
Novel CRISPR–Cas Systems: An Updated Review of the Current Achievements, Applications, and Future Research Perspectives

 , , , and
, , , and
Abstract
:1. Introduction
2. The CRISPR–Cas System
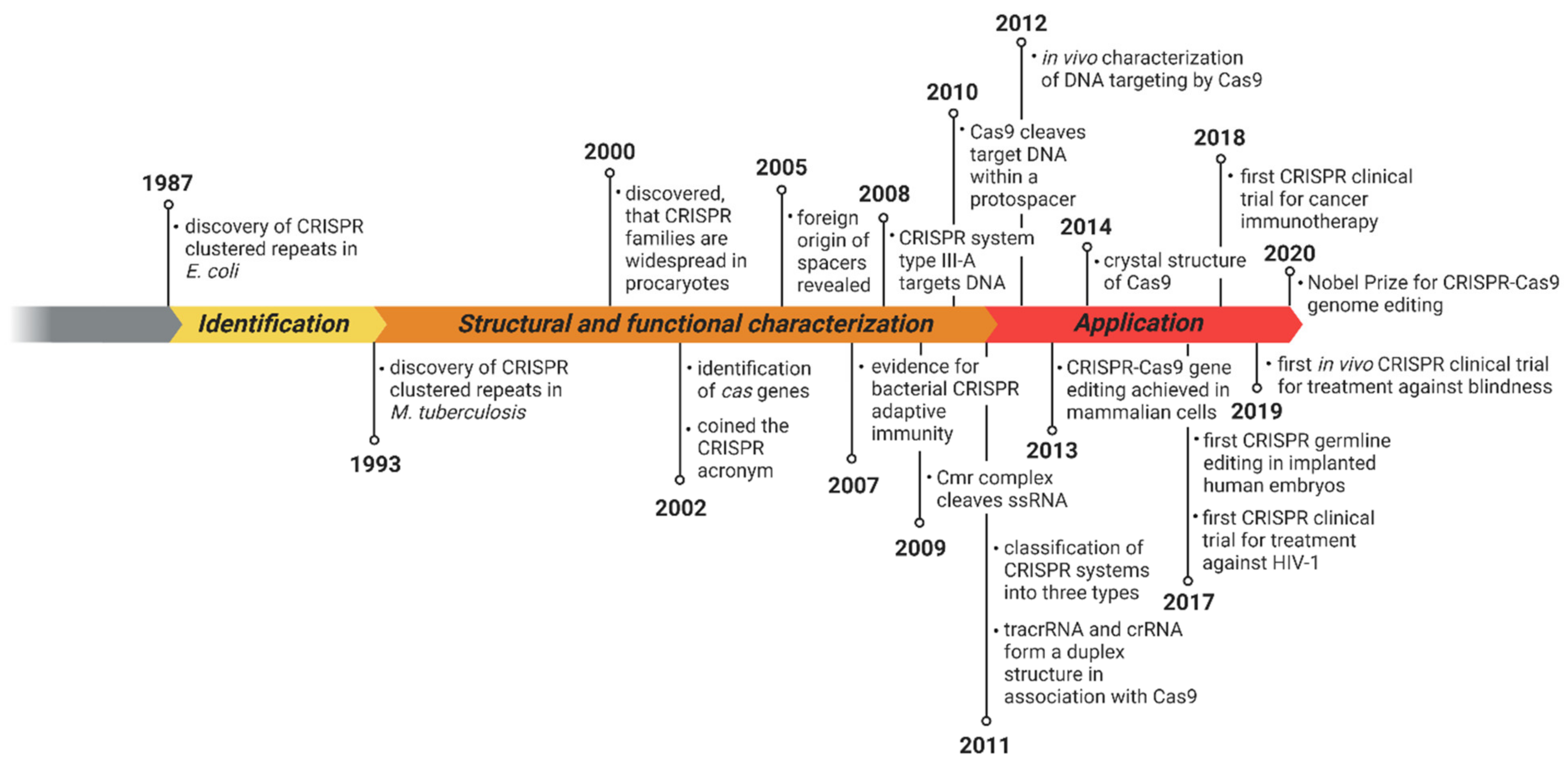
2.1. History of the CRISPR–Cas System
2.2. Function of the CRISPR–Cas System
- (1)
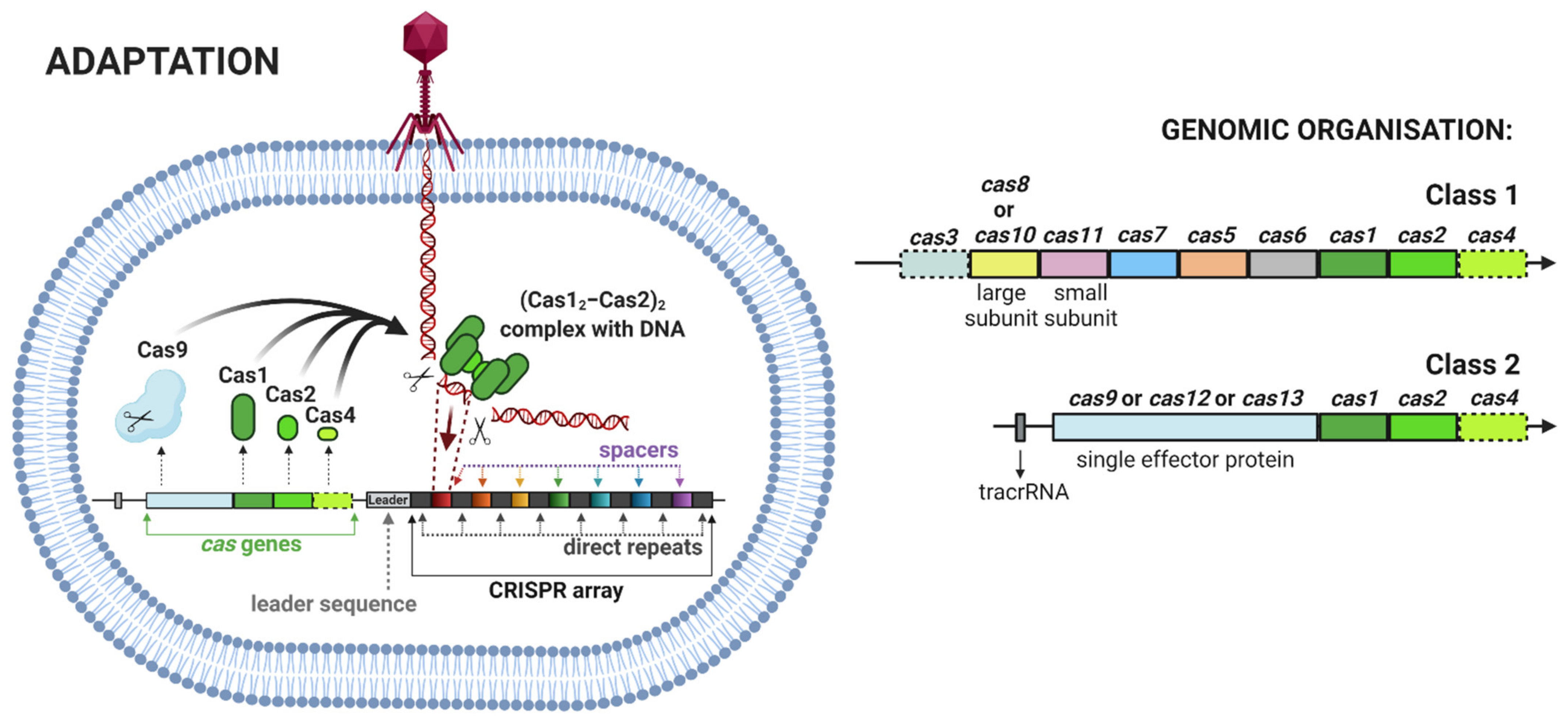
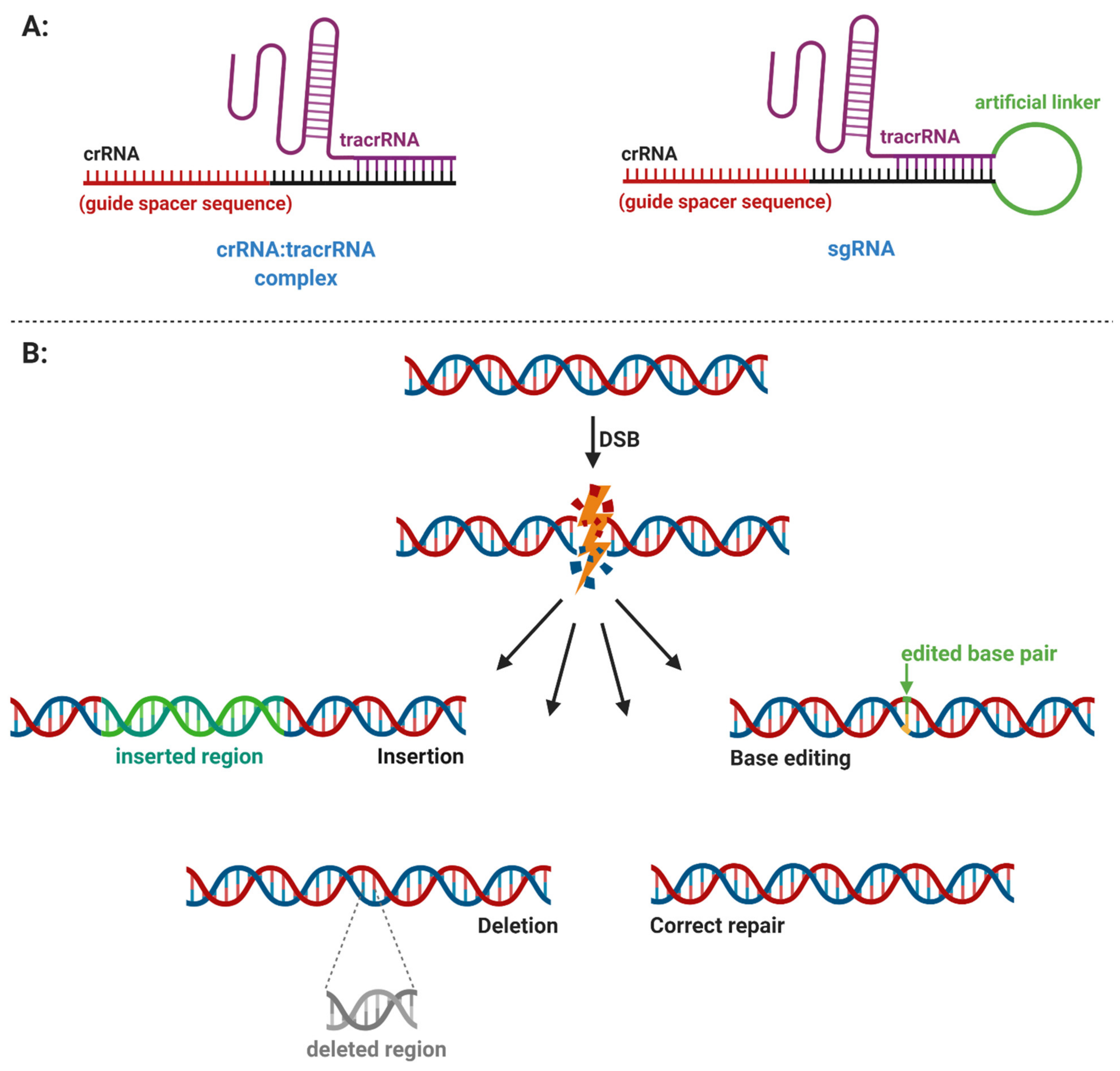
- Adaptation, also termed as insertion or acquisition, is a process of foreign DNA sequence incorporation into CRISPR arrays (Figure 2). Integration of a new spacer is mediated by the heterohexameric protein complex ((Cas12–Cas2)2) to the leader sequence of the CRISPR array. There are two different types of spacer acquisition system named type I and type II. A type I system utilizes integration host factor (IHF) bonded to the leader sequence, which induces DNA bending. This bending enables the (Cas12–Cas2)2 complex to perform an initial cleavage for insertion of the spacer. In a type II system, the leader anchoring sequence (LAS) of the leader is recognized using the Cas1 protein of the (Cas12–Cas2)2 complex and, then, the polar spacer is inserted [13].
- (2)
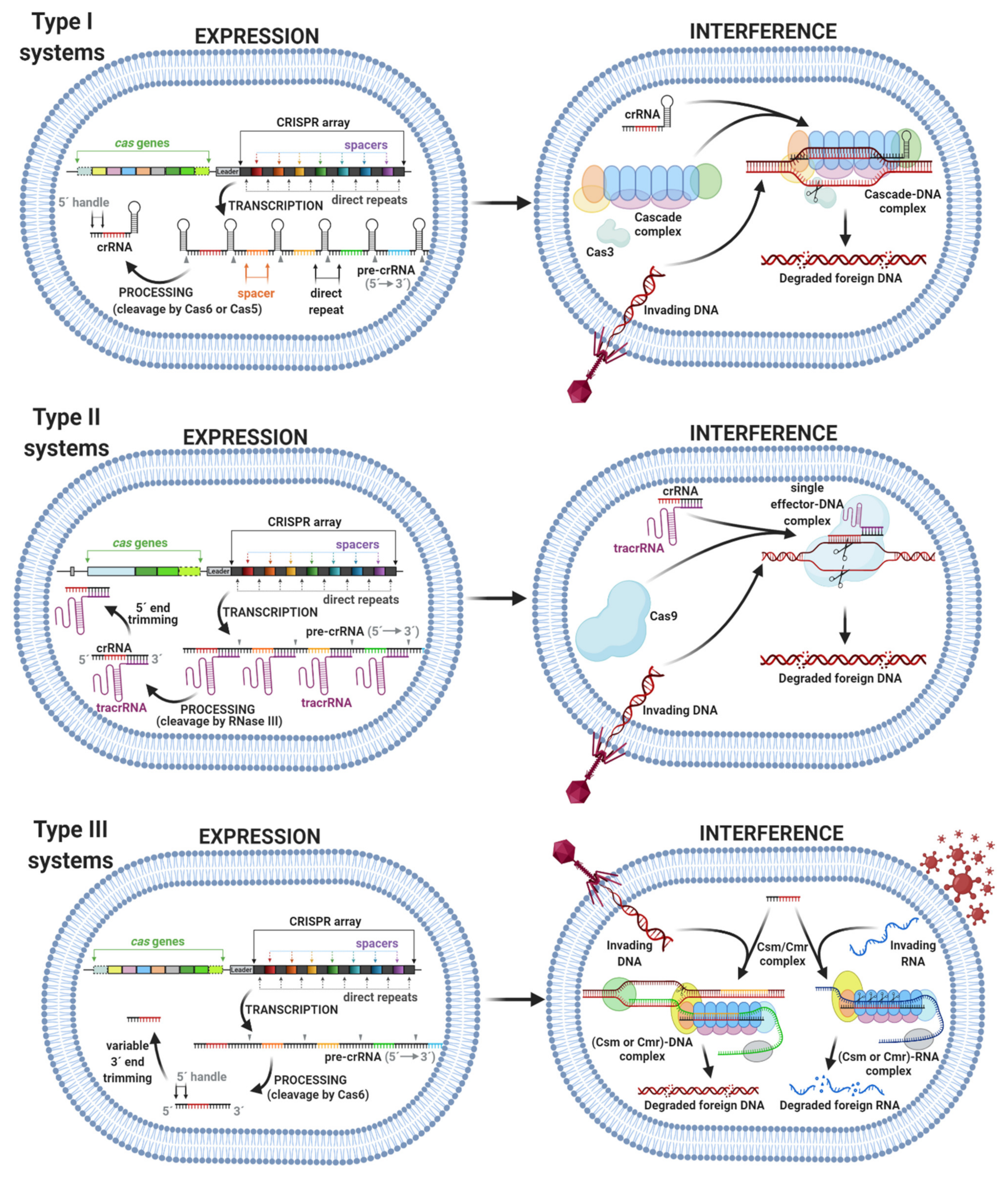
- Expression, also termed CRISPR RNA (crRNA) biogenesis or crRNA processing, represents the transcription of a CRISPR array into a long precursor CRISPR RNA (pre-crRNA). Further processing involves the cleavage of pre-crRNA within each direct repeat sequence to afford shorter, mature CRISPR RNAs (crRNAs). Some crRNAs can further undergo 5′ end or 3′ end trimming. The enzymes involved in crRNA processing differ among types, in some cases, even among subtypes of the CRISPR–Cas system (Figure 3) [15].
- (3)
- Interference involves the formation of a multiprotein effector complex or single effector protein. The multiprotein effector complex is typical for class 1 systems and consists of multiple Cas proteins and crRNA. The single effector protein is utilized in class 2 systems and contains only a single Cas protein with crRNA. The primary purpose of both complexes is to recognize the same or very similar sequences in the genome of the invading virus or plasmid. After recognition, the invading genome is cleaved by the complex and inactivated [14]. The interference step of some systems requires recognition of a protospacer adjacent motif (PAM) in the invading genome. The PAM is a short DNA sequence that is not present in the bacterial host genome. Hence, PAM is an essential targeting sequence to bind for some Cas proteins, followed by cleavage (Figure 3) [16].
2.3. Structure of CRISPR Locus and Classification of Cas Proteins
- The CRISPR array consists of short, direct repeats bordered with spacers. The direct repeats are nucleotide sequences in the genome with identical sequence and length. The sequences of direct repeats can be similar in related species, but overall diversity among the species is wide. The average size of the repeats is 32 bp; however, the size may vary from 21 to 47 bp. The spacers are nucleotide sequences with a fixed length, but they are highly variable in sequences. The average size of spacers is from 20 to 72 bp [18].
- The leader sequence is commonly adjacent to the CRISPR array and is involved in adaptation and transcription. These regions exhibit only limited conservation in sequence. It was observed that leaderless CRISPR loci are inactive in adaptation but still able to contribute to crRNA-directed interference [19].
- CRISPR-associated (cas) genes represent a cluster of genes in varying orientation and order that code corresponding Cas proteins (Table 1). In summary, 93 different cas genes have been identified until now. These genes were classified into 35 families based on the sequence similarities [20]. Cas proteins play a major role in the acquisition and destruction of foreign sequences (Table 2).
2.4. Classification of the CRISPR–Cas System
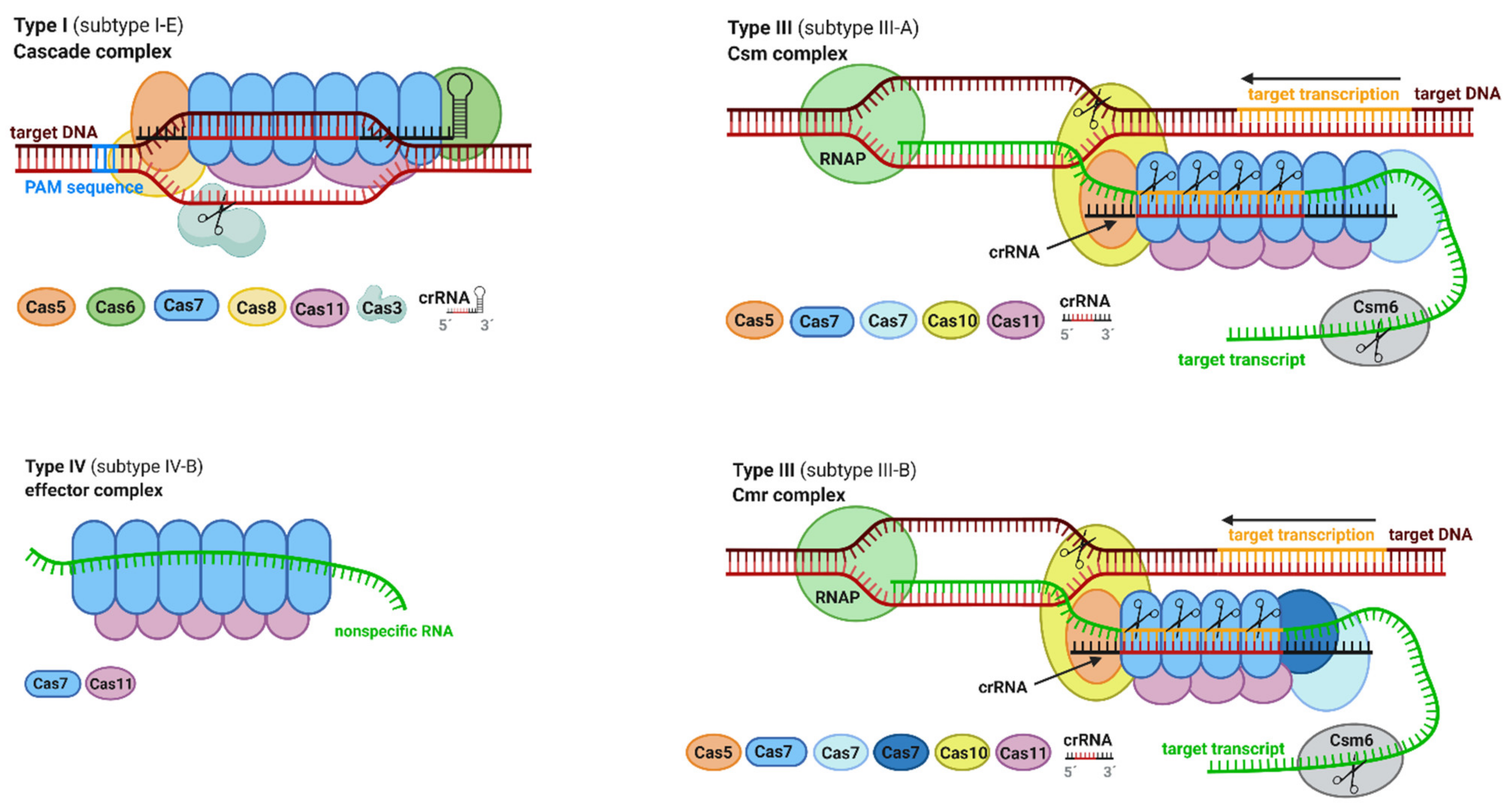
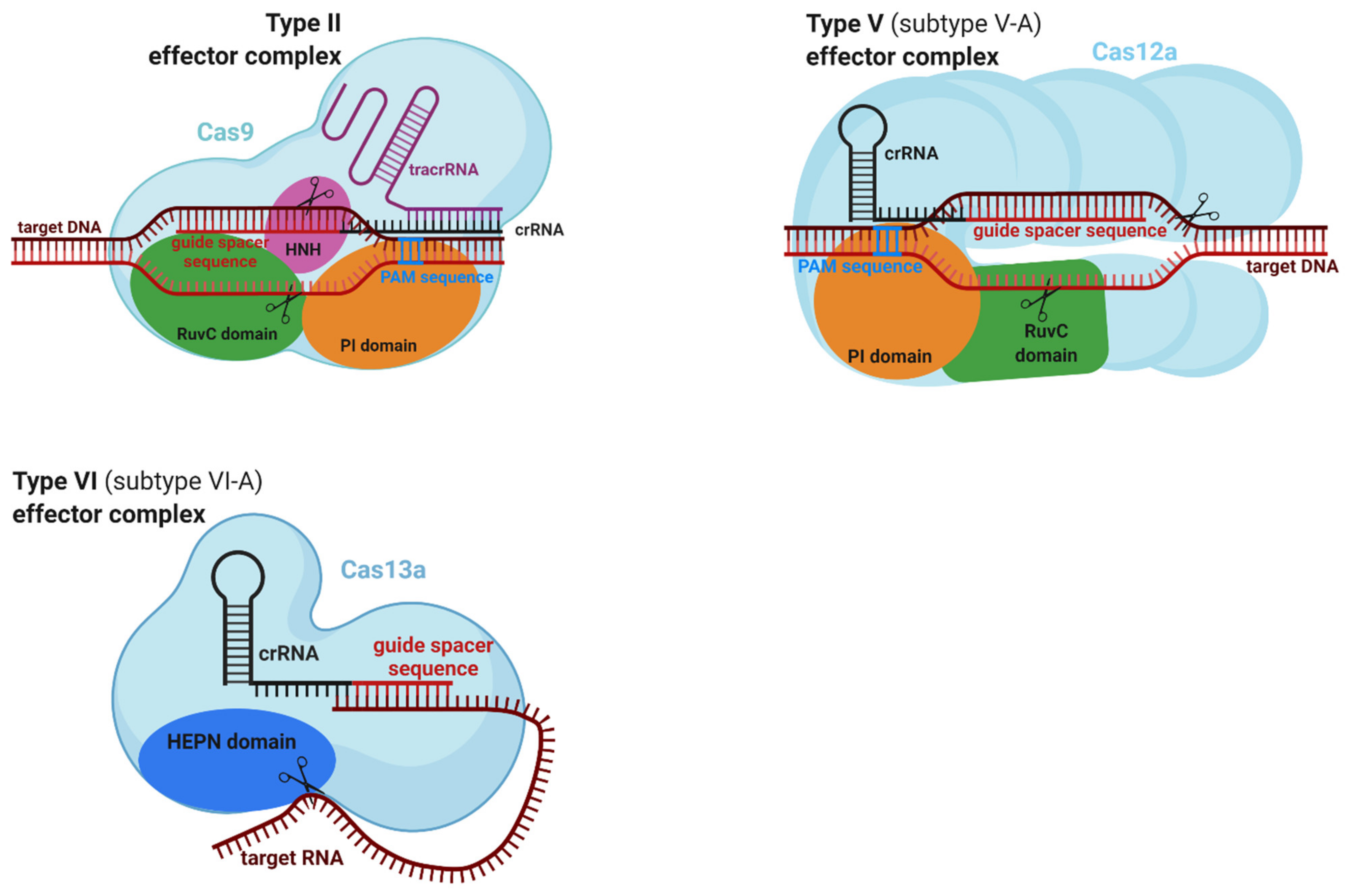
2.5. Structure and Function of Effector Complexes
2.5.1. Effector Complexes of Class 1
2.5.2. Effector Complexes of Class 2
3. Applications of CRISPR–Cas Systems
3.1. In Plant Biotechnology
3.1.1. Resistivity to Stress
3.1.2. Prospects in Plant Gene Editing
3.2. In Therapeutics
3.2.1. For Treating Genetic Diseases
3.2.2. For Management of Infectious Diseases
Control of Human Immunodeficiency Virus (HIV)
Management of Other Infectious Diseases
3.2.3. For Management of Cancers
3.2.4. Prospects in Therapeutics and Management of Infectious Diseases
3.3. In the Food Industry
Prospects in the Food Industry
4. CRISPR–Cas9: Ifs and Buts
5. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakata, A. Nucleotide sequence of the Iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product. J. Bacteriol. 1987, 169, 5429–5433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jore, M.M.; Brouns, S.J.J.; van der Oost, J. RNA in defense: CRISPRs protect prokaryotes against mobile genetic elements. Cold Spring Harb. Perspect. Biol. 2012, 4. [Google Scholar] [CrossRef]
- Jansen, R.; van Embden, J.D.A.; Gaastra, W.; Schouls, L.M. Identification of genes that are associated with DNA repeats in prokaryotes. Mol. Microbiol. 2002, 43, 1565–1575. [Google Scholar] [CrossRef]
- Mojica, F.J.; Juez, G.; Rodríguez-Valera, F. Transcription at different salinities of Haloferax mediterranei sequences adjacent to partially modified PstI sites. Mol. Microbiol. 1993, 9, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Aravind, L.; Grishin, N.V.; Rogozin, I.B.; Koonin, E.V. A DNA repair system specific for thermophilic Archaea and bacteria predicted by genomic context analysis. Nucleic Acids Res. 2002, 30, 482–496. [Google Scholar] [CrossRef] [Green Version]
- Mojica, F.J.M.; Díez-Villaseñor, C.; García-Martínez, J.; Soria, E. Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements. J. Mol. Evol. 2005, 60, 174–182. [Google Scholar] [CrossRef]
- Pourcel, C.; Salvignol, G.; Vergnaud, G. CRISPR Elements in Yersinia pestis acquire new repeats by preferential uptake of bacteriophage DNA, and provide additional tools for evolutionary studies. Microbiology 2005, 151, 653–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makarova, K.S.; Grishin, N.V.; Shabalina, S.A.; Wolf, Y.I.; Koonin, E.V. A Putative RNA-interference-based immune system in prokaryotes: Computational analysis of the predicted enzymatic machinery, functional analogies with eukaryotic RNAi, and hypothetical mechanisms of action. Biol. Direct. 2006, 1, 7. [Google Scholar] [CrossRef] [Green Version]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR provides acquired resistance against viruses in prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef]
- Makarova, K.S.; Haft, D.H.; Barrangou, R.; Brouns, S.J.J.; Charpentier, E.; Horvath, P.; Moineau, S.; Mojica, F.J.M.; Wolf, Y.I.; Yakunin, A.F.; et al. Evolution and classification of the CRISPR-cas systems. Nat. Rev. Microbiol. 2011, 9, 467–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sapranauskas, R.; Gasiunas, G.; Fremaux, C.; Barrangou, R.; Horvath, P.; Siksnys, V. The Streptococcus thermophilus CRISPR/cas system provides immunity in Escherichia Coli. Nucleic Acids Res. 2011, 39, 9275–9282. [Google Scholar] [CrossRef]
- Bikard, D.; Jiang, W.; Samai, P.; Hochschild, A.; Zhang, F.; Marraffini, L.A. Programmable repression and activation of bacterial gene expression using an engineered CRISPR-cas system. Nucleic Acids Res. 2013, 41, 7429–7437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGinn, J.; Marraffini, L.A. Molecular mechanisms of CRISPR-cas spacer acquisition. Nat. Rev. Microbiol. 2019, 17, 7–12. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; Iranzo, J.; Shmakov, S.A.; Alkhnbashi, O.S.; Brouns, S.J.J.; Charpentier, E.; Cheng, D.; Haft, D.H.; Horvath, P.; et al. Evolutionary classification of CRISPR–cas systems: A burst of class 2 and derived variants. Nat. Rev. Microbiol. 2020, 18, 67–83. [Google Scholar] [CrossRef]
- Hochstrasser, M.L.; Doudna, J.A. Cutting it close: CRISPR-associated endoribonuclease structure and function. Trends Biochem. Sci. 2015, 40, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Gleditzsch, D.; Pausch, P.; Müller-Esparza, H.; Özcan, A.; Guo, X.; Bange, G.; Randau, L. PAM identification by CRISPR-cas effector complexes: Diversified mechanisms and structures. RNA Biol. 2018, 16, 504–517. [Google Scholar] [CrossRef] [PubMed]
- Mojica, F.J.; Díez-Villaseñor, C.; Soria, E.; Juez, G. Biological significance of a family of regularly spaced repeats in the genomes of Archaea, Bacteria and Mitochondria. Mol. Microbiol. 2000, 36, 244–246. [Google Scholar] [CrossRef]
- Karginov, F.V.; Hannon, G.J. The CRISPR system: Small RNA-guided defense in Bacteria and Archaea. Mol. Cell 2010, 37, 7. [Google Scholar] [CrossRef] [Green Version]
- Alkhnbashi, O.S.; Shah, S.A.; Garrett, R.A.; Saunders, S.J.; Costa, F.; Backofen, R. Characterizing leader sequences of CRISPR loci. Bioinformatics 2016, 32, i576–i585. [Google Scholar] [CrossRef] [Green Version]
- Makarova, K.S.; Wolf, Y.I.; Alkhnbashi, O.S.; Costa, F.; Shah, S.A.; Saunders, S.J.; Barrangou, R.; Brouns, S.J.J.; Charpentier, E.; Haft, D.H.; et al. An updated evolutionary classification of CRISPR–cas systems. Nat. Rev. Microb. 2015, 13, 722–736. [Google Scholar] [CrossRef] [Green Version]
- Clark, D.P.; Pazdernik, N.J.; McGehee, M.R. Chapter 20-Genome Defense. In Molecular Biology, 3rd ed.; Clark, D.P., Pazdernik, N.J., McGehee, M.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 622–653. ISBN 978-0-12-813288-3. [Google Scholar]
- Charpentier, E.; Richter, H.; van der Oost, J.; White, M.F. Biogenesis pathways of RNA guides in Archaeal and Bacterial CRISPR-cas adaptive immunity. FEMS Microbiol. Rev. 2015, 39, 428–441. [Google Scholar] [CrossRef] [PubMed]
- Özcan, A.; Pausch, P.; Linden, A.; Wulf, A.; Schühle, K.; Heider, J.; Urlaub, H.; Heimerl, T.; Bange, G.; Randau, L. Type IV CRISPR RNA processing and effector complex formation in Aromatoleum aromaticum. Nat. Microbiol. 2019, 4, 89–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hille, F.; Richter, H.; Wong, S.P.; Bratovič, M.; Ressel, S.; Charpentier, E. The biology of CRISPR-cas: Backward and forward. Cell 2018, 172, 1239–1259. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Bravo, J.P.K.; Taylor, H.N.; Steens, J.; Jackson, R.N.; Staals, R.H.J.; Taylor, D.W. Structure of a type IV CRISPR-cas effector complex. bioRxiv 2020. [Google Scholar] [CrossRef]
- Schindele, P.; Wolter, F.; Puchta, H. Transforming plant biology and breeding with CRISPR/Cas9, Cas12 and Cas13. FEBS Lett. 2018, 592, 1954–1967. [Google Scholar] [CrossRef] [Green Version]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Li, Y.; Glass, Z.; Huang, M.; Chen, Z.-Y.; Xu, Q. Ex vivo cell-based CRISPR/Cas9 genome editing for therapeutic applications. Biomaterials 2020, 234, 119711. [Google Scholar] [CrossRef]
- Rees, H.A.; Liu, D.R. Base editing: Precision chemistry on the genome and transcriptome of living cells. Nat. Rev. Genet. 2018, 19, 770–788. [Google Scholar] [CrossRef]
- Miao, J.; Guo, D.; Zhang, J.; Huang, Q.; Qin, G.; Zhang, X.; Wan, J.; Gu, H.; Qu, L.-J. Targeted mutagenesis in rice using CRISPR-cas system. Cell Res. 2013, 23, 1233–1236. [Google Scholar] [CrossRef] [Green Version]
- Shan, Q.; Wang, Y.; Li, J.; Gao, C. Genome editing in rice and wheat using the CRISPR/cas system. Nat. Protoc. 2014, 9, 2395–2410. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, D.; Zhang, D.; Zhao, X.; Cao, X.; Dong, L.; Liu, J.; Chen, K.; Zhang, H.; Gao, C.; et al. Analysis of the functions of TaGW2 homoeologs in wheat grain weight and protein content traits. Plant J. 2018, 94, 857–866. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Zhang, Q.; Zhu, Q.; Liu, W.; Chen, Y.; Qiu, R.; Wang, B.; Yang, Z.; Li, H.; Lin, Y.; et al. A Robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicot plants. Mol. Plant 2015, 8, 1274–1284. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.Z.; Henry, I.M.; Lynagh, P.G.; Comai, L.; Cahoon, E.B.; Weeks, D.P. Significant enhancement of fatty acid composition in seeds of the allohexaploid, cCamelina sativa, using CRISPR/Cas9 gene editing. Plant Biotechnol. J. 2017, 15, 648–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-León, S.; Gil-Humanes, J.; Ozuna, C.V.; Giménez, M.J.; Sousa, C.; Voytas, D.F.; Barro, F. Low-gluten, nontransgenic wheat engineered with CRISPR/Cas9. Plant Biotechnol. J. 2018, 16, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Aznar-Moreno, J.A.; Durrett, T.P. Simultaneous targeting of multiple gene homeologs to alter seed oil production in Camelina sativa. Plant Cell Physiol. 2017, 58, 1260–1267. [Google Scholar] [CrossRef]
- Sun, Y.; Jiao, G.; Liu, Z.; Zhang, X.; Li, J.; Guo, X.; Du, W.; Du, J.; Francis, F.; Zhao, Y.; et al. Generation of high-amylose rice through CRISPR/Cas9-mediated targeted mutagenesis of starch branching enzymes. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Shan, Q.; Wang, Y.; Li, J.; Zhang, Y.; Chen, K.; Liang, Z.; Zhang, K.; Liu, J.; Xi, J.J.; Qiu, J.-L.; et al. Targeted genome modification of crop plants using a CRISPR-cas system. Nat. Biotechnol. 2013, 31, 686–688. [Google Scholar] [CrossRef]
- Klap, C.; Yeshayahou, E.; Bolger, A.M.; Arazi, T.; Gupta, S.K.; Shabtai, S.; Usadel, B.; Salts, Y.; Barg, R. Tomato facultative parthenocarpy results from SlAGAMOUS-LIKE 6 loss of function. Plant Biotechnol. J. 2017, 15, 634–647. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.-P.; Luo, T.; Fu, H.-W.; Wang, L.; Tan, Y.-Y.; Huang, J.-Z.; Wang, Q.; Ye, G.-Y.; Gatehouse, A.M.R.; Lou, Y.-G.; et al. Resistance of Rice to Insect Pests Mediated by Suppression of Serotonin Biosynthesis. Nat. Plants 2018, 4, 338–344. [Google Scholar] [CrossRef]
- Christian, M.; Cermak, T.; Doyle, E.L.; Schmidt, C.; Zhang, F.; Hummel, A.; Bogdanove, A.J.; Voytas, D.F. Targeting DNA Double-Strand Breaks with TAL Effector Nucleases. Genetics 2010, 186, 757–761. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Wang, Y.; Zhang, R.; Zhang, H.; Gao, C. CRISPR/Cas genome editing and precision plant breeding in agriculture. Annu. Rev. Plant Biol. 2019, 70, 667–697. [Google Scholar] [CrossRef]
- Kim, H.; Kim, S.-T.; Ryu, J.; Kang, B.-C.; Kim, J.-S.; Kim, S.-G. CRISPR/Cpf1-mediated DNA-free plant genome editing. Nat. Commun. 2017, 8, 14406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, R.; Li, H.; Qin, R.; Wang, L.; Li, L.; Wei, P.; Yang, J. Gene targeting using the Agrobacterium Ttumefaciens-mediated CRISPR-cas system in rice. Rice 2014, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Yin, K.; Han, T.; Liu, G.; Chen, T.; Wang, Y.; Yu, A.Y.L.; Liu, Y. A Geminivirus-based guide RNA delivery system for CRISPR/Cas9 mediated plant genome editing. Sci. Rep. 2015, 5, 14926. [Google Scholar] [CrossRef] [PubMed]
- Baltes, N.J.; Hummel, A.W.; Konecna, E.; Cegan, R.; Bruns, A.N.; Bisaro, D.M.; Voytas, D.F. Conferring resistance to geminiviruses with the CRISPR-cas prokaryotic immune System. Nat. Plants 2015, 1, 15145. [Google Scholar] [CrossRef]
- Feng, Z.; Mao, Y.; Xu, N.; Zhang, B.; Wei, P.; Yang, D.-L.; Wang, Z.; Zhang, Z.; Zheng, R.; Yang, L.; et al. Multigeneration analysis reveals the inheritance, specificity, and patterns of CRISPR/cas-induced gene modifications in arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 4632–4637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puchta, H. Using CRISPR/cas in three dimensions: Towards synthetic plant genomes, transcriptomes and epigenomes. Plant J. 2016, 87, 5–15. [Google Scholar] [CrossRef] [Green Version]
- Aman, R.; Ali, Z.; Butt, H.; Mahas, A.; Aljedaani, F.; Khan, M.Z.; Ding, S.; Mahfouz, M. RNA Virus interference via CRISPR/Cas13a system in plants. Genome Biol. 2018, 19, 1. [Google Scholar] [CrossRef]
- Abudayyeh, O.O.; Gootenberg, J.S.; Essletzbichler, P.; Han, S.; Joung, J.; Belanto, J.J.; Verdine, V.; Cox, D.B.T.; Kellner, M.J.; Regev, A.; et al. RNA targeting with CRISPR-Cas13. Nature 2017, 550, 280–284. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; He, M.; Li, J.; Chen, L.; Huang, Z.; Zheng, S.; Zhu, L.; Ni, E.; Jiang, D.; Zhao, B.; et al. Development of commercial thermo-sensitive genic male sterile rice accelerates hybrid rice breeding using the CRISPR/Cas9-Mediated TMS5 editing system. Sci. Rep. 2016, 6, 37395. [Google Scholar] [CrossRef] [Green Version]
- Svitashev, S.; Schwartz, C.; Lenderts, B.; Young, J.K.; Mark Cigan, A. Genome editing in maize directed by CRISPR–Cas9 ribonucleoprotein complexes. Nat. Commun. 2016, 7, 13274. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, M.; Albertsen, M.C.; Young, J.K.; Cigan, A.M. Concurrent modifications in the three homeologs of Ms45 gene with CRISPR-Cas9 lead to rapid generation of male sterile bread wheat (Triticum Aaestivum, L.). Plant. Mol. Biol. 2018, 97, 371–383. [Google Scholar] [CrossRef]
- Yao, L.; Zhang, Y.; Liu, C.; Liu, Y.; Wang, Y.; Liang, D.; Liu, J.; Sahoo, G.; Kelliher, T. OsMATL mutation induces haploid seed formation in indica rice. Nat. Plants 2018, 4, 530–533. [Google Scholar] [CrossRef] [PubMed]
- Shimatani, Z.; Kashojiya, S.; Takayama, M.; Terada, R.; Arazoe, T.; Ishii, H.; Teramura, H.; Yamamoto, T.; Komatsu, H.; Miura, K.; et al. Targeted base editing in rice and tomato using a CRISPR-Cas9 cytidine deaminase fusion. Nat. Biotechnol. 2017, 35, 441–443. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, Z.; Ni, H.; Xu, Y.; Chen, Q.; Jiang, L. CRISPR/Cas9-mediated base-editing system efficiently generates gain-of-function mutations in arabidopsis. Sci. China Life Sci. 2017, 60, 520–523. [Google Scholar] [CrossRef]
- Tian, S.; Jiang, L.; Cui, X.; Zhang, J.; Guo, S.; Li, M.; Zhang, H.; Ren, Y.; Gong, G.; Zong, M.; et al. Engineering herbicide-resistant watermelon variety through CRISPR/Cas9-mediated base-editing. Plant Cell Rep. 2018, 37, 1353–1356. [Google Scholar] [CrossRef] [PubMed]
- Malnoy, M.; Viola, R.; Jung, M.-H.; Koo, O.-J.; Kim, S.; Kim, J.-S.; Velasco, R.; Nagamangala Kanchiswamy, C. DNA-free genetically edited grapevine and apple protoplast using CRISPR/Cas9 ribonucleoproteins. Front. Plant Sci. 2016, 7, 1904. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Jordan, W.T.; Shi, X.; Hu, L.; He, C.; Schmitz, R.J. TET-mediated epimutagenesis of the Arabidopsis thaliana methylome. Nat. Commun. 2018, 9, 895. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, L.; Ntui, V.O.; Tripathi, J.N. CRISPR/Cas9-based genome editing of banana for disease resistance. Curr. Opin. Plant Biol. 2020, 56, 118–126. [Google Scholar] [CrossRef]
- Gomez, M.A.; Lin, Z.D.; Moll, T.; Luebbert, C.; Chauhan, R.D.; Vijayaraghavan, A.; Kelley, R.; Beyene, G.; Taylor, N.J.; Carrington, J.; et al. Simultaneous CRISPR/Cas9-mediated editing of cassava EIF4E isoforms NCBP-1 and NCBP-2 confers elevated resistance to cassava brown streak disease. bioRxiv 2017, 209874. [Google Scholar] [CrossRef] [Green Version]
- Hummel, A.W.; Chauhan, R.D.; Cermak, T.; Mutka, A.M.; Vijayaraghavan, A.; Boyher, A.; Starker, C.G.; Bart, R.; Voytas, D.F.; Taylor, N.J. Allele exchange at the EPSPS locus confers glyphosate tolerance in cassava. Plant Biotechnol. J. 2018, 16, 1275–1282. [Google Scholar] [CrossRef] [Green Version]
- Peng, A.; Chen, S.; Lei, T.; Xu, L.; He, Y.; Wu, L.; Yao, L.; Zou, X. Engineering canker-resistant plants through CRISPR/Cas9-targeted editing of the susceptibility gene CsLOB1 promoter in citrus. Plant Biotechnol. J. 2017, 15, 1509–1519. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Zhang, Y.; Orbović, V.; Xu, J.; White, F.F.; Jones, J.B.; Wang, N. Genome editing of the disease susceptibility gene CsLOB1 in citrus confers resistance to citrus canker. Plant Biotechnol. J. 2017, 15, 817–823. [Google Scholar] [CrossRef] [Green Version]
- Fister, A.S.; Landherr, L.; Maximova, S.N.; Guiltinan, M.J. Transient expression of CRISPR/Cas9 machinery targeting TcNPR3 enhances defense response in theobroma cacao. Front. Plant Sci. 2018, 9, 268. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, Z.; Sattar, M.N.; Shafiq, M. CRISPR/Cas9: A tool to circumscribe cotton leaf curl disease. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Ge, X.; Luo, X.; Wang, P.; Fan, Q.; Hu, G.; Xiao, J.; Li, F.; Wu, J. Simultaneous editing of two copies of Gh14-3-3d confers enhanced transgene-clean plant defense against Verticillium Dahliae in allotetraploid upland cotton. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, J.; Brumin, M.; Wolf, D.; Leibman, D.; Klap, C.; Pearlsman, M.; Sherman, A.; Arazi, T.; Gal-On, A. Development of broad virus resistance in non-transgenic cucumber using CRISPR/Cas9 technology. Mol. Plant Pathol. 2016, 17, 1140–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauer, N.J.; Narváez-Vásquez, J.; Mozoruk, J.; Miller, R.B.; Warburg, Z.J.; Woodward, M.J.; Mihiret, Y.A.; Lincoln, T.A.; Segami, R.E.; Sanders, S.L.; et al. Oligonucleotide-mediated genome editing provides precision and function to engineered nucleases and antibiotics in plants. Plant Physiol. 2016, 170, 1917–1928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Guo, R.; Tu, M.; Wang, D.; Guo, C.; Wan, R.; Li, Z.; Wang, X. Ectopic expression of the wild grape WRKY transcription factor VqWRKY52 in Arabidopsis thaliana enhances resistance to the biotrophic pathogen powdery mildew but not to the necrotrophic pathogen Botrytis cinerea. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, M.D.; Das, S.; Tarafdar, S. Effect of loading history on visco-elastic potato starch Gel. Coll. Surf. A Physicochem. Eng. Asp. 2016, 492, 47–53. [Google Scholar] [CrossRef]
- Makhotenko, A.V.; Khromov, A.V.; Snigir, E.A.; Makarova, S.S.; Makarov, V.V.; Suprunova, T.P.; Kalinina, N.O.; Taliansky, M.E. Functional analysis of coilin in virus resistance and stress tolerance of potato Solanum tuberosum using CRISPR-Cas9 editing. Dokl. Biochem. Biophys. 2019, 484, 88–91. [Google Scholar] [CrossRef]
- Ma, J.; Chen, J.; Wang, M.; Ren, Y.; Wang, S.; Lei, C.; Cheng, Z. Sodmergen, null disruption of OsSEC3A increases the content of salicylic acid and induces plant defense responses in rice. J. Exp. Bot. 2018, 69, 1051–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macovei, A.; Sevilla, N.R.; Cantos, C.; Jonson, G.B.; Slamet-Loedin, I.; Čermák, T.; Voytas, D.F.; Choi, I.-R.; Chadha-Mohanty, P. Novel alleles of rice EIF4G generated by CRISPR/Cas9-targeted mutagenesis confer resistance to rice tungro spherical virus. Plant Biotechnol. J. 2018, 16, 1918–1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Meng, X.; Zong, Y.; Chen, K.; Zhang, H.; Liu, J.; Li, J.; Gao, C. Gene replacements and insertions in rice by intron targeting using CRISPR-Cas9. Nat. Plants 2016, 2, 16139. [Google Scholar] [CrossRef]
- Mishra, R.; Joshi, R.K.; Zhao, K. Genome editing in rice: Recent advances, challenges, and future implications. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Cai, Y.; Chen, L.; Liu, X.; Sun, S.; Wu, C.; Jiang, B.; Han, T.; Hou, W. CRISPR/Cas9-mediated genome editing in soybean hairy roots. PLoS ONE 2015, 10, e0136064. [Google Scholar] [CrossRef]
- Ludman, M.; Burgyán, J.; Fátyol, K. Crispr/Cas9 mediated inactivation of argonaute 2 reveals its differential involvement in antiviral responses. Sci. Rep. 2017, 7, 1010. [Google Scholar] [CrossRef] [PubMed]
- Nekrasov, V.; Wang, C.; Win, J.; Lanz, C.; Weigel, D.; Kamoun, S. Rapid generation of a transgene-free powdery mildew resistant tomato by genome deletion. Sci. Rep. 2017, 7, 482. [Google Scholar] [CrossRef] [Green Version]
- Ortigosa, A.; Gimenez-Ibanez, S.; Leonhardt, N.; Solano, R. Design of a bacterial speck resistant tomato by CRISPR/Cas9-mediated editing of SlJAZ2. Plant Biotechnol. J. 2019, 17, 665–673. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Bai, Y.; Wu, G.; Zou, S.; Chen, Y.; Gao, C.; Tang, D. Simultaneous modification of three homoeologs of TaEDR1 by genome editing enhances powdery mildew resistance in wheat. Plant J. 2017, 91, 714–724. [Google Scholar] [CrossRef] [Green Version]
- Acevedo-Garcia, J.; Spencer, D.; Thieron, H.; Reinstädler, A.; Hammond-Kosack, K.; Phillips, A.L.; Panstruga, R. Mlo-based powdery mildew resistance in hexaploid bread wheat generated by a non-transgenic TILLING approach. Plant Biotechnol. J. 2017, 15, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Nguyen, K.H.; Chu, H.D.; Ha, C.V.; Watanabe, Y.; Osakabe, Y.; Leyva-González, M.A.; Sato, M.; Toyooka, K.; Voges, L.; et al. The karrikin receptor KAI2 promotes drought resistance in Arabidopsis thaliana. PLoS Genet. 2017, 13, e1007076. [Google Scholar] [CrossRef] [Green Version]
- Kapusi, E.; Corcuera-Gómez, M.; Melnik, S.; Stoger, E. Heritable Genomic fragment deletions and small indels in the putative ENGase gene induced by CRISPR/Cas9 in barley. Front. Plant Sci. 2017, 8, 540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, C.; Liu, X.; Zhang, Z.; Wang, Y.; Duan, W.; Li, S.; Liang, Z. CRISPR/Cas9-mediated efficient targeted mutagenesis in chardonnay (Vitis Vinifera, L.). Sci. Rep. 2016, 6, 32289. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, L.; Tan, Q.; Fan, Q.; Zhu, H.; Hong, Z.; Zhang, Z.; Duanmu, D. Efficient inactivation of symbiotic nitrogen fixation related genes in Lotus japonicus using CRISPR-Cas9. Front. Plant Sci. 2016, 7, 1333. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Y.; Feng, F.; Qi, W.; Song, R. Dek42 encodes an RNA-binding protein that affects alternative Pre-MRNA splicing and maize kernel development. J. Integr. Plant Biol. 2019, 61, 728–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waltz, E. CRISPR-edited crops free to enter market, skip regulation. Nat. Biotechnol. 2016, 34, 582. [Google Scholar] [CrossRef]
- Alagoz, Y.; Gurkok, T.; Zhang, B.; Unver, T. Manipulating the biosynthesis of bioactive compound alkaloids for next-generation metabolic engineering in opium poppy using CRISPR-Cas 9 genome editing technology. Sci. Rep. 2016, 6, 30910. [Google Scholar] [CrossRef] [Green Version]
- Kui, L.; Chen, H.; Zhang, W.; He, S.; Xiong, Z.; Zhang, Y.; Yan, L.; Zhong, C.; He, F.; Chen, J.; et al. Building a genetic manipulation tool box for orchid biology: Identification of constitutive promoters and application of CRISPR/Cas9 in the orchid, Dendrobium officinale. Front. Plant Sci. 2017, 7. [Google Scholar] [CrossRef]
- Semiarti, E.; Nopitasari, S.; Setiawati, Y.; Lawrie, M.D.; Purwantoro, A.; Widada, J.; Yoshioka, Y.; Matsumoto, S.; Ninomiya, K.; Asano, Y. Application of CRISPR/Cas9 genome editing system for molecular breeding of orchids. Indones. J. Biotechnol. 2020, 25, 61–68. [Google Scholar] [CrossRef]
- Andersson, M.; Turesson, H.; Nicolia, A.; Fält, A.-S.; Samuelsson, M.; Hofvander, P. Efficient targeted multiallelic mutagenesis in tetraploid potato (Solanum Ttuberosum) by transient CRISPR-Cas9 expression in protoplasts. Plant. Cell Rep. 2017, 36, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Zha, M.; Huang, J.; Li, L.; Imran, M.; Zhang, C. StMYB44 negatively regulates phosphate transport by suppressing expression of PHOSPHATE1 in potato. J. Exp. Bot. 2017, 68, 1265–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veillet, F.; Perrot, L.; Chauvin, L.; Kermarrec, M.-P.; Guyon-Debast, A.; Chauvin, J.-E.; Nogué, F.; Mazier, M. Transgene-free genome editing in tomato and potato plants using agrobacterium-mediated delivery of a CRISPR/Cas9 cytidine base editor. Int. J. Mol. Sci. 2019, 20, 402. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Li, X.; Zhou, Z.; Wu, P.; Fang, M.; Pan, X.; Lin, Q.; Luo, W.; Wu, G.; Li, H. Reassessment of the four yield-related genes Gn1a, DEP1, GS3, and IPA1 in rice using a CRISPR/Cas9 system. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Yang, Y.; Qin, R.; Li, H.; Qiu, C.; Li, L.; Wei, P.; Yang, J. Rapid improvement of grain weight via highly efficient CRISPR/Cas9-mediated multiplex genome editing in rice. J. Genet. Genom. 2016, 43, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, J.; Wei, P.; Zhang, B.; Gou, F.; Feng, Z.; Mao, Y.; Yang, L.; Zhang, H.; Xu, N.; et al. The CRISPR/Cas9 system produces specific and homozygous targeted gene editing in rice in one generation. Plant Biotechnol. J. 2014, 12, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Nieves-Cordones, M.; Mohamed, S.; Tanoi, K.; Kobayashi, N.I.; Takagi, K.; Vernet, A.; Guiderdoni, E.; Périn, C.; Sentenac, H.; Véry, A.-A. Production of low-Cs+ rice plants by inactivation of the K+ transporter OsHAK1 with the CRISPR-cas system. Plant J. 2017, 92, 43–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, X.; Zheng, Y.; Xiao, K.; Wei, Y.; Zhu, Y.; Cai, Q.; Chen, L.; Xie, H.; Zhang, J. OsPRX2 contributes to stomatal closure and improves potassium deficiency tolerance in rice. Biochem. Biophys. Res. Commun. 2018, 495, 461–467. [Google Scholar] [CrossRef]
- Shen, C.; Que, Z.; Xia, Y.; Tang, N.; Li, D.; He, R.; Cao, M. Knock out of the annexin gene OsAnn3 via CRISPR/Cas9-mediated genome editing decreased cold tolerance in rice. J. Plant Biol. 2017, 60, 539–547. [Google Scholar] [CrossRef]
- Li, C.; Zong, Y.; Wang, Y.; Jin, S.; Zhang, D.; Song, Q.; Zhang, R.; Gao, C. Expanded base editing in rice and wheat using a Cas9-adenosine deaminase fusion. Genome Biol. 2018, 19, 59. [Google Scholar] [CrossRef] [Green Version]
- Iaffaldano, B.; Zhang, Y.; Cornish, K. CRISPR/Cas9 genome editing of rubber producing dandelion Taraxacum kok-saghyz using Agrobacterium rhizogenes without selection. Ind. Crops Prod. 2016, 89, 356–362. [Google Scholar] [CrossRef] [Green Version]
- Bao, A.; Chen, H.; Chen, L.; Chen, S.; Hao, Q.; Guo, W.; Qiu, D.; Shan, Z.; Yang, Z.; Yuan, S.; et al. CRISPR/Cas9-mediated targeted mutagenesis of GmSPL9 genes alters plant architecture in soybean. BMC Plant Biol. 2019, 19, 131. [Google Scholar] [CrossRef] [Green Version]
- Ueta, R.; Abe, C.; Watanabe, T.; Sugano, S.S.; Ishihara, R.; Ezura, H.; Osakabe, Y.; Osakabe, K. Rapid breeding of parthenocarpic tomato plants using CRISPR/Cas9. Sci. Rep. 2017, 7, 507. [Google Scholar] [CrossRef] [Green Version]
- Ito, Y.; Nishizawa-Yokoi, A.; Endo, M.; Mikami, M.; Toki, S. CRISPR/Cas9-mediated mutagenesis of the RIN locus that regulates tomato fruit ripening. Biochem. Biophys. Res. Commun. 2015, 467, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Tavano, E.C.D.R.; Lammers, M.; Martinelli, A.P.; Angenent, G.C.; de Maagd, R.A. Re-evaluation of transcription factor function in tomato fruit development and ripening with CRISPR/Cas9-mutagenesis. Sci. Rep. 2019, 9, 1696. [Google Scholar] [CrossRef]
- Brooks, C.; Nekrasov, V.; Lippman, Z.B.; Van Eck, J. Efficient gene editing in tomato in the first generation using the clustered regularly interspaced short palindromic repeats/CRISPR-associated9 system1. Plant Physiol. 2014, 166, 1292–1297. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Liu, C.; Zhao, R.; Wang, L.; Chen, L.; Yu, W.; Zhang, S.; Sheng, J.; Shen, L. CRISPR/Cas9-mediated SlNPR1 mutagenesis reduces tomato plant drought tolerance. BMC Plant Biol. 2019, 19, 38. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Zhang, R.; Song, G.; Gao, J.; Li, W.; Han, X.; Chen, M.; Li, Y.; Li, G. Targeted mutagenesis using the Agrobacterium tumefaciens-mediated CRISPR-Cas9 system in common wheat. BMC Plant Biol. 2018, 18, 302. [Google Scholar] [CrossRef]
- Liu, H.; Wang, K.; Jia, Z.; Gong, Q.; Lin, Z.; Du, L.; Pei, X.; Ye, X. Efficient induction of haploid plants in wheat by editing of TaMTL using an optimized agrobacterium-mediated CRISPR System. J. Exp. Bot. 2020, 71, 1337–1349. [Google Scholar] [CrossRef] [PubMed]
- Von Caemmerer, S.; Quick, W.P.; Furbank, R.T. The development of C4 rice: Current progress and future challenges. Science 2012, 336, 1671–1672. [Google Scholar] [CrossRef] [PubMed]
- Salesse-Smith, C.E.; Sharwood, R.E.; Busch, F.A.; Kromdijk, J.; Bardal, V.; Stern, D.B. Overexpression of rubisco subunits with RAF1 increases rubisco content in maize. Nat. Plants 2018, 4, 802–810. [Google Scholar] [CrossRef]
- Li, T.; Yang, X.; Yu, Y.; Si, X.; Zhai, X.; Zhang, H.; Dong, W.; Gao, C.; Xu, C. Domestication of wild tomato is accelerated by genome editing. Nat. Biotechnol. 2018. [Google Scholar] [CrossRef]
- Kosicki, M.; Tomberg, K.; Bradley, A. Repair of double-strand breaks induced by CRISPR-Cas9 leads to large deletions and complex rearrangements. Nat. Biotechnol. 2018, 36, 765–771. [Google Scholar] [CrossRef]
- Khan, M.Z.; Amin, I.; Hameed, A.; Mansoor, S. CRISPR-Cas13a: Prospects for plant virus resistance. Trends Biotechnol. 2018, 36, 1207–1210. [Google Scholar] [CrossRef]
- Min, Y.-L.; Li, H.; Rodriguez-Caycedo, C.; Mireault, A.A.; Huang, J.; Shelton, J.M.; McAnally, J.R.; Amoasii, L.; Mammen, P.P.A.; Bassel-Duby, R.; et al. CRISPR-Cas9 corrects duchenne muscular dystrophy exon 44 deletion mutations in mice and human cells. Sci. Adv. 2019, 5, eaav4324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjursell, M.; Porritt, M.J.; Ericson, E.; Taheri-Ghahfarokhi, A.; Clausen, M.; Magnusson, L.; Admyre, T.; Nitsch, R.; Mayr, L.; Aasehaug, L.; et al. Therapeutic genome editing with CRISPR/Cas9 in a humanized mouse model ameliorates A1-antitrypsin deficiency phenotype. EBioMedicine 2018, 29, 104–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohmori, T.; Mizukami, H.; Ozawa, K.; Sakata, Y.; Nishimura, S. New approaches to gene and cell therapy for hemophilia. J. Thromb. Haemost. 2015, 13, S133–S142. [Google Scholar] [CrossRef] [Green Version]
- Khosravi, M.A.; Abbasalipour, M.; Concordet, J.-P.; Berg, J.V.; Zeinali, S.; Arashkia, A.; Azadmanesh, K.; Buch, T.; Karimipoor, M. Targeted deletion of BCL11A gene by CRISPR-Cas9 system for fetal hemoglobin reactivation: A promising approach for gene therapy of beta thalassemia disease. Eur. J. Pharmacol. 2019, 854, 398–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- György, B.; Nist-Lund, C.; Pan, B.; Asai, Y.; Karavitaki, K.D.; Kleinstiver, B.P.; Garcia, S.P.; Zaborowski, M.P.; Solanes, P.; Spataro, S.; et al. Allele-specific gene editing prevents deafness in a model of dominant progressive hearing loss. Nat. Med. 2019, 25, 1123–1130. [Google Scholar] [CrossRef]
- Dever, D.P.; Bak, R.O.; Reinisch, A.; Camarena, J.; Washington, G.; Nicolas, C.E.; Pavel-Dinu, M.; Saxena, N.; Wilkens, A.B.; Mantri, S.; et al. CRISPR/Cas9 β-globin gene targeting in human haematopoietic stem cells. Nature 2016, 539, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Isgrò, A.; Gaziev, J.; Sodani, P.; Lucarelli, G. Progress in hematopoietic stem cell transplantation as allogeneic cellular gene therapy in thalassemia. Ann. N. Y. Acad. Sci. 2010, 1202, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Traylen, C.M.; Patel, H.R.; Fondaw, W.; Mahatme, S.; Williams, J.F.; Walker, L.R.; Dyson, O.F.; Arce, S.; Akula, S.M. Virus reactivation: A panoramic view in human infections. Future Virol. 2011, 6, 451–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craigie, R.; Bushman, F.D. HIV DNA integration. Cold Spring Harb. Perspect. Med. 2012, 2, a006890. [Google Scholar] [CrossRef] [Green Version]
- Finzi, D.; Blankson, J.; Siliciano, J.D.; Margolick, J.B.; Chadwick, K.; Pierson, T.; Smith, K.; Lisziewicz, J.; Lori, F.; Flexner, C.; et al. Latent infection of CD4+ T cells provides a mechanism for lifelong persistence of HIV-1, even in patients on effective combination therapy. Nat. Med. 1999, 5, 512–517. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Kaminski, R.; Yang, F.; Zhang, Y.; Cosentino, L.; Li, F.; Luo, B.; Alvarez-Carbonell, D.; Garcia-Mesa, Y.; Karn, J.; et al. RNA-directed gene editing specifically eradicates latent and prevents new HIV-1 infection. Proc. Natl. Acad. Sci. USA 2014, 111, 11461–11466. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.W.; Kim, S.; Kim, J.M.; Kim, J.-S. Targeted genome engineering in human cells with the Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31, 230–232. [Google Scholar] [CrossRef]
- Hou, P.; Chen, S.; Wang, S.; Yu, X.; Chen, Y.; Jiang, M.; Zhuang, K.; Ho, W.; Hou, W.; Huang, J.; et al. Genome editing of CXCR4 by CRISPR/Cas9 confers cells resistant to HIV-1 infection. Sci. Rep. 2015, 5, 15577. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, S.; Jin, X.; Wang, Q.; Yang, K.; Li, C.; Xiao, Q.; Hou, P.; Liu, S.; Wu, S.; et al. Genome editing of the HIV co-receptors CCR5 and CXCR4 by CRISPR-Cas9 protects CD4+ T cells from HIV-1 infection. Cell Biosci. 2017, 7, 1–15. [Google Scholar] [CrossRef]
- Yu, S.; Yao, Y.; Xiao, H.; Li, J.; Liu, Q.; Yang, Y.; Adah, D.; Lu, J.; Zhao, S.; Qin, L.; et al. Simultaneous knockout of CXCR4 and CCR5 genes in CD4+ T cells via CRISPR/Cas9 confers resistance to both X4- and R5-tropic human immunodeficiency virus type 1 infection. Hum. Gene. Ther. 2018, 29, 51–67. [Google Scholar] [CrossRef]
- Dash, P.K.; Kaminski, R.; Bella, R.; Su, H.; Mathews, S.; Ahooyi, T.M.; Chen, C.; Mancuso, P.; Sariyer, R.; Ferrante, P.; et al. Sequential LASER ART and CRISPR treatments eliminate HIV-1 in a subset of infected humanized mice. Nat. Commun. 2019, 10, 2753. [Google Scholar] [CrossRef] [Green Version]
- Rusconi, S.; Giacomelli, A. CRISPR in HIV: Dangers of CCR5 deletion. Future Virol. 2020, 15, 207–209. [Google Scholar] [CrossRef]
- Gao, Z.; Fan, M.; Das, A.T.; Herrera-Carrillo, E.; Berkhout, B. Extinction of all infectious HIV in cell culture by the CRISPR-Cas12a system with only a single CrRNA. Nucleic Acids Res. 2020, 48, 5527–5539. [Google Scholar] [CrossRef]
- Nouri, R.; Jiang, Y.; Lian, X.L.; Guan, W. Sequence-specific recognition of HIV-1 DNA with solid-state CRISPR-Cas12a-assisted nanopores (SCAN). ACS Sens. 2020, 5, 1273–1280. [Google Scholar] [CrossRef]
- Ding, X.; Yin, K.; Li, Z.; Liu, C. All-in-one dual CRISPR-Cas12a (AIOD-CRISPR) assay: A case for rapid, ultrasensitive and visual detection of novel Coronavirus SARS-CoV-2 and HIV Virus. bioRxiv 2020. [Google Scholar] [CrossRef]
- Hou, T.; Zeng, W.; Yang, M.; Chen, W.; Ren, L.; Ai, J.; Wu, J.; Liao, Y.; Gou, X.; Li, Y.; et al. Development and evaluation of A CRISPR-based diagnostic for 2019-novel Coronavirus. medRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Roehm, P.C.; Shekarabi, M.; Wollebo, H.S.; Bellizzi, A.; He, L.; Salkind, J.; Khalili, K. Inhibition of HSV-1 replication by gene editing strategy. Sci. Rep. 2016, 6, 23146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, C.; Tang, Y.; Wang, J.; Xiong, F.; Guo, C.; Wang, Y.; Xiang, B.; Zhou, M.; Li, X.; Wu, X.; et al. The emerging role of epstein-barr virus encoded microRNAs in nasopharyngeal carcinoma. J. Cancer 2018, 9, 2852–2864. [Google Scholar] [CrossRef] [Green Version]
- Van Diemen, F.R.; Kruse, E.M.; Hooykaas, M.J.G.; Bruggeling, C.E.; Schürch, A.C.; van Ham, P.M.; Imhof, S.M.; Nijhuis, M.; Wiertz, E.J.H.J.; Lebbink, R.J. CRISPR/Cas9-mediated genome editing of herpesviruses limits productive and latent infections. PLoS Pathog. 2016, 12, e1005701. [Google Scholar] [CrossRef] [PubMed]
- Tso, F.Y.; West, J.T.; Wood, C. Reduction of Kaposi’s sarcoma-associated herpesvirus latency using CRISPR-Cas9 to edit the latency-associated nuclear antigen gene. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Gergen, J.; Coulon, F.; Creneguy, A.; Elain-Duret, N.; Gutierrez, A.; Pinkenburg, O.; Verhoeyen, E.; Anegon, I.; Nguyen, T.H.; Halary, F.A.; et al. Multiplex CRISPR/Cas9 system impairs HCMV replication by excising an essential viral gene. PLoS ONE 2018, 13, e0192602. [Google Scholar] [CrossRef] [Green Version]
- Moffett, H.F.; Harms, C.K.; Fitzpatrick, K.S.; Tooley, M.R.; Boonyaratanakornkit, J.; Taylor, J.J. B cells engineered to express pathogen-specific antibodies protect against infection. Sci. Immunol. 2019, 4. [Google Scholar] [CrossRef]
- Wollebo, H.S.; Bellizzi, A.; Kaminski, R.; Hu, W.; White, M.K.; Khalili, K. CRISPR/Cas9 system as an agent for eliminating polyomavirus JC infection. PLoS ONE 2015, 10, e0136046. [Google Scholar] [CrossRef]
- Tian, X.; Gu, T.; Patel, S.; Bode, A.M.; Lee, M.-H.; Dong, Z. CRISPR/Cas9-an evolving biological tool kit for cancer biology and oncology. NPJ Precis. Oncol. 2019, 3, 8. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-H.; Lee, K.-C.; Doudna, J.A. Applications of CRISPR-Cas enzymes in cancer therapeutics and detection. Trends Cancer 2018, 4, 499–512. [Google Scholar] [CrossRef]
- Morris, L.G.T.; Chan, T.A. Therapeutic targeting of tumor suppressor genes. Cancer 2015, 121, 1357–1368. [Google Scholar] [CrossRef] [Green Version]
- Kodama, M.; Kodama, T.; Murakami, M. Oncogene activation and tumor suppressor gene inactivation find their sites of expression in the changes in time and space of the age-adjusted cancer incidence rate. In Vivo 2000, 14, 725–734. [Google Scholar] [PubMed]
- Bu, X.; Kato, J.; Hong, J.A.; Merino, M.J.; Schrump, D.S.; Lund, F.E.; Moss, J. CD38 knockout suppresses tumorigenesis in mice and clonogenic growth of human lung cancer cells. Carcinogenesis 2018, 39, 242–251. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.H.; Changou, C.A.; Hsieh, T.H.; Lee, Y.C.; Chu, C.Y.; Hsu, K.C.; Wang, H.C.; Lin, Y.C.; Lo, Y.N.; Liu, Y.R.; et al. Dual inhibition of PIK3C3 and FGFR as a new therapeutic approach to treat bladder cancer. Clin. Cancer res. Off. J. Am. Assoc. Cancer Res. 2018, 24, 1176–1189. [Google Scholar] [CrossRef] [Green Version]
- Takeda, H.; Kataoka, S.; Nakayama, M.; Ali, M.A.E.; Oshima, H.; Yamamoto, D.; Park, J.-W.; Takegami, Y.; An, T.; Jenkins, N.A.; et al. CRISPR-Cas9–mediated gene knockout in intestinal tumor organoids provides functional validation for colorectal cancer driver genes. PNAS 2019, 116, 15635–15644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artegiani, B.; van Voorthuijsen, L.; Lindeboom, R.G.H.; Seinstra, D.; Heo, I.; Tapia, P.; López-Iglesias, C.; Postrach, D.; Dayton, T.; Oka, R.; et al. Probing the tumor suppressor function of BAP1 in CRISPR-engineered human liver organoids. Cell Stem. Cell 2019, 24, 927–943.e6. [Google Scholar] [CrossRef] [PubMed]
- Eyquem, J.; Mansilla-Soto, J.; Giavridis, T.; van der Stegen, S.J.C.; Hamieh, M.; Cunanan, K.M.; Odak, A.; Gönen, M.; Sadelain, M. Targeting a CAR to the TRAC Locus with CRISPR/Cas9 enhances tumour rejection. Nature 2017, 543, 113–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, J.; Liu, X.; Fang, C.; Jiang, S.; June, C.H.; Zhao, Y. Multiplex genome editing to generate universal CAR T cells resistant to PD1 inhibition. Clin. Cancer Res. 2017, 23, 2255–2266. [Google Scholar] [CrossRef] [Green Version]
- Rupp, L.J.; Schumann, K.; Roybal, K.T.; Gate, R.E.; Ye, C.J.; Lim, W.A.; Marson, A. CRISPR/Cas9-Mediated PD-1 disruption enhances anti-tumor efficacy of human chimeric antigen receptor T cells. Sci. Rep. 2017, 7, 737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, R.-Y.; Francois, A.; McGray, A.R.; Miliotto, A.; Odunsi, K. Compensatory upregulation of PD-1, LAG-3, and CTLA-4 limits the efficacy of single-agent checkpoint blockade in metastatic ovarian cancer. Oncoimmunology 2017, 6, e1249561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Liu, Y.; Zhou, X.; Zhao, R.; Wang, H. Applications of CRISPR-Cas9 in gynecological cancer research. Clin. Genet 2020, 97, 827–834. [Google Scholar] [CrossRef]
- Firth, A.L.; Menon, T.; Parker, G.S.; Qualls, S.J.; Lewis, B.M.; Ke, E.; Dargitz, C.T.; Wright, R.; Khanna, A.; Gage, F.H.; et al. Functional gene correction for cystic fibrosis in lung epithelial cells generated from patient IPSCs. Cell Rep. 2015, 12, 1385–1390. [Google Scholar] [CrossRef] [Green Version]
- Duchêne, B.L.; Cherif, K.; Iyombe-Engembe, J.-P.; Guyon, A.; Rousseau, J.; Ouellet, D.L.; Barbeau, X.; Lague, P.; Tremblay, J.P. CRISPR-induced deletion with SaCas9 restores dystrophin expression in dystrophic models in vitro and in vivo. Mol. Ther. 2018, 26, 2604–2616. [Google Scholar] [CrossRef] [Green Version]
- Long, C.; Li, H.; Tiburcy, M.; Rodriguez-Caycedo, C.; Kyrychenko, V.; Zhou, H.; Zhang, Y.; Min, Y.-L.; Shelton, J.M.; Mammen, P.P.A.; et al. Correction of diverse muscular dystrophy mutations in human engineered heart muscle by single-site genome editing. Sci. Adv. 2018, 4, eaap9004. [Google Scholar] [CrossRef] [Green Version]
- Frangoul, H.; Altshuler, D.; Cappellini, M.D.; Chen, Y.-S.; Domm, J.; Eustace, B.K.; Foell, J.; de la Fuente, J.; Grupp, S.; Handgretinger, R.; et al. CRISPR-Cas9 gene editing for sickle cell disease and β-thalassemia. N. Engl. J. Med. 2021, 384, 252–260. [Google Scholar] [CrossRef]
- Vilarino, M.; Suchy, F.P.; Rashid, S.T.; Lindsay, H.; Reyes, J.; McNabb, B.R.; van der Meulen, T.; Huising, M.O.; Nakauchi, H.; Ross, P.J. Mosaicism diminishes the value of pre-implantation embryo biopsies for detecting CRISPR/Cas9 induced mutations in sheep. Transgenic Res. 2018, 27, 525–537. [Google Scholar] [CrossRef]
- Haston, S.; Pozzi, S.; Gonzalez-Meljem, J.M. Applications of CRISPR-cas in ageing research. Clin. Genet. Genom. Aging 2020, 213–230. [Google Scholar] [CrossRef]
- Yue, Y.; Kan, Y.; Xu, W.; Zhao, H.-Y.; Zhou, Y.; Song, X.; Wu, J.; Xiong, J.; Goswami, D.; Yang, M.; et al. Extensive mammalian germline genome engineering. bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Skill, N.; Kubal, S.; Fridell, J.; Ekser, B. Identification of novel xenoreactive non-gal antigens: Tetraspanin CD37 and CD81. Xenotransplantation 2017, 24, 27–28. [Google Scholar] [CrossRef] [Green Version]
- Ul Ain, Q.; Chung, J.Y.; Kim, Y.-H. Current and future delivery systems for engineered nucleases: ZFN, TALEN and RGEN. J. Control. Release 2015, 205, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhou, H.; Fan, X.; Zhang, Y.; Zhang, M.; Wang, Y.; Xie, Z.; Bai, M.; Yin, Q.; Liang, D.; et al. Correction of a genetic disease by CRISPR-Cas9-mediated gene editing in mouse spermatogonial stem cells. Cell Res. 2015, 25, 67–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flynn, R.; Grundmann, A.; Renz, P.; Hänseler, W.; James, W.S.; Cowley, S.A.; Moore, M.D. CRISPR-mediated genotypic and phenotypic correction of a chronic granulomatous disease mutation in human IPS cells. Exp. Hematol. 2015, 43, 838–848. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Cheng, Z.-Y.; Zhao, Z.-N.; Quan, X.-Q.; Wei, Y.; Xia, D.-S.; Li, J.-Q.; Hu, J.-L. Correlation of serum PCSK9 in CHD patients with the severity of coronary arterial lesions. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 1135–1139. [Google Scholar]
- Van Agtmaal, E.L.; André, L.M.; Willemse, M.; Cumming, S.A.; van Kessel, I.D.G.; van den Broek, W.J.A.A.; Gourdon, G.; Furling, D.; Mouly, V.; Monckton, D.G.; et al. CRISPR/Cas9-Induced (CTG⋅CAG)n repeat instability in the myotonic dystrophy type 1 locus: Implications for therapeutic genome editing. Mol. Ther. 2017, 25, 24–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.L.; Fujimoto, N.; Sasakawa, N.; Shirai, S.; Ohkame, T.; Sakuma, T.; Tanaka, M.; Amano, N.; Watanabe, A.; Sakurai, H.; et al. Precise correction of the dystrophin gene in duchenne muscular dystrophy patient induced pluripotent stem cells by TALEN and CRISPR-Cas9. Stem. Cell Rep. 2015, 4, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Monteys, A.M.; Ebanks, S.A.; Keiser, M.S.; Davidson, B.L. CRISPR/Cas9 editing of the mutant huntingtin allele in vitro and in vivo. Mol. Ther. 2017, 25, 12–23. [Google Scholar] [CrossRef] [Green Version]
- Canver, M.C.; Smith, E.C.; Sher, F.; Pinello, L.; Sanjana, N.E.; Shalem, O.; Chen, D.D.; Schupp, P.G.; Vinjamur, D.S.; Garcia, S.P.; et al. BCL11A enhancer dissection by Cas9-mediated in situ saturating mutagenesis. Nature 2015, 527, 192–197. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Zhang, Y.-P.; Song, L.; Luo, J.; Qi, W.; Hu, J.; Lu, D.; Yang, Z.; Zhang, J.; Xiao, J.; et al. Genome editing with CRISPR/Cas9 in postnatal mice corrects PRKAG2 cardiac syndrome. Cell Res. 2016, 26, 1099–1111. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yang, Y.; Kang, X.; Lin, B.; Yu, Q.; Song, B.; Gao, G.; Chen, Y.; Sun, X.; Li, X.; et al. One-step biallelic and scarless correction of a β-thalassemia mutation in patient-specific IPSCs without drug selection. Mol. Ther. Nucleic Acids 2017, 6, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Lavin, M.F.; Yeo, A.J.; Kijas, A.W.; Wolvetang, E.; Sly, P.D.; Wainwright, C.; Sinclair, K. Therapeutic targets and investigated treatments for ataxia-telangiectasia. Expert Opin. Orphan Drugs 2016, 4, 1263–1276. [Google Scholar] [CrossRef]
- Zhen, S.; Hua, L.; Takahashi, Y.; Narita, S.; Liu, Y.-H.; Li, Y. In vitro and in vivo growth suppression of human Papillomavirus 16-positive cervical cancer cells by CRISPR/Cas9. Biochem. Biophys. Res. Commun. 2014, 450, 1422–1426. [Google Scholar] [CrossRef]
- Ma, H.; Marti-Gutierrez, N.; Park, S.-W.; Wu, J.; Lee, Y.; Suzuki, K.; Koski, A.; Ji, D.; Hayama, T.; Ahmed, R.; et al. Correction of a pathogenic gene mutation in human embryos. Nature 2017, 548, 413–419. [Google Scholar] [CrossRef]
- Miyamoto, T.; Akutsu, S.N.; Tauchi, H.; Kudo, Y.; Tashiro, S.; Yamamoto, T.; Matsuura, S. Exploration of genetic basis underlying individual differences in radiosensitivity within human populations using genome editing technology. J. Radiat. Res. 2018, 59, ii75–ii82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Ng, W.-L.; Luster, T.A.; Hu, H.; Sviderskiy, V.O.; Dowling, C.M.; Hollinshead, K.E.R.; Zouitine, P.; Zhang, H.; Huang, Q.; et al. Epigenetic CRISPR screens identify Npm1 as a therapeutic vulnerability in non–small cell lung cancer. Cancer Res. 2020, 80, 3556–3567. [Google Scholar] [CrossRef]
- You, L.; Tong, R.; Li, M.; Liu, Y.; Xue, J.; Lu, Y. Advancements and obstacles of CRISPR-Cas9 technology in translational research. Mol. Ther. Methods Clin. Dev. 2019, 13, 359–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Sheng, C.; Wang, S.; Yang, L.; Liang, Y.; Huang, Y.; Liu, H.; Li, P.; Yang, C.; Yang, X.; et al. Removal of integrated hepatitis B virus DNA using CRISPR-Cas9. Front. Cell Infect. Microbiol. 2017, 7, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomaa, A.A.; Klumpe, H.E.; Luo, M.L.; Selle, K.; Barrangou, R.; Beisel, C.L. Programmable removal of bacterial strains by use of genome-targeting CRISPR-cas systems. mBio 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Yosef, I.; Manor, M.; Kiro, R.; Qimron, U. Temperate and lytic bacteriophages programmed to sensitize and kill antibiotic-resistant bacteria. PNAS 2015, 112, 7267–7272. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-S.; Cho, D.-H.; Park, M.; Chung, W.-J.; Shin, D.; Ko, K.S.; Kweon, D.-H. CRISPR/Cas9-mediated re-sensitization of antibiotic-resistant Escherichia coli harboring extended-spectrum β-lactamases. J. Microbiol. Biotechnol. 2016, 26, 394–401. [Google Scholar] [CrossRef]
- Bikard, D.; Euler, C.; Jiang, W.; Nussenzweig, P.M.; Goldberg, G.W.; Duportet, X.; Fischetti, V.A.; Marraffini, L.A. Development of sequence-specific antimicrobials based on programmable CRISPR-cas nucleases. Nat. Biotechnol. 2014, 32, 1146–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Li, X.; Zhou, W.-L.; Huang, Y.; Liang, X.; Jiang, L.; Yang, X.; Sun, J.; Li, Z.; Han, W.-D.; et al. Genetically engineered T cells for cancer immunotherapy. Signal Transduct. Target. Ther. 2019, 4, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Kyrou, K.; Hammond, A.M.; Galizi, R.; Kranjc, N.; Burt, A.; Beaghton, A.K.; Nolan, T.; Crisanti, A. A CRISPR–Cas9 gene drive targeting doublesex causes complete population suppression in caged Anopheles gambiae mosquitoes. Nat. Biotechnol. 2018, 36, 1062–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakočiūnas, T.; Jensen, M.K.; Keasling, J.D. CRISPR/Cas9 advances engineering of microbial cell factories. Metab. Eng. 2016, 34, 44–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakočiūnas, T.; Bonde, I.; Herrgård, M.; Harrison, S.J.; Kristensen, M.; Pedersen, L.E.; Jensen, M.K.; Keasling, J.D. Multiplex metabolic pathway engineering using CRISPR/Cas9 in Saccharomyces cerevisiae. Metab. Eng. 2015, 28, 213–222. [Google Scholar] [CrossRef]
- Li, Y.; Lin, Z.; Huang, C.; Zhang, Y.; Wang, Z.; Tang, Y.-J.; Chen, T.; Zhao, X. Metabolic engineering of Escherichia coli using CRISPR-Cas9 meditated genome editing. Metab. Eng. 2015, 31, 13–21. [Google Scholar] [CrossRef]
- Cho, J.S.; Choi, K.R.; Prabowo, C.P.S.; Shin, J.H.; Yang, D.; Jang, J.; Lee, S.Y. CRISPR/Cas9-coupled recombineering for metabolic engineering of Corynebacterium glutamicum. Metab. Eng. 2017, 42, 157–167. [Google Scholar] [CrossRef]
- Ferreira, R.; David, F.; Nielsen, J. Advancing Biotechnology with CRISPR/Cas9: Recent applications and patent landscape. J. Ind. Microbiol. Biotechnol. 2018, 45, 467–480. [Google Scholar] [CrossRef] [Green Version]
- Kuivanen, J.; Wang, Y.-M.J.; Richard, P. Engineering Aspergillus niger for galactaric acid production: Elimination of galactaric acid catabolism by using RNA sequencing and CRISPR/Cas9. Microb. Cell Factories 2016, 15, 210. [Google Scholar] [CrossRef] [Green Version]
- Siripong, W.; Angela, C.; Tanapongpipat, S.; Runguphan, W. Metabolic engineering of Pichia pastoris for production of isopentanol (3-Methyl-1-Butanol). Enzyme Microb. Technol. 2020, 138, 109557. [Google Scholar] [CrossRef]
- Xu, P.; Li, L.; Zhang, F.; Stephanopoulos, G.; Koffas, M. Improving fatty acids production by engineering dynamic pathway regulation and metabolic control. PNAS 2014, 111, 11299–11304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez, A.; Muñoz-Fernández, G.; Ledesma-Amaro, R.; Buey, R.M.; Revuelta, J.L. One-Vector CRISPR/Cas9 genome engineering of the industrial fungus Ashbya gossypii. Microb. Biotechnol. 2019, 12, 1293–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; An, C.; Shu, X.; Meng, X.; Yao, Y.; Zhang, J.; Chen, F.; Xiang, H.; Yang, S.; Gao, X.; et al. A dual-plasmid CRISPR/cas system for mycotoxin elimination in polykaryotic industrial fungi. ACS Synth. Biol. 2020, 9, 2087–2095. [Google Scholar] [CrossRef]
- Li, C.; Zhang, R.; Meng, X.; Chen, S.; Zong, Y.; Lu, C.; Qiu, J.-L.; Chen, Y.-H.; Li, J.; Gao, C. Targeted, random mutagenesis of plant genes with dual cytosine and adenine base editors. Nat. Biotechnol. 2020, 38, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Mougiakos, I.; Mohanraju, P.; Bosma, E.F.; Vrouwe, V.; Finger Bou, M.; Naduthodi, M.I.S.; Gussak, A.; Brinkman, R.B.L.; van Kranenburg, R.; van der Oost, J. Characterizing a thermostable Cas9 for bacterial genome editing and silencing. Nat. Commun. 2017, 8, 1647. [Google Scholar] [CrossRef] [Green Version]
- Lim, H.; Choi, S.-K. Programmed GRNA removal system for CRISPR-Cas9-mediated multi-round genome editing in Bacillus subtilis. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Altenbuchner, J. Editing of the Bacillus subtilis genome by the CRISPR-Cas9 system. Appl. Environ. Microbiol. 2016, 82, 5421–5427. [Google Scholar] [CrossRef] [Green Version]
- Nagaraju, S.; Davies, N.K.; Walker, D.J.F.; Köpke, M.; Simpson, S.D. Genome editing of Clostridium autoethanogenum using CRISPR/Cas9. Biotechnol. Biofuels 2016, 9, 219. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, Z.-T.; Seo, S.-O.; Lynn, P.; Lu, T.; Jin, Y.-S.; Blaschek, H.P. Gene transcription repression in Clostridium beijerinckii using CRISPR-DCas9. Biotechnol. Bioeng. 2016, 113, 2739–2743. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Huang, R.; Teng, L.; Jing, X.; Hu, J.; Cui, G.; Wang, Y.; Cui, Q.; Xu, J. Cellulosome stoichiometry in clostridium cellulolyticum is regulated by selective RNA processing and stabilization. Nat. Commun. 2015, 6, 6900. [Google Scholar] [CrossRef]
- Huang, H.; Chai, C.; Li, N.; Rowe, P.; Minton, N.P.; Yang, S.; Jiang, W.; Gu, Y. CRISPR/Cas9-based efficient genome editing in Clostridium ljungdahlii, an autotrophic gas-fermenting bacterium. ACS Synth. Biol. 2016, 5, 1355–1361. [Google Scholar] [CrossRef] [PubMed]
- Pyne, M.E.; Sokolenko, S.; Liu, X.; Srirangan, K.; Bruder, M.R.; Aucoin, M.G.; Moo-Young, M.; Chung, D.A.; Chou, C.P. Disruption of the reductive 1,3-propanediol pathway triggers production of 1,2-propanediol for sustained glycerol fermentation by Clostridium Ppasteurianum. Appl. Environ. Microbiol. 2016, 82, 5375–5388. [Google Scholar] [CrossRef] [Green Version]
- Cleto, S.; Jensen, J.V.; Wendisch, V.F.; Lu, T.K. Corynebacterium Gglutamicum metabolic engineering with CRISPR interference (CRISPRi). ACS Synth. Biol. 2016, 5, 375–385. [Google Scholar] [CrossRef]
- Li, H.; Shen, C.R.; Huang, C.-H.; Sung, L.-Y.; Wu, M.-Y.; Hu, Y.-C. CRISPR-Cas9 for the genome engineering of cyanobacteria and succinate production. Metabol. Eng. 2016, 38, 293–302. [Google Scholar] [CrossRef]
- Zhang, S.; Guo, F.; Yan, W.; Dai, Z.; Dong, W.; Zhou, J.; Zhang, W.; Xin, F.; Jiang, M. Recent advances of CRISPR/Cas9-based genetic engineering and transcriptional regulation in industrial biology. Front. Bioeng. Biotechnol. 2020, 7. [Google Scholar] [CrossRef] [Green Version]
- Donohoue, P.D.; Barrangou, R.; May, A.P. Advances in industrial biotechnology using CRISPR-cas systems. Trends Biotechnol. 2018, 36, 134–146. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.-H.; van Pijkeren, J.-P. CRISPR-Cas9-assisted recombineering in Lactobacillus reuteri. Nucleic Acids Res. 2014, 42, e131. [Google Scholar] [CrossRef] [PubMed]
- Hao, M.; Cui, Y.; Qu, X. Analysis of CRISPR-cas system in Streptococcus thermophilus and its application. Front. Microbiol. 2018, 9, 257. [Google Scholar] [CrossRef]
- Zhang, M.M.; Wong, F.T.; Wang, Y.; Luo, S.; Lim, Y.H.; Heng, E.; Yeo, W.L.; Cobb, R.E.; Enghiad, B.; Ang, E.L.; et al. CRISPR–Cas9 strategy for activation of silent Streptomyces biosynthetic gene clusters. Nat. Chem. Biol. 2017, 13, 607–609. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Zheng, G.; Jiang, W.; Hu, H.; Lu, Y. One-step high-efficiency CRISPR/Cas9-mediated genome editing in Streptomyces. Acta Biochim. Biophys. Sin. 2015, 47, 231–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, H.; Zhang, L.; Wang, T.; Han, J.; Tang, H.; Zhang, L. Development of a CRISPR/Cas9-mediated gene-editing tool in Streptomyces rimosus. Microbiology 2017, 163, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.H.; Wong, F.T.; Yeo, W.L.; Ching, K.C.; Lim, Y.W.; Heng, E.; Chen, S.; Tsai, D.-J.; Lauderdale, T.-L.; Shia, K.-S.; et al. Auroramycin: A potent antibiotic from Streptomyces roseosporus by CRISPR-Cas9 activation. ChemBioChem 2018, 19, 1716–1719. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, X.; Wang, Q.; Xu, J.; Dong, F.; Yang, S.; Yang, J.; Zhang, Z.; Qian, Y.; Chen, J.; et al. Multicopy chromosomal integration using CRISPR-associated transposases. ACS Synth. Biol. 2020, 9, 1998–2008. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, B.; Duan, C.; Sun, B.; Yang, J.; Yang, S. Multigene editing in the Escherichia coli genome via the CRISPR-Cas9 system. Appl. Environ. Microbiol. 2015, 81, 2506–2514. [Google Scholar] [CrossRef] [Green Version]
- Wenderoth, M.; Pinecker, C.; Voß, B.; Fischer, R. Establishment of CRISPR/Cas9 in Alternaria alternata. Fungal Genet. Biol. 2017, 101, 55–60. [Google Scholar] [CrossRef]
- Nødvig, C.S.; Nielsen, J.B.; Kogle, M.E.; Mortensen, U.H. A CRISPR-Cas9 system for genetic engineering of filamentous fungi. PLoS ONE 2015, 10, e0133085. [Google Scholar] [CrossRef] [Green Version]
- Nødvig, C.S.; Hoof, J.B.; Kogle, M.E.; Jarczynska, Z.D.; Lehmbeck, J.; Klitgaard, D.K.; Mortensen, U.H. Efficient oligo nucleotide mediated CRISPR-Cas9 gene editing in aspergilli. Fungal Genet. Biol. 2018, 115, 78–89. [Google Scholar] [CrossRef] [Green Version]
- Weyda, I.; Yang, L.; Vang, J.; Ahring, B.K.; Lübeck, M.; Lübeck, P.S. A comparison of agrobacterium-mediated transformation and protoplast-mediated transformation with CRISPR-Cas9 and bipartite gene targeting substrates, as effective gene targeting tools for Aspergillus carbonarius. J. Microbiol. Methods 2017, 135, 26–34. [Google Scholar] [CrossRef]
- Fuller, K.K.; Chen, S.; Loros, J.J.; Dunlap, J.C. Development of the CRISPR/Cas9 system for targeted gene disruption in Aspergillus fumigatus. Eukaryot. Cell 2015, 14, 1073–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadooka, C.; Yamaguchi, M.; Okutsu, K.; Yoshizaki, Y.; Takamine, K.; Katayama, T.; Maruyama, J.-I.; Tamaki, H.; Futagami, T. A CRISPR/Cas9-mediated gene knockout system in Aspergillus luchuensis mut. Kawachii. Biosci. Biotechnol. Biochem. 2020, 84, 2179–2183. [Google Scholar] [CrossRef] [PubMed]
- Katayama, T.; Tanaka, Y.; Okabe, T.; Nakamura, H.; Fujii, W.; Kitamoto, K.; Maruyama, J. Development of a genome editing technique using the CRISPR/Cas9 system in the industrial filamentous fungus Aspergillus oryzae. Biotechnol. Lett. 2016, 38, 637–642. [Google Scholar] [CrossRef]
- Min, K.; Ichikawa, Y.; Woolford, C.A.; Mitchell, A.P. Candida albicans gene deletion with a transient CRISPR-Cas9 System. mSphere 2016, 1. [Google Scholar] [CrossRef] [Green Version]
- Enkler, L.; Richer, D.; Marchand, A.L.; Ferrandon, D.; Jossinet, F. Genome engineering in the yeast pathogen Candida glabrata using the CRISPR-Cas9 System. Sci Rep 2016, 6, 35766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wei, D.; Zhu, X.; Pan, J.; Zhang, P.; Huo, L.; Zhu, X. A ‘Suicide’ CRISPR-Cas9 system to promote gene deletion and restoration by electroporation in Cryptococcus neoformans. Sci. Rep. 2016, 6, 31145. [Google Scholar] [CrossRef] [Green Version]
- Shi, T.-Q.; Gao, J.; Wang, W.-J.; Wang, K.-F.; Xu, G.-Q.; Huang, H.; Ji, X.-J. CRISPR/Cas9-based genome editing in the filamentous Fungus Fusarium fujikuroi and its application in strain engineering for gibberellic acid production. ACS Synth. Biol. 2019, 8, 445–454. [Google Scholar] [CrossRef]
- Qin, H.; Xiao, H.; Zou, G.; Zhou, Z.; Zhong, J.-J. CRISPR-Cas9 assisted gene disruption in the higher fungus Ganoderma species. Process Biochem. 2017, 56, 57–61. [Google Scholar] [CrossRef]
- Wilson, A.M.; Wingfield, B.D. CRISPR-Cas9-mediated genome editing in the Filamentous Ascomycete Huntiella omanensis. J. Vis. Exp. 2020. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, A.A.; Walter, J.M.; Schubert, M.G.; Kung, S.H.; Hawkins, K.; Platt, D.M.; Hernday, A.D.; Mahatdejkul-Meadows, T.; Szeto, W.; Chandran, S.S.; et al. Efficient multiplexed integration of synergistic alleles and metabolic pathways in yeasts via CRISPR-Cas. Cell Syst. 2015, 1, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Gao, R.; Li, J.; Lin, L.; Zhao, J.; Sun, W.; Tian, C. Development of a genome-editing CRISPR/Cas9 system in thermophilic fungal Myceliophthora species and its application to hyper-cellulase production strain engineering. Biotechnol. Biofuels 2017, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Matsu-ura, T.; Baek, M.; Kwon, J.; Hong, C. Efficient gene editing in Neurospora crassa with CRISPR technology. Fungal Biol. Biotechnol. 2015, 2, s40694–s40715. [Google Scholar] [CrossRef] [Green Version]
- Pohl, C.; Kiel, J.A.K.W.; Driessen, A.J.M.; Bovenberg, R.A.L.; Nygård, Y. CRISPR/Cas9 based genome editing of Penicillium chrysogenum. ACS Synth. Biol. 2016, 5, 754–764. [Google Scholar] [CrossRef]
- Fang, Y.; Tyler, B.M. Efficient disruption and replacement of an effector gene in the oomycete Phytophthora sojae using CRISPR/Cas9. Mol. Plant Pathol. 2016, 17, 127–139. [Google Scholar] [CrossRef]
- Jacobs, J.Z.; Ciccaglione, K.M.; Tournier, V.; Zaratiegui, M. Implementation of the CRISPR-Cas9 System in fission yeast. Nat. Commun. 2014, 5, 5344. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.L.; Isbrandt, T.; Rasmussen, K.B.; Thrane, U.; Hoof, J.B.; Larsen, T.O.; Mortensen, U.H. Genes linked to production of secondary metabolites in Talaromyces atroroseus revealed using CRISPR-Cas9. PLoS ONE 2017, 12, e0169712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.; Chen, L.; Jiang, Y.; Zhou, Z.; Zou, G. Efficient genome editing in Filamentous Fungus Trichoderma reesei Uusing the CRISPR/Cas9 System. Cell Discov. 2015, 1, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Schuster, M.; Schweizer, G.; Reissmann, S.; Kahmann, R. Genome editing in Ustilago maydis using the CRISPR-cas system. Fungal. Genet. Biol. 2016, 89, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, C.M.; Hussain, M.S.; Blenner, M.; Wheeldon, I. Synthetic RNA Polymerase III promoters facilitate high-efficiency CRISPR-Cas9-mediated genome editing in Yarrowia lipolytica. ACS Synth. Biol. 2016, 5, 356–359. [Google Scholar] [CrossRef]
- Kim, Y.G.; Cha, J.; Chandrasegaran, S. Hybrid restriction enzymes: Zinc finger fusions to fok I Cleavage domain. PNAS 1996, 93, 1156–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cermak, T.; Doyle, E.L.; Christian, M.; Wang, L.; Zhang, Y.; Schmidt, C.; Baller, J.A.; Somia, N.V.; Bogdanove, A.J.; Voytas, D.F. Efficient design and assembly of custom TALEN and Other TAL effector-based constructs for DNA Targeting. Nucleic Acids Res. 2011, 39, e82. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; La Russa, M.; Qi, L.S. CRISPR/Cas9 in genome editing and beyond. Annu. Rev. Biochem. 2016, 85, 227–264. [Google Scholar] [CrossRef] [Green Version]
- Sakuma, T.; Nishikawa, A.; Kume, S.; Chayama, K.; Yamamoto, T. Multiplex genome engineering in human cells using all-in-one CRISPR/Cas9 Vector System. Sci. Rep. 2014, 4, 5400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirier, J.T. CRISPR libraries and screening. Prog. Mol. Biol. Transl. Sci. 2017, 152, 69–82. [Google Scholar] [CrossRef]
- Zhang, J.-H.; Adikaram, P.; Pandey, M.; Genis, A.; Simonds, W.F. Optimization of genome editing through CRISPR-Cas9 engineering. Bioengineered 2016, 7, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Li, C.; Gao, C. Applications of CRISPR–cas in agriculture and plant biotechnology. Nat. Rev. Mol. Cell Biol. 2020, 21, 661–677. [Google Scholar] [CrossRef]
- Sedeek, K.E.M.; Mahas, A.; Mahfouz, M. Plant genome engineering for targeted improvement of crop traits. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/RL (accessed on 11 March 2021).
- Biotech Crop Highlights in 2018 | ISAAA.Org. Available online: https://www.isaaa.org/resources/publications/pocketk/16/ (accessed on 14 March 2021).
- PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/ (accessed on 14 March 2021).
- Cho, S.W.; Kim, S.; Kim, Y.; Kweon, J.; Kim, H.S.; Bae, S.; Kim, J.-S. Analysis of off-target effects of CRISPR/Cas-Derived RNA-guided endonucleases and nickases. Genome Res. 2014, 24, 132–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soga, K.; Nakamura, K.; Ishigaki, T.; Kimata, S.; Ohmori, K.; Kishine, M.; Mano, J.; Takabatake, R.; Kitta, K.; Nagoya, H.; et al. Development of a novel method for specific detection of genetically modified atlantic salmon, aquadvantage, using real-time polymerase chain reaction. Food Chem. 2020, 305, 125426. [Google Scholar] [CrossRef]
- Caplan, A.L.; Parent, B.; Shen, M.; Plunkett, C. No time to waste—the ethical challenges created by CRISPR. EMBO Rep. 2015, 16, 1421–1426. [Google Scholar] [CrossRef]
- Scudellari, M. Self-destructing mosquitoes and sterilized rodents: The promise of gene drives. Nature 2019, 571, 160–162. [Google Scholar] [CrossRef]
- Schleidgen, S.; Dederer, H.-G.; Sgodda, S.; Cravcisin, S.; Lüneburg, L.; Cantz, T.; Heinemann, T. Human germline editing in the era of CRISPR-Cas: Risk and uncertainty, inter-generational responsibility, therapeutic legitimacy. BMC Med. Ethics 2020, 21, 87. [Google Scholar] [CrossRef]
- Furtado, R.N.; Furtado, R.N. Gene Editing: The risks and benefits of modifying human DNA. Rev. Bioética 2019, 27, 223–233. [Google Scholar] [CrossRef] [Green Version]
- Locke, L.G. The Promise of CRISPR for human germline editing and the perils of “Playing God”. CRISPR J. 2020, 3, 27–31. [Google Scholar] [CrossRef]
- Ihry, R.J.; Worringer, K.A.; Salick, M.R.; Frias, E.; Ho, D.; Theriault, K.; Kommineni, S.; Chen, J.; Sondey, M.; Ye, C.; et al. P53 Inhibits CRISPR-Cas9 engineering in human pluripotent stem cells. Nat. Med. 2018, 24, 939–946. [Google Scholar] [CrossRef]
- Haapaniemi, E.; Botla, S.; Persson, J.; Schmierer, B.; Taipale, J. CRISPR–Cas9 genome editing induces a P53-mediated DNA damage response. Nat. Med. 2018, 24, 927–930. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Foden, J.A.; Khayter, C.; Maeder, M.L.; Reyon, D.; Joung, J.K.; Sander, J.D. High-frequency off-target mutagenesis induced by CRISPR-cas nucleases in human cells. Nat. Biotechnol. 2013, 31, 822–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferdosi, S.R.; Ewaisha, R.; Moghadam, F.; Krishna, S.; Park, J.G.; Ebrahimkhani, M.R.; Kiani, S.; Anderson, K.S. Multifunctional CRISPR-Cas9 with engineered immunosilenced human T cell epitopes. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Pawluk, A.; Davidson, A.R.; Maxwell, K.L. Anti-CRISPR: Discovery, mechanism and function. Nat. Rev. Microbiol. 2018, 16, 12–17. [Google Scholar] [CrossRef]
- DiEuliis, D.; Giordano, J. Why gene editors like CRISPR/Cas may be a game-changer for neuroweapons. Health Secur. 2017, 15, 296–302. [Google Scholar] [CrossRef]
- DiEuliis, D.; Giordano, J. Gene editing using CRISPR/Cas9: Implications for dual-use and biosecurity. Protein Cell 2018, 9, 239–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, R.M.; Gronvall, G.K. CRISPR Cautions: Biosecurity implications of gene editing. Perspect. Biol. Med. 2020, 63, 73–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayanoğlu, F.B.; Elçin, A.E.; Elçin, Y.M. Bioethical issues in genome editing by CRISPR-Cas9 technology. Turk. J. Biol. 2020, 44, 110–120. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Association in Type or Subtype | Function |
|---|---|---|
| Cas1 | I, II, IV, IV (assumed) III-A, III-B | DNA nuclease |
| Cas2 | I, II, V III-A, III-B, VI (some) | RNA nuclease |
| Cas3 | I | DNA nuclease and helicase |
| Cas4 | II, V I (most) | DNA nuclease |
| Cas5 | IV I-C, III (some) | pre-crRNA processing |
| Cas6 | I (most), III-A, III-B | pre-crRNA processing |
| Cas7 | I, III, IV | RNA recognition, crRNA binding |
| Cas8 | I (most) | large subunit of Cascade complex |
| Cas9 | II | DNA nuclease |
| Cas10 | I (some), III (most) | large subunit of Csm or Cmr complex |
| Cas11 | III I (some), IV (some) | small subunit of effector complexes |
| Cas12 | V | crRNA processing, DNA nuclease |
| Cas13 | VI | crRNA processing, RNA nuclease |
| Adaptation | Expression | Interference | |||
|---|---|---|---|---|---|
| Spacer Integration | pre-crRNA Processing | Effector Complex | Target Cleavage | ||
| class 1 | type I | Cas1, Cas2, Cas4 | Cas6 | Cas7, Cas5, SS *, Cas8/LS | Cas3″, Cas3′ |
| type III | Cas1, Cas2, RT | Cas6 | Cas7, Cas5, SS, Cas10/LS | Cas10/LS | |
| type IV | Cas1, Cas2 | Cas6 | Cas7, Cas5, SS, Csf1/LS | ? | |
| class 2 | type II | Cas1, Cas2, Cas4 | RNase III | Cas9 | Cas9 |
| type V | Cas1, Cas2, Cas4 | Cas12 | Cas12 | Cas12 | |
| type VI | Cas1, Cas2 | Cas13 | Cas13 | Cas13 | |
| Class | Type | Subtype | Variant | Native Target | Origin |
|---|---|---|---|---|---|
| 1 | I | I-A | DNA | Archaeoglobus fulgidus (AF1859, AF1870–AF1879) | |
| I-B | DNA | Clostridium kluyveri (CKL_2758–CKL_2751) | |||
| I-C | DNA | Bacillus halodurans (BH0336–BH0342) | |||
| I-D | DNA | Cyanothece sp. 8802 (Cyan8802_0527–Cyan8802_0520) | |||
| I-E | DNA | Escherichia coli K12 (ygcB–ygbF) | |||
| I-F | I-F1 | DNA | Yersinia pseudo-tuberculosis (YPK_1644–YPK-1649) | ||
| I-F2 | DNA | Shewanella putrefaciens CN-32 (Sputcn32_1819–Sputcn32_1823) | |||
| I-F3 | Vibrio crassostreae J5 20 (VCR20J5_310088–VCR20J5_310108) | ||||
| I-G | DNA | Geobacter sulfurreducens (GSU0051–GSU0054, GSU0057–GSU0058) | |||
| III | III-A | DNA + RNA | Staphylococcus epidermidis (SERP2463–SERP2455) | ||
| III-B | DNA + RNA | Pyrococcus furiosus (PF1131–PF1124) | |||
| III-C | DNA + RNA | Methanothermobacter thermautotrophicus (MTH328–MTH323) | |||
| III-D | RNA? | Synechocystis sp. 6803 (sll7067–sll7063) | |||
| III-E | RNA? | Candidatus Scalidua brodae (SCABRO_02601, SCABRO_02597, SCABRO_02593, SCABRO_02595) | |||
| III-F | DNA? | Thermotoga lettingae TMO (Tlet_0097–Tlet_0100) | |||
| IV | IV-A | Thioalkalivibrio sp. K90mix (TK90_2699–TK90_2703) | |||
| IV-B | Rhodococcus jostii RHA1 (RHA1_ro10069–RHA_ro10072) | ||||
| IV-C | DNA? | Thermoflexia bacterium (D6793_05715–D6793_05700) |
| Class | Type | Subtype | Variant | Native Target | Origin |
|---|---|---|---|---|---|
| 2 | II | II-A | DNA | Streptococcus thermophilus (str0657–str0660) | |
| II-B | DNA | Legionella pneumophila str. Paris (lpp0160–lpp0163) | |||
| II-C | II-C1 | DNA | Neisseria lactamica 020-06 (NLA_17660–NLA_17680) | ||
| II-C2 | DNA | Micrarchaeum acidiphilum ARMAN-1 (BK997_03320–BK997_03335) | |||
| V | V-A | DNA | Francisella cf. Novicida Fx1 (FNFX1_1431–FNFX1_1428) | ||
| V-B | V-B1 | DNA | Alicyclobacillus acidoterrestris (N007_06525–N007_06535) | ||
| V-B2 | DNA | Planctomycetes bacterium RGB_13_46_10 (A2167_01675–A2167_01685) | |||
| V-C | DNA | Oleiphilus sp. (A3715_16885–A3715_16890) | |||
| V-D | DNA | Bacterium CG09_39_24 (BK003_02070–BK003_02075) | |||
| V-E | DNA | Deltaproteobacteria bacterium (A2Z89_08250–A2Z89_08265) | |||
| V-F | V-F1 | DNA | Uncultured archaeon (NDOCEIEL_00008–NDOCEIEL_00011) | ||
| V-F1* | DNA | Bacillus thuringiensis HD-771 (BTG_31928) | |||
| V-F2 | DNA | Uncultured archaeon (ICDLJNLD_00049–ICDLJNLD_00052) | |||
| V-F3 | Candidatus Micrarchaeota archaeon (COU37_03050–COU37_03065) | ||||
| V-U1 | Gordonia otitidis (GOOTI_RS19525) | ||||
| V-U2 | Cyanothece sp. PCC 8801 (PCC8801_4127) | ||||
| V-U4 | Rothia dentrocariosa M567 (HMPREF0734_01291) | ||||
| V-G | RNA | Hot springs metagenome FLYL01000025.1 (182949–185252) | |||
| V-H | Hypersaline lake sediment metagenome (JGI) (Ga0180438_100006283) | ||||
| V-I | DNA | Freshwater metagenome (JGI) (Ga0208225_100001036) | |||
| V-K | Cyanothece sp. PCC 8801 (PCC8801_2993–PCC8801_2997) | ||||
| VI | VI-A | RNA | Leptotrichia shahii (B031_RS0110445) | ||
| VI-B | VI-B1 | RNA | Prevotella buccae (HMPREF6485_RS00335–HMPREF6483_RS00340) | ||
| VI-B2 | RNA | Bergeyella zoohelcum (HMPREF9699_02005–HMPREF9699_02006) | |||
| VI-C | RNA? | Fusobacterium perfoetens (T364_RS0105110) | |||
| VI-D | RNA | Ruminococcus bicirculans (RBI_RS12820) |
| Crops | Target Genes | Editing Process | Results | References |
|---|---|---|---|---|
| Biotic Factors | ||||
| Apple | DIPM1, DIPM2, DIPM4 | Gene knockout | Resistance to fire blight disease | [58] |
| Arabidopsis thaliana | Non-coding/coding region of viral genome, eIF(iso)4E | Gene knockout | Virus resistance | [59] |
| Banana | banana streak virus genes | Gene inactivation | Virus resistant | [60] |
| Cassava | eIF4E isoforms nCBP-1, nCBP-2 | Gene knockout | Partial resistance to CBSD | [61] |
| Cassava | EPSPS | Gene insertion and replacement | Herbicide resistant | [62] |
| Citrus | PthA4, CsLOB1 | Gene knockout | Resistance to citrus canker | [63] |
| Citrus | CsLOB1 | Gene knockout | Resistance to citrus canker | [64] |
| Cocoa | TcNPR3 | Gene knockout | Increased resistance to Phytophthora tropicalis | [65] |
| Cotton | clcud | Gene disruption | Leaf curl disease resistant | [66] |
| Cotton | Gh14-3-3d | Gene knockout | Resistance to Verticillium dahlia | [67] |
| Cucumber | eIF4E | Gene knockout | Broad virus resistant | [68] |
| Flax | EPSPS | Gene insertion and replacement | Herbicide resistant | [69] |
| Grape vine | VvWRKY52 | Gene knockout | Resistance to Botrytis cinerea | [70] |
| Grape vine | MLO7 | Gene knockout | Resistance to powdery mildew | [58] |
| Potato | ALS | Gene insertion and replacement | Herbicide resistant | [71] |
| Potato | Coilin | Gene knockout | Increased resistance to potato virus Y | [72] |
| Rice | OsERF922, OsSEC3A, OsSWEET13 | Gene mutations | Resistant to blast and bacterial blight | [73] |
| Rice | CYP71A1 | Gene knockout | Broad-spectrum resistance to insect pests | [40] |
| Rice | eIF4G | Gene mutation | Resistance to rice tungro spherical virus | [74] |
| Rice | ALS | Gene insertion and replacement | Herbicide resistant | [42] |
| Rice | EPSPS | Gene insertion and replacement | Herbicide resistant | [75] |
| Rice | C287T, ALS | Multiplex genome editing | Herbicide resistance | [55] |
| Rice | UVb1-1 | Multiplex genome editing | Resistance against false smut | [76] |
| Rice | ALS | Base editing | Herbicide resistant | [55] |
| Soybean | ALS | Gene insertion and replacement | Herbicide resistant | [77] |
| Tobacco | AGO2 | Gene knockout | Virus resistance | [78] |
| Tomato | SlMLO1 | Gene deletion | Powdery mildew resistant | [79] |
| Tomato | SlJAZ2 | Gene truncation | Bacterial speck resistant | [80] |
| Watermelon | ALS | Base editing | Herbicide resistant | [57] |
| Wheat | EDR1 | Gene knockout | Resistant to powdery mildew | [81] |
| Wheat | TaMLO-A1, TaMLO-B1, TaMLO-D1 | Gene knockout | Resistant to powdery mildew | [82] |
| Abiotic Factors | ||||
| Arabidopsis thaliana | UGT79B2 and UGT79B3 | Gene knockout | Cold, salt, and drought resistance | [83] |
| Barley | ENGase, HvPM19, BolC.GA4.a | Gene knockout | Grain number increase and dormancy control | [84] |
| False flax (Camelina sativa) | FAD2, CsDGAT1, CsPDAT1 | Multiplex genome editing using CRISPR | Increased oleic acid and decreased PUFA | [34,36] |
| Grape vine | IdnDH | Gene insertion and replacement | Failure of tartaric acid biosynthesis or accumulation | [85] |
| Lotus | SNF, SYMRK, LjLb1, LjLb2 | Gene knockout | Increased nitrogen accumulation | [86] |
| Maize | Wx1 | Gene knockout | Increased amylose | [52] |
| Maize | TMS5 | Gene knockout | Male sterile lines | [52] |
| Maize | ARGOS8 | Gene insertion and replacement | Drought tolerance | [52] |
| Maize | Dek42 | Gene knockout | Regulation of kernel development | [87] |
| Mushroom | PPO | Gene knockout | Browning-resistant mushroom | [88] |
| Opium poppy | 4′OMT2 | Gene knockout | Biomolecule synthesis | [89] |
| Orchids | C3H, C4H, 4CL, CCR, and IRX | Gene knockout | Increased lignocellulose biosynthesis | [90] |
| Orchids | PDS | Gene insertion and replacement | Increasing quality and quantity | [91] |
| Potato | StGBSS | Gene knockout | Amylose synthesis | [92] |
| Potato | StMYB44 | Gene knockout | Phosphate stress response | [93] |
| Potato | ALS | Gene knockout | Chlorsulfuron resistance | [94] |
| Rice | Gn1a, DEP1, GS3 | Gene knockout | Enhanced grain size and number and dense erect panicles | [95] |
| Rice | OsGn1a | Gene knockout | Grain number | [30,31] |
| Rice | OsGS3 | Gene knockout | Grain size | [30,31] |
| Rice | TaGW2, OsGW5, OsGLW2, or TaGASR7 | Gene knockout | Grain weight | [30,31] |
| Rice | OsDEP1, TaDEP1 | Gene knockout | Panicle size | [30,31] |
| Rice | LAZY1 | Gene knockout | Tiller spreading | [30,31] |
| Rice | OsAAP3 | Gene knockout | Increased tiller number | [30,31] |
| Rice | GW2, GW5, and TGW6 | Gene knockout | Gain weight improvement | [96] |
| Rice | OsPDS, OsMPK2, OsBADH2 | Gene knockout, Indels | Edited abiotic stress tolerance | [38] |
| Rice | OsDERF1, OsPMS3, OsEPSPS, OsMSH1, OsMYB5 | Gene knockout | Edited drought tolerance | [97] |
| Rice | OsHAK-1 | Gene knockout | Strongly reduced content of 137Cs+ | [98] |
| Rice | OsPRX2 | Gene knockout | Serious defects in leaves, stomal opening under K+-deficient conditions | [99] |
| Rice | OsAnn3 | Gene knockout | Decreased cold tolerance | [100] |
| Rice | SBEIIb | Gene knockout | Higher amylose content | [37] |
| Rice | TMS5 | Gene knockout | Thermosensitive male sterile lines | [51] |
| Rice | csa | Gene knockout | Photosensitive male sterile lines | [95] |
| Rice | OsMATL | Gene knockout | Haploid seed formation | [54] |
| Rice | ACCase gene | Base editing | Haloxyfop-R-methyl resistant | [101] |
| Rubber | TK 1-FFT | Gene knockout | Rubber biosynthesis | [102] |
| Soybean | GmSPL9a, b, c | Gene insertion and replacement | Improved yield | [103] |
| Tomato | SlAGL6 | Gene knockout | Parthenocarpic fruit production under heat stress conditions | [39] |
| Tomato | SlIAA9 | Gene knockout | Production of parthenocarpic plants | [104] |
| Tomato | RIN | Gene knockout | Improved shelf life | [105] |
| Tomato | AP2a, NOR, TDR4, MBP7 | Gene knockout | Delayed fruit ripening | [106] |
| Tomato | SlAGO7 | Gene knockout | Increased growth | [107] |
| Tomato | SlNPR1 | Gene knockout | Role of SINPR1 in drought resistance | [108] |
| Wheat | GW2 | Base editing | Increased grain and protein content | [32] |
| Wheat | α-gliadin family members | Indels | Decreased gluten | [35] |
| Wheat | PinB | Gene insertion and replacement | Genetic improvement | [109] |
| Wheat | TaWaxy, TaMTL | Gene insertion and replacement | Induction of haploid plants | [110] |
| Wheat | Ms45 | Base editing | Male-sterility | [53] |
| Disease | Target Genes | Editing Process | Results | Reference |
|---|---|---|---|---|
| Cataracts | EGFP, Crygc | Indels | Gene correction of Crygc gene | [166] |
| Chronic granulomatous disease (CGD) | CYBB | Single point mutation | Restoration of ROS activity on phagocytic cells | [167] |
| Coronary heart disease | PCSK9 | Insertion/deletion | Prevent coronary heart disease | [168] |
| Cystic fibrosis (CF) | CFTR | Base editing | Normal recovery of CFTR expression | [157] |
| Diabetes mellitus type 1 (DM1) | DMPK | Gene editing | Play important role in disease etiology | [169] |
| Duchenne muscular dystrophy (DMD) | exon44 | Gene deletion | Restoration of dystrophin protein | [116] |
| Duchenne muscular dystrophy (DMD) | Dmd | Gene deletion | Restoration of dystrophin protein | [170] |
| Hemophilia B andA | F9 and F8 | Gene knock-in and alteration | Control bleeding process | [118] |
| Hearing loss | Tmc1 | Gene disruption | Prevention of deafness | [120] |
| Hematopoietic diseases | BCL11A | Gene deletion | Increased production of γ-hemoglobin | [119] |
| Huntington disease (HD) | HTT | Small targeted deletions | Terminating HTT expression | [171] |
| Sickle cell disease (SCD) | HBB | Indels | RBC formation | [121] |
| Sickle cell disease (SCD) | BCL11A | Gene interference | RBC formation | [172] |
| WPW syndrome | PRKAG2 | Insertion/deletion | Correction of PRKAG2 cardiac syndrome | [173] |
| α1-antitrypsin | hSERPINA1 | Gene disruption | Reduced expression of liver fibrosis markers | [117] |
| β-thalassemia | HBB | Mutation deletion | RBC formation | [174] |
| Cancer Tissue Therapy | ||||
| Ataxia-telangiectasia | ATM | Gene knockout | Increased ATM-independent repair mechanism | [175] |
| Bladder cancer | ATG5 | Gene knockout | Cytotoxicity suppression | [149] |
| Cervical cancer | E6 and E7 | Gene knockout | Anti-tumor activities | [176] |
| Cholangiocarcinoma | BAP1 | Loss of malignancy | [151] | |
| Colorectal cancer | Acvr1b, Acvr2a, and Arid2 | Gene knockout | Suppression of cancer | [150] |
| Human lung adenocarcinoma A549 cells | CD38 | Gene knockout | Inhibited anchorage-independent cell growth | [148] |
| Hypertrophic cardiomyopathy (HCM) | MYBPC3 | Mutation correction | Maintain sarcomere structure and regulate relaxation/contraction | [177] |
| Leukemia and lymphoma | TRAC locus | Knock-in | Enhanced anti-tumor capability | [152] |
| Melanoma | PD-1, LAG3, and CTLA4 | Anti-tumor activities | [155] | |
| Nijmegen breakage syndrome | NBS1 | Mutation correction | Decreased susceptibility of cancer | [178] |
| Non-small cell lung cancer | NPM1 | Gene knockout | Anti-tumor activities | [179] |
| Tumor | B2M | Gene disruption | Anti-tumor activities | [153] |
| Tumor | PD-1 | Gene disruption | Anti-tumor activities | [154] |
| Cell Therapies | ||||
| Lung and esophageal cancer | PD-1 of T cells | Gene knockout | Clinical trials | [180] |
| Viral Infection | ||||
| Chimeric hepatitis B | Repeat regions | Gene inactivation | Eradication of HBV infection | [181] |
| HIV | LTR U3 region | Gene knockout | Loss of LTR expression | [126] |
| HIV | CCR5 on T cells | Gene knockout | Reduced entry of virus | [127] |
| HIV | CXCR4 | Gene knockout | Resistant to X4 HIV virus | [128] |
| HIV | CXCR4 and CCR5 | Gene knockout | Cytotoxicity of cells | [129,130] |
| HIV | Subgenomic particles | Gene knockout | Removal of proviral DNA | [131] |
| HSV-1 | ICP0, ICP4, ICP27 | Abrogation of viral infection | [137] | |
| Human cytomegalovirus | UL122/123 | Decreased IE protein expression, reduced production of new virions | [141] | |
| Human polyomavirus (JCV) | N-terminal of T-antigens | Suppressed viral replication | [143] | |
| Kaposi’s sarcoma-associated herpesvirus | LANA | Decreased episomal load | [140] | |
| Bacterial Infection | ||||
| Burkitt lymphoma Hodgkin’s disease | BART5, BART6 | Gene deletion | Suppressing the viral replication | [139] |
| Escherichia coli | ftsA, asd, msbA, nusB | Gene knockout | Antibiotic resistance | [182] |
| Escherichia coli | NDM-1, CTX-M-15 | Gene truncation | Carbapenem resistance | [183] |
| Escherichia coli | blaTEM, blaSHV | Gene truncation | Re-sensitization to β-lactam resistance | [184] |
| Staphylococcus aureus | aph-3, mecA | Gene deletion | Confer resistance against kanamycin | [185] |
| Target Species | Target Gene(s) | Editing Process | Results | References |
|---|---|---|---|---|
| Bacteria | ||||
| Ashbya gossypii | ADE2 | Nucleotide deletions | Production of folic acid, biolipids, and nucleoside | [196] |
| Bacillus smithii | pyrF, amyE, trpC2 | Gene deletion and insertion/recombination | Capable of using five and six carbon containing sugars | [199,200] |
| Bacillus subtilis | cypX, yvmA | Gene deletion and insertion | Production of several enzymes and low molecular weight substances | [201] |
| Clostridium autoethanogenum | adh, 2, 3-bdh | Gene deletion and insertion | Production biofuels and 2,3-butanediol utilizing CO, CO2, and H2 | [202] |
| Clostridium beijerinckii | Ack, adhE | Gene deletion and insertion | Biofuels and biochemicals production | [203] |
| Clostridium cellulolyticum | pyrF, MspI | Gene deletion and insertion | Model for production of renewable biochemicals | [204] |
| Clostridium ljungdahlii | pta, adhE1, ctf and pyrE | Gene deletion | Production of ethanol from synthesis gas | [205] |
| Clostridium pasteurianum | cpaAIR | Gene deletion | Potential biofuel from conversion of waste glycerol into ethanol | [206] |
| Corynebacterium glutamicum | Pgi, pck | CRISPRi | Increased production of γ-aminobutyric acid | [207] |
| Cyanobacteria | glgC, gltA, ppc | Gene deletion and insertion | Increased production of succinate | [208] |
| Escherichia coli | Growth-related genes | CRISPRi, multiplexed CRISPRi, multiplexed RNA, RNA targeting, MUCICAT | Enhanced model for production of bioproducts and enzymes | [209,210] |
| Lactobacillus reuteri | lacL | Gene deletion and insertion | Model for new single stranded DNA editing | [211] |
| Streptococcus thermophiles | Growth-related genes | Genome editing | Pro-biotic activity and fermentation | [212] |
| Streptomyces albus | redF | Gene deletion | Bioactive products | [213] |
| Streptomyces coelicolor | actII-orf4, glnR, redF | Gene deletion and insertion, CRISPRi | Improvement in genome editing efficiency as a model | [214] |
| Streptomyces rimosus | zwf2, devB | Gene mutation and disruption | Increased yield of oxytetracycline | [215] |
| Streptomyces roseosporus | PKS gene cluster | Gene cluster activation | Production of auroramycin and its aglycon | [216] |
| Streptomyces viridochromogenes | Biosynthetic gene cluster | Gene knock-in | Production of pentangular type II polyketide | [213] |
| Tatumella citrea | tkrA, glk | MUCICAT, allelic exchange | Successful application of the method, multiplex genome editing system | [217,218] |
| Yeast | ||||
| Agaricus bisporus | PPO | Gene disruption | Non-transgenic variety | [88] |
| Alternaria alternata | pksA, brm2 | Gene deletion/insertion | Established pyr4 as selection marker and GFP for protein tagging | [219] |
| Aspergillus aculeatus | alba | Gene mutation | Potential genome editor in filamentous fungi | [220] |
| Aspergillus brasiliensis | pyrG | Gene mutation | Potential genome editor in filamentous fungi | [221] |
| Aspergillus carbonarius | ayg1 | Gene mutation | Potential genome editor in filamentous fungi | [220,222] |
| Aspergillus fumigatus | pksP | Gene cleavage | Potential toolbox for decreasing pathogenicity | [223] |
| Aspergillus luchuensis | brlA, niaD, amyA | Gene mutation | Potential genome editor in filamentous fungi | [220,224] |
| Aspergillus nidulans | yA | Gene mutation | Potential genome editor in filamentous fungi | [220] |
| Aspergillus niger | udh | Gene deletion/insertion | Increased production of galactaric acid | [193] |
| Aspergillus oryzae | Ku-70, ligD | Gene deletion/insertion | Construct plasmid for targeted mutagenesis | [225] |
| Candida albicans | ADE2 | Recombination and multiplexed recombination | Genetic manipulation for increased biproducts | [226] |
| Candida glabrata | a GPI-anchored aspartyl protease, a putative serine/threonine kinase | Gene cleavage | Understanding C. glabrata virulence in vivo | [227] |
| Cryptococcus Neoformans | ADE2, L41, Tsp2 | Gene cleavage | Genome engineering in higher fungi | [228] |
| Fusarium fujikuroi | tHMGR, Cps/Ks | Gene knockout | Improved gibberellic acid production | [229] |
| Ganoderma species | ura3 | Gene cleavage | Codon optimization | [230] |
| Huntiella omanensis | MAT1-2-7 gene | Gene inactivation | Role in sexual reproduction | [231] |
| Kluyveromyces lactis | GAL80, YKU80 | Gene deletion and insertion | Enhanced production strain | [232] |
| Myceliophthora thermophila | cre-1, res-1, gh1-1, and alp-1 | Multigene disruption | Enhanced hypercellulase production | [233] |
| Neurospora crassa | CLR-2, GSY-1 | Gene replacement | Increased expression of cellulase and luciferase | [234] |
| Penicillium chrysogenum | pks17, amdS | Gene cleavage | Potential genome editor in filamentous fungi | [235] |
| Phytophthora sojae | RXLR, Avr4/6 | Gene replacement | Control pathogenicity | [236] |
| Pichia pastoris | AOX1, MPPI | Gene deletion | Production of iso-pentanol | [194] |
| Saccharomyces cerevisiae | Growth-related genes | CRISPRa, CRISPRi, Multiplex metabolic engineering | Increased bioproducts | [189,232] |
| Schizosaccharomyces pombe | rrk1 | Gene cleavage | CRISPR toolbox in S. pombe | [237] |
| Talaromyces atroroseus | PKS | Gene cleavage | Production of polyketide-nonribosomal peptide | [238] |
| Trichoderma reesei | ura5 | Homologous recombination | Tool for genome engineering | [239] |
| Ustilago maydis | bE1, bW2 | Gene disruption | Increased DNA repair system | [240] |
| Yarrowia lipolytica | PEX10, KU70, and MFE1 | Gene disruption | Increased synthesis and storage of lipids | [241] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nidhi, S.; Anand, U.; Oleksak, P.; Tripathi, P.; Lal, J.A.; Thomas, G.; Kuca, K.; Tripathi, V. Novel CRISPR–Cas Systems: An Updated Review of the Current Achievements, Applications, and Future Research Perspectives. Int. J. Mol. Sci. 2021, 22, 3327. https://doi.org/10.3390/ijms22073327
Nidhi S, Anand U, Oleksak P, Tripathi P, Lal JA, Thomas G, Kuca K, Tripathi V. Novel CRISPR–Cas Systems: An Updated Review of the Current Achievements, Applications, and Future Research Perspectives. International Journal of Molecular Sciences. 2021; 22(7):3327. https://doi.org/10.3390/ijms22073327
Chicago/Turabian StyleNidhi, Sweta, Uttpal Anand, Patrik Oleksak, Pooja Tripathi, Jonathan A. Lal, George Thomas, Kamil Kuca, and Vijay Tripathi. 2021. "Novel CRISPR–Cas Systems: An Updated Review of the Current Achievements, Applications, and Future Research Perspectives" International Journal of Molecular Sciences 22, no. 7: 3327. https://doi.org/10.3390/ijms22073327