
TFF1 Induces Aggregation and Reduces Motility of Helicobacter pylori

 , , , ,
, , , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
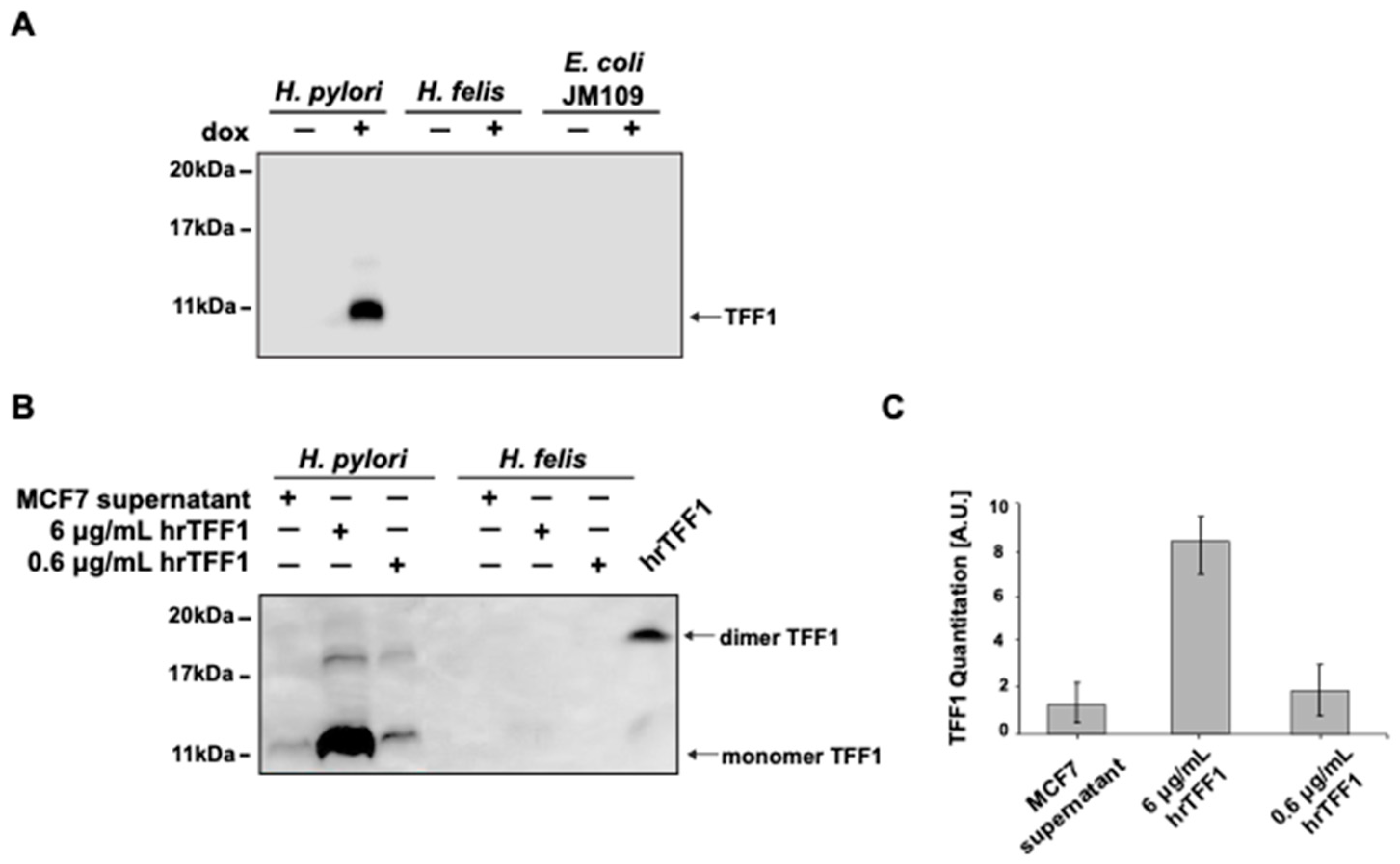
2.1. hTFF1 Binds H. pylori but Not H. felis
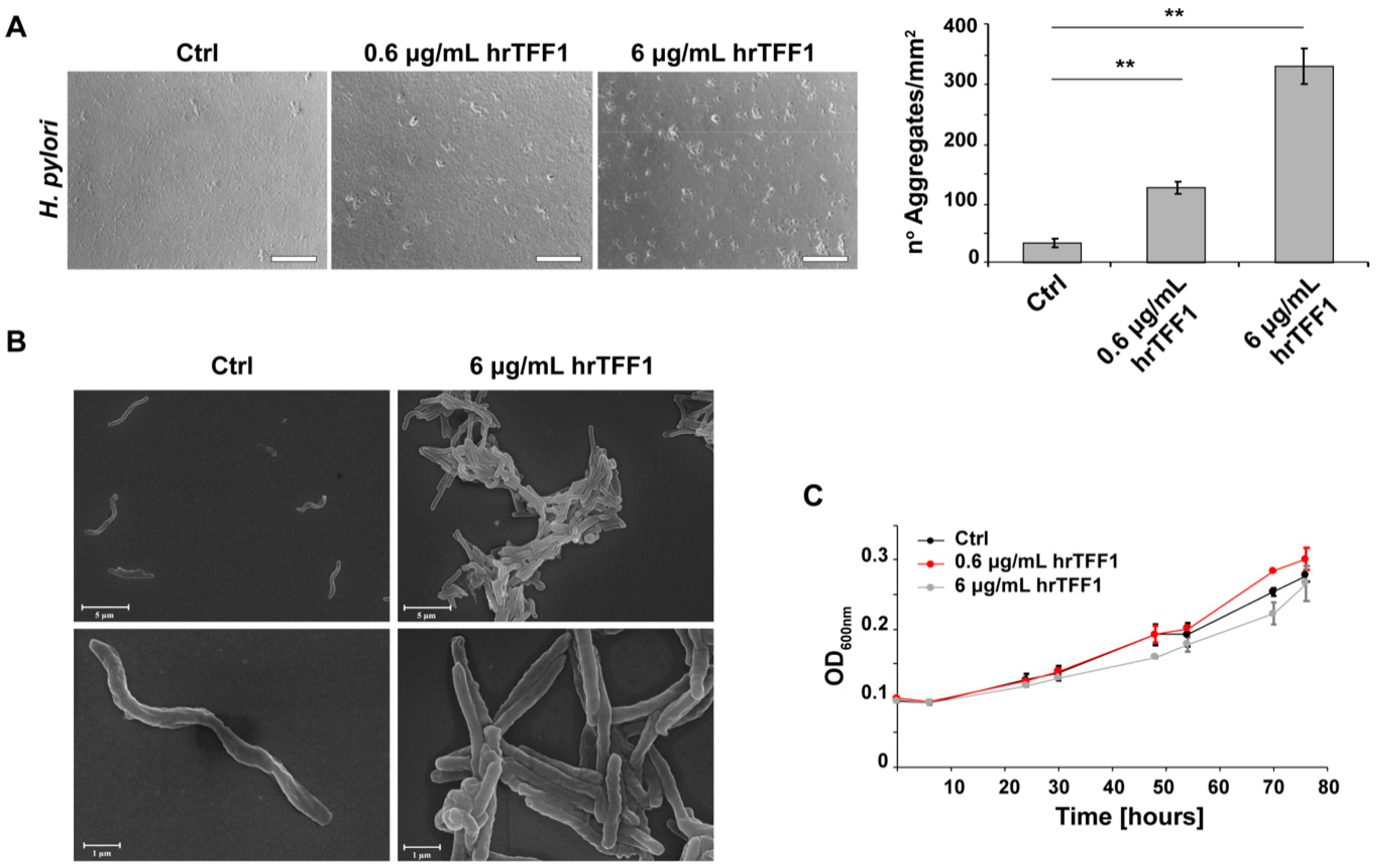
2.2. hrTFF1 Induces H. pylori Aggregation
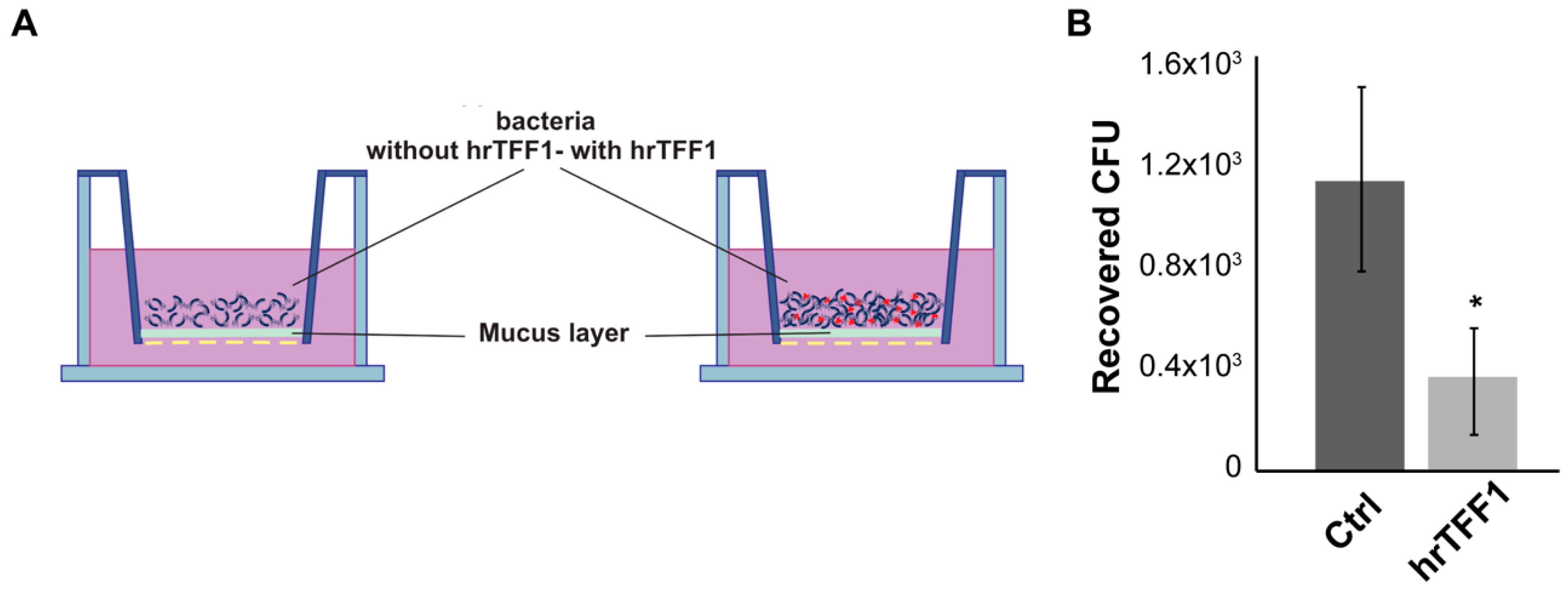
2.3. hrTFF1 Affects H. pylori Motility
2.4. Transcriptional Regulation in hrTFF1-H. pylori Aggregates
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. Bacterial Strains and Culture Conditions
4.3. Human Recombinant TFF1 Production and Purification
4.4. Pull-down Interaction Experiment
4.5. Helicobacter pylori Growth Curve
4.6. Bacterial Aggregation Observation
4.7. Scanning Electron Microscopy Analysis
4.8. Chemotaxis Assay
4.8.1. Analysis of TFF1 Chemiotactic Properties
4.8.2. Analysis of TFF1 Influence on H. pylori Chemotactic Behavior
4.9. Motility Assay
4.10. Real Time-PCR
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018 GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Peek, R.M., Jr.; Blaser, M.J. Helicobacter pylori and gastrointestinal tract adenocarcinomas. Nat. Rev. Cancer 2002, 2, 28–37. [Google Scholar] [CrossRef]
- Graham, D.Y. Helicobacter pylori update: Gastric cancer, reliable therapy, and possible benefits. Gastroenterology 2015, 148, 719–731.e3. [Google Scholar] [CrossRef] [Green Version]
- Algood, H.M.; Cover, T.L. Helicobacter pylori persistence: An overview of interactions between H. pylori and host immune defenses. Clin. Microbiol. Rev. 2006, 19, 597–613. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Meng, W.; Wang, B.; Qiao, L. Helicobacter pylori-induced gastric inflammation and gastric cancer. Cancer Lett. 2014, 345, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Mentis, A.A.; Boziki, M.; Grigoriadis, N.; Papavassiliou, A.G. Helicobacter pylori infection and gastric cancer biology: Tempering a double-edged sword. Cell. Mol. Life Sci. CMLS 2019, 76, 2477–2486. [Google Scholar] [CrossRef] [Green Version]
- Rolig, A.S.; Shanks, J.; Carter, J.E.; Ottemann, K.M. Helicobacter pylori requires TlpD-driven chemotaxis to proliferate in the antrum. Infect. Immun. 2012, 80, 3713–3720. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.E.; Sjovall, H.; Hansson, G.C. The gastrointestinal mucus system in health and disease. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 352–361. [Google Scholar] [CrossRef] [Green Version]
- Boccellato, F.; Woelffling, S.; Aki, I.M.; Sanchez, G.; Goosmann, C.; Schmid, M.; Berger, H.; Morey, P.; Denecke, C.; Ordemann, J.; et al. Polarised epithelial monolayers of the gastric mucosa reveal insights into mucosal homeostasis and defence against infection. Gut 2018, 68, 400–413. [Google Scholar] [CrossRef] [PubMed]
- Van den Brink, G.R.; Tytgat, K.M.; Van der Hulst, R.W.; Van der Loos, C.M.; Einerhand, A.W.; Buller, H.A.; Dekker, J.H. pylori colocalises with MUC5AC in the human stomach. Gut 2000, 46, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Van de Bovenkamp, J.H.; Mahdavi, J.; Korteland-Van Male, A.M.; Buller, H.A.; Einerhand, A.W.; Boren, T.; Dekker, J. The muc5ac glycoprotein is the primary receptor for Helicobacter pylori in the human stomach. Helicobacter 2003, 8, 521–532. [Google Scholar] [CrossRef]
- May, F.E.; Westley, B.R. Trefoil proteins: Their role in normal and malignant cells. J. Pathol. 1997, 183, 4–7. [Google Scholar] [CrossRef]
- Thim, L.; Madsen, F.; Poulsen, S.S. Effect of trefoil factors on the viscoelastic properties of mucus gels. Eur. J. Clin. Investig. 2002, 32, 519–527. [Google Scholar] [CrossRef]
- Heuer, J.; Heuer, F.; Stürmer, R.; Harder, S.; Schlüter, H.; Braga Emidio, N.; Muttenthaler, M.; Jechorek, D.; Meyer, F.; Hoffmann, W. The Tumor Suppressor TFF1 Occurs in Different Forms and Interacts with Multiple Partners in the Human Gastric Mucus Barrier: Indications for Diverse Protective Functions. Int. J. Mol. Sci. 2020, 21, 2508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clyne, M.; Dillon, P.; Daly, S.; O’Kennedy, R.; May, F.E.; Westley, B.R.; Drumm, B. Helicobacter pylori interacts with the human single-domain trefoil protein TFF1. Proc. Natl. Acad. Sci. USA 2004, 101, 7409–7414. [Google Scholar] [CrossRef] [Green Version]
- Montefusco, S.; Esposito, R.; D’Andrea, L.; Monti, M.C.; Dunne, C.; Dolan, B.; Tosco, A.; Marzullo, L.; Clyne, M. Copper promotes TFF1-mediated Helicobacter pylori colonization. PLoS ONE 2013, 8, e79455. [Google Scholar] [CrossRef]
- Tosco, A.; Monti, M.C.; Fontanella, B.; Montefusco, S.; D’Andrea, L.; Ziaco, B.; Baldantoni, D.; Rio, M.C.; Marzullo, L. Copper binds the carboxy-terminus of trefoil protein 1 (TFF1), favoring its homodimerization and motogenic activity. Cell. Mol. Life Sci. 2010, 67, 1943–1955. [Google Scholar] [CrossRef] [PubMed]
- Reeves, E.P.; Ali, T.; Leonard, P.; Hearty, S.; O’Kennedy, R.; May, F.E.; Westley, B.R.; Josenhans, C.; Rust, M.; Suerbaum, S.; et al. Helicobacter pylori lipopolysaccharide interacts with TFF1 in a pH-dependent manner. Gastroenterology 2008, 135, 2043–2054. [Google Scholar] [CrossRef]
- Dunne, C.; Naughton, J.; Duggan, G.; Loughrey, C.; Kilcoyne, M.; Joshi, L.; Carrington, S.; Earley, H.; Backert, S.; Robbe Masselot, C.; et al. Binding of Helicobacter pylori to Human Gastric Mucins Correlates with Binding of TFF1. Microorganisms 2018, 6, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomita, H.; Takaishi, S.; Menheniott, T.R.; Yang, X.; Shibata, W.; Jin, G.; Betz, K.S.; Kawakami, K.; Minamoto, T.; Tomasetto, C.; et al. Inhibition of gastric carcinogenesis by the hormone gastrin is mediated by suppression of TFF1 epigenetic silencing. Gastroenterology 2011, 140, 879–891. [Google Scholar] [CrossRef] [Green Version]
- Van De Bovenkamp, J.H.; Korteland-Van Male, A.M.; Büller, H.A.; Einerhand, A.W.; Dekker, J. Infection with Helicobacter pylori affects all major secretory cell populations in the human antrum. Dig. Dis. Sci. 2005, 50, 1078–1086. [Google Scholar] [CrossRef] [PubMed]
- Esposito, R.; Morello, S.; Vllahu, M.; Eletto, D.; Porta, A.; Tosco, A. Gastric TFF1 Expression from Acute to Chronic Helicobacter infection. Front. Cell. Infect. Microbiol. 2017, 7–434. [Google Scholar] [CrossRef]
- Matsuda, K.; Yamauchi, K.; Matsumoto, T.; Sano, K.; Yamaoka, Y.; Ota, H. Quantitative analysis of the effect of Helicobacter pylori on the expressions of SOX2, CDX2, MUC2, MUC5AC, MUC6, TFF1, TFF2, and TFF3 mRNAs in human gastric carcinoma cells. Scand. J. Gastroenterol. 2008, 43, 25–33. [Google Scholar] [CrossRef]
- Soutto, M.; Chen, Z.; Katsha, A.M.; Romero-Gallo, J.; Krishna, U.S.; Piazuelo, M.B.; Washington, M.K.; Peek, R.M., Jr.; Belkhiri, A.; El-Rifai, W.M. Trefoil factor 1 expression suppresses Helicobacter pylori-induced inflammation in gastric carcinogenesis. Cancer 2015, 121, 4348–4358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soutto, M.; Romero-Gallo, J.; Krishna, U.; Piazuelo, M.B.; Washington, M.K.; Belkhiri, A.; Peek, R.M., Jr.; El-Rifai, W. Loss of TFF1 promotes Helicobacter pylori-induced b-catenin activation and gastric tumorigenesis. Oncotarget 2015, 6, 17911–17922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchbank, T.; Westley, B.R.; May, F.E.; Calnan, D.P.; Playford, R.J. Dimerization of human pS2 (TFF1) plays a key role in its protective/healing effects. J. Pathol. 1998, 185, 153–158. [Google Scholar] [CrossRef]
- Tosco, A.; Monti, M.C.; Fontanella, B.; Rio, M.C.; Gomez-Paloma, L.; Leone, A.; Marzullo, L. Copper-binding activity of Trefoil factor 1 (TFF1): A new perspective in the study of the multifunctional roles of TFFs. Peptides 2007, 28, 1461–1469. [Google Scholar] [CrossRef]
- Cerda, O.A.; Núñez-Villena, F.; Soto, S.E.; Ugalde, J.M.; López-Solís, R.; Toledo, H. tlpA gene expression is required for arginine and bicarbonate chemotaxis in Helicobacter pylori. Biol. Res. 2011, 44, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Dolan, B.; Naughton, J.; Tegtmeyer, N.; May, F.E.; Clyne, M. The interaction of Helicobacter pylori with the adherent mucus gel layer secreted by polarized HT29-MTX-E12 cells. PLoS ONE 2012, 7, e47300. [Google Scholar] [CrossRef] [Green Version]
- Moodley, Y.; Linz, B.; Bond, R.P.; Nieuwoudt, M.; Soodyall, H.; Schlebusch, C.M.; Bernhöft, S.; Hale, J.; Suerbaum, S.; Mugisha, L.; et al. Age of the association between Helicobacter pylori and man. PLoS Pathog. 2012, 8, e1002693. [Google Scholar] [CrossRef] [Green Version]
- Clyne, M.; May, F.E.B. The Interaction of Helicobacter pylori with TFF1 and Its Role in Mediating the Tropism of the Bacteria within the Stomach. Int. J. Mol. Sci. 2019, 20, 4400. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yang, T.; Liao, T.; Debowski, A.W.; Nilsson, H.-O.; Fulurija, A.; Haslam, S.M.; Mulloy, B.; Dell, A.; Stubbs, K.A.; et al. The redefinition of Helicobacter pylori lipopolysaccharide O-antigen and core- oligosaccharide domains. PLoS Pathog. 2017, 13, e1006280. [Google Scholar] [CrossRef] [PubMed]
- Hynes, S.; Ferris, J.A.; Szponar, B.; Wadström, T.; Fox, J.G.; O’Rourke, J.; Larsson, L.; Yaquian, E.; Ljungh, A.; Clyne, M.; et al. Comparative chemical and biological characterization of the lipopolysaccharides of gastric and enterohepatic helicobacters. Helicobacter 2004, 9, 313–323. [Google Scholar] [CrossRef] [Green Version]
- Skoog, E.; Padra, M.; Åberg, A.; Gideonsson, P.; Obi, I.; Quintana-Hayashi, M.P.; Arnqvist, A.; Lindén, S.K. BabA dependent binding of Helicobacter pylori to human gastric mucins cause aggregation that inhibits proliferation and is regulated via ArsS. Sci. Rep. 2017, 7, 40656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, T.; Saito, R.; Oyama, M.; Takeuchi, T.; Tanaka, T.; Natsume, H.; Tamura, M.; Arata, Y.; Hatanaka, T. Galectin-2 Has Bactericidal Effects against Helicobacter pylori in a β-galactoside-Dependent Manner. Int. J. Mol. Sci. 2020, 21, 2697. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Padra, M.; Constantino, M.A.; Sharba, S.; Thorell, A.; Lindén, S.K.; Bansil, R. Influence of the viscosity of healthy and diseased human mucins on the motility of Helicobacter pylori. Sci Rep. 2018, 8, 9710. [Google Scholar] [CrossRef]
- Ottemann, K.M.; Lowenthal, A.C. Helicobacter pylori uses motility for initial colonization and to attain robust infection. Infect Immun. 2002, 70, 1984–1990. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, P.W.; Kostrzynska, M.; Trust, T.J. Non-motile mutants of Helicobacter pylori and Helicobacter mustelae defective in flagellar hook production. Mol. Microbiol. 1994, 14, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Josenhans, C.; Labigne, A.; Suerbaum, S. Comparative ultrastructural and functional studies of Helicobacter pylori and Helicobacter mustelae flagellin mutants: Both flagellin subunits, FlaA and FlaB, are necessary for full motility in Helicobacter species. J. Bacteriol. 1995, 177, 3010–3020. [Google Scholar] [CrossRef] [Green Version]
- Romano, E.; Vllahu, M.; Bizzarro, V.; Belvedere, R.; Esposito, R.; Petrella, A.; Tosco, A. TFF1 Promotes EMT-Like Changes through an Auto-Induction Mechanism. Int. J. Mol. Sci. 2018, 19, 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazumder, R.; Phelps, T.J.; Krieg, N.R.; Benoit, R.E. Determining chemotactic responses by two subsurface microaerophiles using a simplified capillary assay method. J. Microbiol. Methods 1999, 37, 255–263. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eletto, D.; Vllahu, M.; Mentucci, F.; Del Gaudio, P.; Petrella, A.; Porta, A.; Tosco, A. TFF1 Induces Aggregation and Reduces Motility of Helicobacter pylori. Int. J. Mol. Sci. 2021, 22, 1851. https://doi.org/10.3390/ijms22041851
Eletto D, Vllahu M, Mentucci F, Del Gaudio P, Petrella A, Porta A, Tosco A. TFF1 Induces Aggregation and Reduces Motility of Helicobacter pylori. International Journal of Molecular Sciences. 2021; 22(4):1851. https://doi.org/10.3390/ijms22041851
Chicago/Turabian StyleEletto, Daniela, Megi Vllahu, Fatima Mentucci, Pasquale Del Gaudio, Antonello Petrella, Amalia Porta, and Alessandra Tosco. 2021. "TFF1 Induces Aggregation and Reduces Motility of Helicobacter pylori" International Journal of Molecular Sciences 22, no. 4: 1851. https://doi.org/10.3390/ijms22041851