
Oxidative Stress-Induced Sirtuin1 Downregulation Correlates to HIF-1α, GLUT-1, and VEGF-A Upregulation in Th1 Autoimmune Hashimoto’s Thyroiditis

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
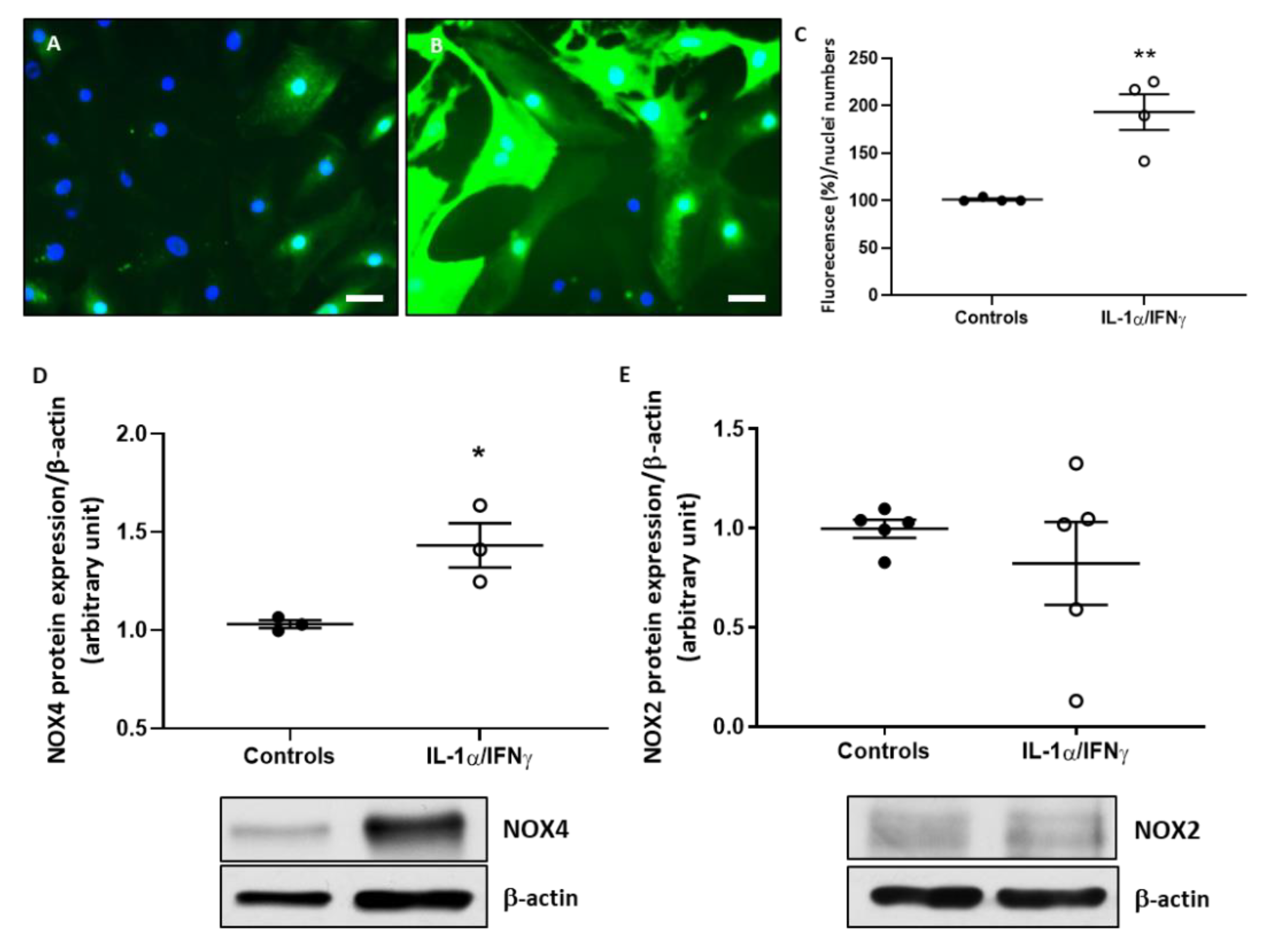
2.1. Th1 Cytokines Increase ROS Production and NOX4 Protein Expression in Human Thyroid Cells, without Changing NOX2 Protein Expression
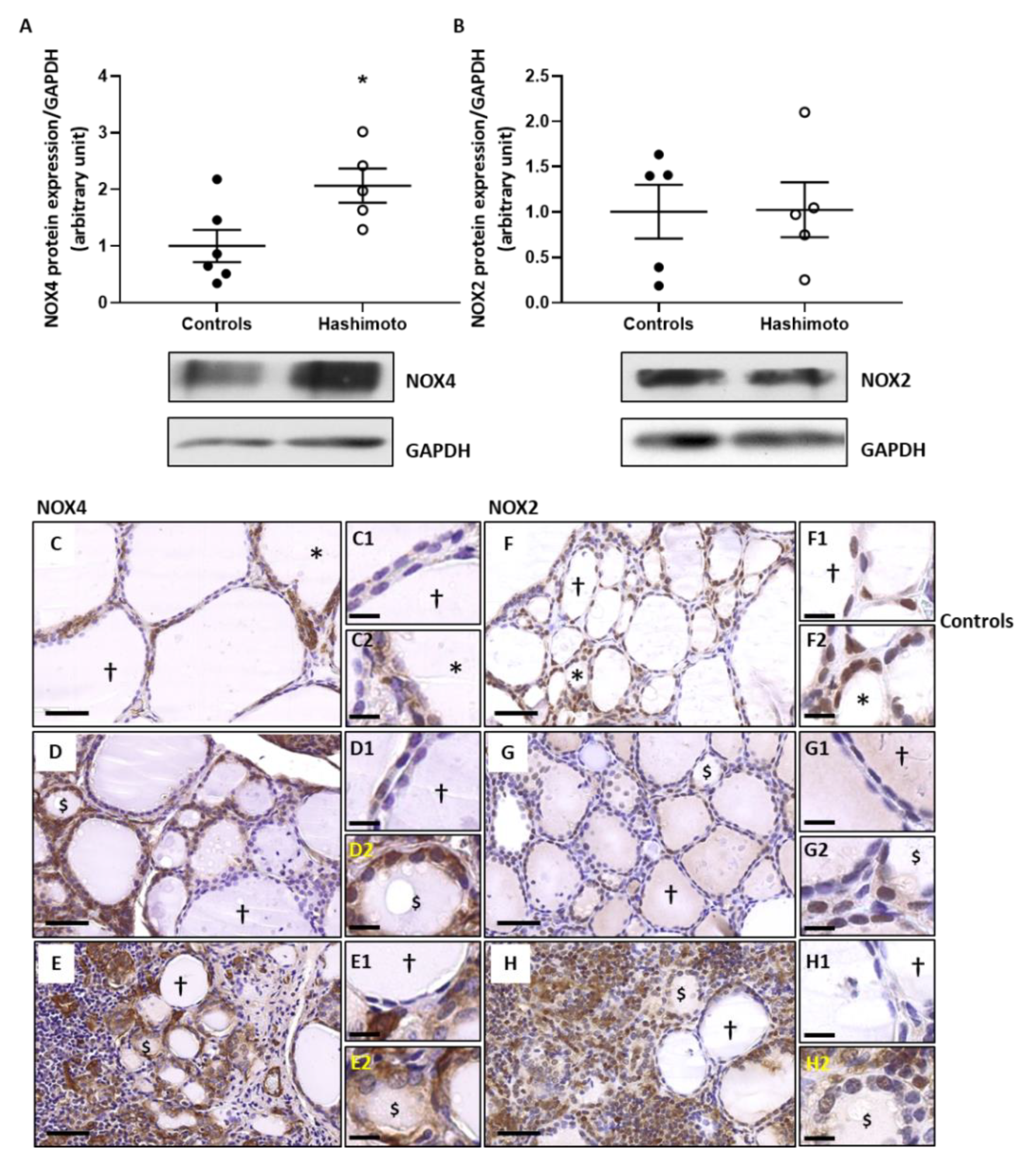
2.2. NOX4 Protein Expression Is Increased in HT Thyroid Samples without Any Change of NOX2 Protein Expression
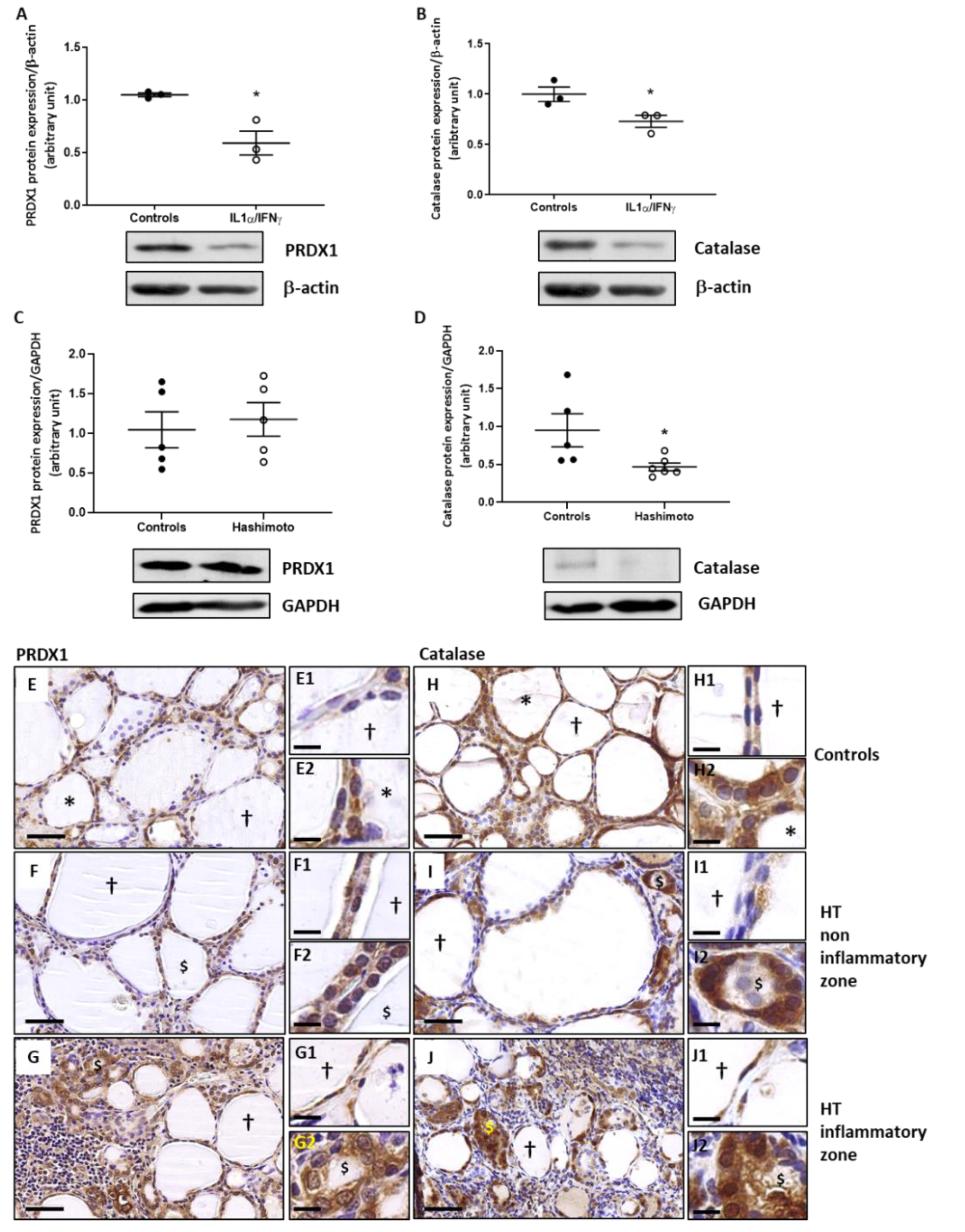
2.3. The Expression of PRDX1 and Catalase Is Decreased by Th1 Cytokines in Human Thyroid Cells and the Expression of Catalase Is Lowered in HT Thyroid Samples
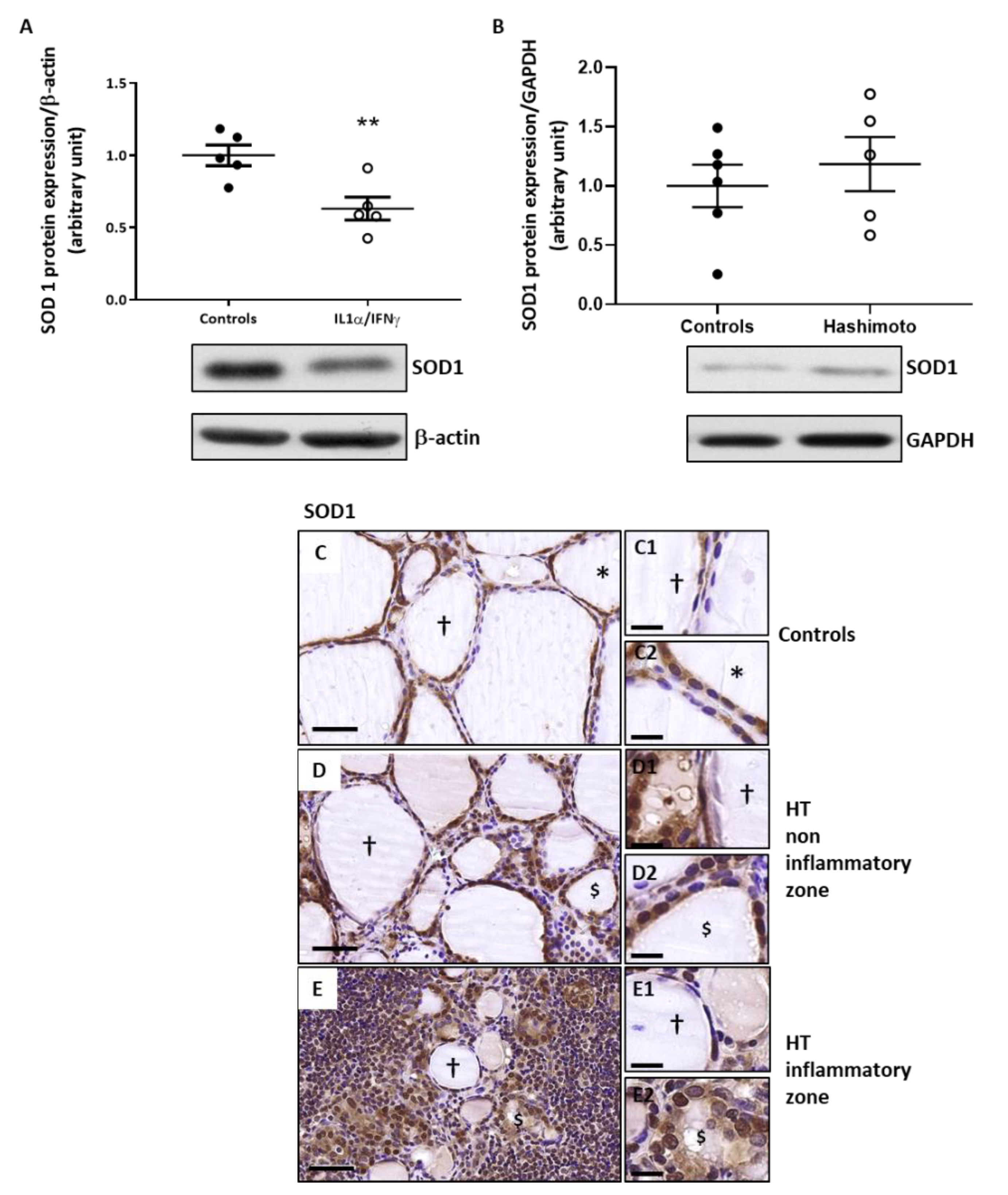
2.4. The Expression of SOD1 Is Decreased by Th1 Cytokines in Human Thyroid Cells and Is Unchanged in HT Thyroid Samples
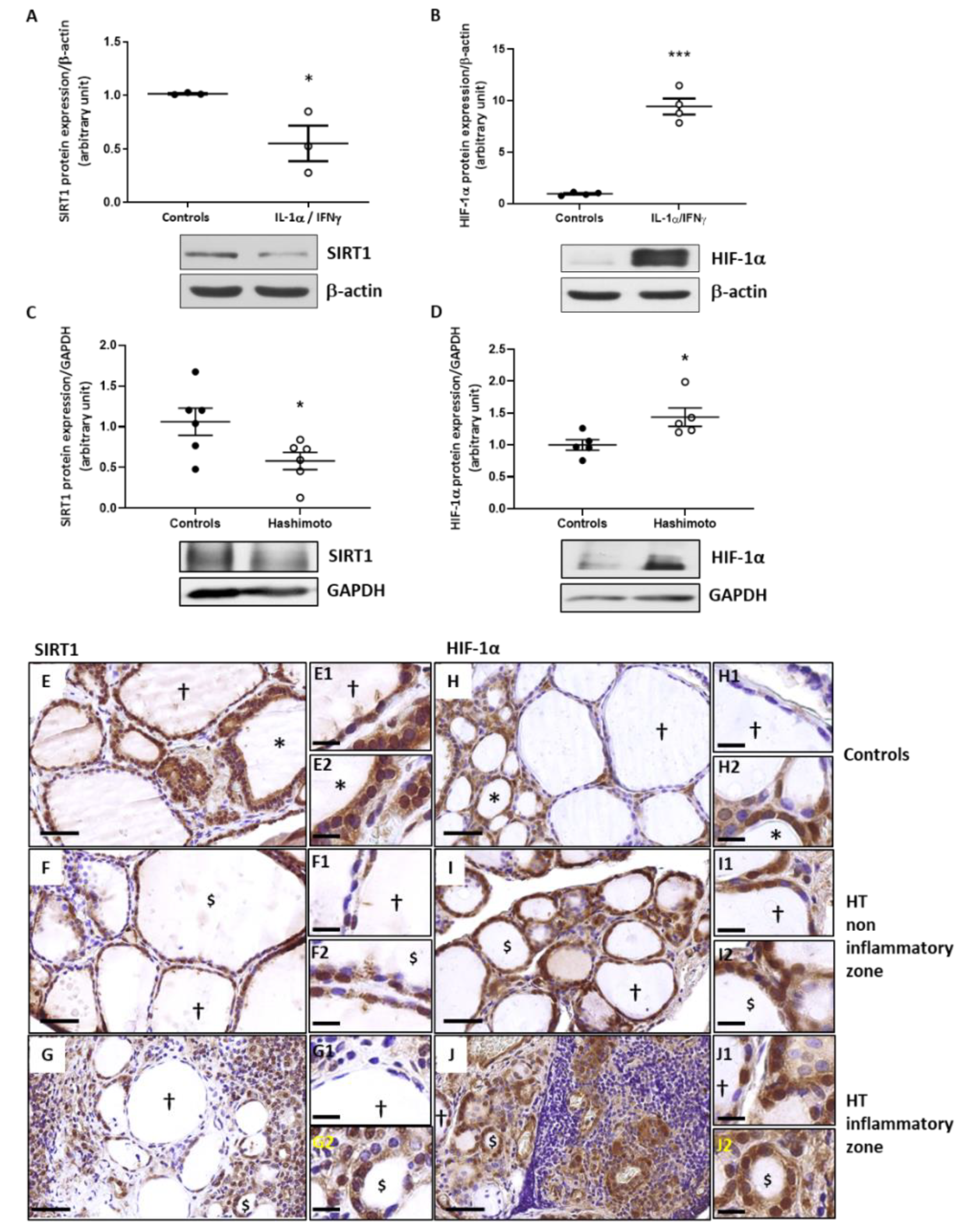
2.5. In Th1 Cytokines-Treated Human Thyroid Cells, as Well as in HT Thyroid Samples, the Expression of SIRT1 Is Decreased While the Expression of HIF-1α Is Increased
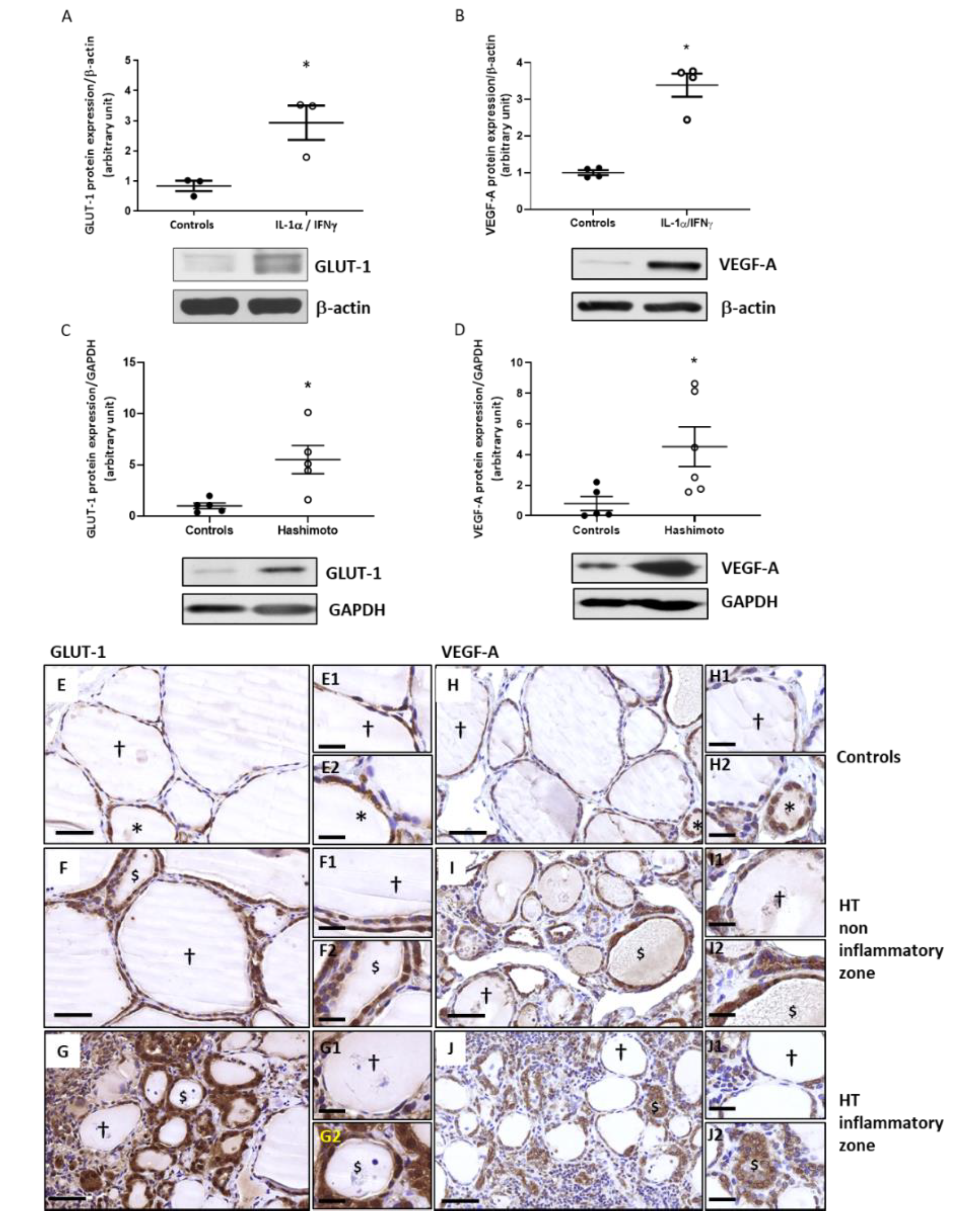
2.6. The Expression of GLUT-1 and VEGF-A Is Increased by Th1 Cytokines in Primary Cultures of Human Thyrocytes and in HT Thyroid Samples
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Primary Cell Cultures
4.3. Immunohistochemistry
4.4. Western Blotting
4.5. Quantification of ROS Levels Using CM-H2DCFDA
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Cav-1 | Caveolin-1 (Cav-1) |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| GLUT | Glucose Transporter (GLUT) |
| HIF | Hypoxia-inducible-factor (HIF) |
| HT | Hashimoto’s thyroiditis (HT) |
| IFN | Interferon (IFN) |
| IHC | Immunohistochemistry (IHC) |
| IL | Interleukin (IL) |
| NOX | NADPH-oxidase (NOX) |
| OS | Oxidative stress (OS) |
| PRDX | Peroxiredoxin (PRDX) |
| PTC | Papillary thyroid cancers (PTC) |
| ROS | Reactive oxygen species (ROS) |
| SEM | Standard error of the mean |
| SIRT | Sirtuin (SIRT) |
| SOD | Superoxide dismutase (SOD) |
| Th1 | T helper type one (Th1) |
| TNF | Tumor necrosis factor (TNF) |
| VEGF | Vascular endothelial growth factor (VEGF) |
| WB | Western Blot (WB) |
References
- Pearce, E.N.; Farwell, A.P.; Braverman, L.E. Thyroiditis. N. Engl. J. Med. 2003, 348, 2646–2655. [Google Scholar] [CrossRef] [PubMed]
- Singer, P.A. Thyroiditis. Acute, subacute, and chronic. Med. Clin. N. Am. 1991, 75, 61–77. [Google Scholar] [CrossRef]
- Phenekos, C.; Vryonidou, A.; Gritzapis, A.D.; Baxevanis, C.N.; Goula, M.; Papamichail, M. Th1 and Th2 serum cytokine profiles characterize patients with Hashimoto’s thyroiditis (Th1) and Graves’ disease (Th2). Neuroimmunomodulation 2004, 11, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Giannakou, M.; Saltiki, K.; Mantzou, E.; Loukari, E.; Philippou, G.; Terzidis, K.; Lili, K.; Stavrianos, C.; Kyprianou, M.; Alevizaki, M. RAGE polymorphisms and oxidative stress levels in Hashimoto’s thyroiditis. Eur. J. Clin. Investig. 2017, 47, 341–347. [Google Scholar] [CrossRef]
- Ruggeri, R.M.; Vicchio, T.M.; Cristani, M.; Certo, R.; Caccamo, D.; Alibrandi, A.; Giovinazzo, S.; Saija, A.; Campenni, A.; Trimarchi, F.; et al. Oxidative stress and advanced glycation end products in Hashimoto’s thyroiditis. Thyroid 2016, 26, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Ates, I.; Yilmaz, F.M.; Altay, M.; Yilmaz, N.; Berker, D.; Guler, S. The relationship between oxidative stress and autoimmunity in Hashimoto’s thyroiditis. Eur. J. Endocrinol. 2015, 173, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Szanto, I.; Pusztaszeri, M.; Mavromati, M. H(2)O(2) Metabolism in normal thyroid cells and in thyroid tumorigenesis: Focus on NADPH oxidases. Antioxidants 2019, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Marique, L.; Van Regemorter, V.; Gérard, A.C.; Craps, J.; Senou, M.; Marbaix, E.; Rahier, J.; Daumerie, C.; Mourad, M.; Lengelé, B.; et al. The expression of dual oxidase, thyroid peroxidase, and caveolin-1 differs according to the type of immune response (TH1/TH2) involved in thyroid autoimmune disorders. J. Clin. Endocrinol. Metab. 2014, 99, 1722–1732. [Google Scholar] [CrossRef] [Green Version]
- Werion, A.; Joris, V.; Hepp, M.; Papasokrati, L.; Marique, L.; de Ville de Goyet, C.; Van Regemorter, V.; Mourad, M.; Lengelé, B.; Daumerie, C.; et al. Pioglitazone, a PPARgamma agonist, upregulates the expression of Caveolin-1 and Catalase, essential for thyroid cell homeostasis: A clue to the pathogenesis of Hashimoto’s thyroiditis. Thyroid 2016, 26, 1320–1331. [Google Scholar] [CrossRef]
- Colin, I.M.; Poncin, S.; Leveque, P.; Gallez, B.; Gérard, A.C. Differential regulation of the production of reactive oxygen species in Th1 cytokine-treated thyroid cells. Thyroid 2014, 24, 441–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weyemi, U.; Caillou, B.; Talbot, M.; Ameziane-El-Hassani, R.; Lacroix, L.; Lagent-Chevallier, O.; Al Ghuzlan, A.; Roos, D.; Bidart, J.M.; Virion, A.; et al. Intracellular expression of reactive oxygen species-generating NADPH oxidase NOX4 in normal and cancer thyroid tissues. Endocr. Relat. Cancer 2010, 17, 27–37. [Google Scholar] [CrossRef]
- Weyemi, U.; Lagente-Chevallier, O.; Boufraqech, M.; Prenois, F.; Courtin, F.; Caillou, B.; Talbot, M.; Dardalhon, M.; Al Ghuzlan, A.; Bidart, J.M.; et al. ROS-generating NADPH oxidase NOX4 is a critical mediator in oncogenic H-Ras-induced DNA damage and subsequent senescence. Oncogene 2012, 31, 1117–1129. [Google Scholar] [CrossRef] [Green Version]
- Subhi, O.; Schulten, H.J.; Bagatian, N.; Al-Dayini, R.; Karim, S.; Bakhashab, S.; Alotibi, R.; Al-Ahmadi, A.; Ata, M.; Elaimi, A.; et al. relationship between Hashimoto’s thyroiditis and papillary thyroid carcinoma with coexisting Hashimoto’s thyroiditis. PLoS ONE 2020, 15, e0234566. [Google Scholar] [CrossRef] [PubMed]
- Tang, P.; Dang, H.; Huang, J.; Xu, T.; Yuan, P.; Hu, J.; Sheng, J.F. NADPH oxidase NOX4 is a glycolytic regulator through mROS-HIF1alpha axis in thyroid carcinomas. Sci. Rep. 2018, 8, 15897. [Google Scholar] [CrossRef]
- Gérard, A.C.; Poncin, S.; Audinot, J.N.; Denef, J.F.; Colin, I.M. Iodide deficiency-induced angiogenic stimulus in the thyroid occurs via HIF- and ROS-dependent VEGF-A secretion from thyrocytes. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E1414–E1422. [Google Scholar] [CrossRef]
- Craps, J.; Wilvers, C.; Joris, V.; De Jongh, B.; Vanderstraeten, J.; Lobysheva, I.; Balligand, J.L.; Sonveaux, P.; Gilon, P.; Many, M.C.; et al. Involvement of nitric oxide in iodine deficiency-induced microvascular remodeling in the thyroid gland: Role of nitric oxide synthase 3 and ryanodine receptors. Endocrinology 2015, 156, 707–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, J.M.; Santos, S.C.; Espadinha, C.; Correia, I.; Vag, T.; Casalou, C.; Cavaco, B.M.; Catarino, A.L.; Dias, S.; Leite, V. Expression of vascular endothelial growth factor (VEGF) and its receptors in thyroid carcinomas of follicular origin: A potential autocrine loop. Eur. J. Endocrinol. 2005, 153, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Crosstalk between oxidative stress and SIRT1: Impact on the aging process. Int. J. Mol. Sci. 2013, 14, 3834–3859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, C.; Morrison, A.; Mattison, S.; Qian, S.; Bryniarski, M.; Rankin, B.; Wang, J.; Thomas, D.P.; Li, J. Impaired SIRT1 nucleocytoplasmic shuttling in the senescent heart during ischemic stress. FASEB J. 2013, 27, 4332–4342. [Google Scholar] [CrossRef] [Green Version]
- Chung, S.; Yao, H.; Caito, S.; Hwang, J.W.; Arunachalam, G.; Rahman, I. Regulation of SIRT1 in cellular functions: Role of polyphenols. Arch. Biochem. Biophys. 2010, 501, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Al-Khaldi, A.; Sultan, S. The expression of sirtuins, superoxide dismutase, and lipid peroxidation status in peripheral blood from patients with diabetes and hypothyroidism. BMC. Endocr. Disord. 2019, 19, 19. [Google Scholar] [CrossRef]
- Roehlen, N.; Doering, C.; Hansmann, M.L.; Gruenwald, F.; Vorlaender, C.; Bechstein, W.O.; Holzer, K.; Badenhoop, K.; Penna-Martinez, M. Vitamin D, FOXO3a, and Sirtuin1 in Hashimoto’s thyroiditis and differentiated thyroid cancer. Front. Endocrinol. 2018, 9, 527. [Google Scholar] [CrossRef] [PubMed]
- Gérard, A.C.; Many, M.C.; Daumerie, C.; Costagliola, S.; Miot, F.; de Vijlder, J.J.; Colin, I.M.; Denef, J.F. Structural changes in the angiofollicular units between active and hypofunctioning follicles align with differences in the epithelial expression of newly discovered proteins involved in iodine transport and organification. J. Clin. Endocrinol. Metab. 2002, 87, 1291–1299. [Google Scholar] [CrossRef] [Green Version]
- Zarzuelo, M.J.; López-Sepúlveda, R.; Sánchez, M.; Romero, M.; Gómez-Guzmán, M.; Ungvary, Z.; Pérez-Vizcaíno, F.; Jiménez, R.; Duarte, J. SIRT1 inhibits NADPH oxidase activation and protects endothelial function in the rat aorta: Implications for vascular aging. Biochem. Pharmacol. 2013, 85, 1288–1296. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Huang, Q.; Zeng, Z.; Wu, J.; Zhang, Y.; Chen, Z. Sirt1 inhibits oxidative stress in vascular endothelial cells. Oxid. Med. Cell Longev. 2017, 2017, 7543973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmatz, R.; Perreira, L.B.; Stefanello, N.; Mazzanti, C.; Spanevello, R.; Gutierres, J.; Bagatini, M.; Martins, C.C.; Abdalla, F.H.; Daci da Silva, S.J.; et al. Effects of resveratrol on biomarkers of oxidative stress and on the activity of delta aminolevulinic acid dehydratase in liver and kidney of streptozotocin-induced diabetic rats. Biochimie 2012, 94, 374–383. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.M.; Shaw, L.H.; Chang, P.J.; Tung, S.Y.; Chang, T.S.; Shen, C.H.; Hsieh, Y.Y.; Wei, K.L. Hepatoprotective effect of resveratrol against ethanol-induced oxidative stress through induction of superoxide dismutase in vivo and in vitro. Exp. Ther. Med. 2016, 11, 1231–1238. [Google Scholar] [CrossRef] [Green Version]
- Olmos, Y.; Sánchez-Gómez, F.J.; Wild, B.; García-Quintans, N.; Cabezudo, S.; Lamas, S.; Monsalve, M. SirT1 regulation of antioxidant genes is dependent on the formation of a FoxO3a/PGC-1alpha complex. Antioxid. Redox. Signal. 2013, 19, 1507–1521. [Google Scholar] [CrossRef]
- Li, N.; Muthusamy, S.; Liang, R.; Sarojini, H.; Wang, E. Increased expression of miR-34a and miR-93 in rat liver during aging, and their impact on the expression of Mgst1 and Sirt1. Mech. Ageing Dev. 2011, 132, 75–85. [Google Scholar] [CrossRef]
- Kupis, W.; Palyga, J.; Tomal, E.; Niewiadomska, E. The role of sirtuins in cellular homeostasis. J. Physiol. Biochem. 2016, 72, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.H.; Lee, Y.M.; Chun, Y.S.; Chen, J.; Kim, J.E.; Park, J.W. Sirtuin 1 modulates cellular responses to hypoxia by deacetylating hypoxia-inducible factor 1alpha. Mol. Cell 2010, 38, 864–878. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Shin, S.H.; Shin, D.H.; Chun, Y.S.; Park, J.W. Differential roles of Sirt1 in HIF-1alpha and HIF-2alpha mediated hypoxic responses. Biochem. Biophys. Res. Commun. 2014, 444, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Ryu, D.R.; Yu, M.R.; Kong, K.H.; Kim, H.; Kwon, S.H.; Jeon, J.S.; Han, D.C.; Noh, H. Sirt1-hypoxia-inducible factor-1alpha interaction is a key mediator of tubulointerstitial damage in the aged kidney. Aging Cell 2019, 18, e12904. [Google Scholar] [CrossRef] [PubMed]
- Ke, Q.; Costa, M. Hypoxia-inducible factor-1 (HIF-1). Mol. Pharmacol. 2006, 70, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. AMPK and HIF signaling pathways regulate both longevity and cancer growth: The good news and the bad news about survival mechanisms. Biogerontology 2016, 17, 655–680. [Google Scholar] [CrossRef]
- Jung, Y.J.; Isaacs, J.S.; Lee, S.; Trepel, J.; Neckers, L. IL-1beta-mediated up-regulation of HIF-1alpha via an NFkappaB/COX-2 pathway identifies HIF-1 as a critical link between inflammation and oncogenesis. FASEB J. 2003, 17, 2115–2117. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, K.M.; Le, A.; Duong, H.; Wu, Y.; Zhang, Q.; Messadi, D.V. Correlation between VEGF and HIF-1alpha expression in human oral squamous cell carcinoma. Exp. Mol. Pathol. 2004, 76, 143–152. [Google Scholar] [CrossRef]
- Koperek, O.; Akin, E.; Asari, R.; Niederle, B.; Neuhold, N. Expression of hypoxia-inducible factor 1 alpha in papillary thyroid carcinoma is associated with desmoplastic stromal reaction and lymph node metastasis. Virchows Arch. 2013, 463, 795–802. [Google Scholar] [CrossRef]
- Lodewijk, L.; van Diest, P.; van der Groep, P.; Ter Hoeve, N.; Schepers, A.; Morreau, J.; Bonenkamp, J.; van Engen-van Grunsven, A.; Kruijff, S.; van Hemel, B.; et al. Expression of HIF-1alpha in medullary thyroid cancer identifies a subgroup with poor prognosis. Oncotarget 2017, 8, 28650–28659. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Pore, N.; Behrooz, A.; Ismail-Beigi, F.; Maity, A. Regulation of glut1 mRNA by hypoxia-inducible factor-1. Interaction between H-ras and hypoxia. J. Biol. Chem. 2001, 276, 9519–9525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, R.G.; Cazarin, J.M.; de Albuquerque, J.P.C.; de Andrade, B.M.; Carvalho, D.P. Differential glycolytic profile and Warburg effect in papillary thyroid carcinoma cell lines. Oncol. Rep. 2016, 36, 3673–3681. [Google Scholar] [CrossRef]
- Abbasalizad, F.M.; Tajmiri, S. The correlation between inflammatory and metabolic parameters with thyroid function in patients with Hashimoto’s thyroiditis: The potential role of interleukin 23 (IL-23) and vascular endothelial growth factor (VEGF)-1. Acta Endocrinol. 2018, 14, 163–168. [Google Scholar]
- Chen, Y.K.; Lin, C.L.; Cheng, F.T.; Sung, F.C.; Kao, C.H. Cancer risk in patients with Hashimoto’s thyroiditis: A nationwide cohort study. Br. J. Cancer 2013, 109, 2496–2501. [Google Scholar] [CrossRef] [Green Version]
- Guarino, V.; Castellone, M.D.; Avilla, E.; Melillo, R.M. Thyroid cancer and inflammation. Mol. Cell Endocrinol. 2010, 321, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Fugazzola, L.; Colombo, C.; Perrino, M.; Muzza, M. Papillary thyroid carcinoma and inflammation. Front. Endocrinol. 2011, 2, 88. [Google Scholar] [CrossRef] [Green Version]
- Paparodis, R.D.; Karvounis, E.; Bantouna, D.; Chourpiliadis, C.; Chourpiliadi, H.; Livadas, S.; Imam, S.; Jaume, J.C. Incidentally discovered papillary thyroid microcarcinomas are more frequently found in patients with chronic lymphocytic thyroiditis than with multinodular goiter or Graves’ disease. Thyroid 2020, 30, 531–535. [Google Scholar] [CrossRef]
- Joris, V.; Gomez, E.L.; Menchi, L.; Lobysheva, I.; Di Mauro, V.; Esfahani, H.; Condorelli, G.; Balligand, J.L.; Catalucci, D.; Dessy, C. MicroRNA-199a-3p and microRNA-199a-5p take part to a redundant network of regulation of the NOS (NO Synthase)/NO pathway in the endothelium. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 2345–2357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.A.; Tyagi, M.; Sharma, A.K.; Barreto, S.G.; Sirohi, B.; Ramadwar, M.; Shrikhande, S.V.; Gupta, S. Cell-type specificity of beta-actin expression and its clinicopathological correlation in gastric adenocarcinoma. World J. Gastroenterol. 2014, 20, 12202–12211. [Google Scholar] [CrossRef]
- Zhang, B.; Wu, X.; Liu, J.; Song, L.; Song, Q.; Wang, L.; Yuan, D.; Wu, Z. Beta-Actin: Not a suitable internal control of hepatic fibrosis caused by Schistosoma japonicum. Front. Microbiol. 2019, 10, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Software Image J, Version 1.52a, Java 1.8.0_112; Wayne Rasband National Institutes of Health, USA. 2018. Available online: https://imagej.nih.gov/ij/download.html (accessed on 1 April 2021).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Hashimoto’s Patients | n = 6 | |
|---|---|---|
| Mean (range) age | 46.5 (24–58) years | |
| Sex | Female Male | n = 5 n = 1 |
| Mean (range) HT duration | 53.2 (3–156) months | |
| Mean (range) free T4 levels at biopsy time | 13.78 (12.1–15.6) pmol/L; n = 5 | |
| Mean (range) TSH levels at biopsy time | 2.39 (0.01–4.22) µU/mL: n = 6 | |
| Mean (range) anti-TPO levels at biopsy time | 232.675 (67–491.8) U/l; n = 4 | |
| L-thyroxine treatment | n = 6 | |
| Medical imagery | Suspicious nodules; n = 4 |
| Proteins | Primary Antibodies | References | Incubation Conditions | |
|---|---|---|---|---|
| IHC | WB | |||
| Catalase | Mouse Monoclonal Antibody (Sigma, Merck KGaA, Darmstadt, Germany) | C-0979 | 1 µg/mL, 1 h | 2 µg/mL, overnight |
| GAPDH | Rabbit Monoclonal Antibody (Cell signaling, Leiden, The Netherlands) | 2118S | / | 1/3000, overnight |
| GLUT-1 | Rabbit Polyclonal Antibody (Abcam, Cambridge, UK) | Ab32551 | 1/200, overnight | 1/1000, overnight |
| HIF-1α | Rabbit Polyclonal Antibody (Novus Bio, Oxon, UK) | NB 100449 | 4 µg/mL, overnight | 0.4 µg/mL, overnight |
| HNE | Rabbit Polyclonal Antibody (Sigma, Merck KGaA, Darmstadt, Germany) | 393207-100 | 1/200, overnight | / |
| NOX2 | Rabbit Polyclonal Antibody (Abcam, Cambridge, UK) | Ab31092 | 5 µg/mL, overnight | 0.5 µg/mL, overnight |
| NOX4 | Rabbit Polyclonal Antibody (Proteintech, Manchester, UK) | 14347-1-AP | 1/200, overnight | 1/1000, overnight |
| PRDX1 | Rabbit Polyclonal Antibody (Abcam, Cambridge, UK) | Ab15571 | 1/100, overnight | 1/1000, overnight |
| SIRT1 | Rabbit Monoclonal Antibody (Cell signaling, Leiden, The Netherlands) | 9475S | / | 1/1000, overnight |
| SIRT1 | Mouse Monoclonal Antibody (ThermoFischer, Merelbeke, Belgium) | MA5-15677 | 1/150, overnight | / |
| SOD1 | Rabbit Polyclonal Antibody (Abcam, Cambridge, UK) | Ab13498 | 1/500, overnight | 1/1000, overnight |
| T4 | Mouse Monoclonal Antibody (MyBiosource, San Diego, CA, USA) | MBS 5920 059 | 1/400, overnight | / |
| VEGF-A | Rabbit Monoclonal Antibody (Abcam, Cambridge, UK) | Ab46154 | 5 µg/mL, overnight | 0.5 µg/mL, overnight |
| β-actin | Mouse Monoclonal Antibody (Sigma, Merck KGaA, Darmstadt, Germany) | A5441 | / | 1/200,000, overnight |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hepp, M.; Werion, A.; De Greef, A.; de Ville de Goyet, C.; de Bournonville, M.; Behets, C.; Lengelé, B.; Daumerie, C.; Mourad, M.; Ludgate, M.; et al. Oxidative Stress-Induced Sirtuin1 Downregulation Correlates to HIF-1α, GLUT-1, and VEGF-A Upregulation in Th1 Autoimmune Hashimoto’s Thyroiditis. Int. J. Mol. Sci. 2021, 22, 3806. https://doi.org/10.3390/ijms22083806
Hepp M, Werion A, De Greef A, de Ville de Goyet C, de Bournonville M, Behets C, Lengelé B, Daumerie C, Mourad M, Ludgate M, et al. Oxidative Stress-Induced Sirtuin1 Downregulation Correlates to HIF-1α, GLUT-1, and VEGF-A Upregulation in Th1 Autoimmune Hashimoto’s Thyroiditis. International Journal of Molecular Sciences. 2021; 22(8):3806. https://doi.org/10.3390/ijms22083806
Chicago/Turabian StyleHepp, Michaël, Alexis Werion, Axel De Greef, Christine de Ville de Goyet, Marc de Bournonville, Catherine Behets, Benoit Lengelé, Chantal Daumerie, Michel Mourad, Marian Ludgate, and et al. 2021. "Oxidative Stress-Induced Sirtuin1 Downregulation Correlates to HIF-1α, GLUT-1, and VEGF-A Upregulation in Th1 Autoimmune Hashimoto’s Thyroiditis" International Journal of Molecular Sciences 22, no. 8: 3806. https://doi.org/10.3390/ijms22083806