007, 60, 1124–1130. [Google Scholar] [CrossRef] [Green Version]
- Nordmann, P.; Cuzon, G.; Naas, T. The real threat of Klebsiella pneumoniae carbapenemase-producing bacteria. Lancet Infect. Dis. 2009, 9, 228–236. [Google Scholar] [CrossRef]
- Yigit, H.; Queenan, A.M.; Anderson, G.J.; Domenech-Sanchez, A.; Biddle, J.W.; Steward, C.D.; Alberti, S.; Bush, K.; Tenover, F.C. Novel carbapenem-hydrolyzing beta-lactamase, KPC-1, from a carbapenem-resistant strain of Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2001, 45, 1151–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Duin, D.; Perez, F.; Rudin, S.D.; Cober, E.; Hanrahan, J.; Ziegler, J.; Webber, R.; Fox, J.; Mason, P.; Richter, S.S.; et al. Surveillance of carbapenem-resistant Klebsiella pneumoniae: Tracking molecular epidemiology and outcomes through a regional network. Antimicrob. Agents Chemother. 2014, 58, 4035–4041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, D.; Toleman, M.A.; Giske, C.G.; Cho, H.S.; Sundman, K.; Lee, K.; Walsh, T.R. Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob. Agents Chemother. 2009, 53, 5046–5054. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, C.; Cormican, M.; Boo, T.W.; McGrath, E.; Slevin, B.; O’Gorman, A.; Commane, M.; Mahony, S.; O’Donovan, E.; Powell, J.; et al. An Irish outbreak of New Delhi metallo-beta-lactamase (NDM)-1 carbapenemase-producing Enterobacteriaceae: Increasing but unrecognized prevalence. J. Hosp. Infect. 2016, 94, 351–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch, T.; Lutgens, S.P.M.; Hermans, M.H.A.; Wever, P.C.; Schneeberger, P.M.; Renders, N.H.M.; Leenders, A.; Kluytmans, J.; Schoffelen, A.; Notermans, D.; et al. Outbreak of NDM-1-Producing Klebsiella pneumoniae in a Dutch Hospital, with Interspecies Transfer of the Resistance Plasmid and Unexpected Occurrence in Unrelated Health Care Centers. J. Clin. Microbiol. 2017, 55, 2380–2390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavoschi, L.; Forni, S.; Porretta, A.; Righi, L.; Pieralli, F.; Menichetti, F.; Falcone, M.; Gemignani, G.; Sani, S.; Vivani, P.; et al. Prolonged outbreak of New Delhi metallo-beta-lactamase-producing carbapenem-resistant Enterobacterales (NDM-CRE), Tuscany, Italy, 2018 to 2019. Eur. Surveill 2020, 25, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrer, A.; Poirel, L.; Eraksoy, H.; Cagatay, A.A.; Badur, S.; Nordmann, P. Spread of OXA-48-positive carbapenem-resistant Klebsiella pneumoniae isolates in Istanbul, Turkey. Antimicrob. Agents Chemother. 2008, 52, 2950–2954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avgoulea, K.; Di Pilato, V.; Zarkotou, O.; Sennati, S.; Politi, L.; Cannatelli, A.; Themeli-Digalaki, K.; Giani, T.; Tsakris, A.; Rossolini, G.M.; et al. Characterization of Extensively Drug-Resistant or Pandrug-Resistant Sequence Type 147 and 101 OXA-48-Producing Klebsiella pneumoniae Causing Bloodstream Infections in Patients in an Intensive Care Unit. Antimicrob. Agents Chemother. 2018, 62, 457. [Google Scholar] [CrossRef] [Green Version]
- Wrenn, C.; O’Brien, D.; Keating, D.; Roche, C.; Rose, L.; Ronayne, A.; Fenelon, L.; Fitzgerald, S.; Crowley, B.; Schaffer, K. Investigation of the first outbreak of OXA-48-producing Klebsiella pneumoniae in Ireland. J. Hosp. Infect. 2014, 87, 41–46. [Google Scholar] [CrossRef]
- Semin-Pelletier, B.; Cazet, L.; Bourigault, C.; Juvin, M.E.; Boutoille, D.; Raffi, F.; Hourmant, M.; Blancho, G.; Agard, C.; Connault, J.; et al. Challenges of controlling a large outbreak of OXA-48 carbapenemase-producing Klebsiella pneumoniae in a French university hospital. J. Hosp. Infect. 2015, 89, 248–253. [Google Scholar] [CrossRef]
- Borgmann, S.; Pfeifer, Y.; Becker, L.; Riess, B.; Siegmund, R.; Sagel, U. Findings from an outbreak of carbapenem-resistant Klebsiella pneumoniae emphasize the role of antibiotic treatment for cross transmission. Infection 2018, 46, 103–112. [Google Scholar] [CrossRef]
- Guo, L.; An, J.; Ma, Y.; Ye, L.; Luo, Y.; Tao, C.; Yang, J. Nosocomial Outbreak of OXA-48-Producing Klebsiella pneumoniae in a Chinese Hospital: Clonal Transmission of ST147 and ST383. PLoS ONE 2016, 11, e0160754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.M.; Guo, M.K.; Ke, S.C.; Lin, Y.P.; Li, C.R.; Vy Nguyen, H.T.; Wu, L.T. Emergence and nosocomial spread of ST11 carbapenem-resistant Klebsiella pneumoniae co-producing OXA-48 and KPC-2 in a regional hospital in Taiwan. J. Med. Microbiol. 2018, 67, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.C.; Cheng, D.L.; Lin, C.L. Klebsiella pneumoniae liver abscess associated with septic endophthalmitis. Arch. Intern. Med. 1986, 146, 1913–1916. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Liu, Y.C.; Lee, S.S.; Yen, M.Y.; Chen, Y.S.; Wang, J.H.; Wann, S.R.; Lin, H.H. Primary liver abscess due to Klebsiella pneumoniae in Taiwan. Clin. Infect. Dis. 1998, 26, 1434–1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lok, K.H.; Li, K.F.; Li, K.K.; Szeto, M.L. Pyogenic liver abscess: Clinical profile, microbiological characteristics, and management in a Hong Kong hospital. J. Microbiol. Immunol. Infect. 2008, 41, 483–490. [Google Scholar]
- Chung, D.R.; Lee, S.S.; Lee, H.R.; Kim, H.B.; Choi, H.J.; Eom, J.S.; Kim, J.S.; Choi, Y.H.; Lee, J.S.; Chung, M.H.; et al. Emerging invasive liver abscess caused by K1 serotype Klebsiella pneumoniae in Korea. J. Infect. 2007, 54, 578–583. [Google Scholar] [CrossRef]
- Russo, T.A.; Olson, R.; Fang, C.T.; Stoesser, N.; Miller, M.; MacDonald, U.; Hutson, A.; Barker, J.H.; La Hoz, R.M.; Johnson, J.R. Identification of Biomarkers for Differentiation of Hypervirulent Klebsiella pneumoniae from Classical K. pneumoniae. J. Clin. Microbiol. 2018, 56, 18. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.R.; Lin, T.L.; Chen, Y.C.; Chou, H.C.; Wang, J.T. The role of Klebsiella pneumoniae rmpA in capsular polysaccharide synthesis and virulence revisited. Microbiology 2011, 157, 3446–3457. [Google Scholar] [CrossRef]
- Holt, K.E.; Wertheim, H.; Zadoks, R.N.; Baker, S.; Whitehouse, C.A.; Dance, D.; Jenney, A.; Connor, T.R.; Hsu, L.Y.; Severin, J.; et al. Genomic analysis of diversity, population structure, virulence, and antimicrobial resistance in Klebsiella pneumoniae, an urgent threat to public health. Proc. Natl. Acad. Sci. USA 2015, 112, E3574–E3581. [Google Scholar] [CrossRef] [Green Version]
- Walker, K.A.; Miller, V.L. The intersection of capsule gene expression, hypermucoviscosity and hypervirulence in Klebsiella pneumoniae. Curr. Opin. Microbiol. 2020, 54, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.C.; Lu, M.C.; Hsueh, P.R. Hypervirulence and carbapenem resistance: Two distinct evolutionary directions that led high-risk Klebsiella pneumoniae clones to epidemic success. Expert Rev. Mol. Diagn. 2019, 19, 825–837. [Google Scholar] [CrossRef] [PubMed]
- Gu, D.; Dong, N.; Zheng, Z.; Lin, D.; Huang, M.; Wang, L.; Chan, E.W.; Shu, L.; Yu, J.; Zhang, R.; et al. A fatal outbreak of ST11 carbapenem-resistant hypervirulent Klebsiella pneumoniae in a Chinese hospital: A molecular epidemiological study. Lancet Infect. Dis. 2018, 18, 37–46. [Google Scholar] [CrossRef]
- Yao, B.; Xiao, X.; Wang, F.; Zhou, L.; Zhang, X.; Zhang, J. Clinical and molecular characteristics of multi-clone carbapenem-resistant hypervirulent (hypermucoviscous) Klebsiella pneumoniae isolates in a tertiary hospital in Beijing, China. Int. J. Infect. Dis. 2015, 37, 107–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson, M.; Stanton, R.A.; Ansari, U.; McAllister, G.; Chan, M.Y.; Sula, E.; Grass, J.E.; Duffy, N.; Anacker, M.L.; Witwer, M.L.; et al. Identification of a Carbapenemase-Producing Hypervirulent Klebsiella pneumoniae Isolate in the United States. Antimicrob. Agents Chemother. 2019, 63, 519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frost, I.; Van Boeckel, T.P.; Pires, J.; Craig, J.; Laxminarayan, R. Global geographic trends in antimicrobial resistance: The role of international travel. J. Travel Med. 2019, 26, 36. [Google Scholar] [CrossRef]
- Khan, M.S.; Durrance-Bagale, A.; Legido-Quigley, H.; Mateus, A.; Hasan, R.; Spencer, J.; Hanefeld, J. ‘LMICs as reservoirs of AMR’: A comparative analysis of policy discourse on antimicrobial resistance with reference to Pakistan. Health Policy Plan. 2019, 34, 178–187. [Google Scholar] [CrossRef]
- Founou, R.C.; Founou, L.L.; Essack, S.Y. Clinical and economic impact of antibiotic resistance in developing countries: A systematic review and meta-analysis. PLoS ONE 2017, 12, e0189621. [Google Scholar] [CrossRef] [Green Version]
- Zaidi, A.K.; Huskins, W.C.; Thaver, D.; Bhutta, Z.A.; Abbas, Z.; Goldmann, D.A. Hospital-acquired neonatal infections in developing countries. Lancet 2005, 365, 1175–1188. [Google Scholar] [CrossRef]
- Liu, L.; Oza, S.; Hogan, D.; Chu, Y.; Perin, J.; Zhu, J.; Lawn, J.E.; Cousens, S.; Mathers, C.; Black, R.E. Global, regional, and national causes of under-5 mortality in 2000-15: An updated systematic analysis with implications for the Sustainable Development Goals. Lancet 2016, 388, 3027–3035. [Google Scholar] [CrossRef] [Green Version]
- Okomo, U.; Akpalu, E.N.K.; Le Doare, K.; Roca, A.; Cousens, S.; Jarde, A.; Sharland, M.; Kampmann, B.; Lawn, J.E. Aetiology of invasive bacterial infection and antimicrobial resistance in neonates in sub-Saharan Africa: A systematic review and meta-analysis in line with the STROBE-NI reporting guidelines. Lancet Infect. Dis. 2019, 19, 1219–1234. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.W.; Blau, D.M.; Bassat, Q.; Onyango, D.; Kotloff, K.L.; Arifeen, S.E.; Mandomando, I.; Chawana, R.; Baillie, V.L.; Akelo, V.; et al. Initial findings from a novel population-based child mortality surveillance approach: A descriptive study. Lancet Glob. Health 2020, 8, e909–e919. [Google Scholar] [CrossRef]
- Bagley, S.T. Habitat association of Klebsiella species. Infect. Control. 1985, 6, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Brisse, S.; Verhoef, J. Phylogenetic diversity of Klebsiella pneumoniae and Klebsiella oxytoca clinical isolates revealed by randomly amplified polymorphic DNA, gyrA and parC genes sequencing and automated ribotyping. Int. J. Syst. Evol. Microbiol. 2001, 51, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Long, S.W.; Linson, S.E.; Ojeda Saavedra, M.; Cantu, C.; Davis, J.J.; Brettin, T.; Olsen, R.J. Whole-Genome Sequencing of Human Clinical Klebsiella pneumoniae Isolates Reveals Misidentification and Misunderstandings of Klebsiella pneumoniae, Klebsiella variicola, and Klebsiella quasipneumoniae. mSphere 2017, 2, 290. [Google Scholar] [CrossRef]
- Diancourt, L.; Passet, V.; Verhoef, J.; Grimont, P.A.; Brisse, S. Multilocus sequence typing of Klebsiella pneumoniae nosocomial isolates. J. Clin. Microbiol. 2005, 43, 4178–4182. [Google Scholar] [CrossRef] [Green Version]
- Bialek-Davenet, S.; Criscuolo, A.; Ailloud, F.; Passet, V.; Jones, L.; Delannoy-Vieillard, A.S.; Garin, B.; Le Hello, S.; Arlet, G.; Nicolas-Chanoine, M.H.; et al. Genomic definition of hypervirulent and multidrug-resistant Klebsiella pneumoniae clonal groups. Emerg Infect. Dis. 2014, 20, 1812–1820. [Google Scholar] [CrossRef]
- Wyres, K.L.; Holt, K.E. Klebsiella pneumoniae Population Genomics and Antimicrobial-Resistant Clones. Trends Microbiol. 2016, 24, 944–956. [Google Scholar] [CrossRef] [Green Version]
- Wyres, K.L.; Hawkey, J.; Hetland, M.A.K.; Fostervold, A.; Wick, R.R.; Judd, L.M.; Hamidian, M.; Howden, B.P.; Lohr, I.H.; Holt, K.E. Emergence and rapid global dissemination of CTX-M-15-associated Klebsiella pneumoniae strain ST307. J. Antimicrob. Chemother. 2019, 74, 577–581. [Google Scholar] [CrossRef] [Green Version]
- Shi, Q.; Lan, P.; Huang, D.; Hua, X.; Jiang, Y.; Zhou, J.; Yu, Y. Diversity of virulence level phenotype of hypervirulent Klebsiella pneumoniae from different sequence type lineage. BMC Microbiol. 2018, 18, 94. [Google Scholar] [CrossRef]
- Lin, J.C.; Koh, T.H.; Lee, N.; Fung, C.P.; Chang, F.Y.; Tsai, Y.K.; Ip, M.; Siu, L.K. Genotypes and virulence in serotype K2 Klebsiella pneumoniae from liver abscess and non-infectious carriers in Hong Kong, Singapore and Taiwan. Gut Pathog. 2014, 6, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyres, K.L.; Lam, M.M.C.; Holt, K.E. Population genomics of Klebsiella pneumoniae. Nat. Rev. Microbiol. 2020, 18, 344–359. [Google Scholar] [CrossRef] [PubMed]
- Follador, R.; Heinz, E.; Wyres, K.L.; Ellington, M.J.; Kowarik, M.; Holt, K.E.; Thomson, N.R. The diversity of Klebsiella pneumoniae surface polysaccharides. Microb. Genom. 2016, 2, e000073. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Wick, R.R.; Gorrie, C.; Jenney, A.; Follador, R.; Thomson, N.R.; Holt, K.E. Identification of Klebsiella capsule synthesis loci from whole genome data. Microb. Genom. 2016, 2, e000102. [Google Scholar] [CrossRef]
- Patro, L.P.P.; Sudhakar, K.U.; Rathinavelan, T. K-PAM: A unified platform to distinguish Klebsiella species K- and O-antigen types, model antigen structures and identify hypervirulent strains. Sci. Rep. 2020, 10, 16732. [Google Scholar] [CrossRef]
- Choi, M.; Hegerle, N.; Nkeze, J.; Sen, S.; Jamindar, S.; Nasrin, S.; Sen, S.; Permala-Booth, J.; Sinclair, J.; Tapia, M.D.; et al. The Diversity of Lipopolysaccharide (O) and Capsular Polysaccharide (K) Antigens of Invasive Klebsiella pneumoniae in a Multi-Country Collection. Front. Microbiol. 2020, 11, 1249. [Google Scholar] [CrossRef]
- Pennini, M.E.; De Marco, A.; Pelletier, M.; Bonnell, J.; Cvitkovic, R.; Beltramello, M.; Cameroni, E.; Bianchi, S.; Zatta, F.; Zhao, W.; et al. Immune stealth-driven O2 serotype prevalence and potential for therapeutic antibodies against multidrug resistant Klebsiella pneumoniae. Nat. Commun. 2017, 8, 1991. [Google Scholar] [CrossRef]
- Amako, K.; Meno, Y.; Takade, A. Fine structures of the capsules of Klebsiella pneumoniae and Escherichia coli K1. J. Bacteriol. 1988, 170, 4960–4962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, R.M.; Bachman, M.A. Colonization, Infection, and the Accessory Genome of Klebsiella pneumoniae. Front. Cell Infect. Microbiol. 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Y.J.; Lin, T.L.; Lin, Y.T.; Su, P.A.; Chen, C.T.; Hsieh, P.F.; Hsu, C.R.; Chen, C.C.; Hsieh, Y.C.; Wang, J.T. Identification of capsular types in carbapenem-resistant Klebsiella pneumoniae strains by wzc sequencing and implications for capsule depolymerase treatment. Antimicrob. Agents Chemother. 2015, 59, 1038–1047. [Google Scholar] [CrossRef] [Green Version]
- Patro, L.P.P.; Rathinavelan, T. Targeting the Sugary Armor of Klebsiella Species. Front. Cell Infect. Microbiol. 2019, 9, 367. [Google Scholar] [CrossRef]
- Rahn, A.; Drummelsmith, J.; Whitfield, C. Conserved organization in the cps gene clusters for expression of Escherichia coli group 1K antigens: Relationship to the colanic acid biosynthesis locus and the cps genes from Klebsiella pneumoniae. J. Bacteriol. 1999, 181, 2307–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachdeva, S.; Palur, R.V.; Sudhakar, K.U.; Rathinavelan, T. E. coli Group 1 Capsular Polysaccharide Exportation Nanomachinary as a Plausible Antivirulence Target in the Perspective of Emerging Antimicrobial Resistance. Front. Microbiol. 2017, 8, 70. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, C. Biosynthesis and assembly of capsular polysaccharides in Escherichia coli. Annu. Rev. Biochem. 2006, 75, 39–68. [Google Scholar] [CrossRef]
- Shu, H.Y.; Fung, C.P.; Liu, Y.M.; Wu, K.M.; Chen, Y.T.; Li, L.H.; Liu, T.T.; Kirby, R.; Tsai, S.F. Genetic diversity of capsular polysaccharide biosynthesis in Klebsiella pneumoniae clinical isolates. Microbiology 2009, 155, 4170–4183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fung, C.P.; Chang, F.Y.; Lee, S.C.; Hu, B.S.; Kuo, B.I.; Liu, C.Y.; Ho, M.; Siu, L.K. A global emerging disease of Klebsiella pneumoniae liver abscess: Is serotype K1 an important factor for complicated endophthalmitis? Gut 2002, 50, 420–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Struve, C.; Roe, C.C.; Stegger, M.; Stahlhut, S.G.; Hansen, D.S.; Engelthaler, D.M.; Andersen, P.S.; Driebe, E.M.; Keim, P.; Krogfelt, K.A. Mapping the Evolution of Hypervirulent Klebsiella pneumoniae. mBio 2015, 6, e00630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pomakova, D.K.; Hsiao, C.B.; Beanan, J.M.; Olson, R.; MacDonald, U.; Keynan, Y.; Russo, T.A. Clinical and phenotypic differences between classic and hypervirulent Klebsiella pneumonia: An emerging and under-recognized pathogenic variant. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Nassif, X.; Fournier, J.M.; Arondel, J.; Sansonetti, P.J. Mucoid phenotype of Klebsiella pneumoniae is a plasmid-encoded virulence factor. Infect. Immun. 1989, 57, 546–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.; Lv, J.; Niu, S.; Du, H.; Tang, Y.W.; Pitout, J.D.D.; Bonomo, R.A.; Kreiswirth, B.N.; Chen, L. Multiplex PCR Analysis for Rapid Detection of Klebsiella pneumoniae Carbapenem-Resistant (Sequence Type 258 [ST258] and ST11) and Hypervirulent (ST23, ST65, ST86, and ST375) Strains. J. Clin. Microbiol. 2018, 56, 731. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.L.; Ko, W.C.; Cheng, K.C.; Lee, C.C.; Lai, C.C.; Chuang, Y.C. Comparison of prevalence of virulence factors for Klebsiella pneumoniae liver abscesses between isolates with capsular K1/K2 and non-K1/K2 serotypes. Diagn Microbiol. Infect. Dis. 2008, 62, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Shon, A.S.; Bajwa, R.P.; Russo, T.A. Hypervirulent (hypermucoviscous) Klebsiella pneumoniae: A new and dangerous breed. Virulence 2013, 4, 107–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, B.R.; Ovchinnikova, O.G.; Kelly, S.D.; Williamson, M.L.; Butler, J.E.; Liu, B.; Wang, L.; Gou, X.; Follador, R.; Lowary, T.L.; et al. Molecular basis for the structural diversity in serogroup O2-antigen polysaccharides in Klebsiella pneumoniae. J. Biol. Chem. 2018, 293, 4666–4679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trautmann, M.; Ruhnke, M.; Rukavina, T.; Held, T.K.; Cross, A.S.; Marre, R.; Whitfield, C. O-antigen seroepidemiology of Klebsiella clinical isolates and implications for immunoprophylaxis of Klebsiella infections. Clin. Diagn. Lab. Immunol. 1997, 4, 550–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, D.S.; Mestre, F.; Alberti, S.; Hernandez-Alles, S.; Alvarez, D.; Domenech-Sanchez, A.; Gil, J.; Merino, S.; Tomas, J.M.; Benedi, V.J. Klebsiella pneumoniae lipopolysaccharide O typing: Revision of prototype strains and O-group distribution among clinical isolates from different sources and countries. J. Clin. Microbiol. 1999, 37, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Kalynych, S.; Morona, R.; Cygler, M. Progress in understanding the assembly process of bacterial O-antigen. FEMS Microbiol. Rev. 2014, 38, 1048–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regue, M.; Izquierdo, L.; Fresno, S.; Pique, N.; Corsaro, M.M.; Naldi, T.; De Castro, C.; Waidelich, D.; Merino, S.; Tomas, J.M. A second outer-core region in Klebsiella pneumoniae lipopolysaccharide. J. Bacteriol. 2005, 187, 4198–4206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fresno, S.; Jimenez, N.; Canals, R.; Merino, S.; Corsaro, M.M.; Lanzetta, R.; Parrilli, M.; Pieretti, G.; Regue, M.; Tomas, J.M. A second galacturonic acid transferase is required for core lipopolysaccharide biosynthesis and complete capsule association with the cell surface in Klebsiella pneumoniae. J. Bacteriol. 2007, 189, 1128–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuda, S.; Sherman, D.J.; Silhavy, T.J.; Ruiz, N.; Kahne, D. Lipopolysaccharide transport and assembly at the outer membrane: The PEZ model. Nat. Rev. Microbiol. 2016, 14, 337–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, B.R.; Whitfield, C. Molecular cloning of the rfb region of Klebsiella pneumoniae serotype O1:K20: The rfb gene cluster is responsible for synthesis of the D-galactan I O polysaccharide. J. Bacteriol. 1992, 174, 4614–4621. [Google Scholar] [CrossRef] [Green Version]
- Kelly, R.F.; Perry, M.B.; MacLean, L.L.; Whitfield, C. Structures of the O-antigens of Klebsiella serotypes 02 (2a,2e), 02 (2a,2e,2h), and 02 (2a,2f,2g), members of a family of related D-galactan O-antigens in Klebsiella spp. Innate Immun. 1995, 2, 131–140. [Google Scholar] [CrossRef]
- Kelly, R.F.; Severn, W.B.; Richards, J.C.; Perry, M.B.; MacLean, L.L.; Tomás, J.M.; Merino, S.; Whitfield, C. Structural variation in the O-specific polysaccharides of Klebsiella pneumoniae serotype O1 and O8 lipopolysaccharide: Evidence for clonal diversity in rfb genes. Mol. Microbiol. 1993, 10, 615–625. [Google Scholar] [CrossRef]
- Hsieh, P.F.; Wu, M.C.; Yang, F.L.; Chen, C.T.; Lou, T.C.; Chen, Y.Y.; Wu, S.H.; Sheu, J.C.; Wang, J.T. D-galactan II is an immunodominant antigen in O1 lipopolysaccharide and affects virulence in Klebsiella pneumoniae: Implication in vaccine design. Front. Microbiol. 2014, 5, 608. [Google Scholar] [CrossRef]
- Prehm, P.; Jann, B.; Jann, K. The O9 antigen of Escherichia coli. Structure of the polysaccharide chain. Eur. J. Biochem. 1976, 67, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Jansson, P.E.e.a. Structural studies of the O-antigen polysaccharides of Klebsiella O5 and Escherichia coli O8. Carbohydr. Res. 1985, 145, 59–66. [Google Scholar] [CrossRef]
- Greenfield, L.K.E.A. Biosynthesis of the polymannose lipopolysaccharide O-antigens from Escherichia coli serotypes O8 and O9a requires a unique combination of single- and multiple-active site mannosyltransferases. J. Biol. Chem. 2012, 287, 35078–35091. [Google Scholar] [CrossRef] [Green Version]
- Hagelueken, G.E.A. Structure of WbdD: A bifunctional kinase and methyltransferase that regulates the chain length of the O antigen in Escherichia coli O9a. Mol. Microbiol. 2012, 86, 730–742. [Google Scholar] [CrossRef] [Green Version]
- Vinogradov, E.E.A. Structures of lipopolysaccharides from Klebsiella pneumoniae. Eluicidation of the structure of the linkage region between core and polysaccharide O chain and identification of the residues at the non-reducing termini of the O chains. J. Biol. Chem. 2002, 277, 25070–25081. [Google Scholar] [CrossRef] [Green Version]
- Rollenske, T.; Szijarto, V.; Lukasiewicz, J.; Guachalla, L.M.; Stojkovic, K.; Hartl, K.; Stulik, L.; Kocher, S.; Lasitschka, F.; Al-Saeedi, M.; et al. Cross-specificity of protective human antibodies against Klebsiella pneumoniae LPS O-antigen. Nat. Immunol. 2018, 19, 617–624. [Google Scholar] [CrossRef]
- Duguid, J.P.; Smith, I.W.; Dempster, G.; Edmunds, P.N. Non-flagellar filamentous appendages (fimbriae) and haemagglutinating activity in Bacterium coli. J. Pathol. Bacteriol. 1955, 70, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Althouse, C.; Patterson, S.; Fedorka-Cray, P.; Isaacson, R.E. Type 1 fimbriae of Salmonella enterica serovar Typhimurium bind to enterocytes and contribute to colonization of swine in vivo. Infect. Immun. 2003, 71, 6446–6452. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.G.; Murphy, C.N.; Sippy, J.; Johnson, T.J.; Clegg, S. Type 3 fimbriae and biofilm formation are regulated by the transcriptional regulators MrkHI in Klebsiella pneumoniae. J. Bacteriol. 2011, 193, 3453–3460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, W.J.; Phan, G.; Waksman, G. Pilus biogenesis at the outer membrane of Gram-negative bacterial pathogens. Curr. Opin. Struct. Biol. 2012, 22, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Clegg, S.; Wilson, J.; Johnson, J. More than one way to control hair growth: Regulatory mechanisms in enterobacteria that affect fimbriae assembled by the chaperone/usher pathway. J. Bacteriol. 2011, 193, 2081–2088. [Google Scholar] [CrossRef] [Green Version]
- Morrissey, B.; Leney, A.C.; Toste Rego, A.; Phan, G.; Allen, W.J.; Verger, D.; Waksman, G.; Ashcroft, A.E.; Radford, S.E. The role of chaperone-subunit usher domain interactions in the mechanism of bacterial pilus biogenesis revealed by ESI-MS. Mol. Cell Proteom. 2012, 11, 15289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.C.; Huang, Y.J.; Fung, C.P.; Peng, H.L. Regulation of the Klebsiella pneumoniae Kpc fimbriae by the site-specific recombinase KpcI. Microbiology 2010, 156, 1983–1992. [Google Scholar] [CrossRef] [Green Version]
- Di Martino, P.; Livrelli, V.; Sirot, D.; Joly, B.; Darfeuille-Michaud, A. A new fimbrial antigen harbored by CAZ-5/SHV-4-producing Klebsiella pneumoniae strains involved in nosocomial infections. Infect. Immun. 1996, 64, 2266–2273. [Google Scholar] [CrossRef] [Green Version]
- Di Martino, P.; Sirot, D.; Joly, B.; Rich, C.; Darfeuille-Michaud, A. Relationship between adhesion to intestinal Caco-2 cells and multidrug resistance in Klebsiella pneumoniae clinical isolates. J. Clin. Microbiol. 1997, 35, 1499–1503. [Google Scholar] [CrossRef] [Green Version]
- Brinton, C.C., Jr. The structure, function, synthesis and genetic control of bacterial pili and a molecular model for DNA and RNA transport in gram negative bacteria. Trans. N. Y. Acad. Sci. 1965, 27, 1003–1054. [Google Scholar] [CrossRef]
- Zyla, D.S.; Prota, A.E.; Capitani, G.; Glockshuber, R. Alternative folding to a monomer or homopolymer is a common feature of the type 1 pilus subunit FimA from enteroinvasive bacteria. J. Biol. Chem. 2019, 294, 10553–10563. [Google Scholar] [CrossRef] [Green Version]
- Busch, A.; Phan, G.; Waksman, G. Molecular mechanism of bacterial type 1 and P pili assembly. Philos. Trans. A Math. Phys. Eng. Sci. 2015, 373, 153. [Google Scholar] [CrossRef] [PubMed]
- Lillington, J.; Geibel, S.; Waksman, G. Biogenesis and adhesion of type 1 and P pili. Biochim. Biophys. Acta 2014, 1840, 2783–2793. [Google Scholar] [CrossRef] [PubMed]
- Firon, N.; Ashkenazi, S.; Mirelman, D.; Ofek, I.; Sharon, N. Aromatic alpha-glycosides of mannose are powerful inhibitors of the adherence of type 1 fimbriated Escherichia coli to yeast and intestinal epithelial cells. Infect. Immun. 1987, 55, 472–476. [Google Scholar] [CrossRef] [Green Version]
- Firon, N.; Ofek, I.; Sharon, N. Carbohydrate-binding sites of the mannose-specific fimbrial lectins of enterobacteria. Infect. Immun. 1984, 43, 1088–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernhard, W.; Gbarah, A.; Sharon, N. Lectinophagocytosis of type 1 fimbriated (mannose-specific) Escherichia coli in the mouse peritoneum. J. Leukoc. Biol. 1992, 52, 343–348. [Google Scholar] [CrossRef]
- Sharon, N. Bacterial lectins, cell-cell recognition and infectious disease. FEBS Lett. 1987, 217, 145–157. [Google Scholar] [CrossRef] [Green Version]
- Struve, C.; Bojer, M.; Krogfelt, K.A. Characterization of Klebsiella pneumoniae type 1 fimbriae by detection of phase variation during colonization and infection and impact on virulence. Infect. Immun. 2008, 76, 4055–4065. [Google Scholar] [CrossRef] [Green Version]
- Rosen, D.A.; Pinkner, J.S.; Walker, J.N.; Elam, J.S.; Jones, J.M.; Hultgren, S.J. Molecular variations in Klebsiella pneumoniae and Escherichia coli FimH affect function and pathogenesis in the urinary tract. Infect. Immun. 2008, 76, 3346–3356. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.J.; Littmann, E.R.; Seok, R.; Leiner, I.M.; Taur, Y.; Peled, J.; van den Brink, M.; Ling, L.; Chen, L.; Kreiswirth, B.N.; et al. Genome-Wide Screening for Enteric Colonization Factors in Carbapenem-Resistant ST258 Klebsiella pneumoniae. mBio 2019, 10, 663. [Google Scholar] [CrossRef] [Green Version]
- Young, T.M.; Bray, A.S.; Nagpal, R.K.; Caudell, D.L.; Yadav, H.; Zafar, M.A. Animal Model To Study Klebsiella pneumoniae Gastrointestinal Colonization and Host-to-Host Transmission. Infect. Immun. 2020, 88, 128. [Google Scholar] [CrossRef]
- Allen, B.L.; Gerlach, G.F.; Clegg, S. Nucleotide sequence and functions of mrk determinants necessary for expression of type 3 fimbriae in Klebsiella pneumoniae. J. Bacteriol. 1991, 173, 916–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duguid, J.P. Fimbriae and adhesive properties in Klebsiella strains. J. Gen. Microbiol. 1959, 21, 271–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.J.; Liao, H.W.; Wu, C.C.; Peng, H.L. MrkF is a component of type 3 fimbriae in Klebsiella pneumoniae. Res. Microbiol. 2009, 160, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Sebghati, T.A.; Korhonen, T.K.; Hornick, D.B.; Clegg, S. Characterization of the type 3 fimbrial adhesins of Klebsiella strains. Infect. Immun. 1998, 66, 2887–2894. [Google Scholar] [CrossRef] [Green Version]
- Burmolle, M.; Bahl, M.I.; Jensen, L.B.; Sorensen, S.J.; Hansen, L.H. Type 3 fimbriae, encoded by the conjugative plasmid pOLA52, enhance biofilm formation and transfer frequencies in Enterobacteriaceae strains. Microbiology 2008, 154, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Ong, C.L.; Beatson, S.A.; McEwan, A.G.; Schembri, M.A. Conjugative plasmid transfer and adhesion dynamics in an Escherichia coli biofilm. Appl. Environ. Microbiol. 2009, 75, 6783–6791. [Google Scholar] [CrossRef] [Green Version]
- Tarkkanen, A.M.; Allen, B.L.; Westerlund, B.; Holthofer, H.; Kuusela, P.; Risteli, L.; Clegg, S.; Korhonen, T.K. Type V collagen as the target for type-3 fimbriae, enterobacterial adherence organelles. Mol. Microbiol. 1990, 4, 1353–1361. [Google Scholar] [CrossRef]
- Schroll, C.; Barken, K.B.; Krogfelt, K.A.; Struve, C. Role of type 1 and type 3 fimbriae in Klebsiella pneumoniae biofilm formation. BMC Microbiol. 2010, 10, 179. [Google Scholar] [CrossRef] [Green Version]
- Boddicker, J.D.; Anderson, R.A.; Jagnow, J.; Clegg, S. Signature-tagged mutagenesis of Klebsiella pneumoniae to identify genes that influence biofilm formation on extracellular matrix material. Infect. Immun. 2006, 74, 4590–4597. [Google Scholar] [CrossRef] [Green Version]
- Jagnow, J.; Clegg, S. Klebsiella pneumoniae MrkD-mediated biofilm formation on extracellular matrix- and collagen-coated surfaces. Microbiology 2003, 149, 2397–2405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foroohimanjili, F.; Mirzaie, A.; Hamdi, S.M.M.; Noorbazargan, H.; Hedayati Ch, M.; Dolatabadi, A.; Rezaie, H.; Bishak, F.M. Antibacterial, antibiofilm, and antiquorum sensing activities of phytosynthesized silver nanoparticles fabricated from Mespilus germanica extract against multidrug resistance of Klebsiella pneumoniae clinical strains. J. Basic. Microbiol. 2020, 60, 216–230. [Google Scholar] [CrossRef] [PubMed]
- Wilksch, J.J.; Yang, J.; Clements, A.; Gabbe, J.L.; Short, K.R.; Cao, H.; Cavaliere, R.; James, C.E.; Whitchurch, C.B.; Schembri, M.A.; et al. MrkH, a novel c-di-GMP-dependent transcriptional activator, controls Klebsiella pneumoniae biofilm formation by regulating type 3 fimbriae expression. PLoS Pathog. 2011, 7, e1002204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, D.A.; Twentyman, J.; Hunstad, D.A. High Levels of Cyclic Di-GMP in Klebsiella pneumoniae Attenuate Virulence in the Lung. Infect. Immun. 2018, 86, 647. [Google Scholar] [CrossRef] [Green Version]
- Arato, V.; Gasperini, G.; Giusti, F.; Ferlenghi, I.; Scarselli, M.; Leuzzi, R. Dual role of the colonization factor CD2831 in Clostridium difficile pathogenesis. Sci. Rep. 2019, 9, 5554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.C.; Zamorano-Sanchez, D.; Pagliai, F.A.; Park, J.H.; Floyd, K.A.; Lee, C.K.; Kitts, G.; Rose, C.B.; Bilotta, E.M.; Wong, G.C.L.; et al. Reciprocal c-di-GMP signaling: Incomplete flagellum biogenesis triggers c-di-GMP signaling pathways that promote biofilm formation. PLoS Genet. 2020, 16, e1008703. [Google Scholar] [CrossRef] [Green Version]
- Zamorano-Sanchez, D.; Xian, W.; Lee, C.K.; Salinas, M.; Thongsomboon, W.; Cegelski, L.; Wong, G.C.L.; Yildiz, F.H. Functional Specialization in Vibrio cholerae Diguanylate Cyclases: Distinct Modes of Motility Suppression and c-di-GMP Production. mBio 2019, 10, 670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miethke, M.; Marahiel, M.A. Siderophore-based iron acquisition and pathogen control. Microbiol. Mol. Biol. Rev. 2007, 71, 413–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neilands, J.B. Iron absorption and transport in microorganisms. Annu Rev. Nutr. 1981, 1, 27–46. [Google Scholar] [CrossRef]
- Russo, T.A.; Marr, C.M. Hypervirulent Klebsiella pneumoniae. Clin. Microbiol. Rev. 2019, 32, 1128. [Google Scholar] [CrossRef] [Green Version]
- Russo, T.A.; Olson, R.; Macdonald, U.; Metzger, D.; Maltese, L.M.; Drake, E.J.; Gulick, A.M. Aerobactin mediates virulence and accounts for increased siderophore production under iron-limiting conditions by hypervirulent (hypermucoviscous) Klebsiella pneumoniae. Infect. Immun. 2014, 82, 2356–2367. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, P.F.; Lin, T.L.; Lee, C.Z.; Tsai, S.F.; Wang, J.T. Serum-induced iron-acquisition systems and TonB contribute to virulence in Klebsiella pneumoniae causing primary pyogenic liver abscess. J. Infect. Dis. 2008, 197, 1717–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, M.; Tennant, S.M.; Simon, R.; Cross, A.S. Progress towards the development of Klebsiella vaccines. Expert Rev. Vaccines 2019, 18, 681–691. [Google Scholar] [CrossRef]
- Zurawski, D.V.; McLendon, M.K. Monoclonal Antibodies as an Antibacterial Approach Against Bacterial Pathogens. Antibiotics 2020, 9, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizgerd, J.P.; Skerrett, S.J. Animal models of human pneumonia. Am. J. Physiol. Lung Cell Mol. Physiol. 2008, 294, L387–L398. [Google Scholar] [CrossRef]
- Bengoechea, J.A.; Pessoa, J.S. Klebsiella pneumoniae infection biology: Living to counteract host defences. FEMS Microbiol. Rev. 2019, 43, 123–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berendt, R.F.; Magruder, R.D.; Frola, F.R. Treatment of Klebsiella pneumoniae respiratory tract infection of squirrel monkeys with aerosol administration of kanamycin. Am. J. Vet. Res. 1980, 41, 1492–1494. [Google Scholar] [PubMed]
- Berendt, R.F.; Knutsen, G.L.; Powanda, M.C. Nonhuman primate model for the study of respiratory Klebsiella pneumoniae infection. Infect. Immun. 1978, 22, 275–281. [Google Scholar] [CrossRef] [Green Version]
- Micoli, F.; Costantino, P.; Adamo, R. Potential targets for next generation antimicrobial glycoconjugate vaccines. FEMS Microbiol. Rev. 2018, 42, 388–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, J.M.; Rowley, D. Resistance to Klebsiella pneumoniae and the importance of two bacterial antigens. Aust. J. Exp. Biol. Med. Sci. 1982, 60, 629–641. [Google Scholar] [CrossRef]
- Cryz, S.J.F.E.; Germanier, R. Protection against fatal Klebsiella pneumoniae burn wound sepsis by passive transfer of anticapsular polysaccharide. Infect. Immun. 1984, 45, 139–142. [Google Scholar] [CrossRef] [Green Version]
- Cryz, S.J., Jr.; Furer, E.; Germanier, R. Safety and immunogenicity of Klebsiella pneumoniae K1 capsular polysaccharide vaccine in humans. J. Infect. Dis. 1985, 151, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Trautmann, M.; Cryz, S.J., Jr.; Sadoff, J.C.; Cross, A.S. A murine monoclonal antibody against Klebsiella capsular polysaccharide is opsonic in vitro and protects against experimental Klebsiella pneumoniae infection. Microb. Pathog. 1988, 5, 177–187. [Google Scholar] [CrossRef]
- Cryz, S.J., Jr.; Mortimer, P.; Cross, A.S.; Furer, E.; Germanier, R. Safety and immunogenicity of a polyvalent Klebsiella capsular polysaccharide vaccine in humans. Vaccine 1986, 4, 15–20. [Google Scholar] [CrossRef]
- Granstrom, M.; Wretlind, B.; Markman, B.; Cryz, S. Enzyme-linked immunosorbent assay to evaluate the immunogenicity of a polyvalent Klebsiella capsular polysaccharide vaccine in humans. J. Clin. Microbiol. 1988, 26, 2257–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cryz, S.J., Jr.; Mortimer, P.M.; Mansfield, V.; Germanier, R. Seroepidemiology of Klebsiella bacteremic isolates and implications for vaccine development. J. Clin. Microbiol. 1986, 23, 687–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cryz, S.J., Jr.; Furer, E.; Sadoff, J.C.; Fredeking, T.; Que, J.U.; Cross, A.S. Production and characterization of a human hyperimmune intravenous immunoglobulin against Pseudomonas aeruginosa and Klebsiella species. J. Infect. Dis. 1991, 163, 1055–1061. [Google Scholar] [CrossRef]
- Edelman, R.; Taylor, D.N.; Wasserman, S.S.; McClain, J.B.; Cross, A.S.; Sadoff, J.C.; Que, J.U.; Cryz, S.J. Phase 1 trial of a 24-valent Klebsiella capsular polysaccharide vaccine and an eight-valent Pseudomonas O-polysaccharide conjugate vaccine administered simultaneously. Vaccine 1994, 12, 1288–1294. [Google Scholar] [CrossRef]
- Donta, S.T.; Peduzzi, P.; Cross, A.S.; Sadoff, J.; Haakenson, C.; Cryz, S.J., Jr.; Kauffman, C.; Bradley, S.; Gafford, G.; Elliston, D.; et al. Immunoprophylaxis against klebsiella and pseudomonas aeruginosa infections. The Federal Hyperimmune Immunoglobulin Trial Study Group. J. Infect. Dis. 1996, 174, 537–543. [Google Scholar] [CrossRef] [Green Version]
- Zigterman, J.W.; van Dam, J.E.; Snippe, H.; Rotteveel, F.T.; Jansze, M.; Willers, J.M.; Kamerling, J.P.; Vliegenthart, J.F. Immunogenic properties of octasaccharide-protein conjugates derived from Klebsiella serotype 11 capsular polysaccharide. Infect. Immun. 1985, 47, 421–428. [Google Scholar] [CrossRef] [Green Version]
- Seeberger, P.H.; Pereira, C.L.; Khan, N.; Xiao, G.; Diago-Navarro, E.; Reppe, K.; Opitz, B.; Fries, B.C.; Witzenrath, M. A Semi-Synthetic Glycoconjugate Vaccine Candidate for Carbapenem-Resistant Klebsiella pneumoniae. Angew Chem. Int. Ed. Engl. 2017, 56, 13973–13978. [Google Scholar] [CrossRef] [Green Version]
- Feldman, M.F.; Bridwell, A.E.M.; Scott, N.E.; Vinogradov, E.; McKee, S.R.; Chavez, S.M.; Twentyman, J.; Stallings, C.L.; Rosen, D.A.; Harding, C.M. A promising bioconjugate vaccine against hypervirulent Klebsiella pneumoniae. Proc. Natl. Acad. Sci. USA 2019, 116, 18655–18663. [Google Scholar] [CrossRef] [Green Version]
- Diago-Navarro, E.; Motley, M.P.; Ruiz-Perez, G.; Yu, W.; Austin, J.; Seco, B.M.S.; Xiao, G.; Chikhalya, A.; Seeberger, P.H.; Fries, B.C. Novel, Broadly Reactive Anticapsular Antibodies against Carbapenem-Resistant Klebsiella pneumoniae Protect from Infection. mBio 2018, 9, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trautmann, M.; Held, T.K.; Cross, A.S. O-antigen seroepidemiology of Klebsiella clinical isolates and implications for immunoprophylaxis of Klebsiella infections. Vaccine 2004, 22, 818–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clements, A.; Jenney, A.W.; Farn, J.L.; Brown, L.E.; Deliyannis, G.; Hartland, E.L.; Pearse, M.J.; Maloney, M.B.; Wesselingh, S.L.; Wijburg, O.L.; et al. Targeting subcapsular antigens for prevention of Klebsiella pneumoniae infections. Vaccine 2008, 26, 5649–5653. [Google Scholar] [CrossRef] [PubMed]
- Chhibber, S.; Wadhwa, S.; Yadav, V. Protective role of liposome incorporated lipopolysaccharide antigen of Klebsiella pneumoniae in a rat model of lobar pneumonia. Jpn J. Infect. Dis. 2004, 57, 150–155. [Google Scholar] [PubMed]
- Chhibber, S.; Rani, M.; Vanashree, Y. Immunoprotective potential of polysaccharide-tetanus toxoid conjugate in Klebsiella pneumoniae induced lobar pneumonia in rats. Indian. J. Exp. Biol. 2005, 43, 40–45. [Google Scholar]
- Hegerle, N.; Choi, M.; Sinclair, J.; Amin, M.N.; Ollivault-Shiflett, M.; Curtis, B.; Laufer, R.S.; Shridhar, S.; Brammer, J.; Toapanta, F.R.; et al. Development of a broad spectrum glycoconjugate vaccine to prevent wound and disseminated infections with Klebsiella pneumoniae and Pseudomonas aeruginosa. PLoS ONE 2018, 13, e0203143. [Google Scholar] [CrossRef]
- Harduin-Lepers, A. Glyco25, XXV International Symposium on Glycoconjugates. Glycoconj. J. 2019, 36, 267–397. [Google Scholar] [CrossRef]
- Zhang, L.; Pan, C.; Feng, E.; Hua, X.; Yu, Y.; Wang, H.; Zhu, L. Biosynthesis of polysaccharide conjugate vaccines against Klebsiella pneumoniae serotype O2 strains. Sheng Wu Gong Cheng Xue Bao 2020, 36, 1899–1907. [Google Scholar] [CrossRef]
- Cohen, T.S.; Pelletier, M.; Cheng, L.; Pennini, M.E.; Bonnell, J.; Cvitkovic, R.; Chang, C.S.; Xiao, X.; Cameroni, E.; Corti, D.; et al. Anti-LPS antibodies protect against Klebsiella pneumoniae by empowering neutrophil-mediated clearance without neutralizing TLR4. JCI Insight 2017, 2, 774. [Google Scholar] [CrossRef] [Green Version]
- Guachalla, L.M.; Stojkovic, K.; Hartl, K.; Kaszowska, M.; Kumar, Y.; Wahl, B.; Paprotka, T.; Nagy, E.; Lukasiewicz, J.; Nagy, G.; et al. Discovery of monoclonal antibodies cross-reactive to novel subserotypes of K. pneumoniae O3. Sci. Rep. 2017, 7, 6635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavender, H.; Jagnow, J.J.; Clegg, S. Klebsiella pneumoniae type 3 fimbria-mediated immunity to infection in the murine model of respiratory disease. Int. J. Med. Microbiol. 2005, 295, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Chang, C.S.; Pennini, M.; Pelletier, M.; Rajan, S.; Zha, J.; Chen, Y.; Cvitkovic, R.; Sadowska, A.; Heidbrink, T.J.; et al. Target-Agnostic Identification of Functional Monoclonal Antibodies Against Klebsiella pneumoniae Multimeric MrkA Fimbrial Subunit. J. Infect. Dis. 2016, 213, 1800–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witkowska, D.; Mieszała, M.; Gamian, A.; Staniszewska, M.; Czarny, A.; Przondo-Mordarska, A.; Jaquinod, M.; Forest, E. Major structural proteins of type 1 and type 3 Klebsiella fimbriae are effective protein carriers and immunogens in conjugates as revealed from their immunochemical characterization. FEMS Immunol. Med. Microbiol. 2005, 45, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Zargaran, F.N.; Akya, A.; Rezaeian, S.; Ghadiri, K.; Lorestani, R.C.; Madanchi, H.; Safaei, S.; Rostamian, M. B Cell Epitopes of Four Fimbriae Antigens of Klebsiella pneumoniae: A Comprehensive In Silico Study for Vaccine Development. Int. J. Pept. Res. Ther. 2020, 134, 1–12. [Google Scholar] [CrossRef]
- Dar, H.A.; Zaheer, T.; Shehroz, M.; Ullah, N.; Naz, K.; Muhammad, S.A.; Zhang, T.; Ali, A. Immunoinformatics-Aided Design and Evaluation of a Potential Multi-Epitope Vaccine against Klebsiella Pneumoniae. Vaccines 2019, 7, 88. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.X.; Yang, Y.; de Souza, M.E.B., Jr.; do Carmo, S.J.; Bicalho, R.C. Development and evaluation of a new recombinant protein vaccine (YidR) against Klebsiella pneumoniae infection. Vaccine 2020, 38, 4640–4648. [Google Scholar] [CrossRef]
- Micoli, F.; Bagnoli, F.; Rappuoli, R.; Serruto, D. The role of vaccines in combatting antimicrobial resistance. Nat. Rev. Microbiol. 2021, 3, 506. [Google Scholar] [CrossRef]
- Szijarto, V.; Guachalla, L.M.; Hartl, K.; Varga, C.; Badarau, A.; Mirkina, I.; Visram, Z.C.; Stulik, L.; Power, C.A.; Nagy, E.; et al. Endotoxin neutralization by an O-antigen specific monoclonal antibody: A potential novel therapeutic approach against Klebsiella pneumoniae ST258. Virulence 2017, 8, 1203–1215. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.H.; Choi, H.I.; Hong, S.W.; Kim, K.S.; Gho, Y.S.; Jeon, S.G. Vaccination with Klebsiella pneumoniae-derived extracellular vesicles protects against bacteria-induced lethality via both humoral and cellular immunity. Exp. Mol. Med. 2015, 47, e183. [Google Scholar] [CrossRef] [Green Version]
- Micoli, F.; Alfini, R.; Di Benedetto, R.; Necchi, F.; Schiavo, F.; Mancini, F.; Carducci, M.; Palmieri, E.; Balocchi, C.; Gasperini, G.; et al. GMMA Is a Versatile Platform to Design Effective Multivalent Combination Vaccines. Vaccines 2020, 8, 540. [Google Scholar] [CrossRef] [PubMed]
- Opoku-Temeng, C.; Kobayashi, S.D.; DeLeo, F.R. Klebsiella pneumoniae capsule polysaccharide as a target for therapeutics and vaccines. Comput. Struct. Biotechnol. J. 2019, 17, 1360–1366. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arato, V.; Raso, M.M.; Gasperini, G.; Berlanda Scorza, F.; Micoli, F. Prophylaxis and Treatment against Klebsiella pneumoniae: Current Insights on This Emerging Anti-Microbial Resistant Global Threat. Int. J. Mol. Sci. 2021, 22, 4042. https://doi.org/10.3390/ijms22084042
Arato V, Raso MM, Gasperini G, Berlanda Scorza F, Micoli F. Prophylaxis and Treatment against Klebsiella pneumoniae: Current Insights on This Emerging Anti-Microbial Resistant Global Threat. International Journal of Molecular Sciences. 2021; 22(8):4042. https://doi.org/10.3390/ijms22084042
Chicago/Turabian StyleArato, Vanessa, Maria Michelina Raso, Gianmarco Gasperini, Francesco Berlanda Scorza, and Francesca Micoli. 2021. "Prophylaxis and Treatment against Klebsiella pneumoniae: Current Insights on This Emerging Anti-Microbial Resistant Global Threat" International Journal of Molecular Sciences 22, no. 8: 4042. https://doi.org/10.3390/ijms22084042