
Interplay between Humoral and CLA+ T Cell Response against Candida albicans in Psoriasis

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Description of Patients and Controls Samples Used in Different Experiments
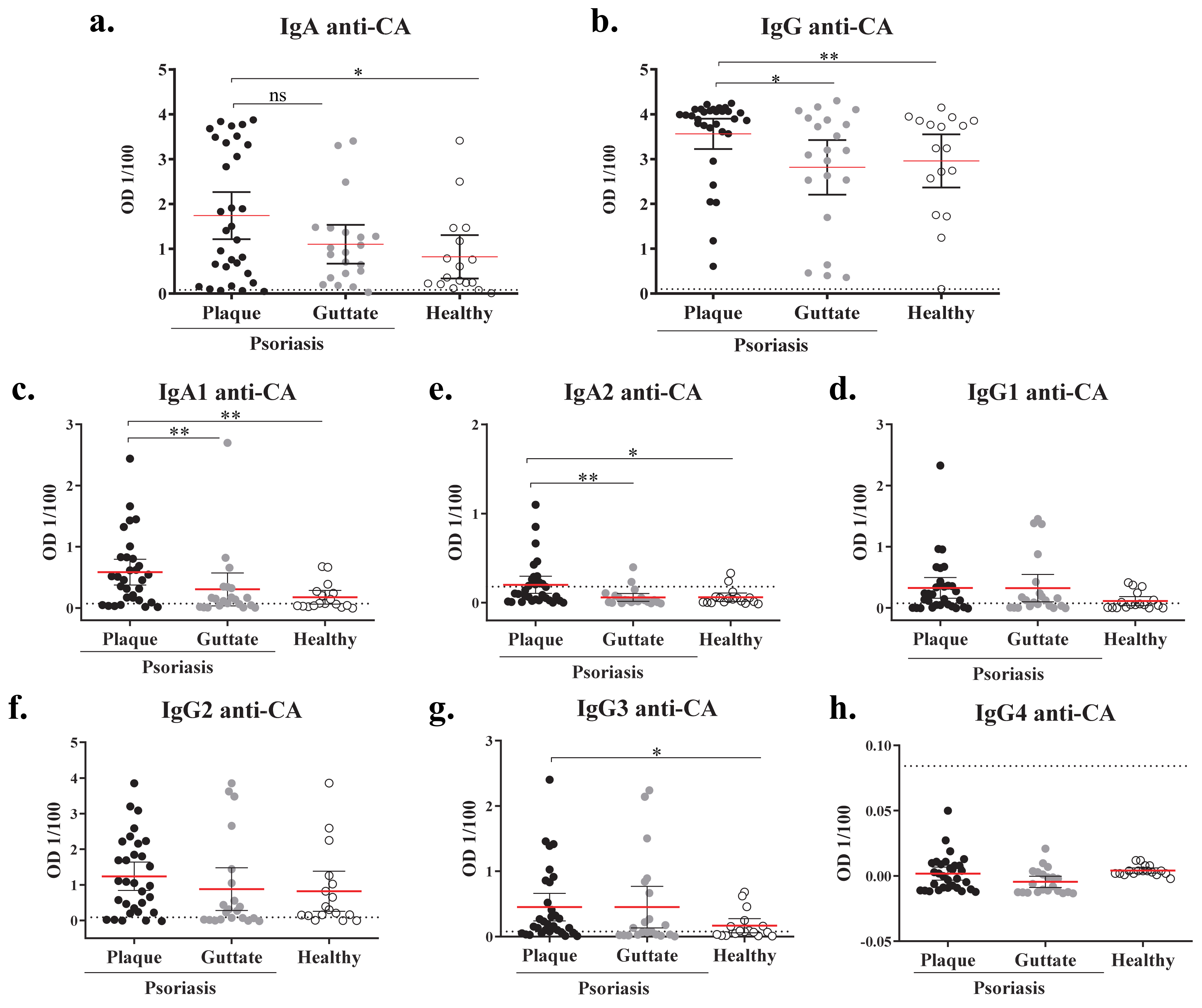
2.2. Plaque Psoriasis Patients Have Increased Levels Candida Albicans-Specific IgA and IgG
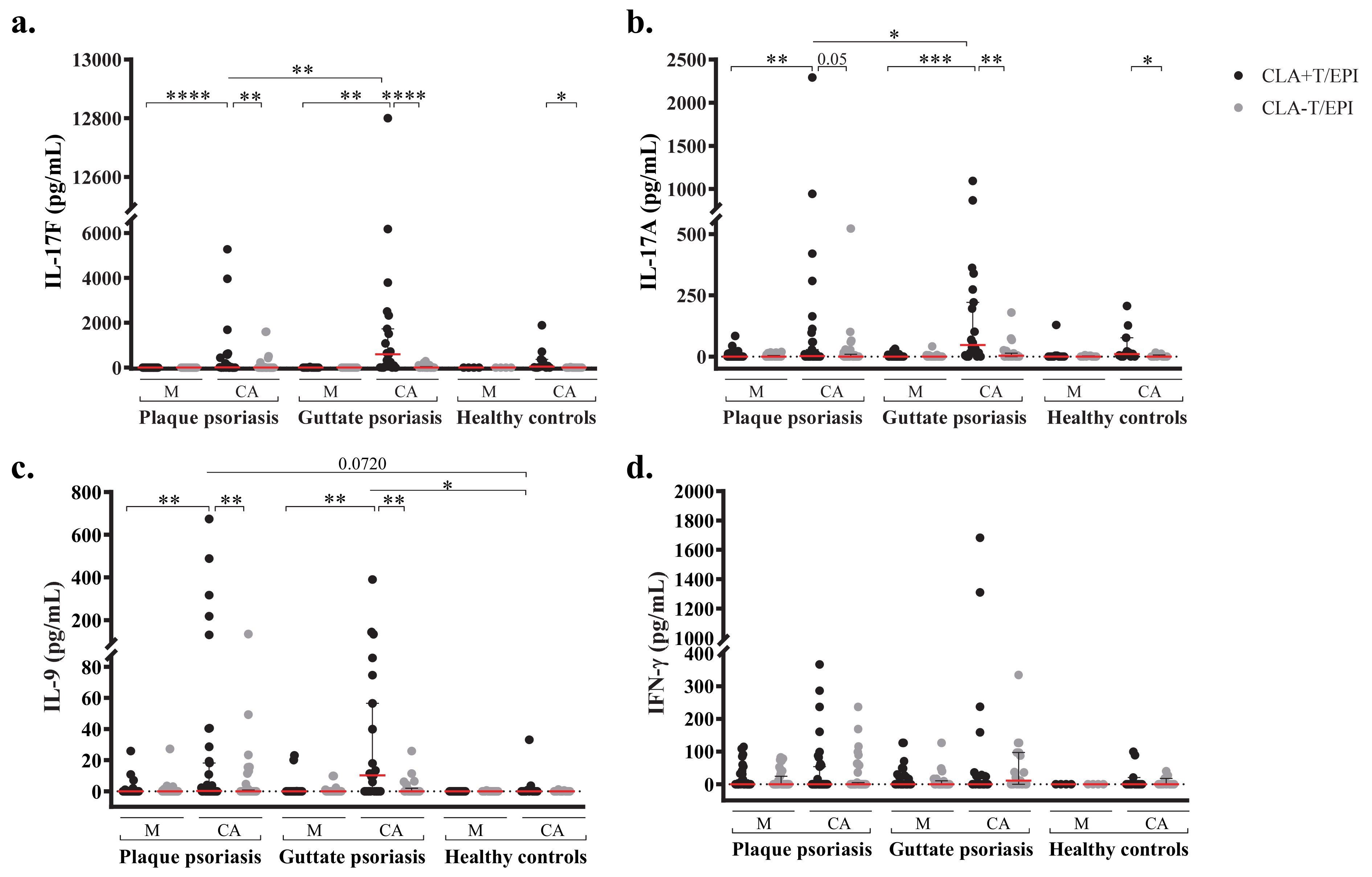
2.3. Candida Albicans-Induced Th17 and Th9 Responses Are Confined to CLA+ T-cells and Dominated by IL-17F
2.4. Candida Albicans’ Specific IgA Plasma Levels and IL-17 T cell Responses Are Directly Associated in Plaque Psoriasis Patients
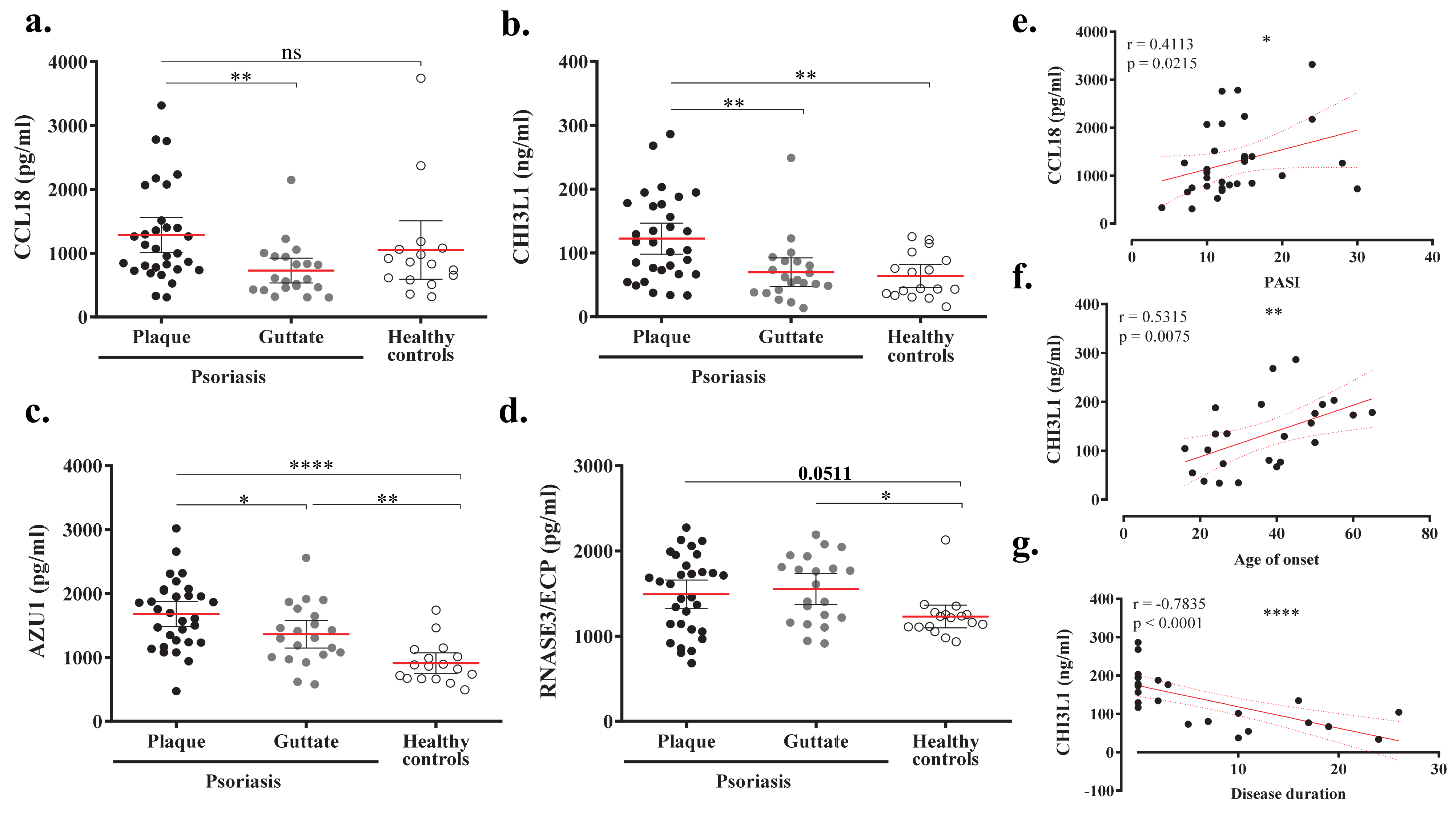
2.5. Proteomic Profile of Plasma from Psoriasis Patients according to Anti-CA IgA Levels
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. ELISA
4.3. Circulating Memory T-cell and Epidermal Cell Isolation
4.4. Co-Cultures
4.5. Cytokine Quantification
4.6. Proteomic Study
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hawkes, J.E.; Chan, T.C.; Krueger, J.G. Psoriasis pathogenesis and the development of novel targeted immune therapies. J. Allergy Clin. Immunol. 2017, 140, 645–653. [Google Scholar] [CrossRef] [Green Version]
- Fry, L.; Baker, B.S. Triggering psoriasis: The role of infections and medications. Clin. Dermatol. 2007, 25, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Waldman, A.; Gilhar, A.; Duek, L.; Berdicevsky, I. Incidence of Candida in psoriasis—A study on the fungal flora of psoriatic patients. Mycoses 2001, 44, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Leibovici, V.; Alkalay, R.; Hershko, K.; Ingber, A.; Westerman, M.; Leviatan-Strauss, N.; Hochberg, M. Prevalence of Candida on the tongue and intertriginous areas of psoriatic and atopic dermatitis patients. Mycoses 2008, 51, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Picciani, B.L.S.; Michalski-Santos, B.; Carneiro, S.; Sampaio, A.L.; Avelleira, J.C.R.; Azulay, D.R.; Pinto, J.M.N.; Dias, E.P. Oral candidiasis in patients with psoriasis: Correlation of oral examination and cytopathological evaluation with psoriasis disease severity and treatment. J. Am. Acad. Dermatol. 2013, 68, 986–991. [Google Scholar] [CrossRef] [PubMed]
- Taheri Sarvtin, M.; Shokohi, T.; Hajheydari, Z.; Yazdani, J.; Hedayati, M.T. Evaluation of candidal colonization and specific humoral responses against Candida albicans in patients with psoriasis. Int. J. Dermatol. 2014, 53, e555–e560. [Google Scholar] [CrossRef] [PubMed]
- Lesan, S.; Toosi, R.; Aliakbarzadeh, R.; Daneshpazhooh, M.; Mahmoudi, L.; Tavakolpour, S.; Mahmoudi, H. Oral Candida colonization and plaque type psoriasis: Is there any relationship? J. Investig. Clin. Dent. 2018, 9, e12335. [Google Scholar] [CrossRef]
- Buslau, M.; Menzel, I.; Holzmann, H. Fungal flora of human faeces in psoriasis and atopic dermatitis. Mycoses 1990, 33, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Bedair, A.A.; Darwazeh, A.M.G.; Al-Aboosi, M.M. Oral Candida colonization and candidiasis in patients with psoriasis. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2012, 114, 610–615. [Google Scholar] [CrossRef]
- Soyuer, U.; Kilic, H.; Alpan, O. Anti-Candida antibody levels in psoriasis vulgaris. Cent. Afr. J. Med. 1990, 36, 190–192. [Google Scholar]
- Flytström, I.; Bergbrant, I.M.; Bråred, J.; Brandberg, L.L. Microorganisms in intertriginous psoriasis: No evidence of Candida. Acta Derm. Venereol. 2003, 83, 121–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jagielski, T.; Rup, E.; Ziółkowska, A.; Roeske, K.; Macura, A.B.; Bielecki, J. Distribution of Malassezia species on the skin of patients with atopic dermatitis, psoriasis, and healthy volunteers assessed by conventional and molecular identification methods. BMC Dermatol. 2014, 14, 3. [Google Scholar] [CrossRef]
- Takemoto, A.; Cho, O.; Morohoshi, Y.; Sugita, T.; Muto, M. Molecular characterization of the skin fungal microbiome in patients with psoriasis. J. Dermatol. 2015, 42, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Paulino, L.C.; Tseng, C.H.; Strober, B.E.; Blaser, M.J. Molecular analysis of fungal microbiota in samples from healthy human skin and psoriatic lesions. J. Clin. Microbiol. 2006, 44, 2933–2941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulino, L.C.; Tseng, C.H.; Blaser, M.J. Analysis of Malassezia microbiota in healthy superficial human skin and in psoriatic lesions by multiplex real-time PCR. FEMS Yeast Res. 2008, 8, 460–471. [Google Scholar] [CrossRef] [Green Version]
- Koike, Y.; Kuwatsuka, S.; Nishimoto, K.; Motooka, D.; Murota, H. Skin mycobiome of psoriasis patients is retained during treatment with TNF and IL-17 inhibitors. Int. J. Mol. Sci. 2020, 21, 3892. [Google Scholar] [CrossRef]
- Salem, I.; Schrom, K.P.; Chu, S.; Retuerto, M.; Richardson, B.; Margvicius, S.; Cameron, M.; Ghannoum, M.; McCormick, T.; Cooper, K. 362 Psoriatic fungal and bacterial microbiomes identify patient endotypes. J. Invest. Dermatol. 2020, 140, S45. [Google Scholar] [CrossRef]
- Lewis, D.J.; Chan, W.H.; Hinojosa, T.; Hsu, S.; Feldman, S.R. Mechanisms of microbial pathogenesis and the role of the skin microbiome in psoriasis: A review. Clin. Dermatol. 2019, 37, 160–166. [Google Scholar] [CrossRef]
- Pakkanen, S.H.; Kantele, J.M.; Moldoveanu, Z.; Hedges, S.; Häkkinen, M.; Mestecky, J.; Kantele, A. Expression of homing receptors on IgA1 and IgA2 plasmablasts in blood reflects differential distribution of IgA1 and IgA2 in various body fluids. Clin. Vaccine Immunol. 2010, 17, 393–401. [Google Scholar] [CrossRef] [Green Version]
- Squiquera, L.; Galimberti, R.; Morelli, L.; Plotkin, L.; Milicich, R.; Kowalckzuk, A.; Leoni, J. Antibodies to proteins from Pityrosporum ovale in the sera from patients with psoriasis. Clin. Exp. Dermatol. 1994, 19, 289–293. [Google Scholar] [CrossRef]
- Liang, Y.S.; Wen, H.Q.; Xiao, R. Serum levels of antibodies for IgG, IgA, and IgM against the fungi antigen in psoriasis vulgaris. Bull. Hunan Med. Univ. 2003, 28, 638–640. [Google Scholar]
- Sparber, F.; Leibundgut-Landmann, S. Interleukin-17 in antifungal immunity. Pathogens 2019, 8, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, B.S.; Powles, A.V.; Malkani, A.K.; Lewis, H.; Valdimarsson, H.; Fry, L. Altered cell-mediated immunity to group A haemolytic streptococcal antigens in chronic plaque psoriasis. Br. J. Dermatol. 1991, 125, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.S.; Powles, A.; Garioch, J.J.; Hardman, C.; Fry, L. Differential T-cell reactivity to the round and oval forms of Pityrosporum in the skin of patients with psoriasis. Br. J. Dermatol. 1997, 136, 319–325. [Google Scholar] [CrossRef]
- Kanda, N.; Tani, K.; Enomoto, U.; Nakai, K.; Watanabe, S. The skin fungus-induced Th1- and Th2-related cytokine, chemokine and prostaglandin E2 production in peripheral blood mononuclear cells from patients with atopic dermatitis and psoriasis vulgaris. Clin. Exp. Allergy 2002, 32, 1243–1250. [Google Scholar] [CrossRef]
- Schlapbach, C.; Gehad, A.; Yang, C.; Watanabe, R.; Guenova, E.; Teague, J.E.; Campbell, L.; Yawalkar, N.; Kupper, T.S.; Clark, R.A. Human TH9 cells are skin-tropic and have autocrine and paracrine proinflammatory capacity. Sci. Transl. Med. 2014, 6, 219ra8. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Romeu, E.; Ferran, M.; de Jesús-Gil, C.; García, P.; Sagristà, M.; Casanova, J.M.; Fernández, J.M.; Chiriac, A.; Hóllo, P.; Celada, A.; et al. Microbe-Dependent Induction of IL-9 by CLA+T Cells in Psoriasis and Relationship with IL-17A. J. Invest. Dermatol. 2018, 138, 580–587. [Google Scholar] [CrossRef] [Green Version]
- De Jesús-Gil, C.; San Nicolás, L.S.; Ruiz-Romeu, E.; Ferran, M.; Soria-Martinez, L.; Chiriac, A.; Celada, A.; Pujol, R.M.; Santamaria-Babí, L.F. Specific IgA and CLA+ T-cell IL-17 response to Streptococcus pyogenes in psoriasis. J. Invest. Dermatol. 2020, 140, 1364–1370. [Google Scholar] [CrossRef]
- Rademaker, M.; Agnew, K.; Anagnostou, N.; Andrews, M.; Armour, K.; Baker, C.; Foley, P.; Gebauer, K.; Gupta, M.; Marshman, G.; et al. Psoriasis and infection. A clinical practice narrative. Australas. J. Dermatol. 2019, 60, 91–98. [Google Scholar] [CrossRef]
- Thomas, J.; Küpper, M.; Batra, R.; Jargosch, M.; Atenhan, A.; Baghin, V.; Krause, L.; Lauffer, F.; Biedermann, T.; Theis, F.J.; et al. Is the humoral immunity dispensable for the pathogenesis of psoriasis? J. Eur. Acad. Dermatology Venereol. 2019, 33, 115–122. [Google Scholar] [CrossRef]
- Tarkowski, A.; Moldoveanu, Z.; Koopman, W.J.; Radl, J.; Haaijman, J.J.; Mestecky, J. Cellular origins of human polymeric and monomeric IgA: Enumeration of single cells secreting polymeric IgA1 and IgA2 in peripheral blood, bone marrow, spleen, gingiva and synovial tissue. Clin. Exp. Immunol. 1991, 85, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Vossenkämper, A.; Blair, P.A.; Safinia, N.; Fraser, L.D.; Das, L.; Sanders, T.J.; Stagg, A.J.; Sanderson, J.D.; Taylor, K.; Chang, F.; et al. A role for gut-associated lymphoid tissue in shaping the human b cell repertoire. J. Exp. Med. 2013, 210, 1665–1674. [Google Scholar] [CrossRef] [Green Version]
- Pietrzak, A.; Grywalska, E.; Socha, M.; Roli, J.; Franciszkiewicz-pietrzak, K.; Rudnicka, L.; Rudzki, M.; Krasowska, D. Prevalence and Possible Role of Candida Species in Patients with Psoriasis: A Systematic Review and Meta-Analysis. Mediators Inflamm. 2018, 2018, 9602362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferran, M.; Romeu, E.R.; Rincón, C.; Sagristà, M.; Giménez Arnau, A.M.; Celada, A.; Pujol, R.M.; Holló, P.; Jókai, H.; Santamaria-Babí, L.F. Circulating CLA+ T lymphocytes as peripheral cell biomarkers in T-cell-mediated skin diseases. Exp. Dermatol. 2013, 22, 439–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurabielle, C.; Link, V.M.; Bouladoux, N.; Han, S.; Dean, E. Immunity to commensal skin fungi promotes psoriasiform skin inflammation. Proc. Natl. Acad. Sci. USA 2020, 117, 16465–16474. [Google Scholar] [CrossRef]
- Nakae, S.; Komiyama, Y.; Nambu, A.; Sudo, K.; Iwase, M.; Homma, I.; Sekikawa, K.; Asano, M.; Iwakura, Y. Antigen-specific T cell sensitization is impaired in Il-17-deficient mice, causing suppression of allergic cellular and humoral responses. Immunity 2002, 17, 375–387. [Google Scholar] [CrossRef] [Green Version]
- Nakae, S.; Nambu, A.; Sudo, K.; Iwakura, Y. Suppression of Immune Induction of Collagen-Induced Arthritis in IL-17-Deficient Mice. J. Immunol. 2003, 171, 6173–6177. [Google Scholar] [CrossRef] [Green Version]
- Shibui, A.; Shimura, E.; Nambu, A.; Yamaguchi, S.; Leonard, W.J.; Okumura, K.; Sugano, S.; Sudo, K.; Nakae, S. Th17 cell-derived IL-17 is dispensable for B cell antibody production. Cytokine 2012, 59, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, E.; Ponzoni, M.; Doglioni, C.; Pistoia, V. IL-17 superfamily cytokines modulate normal germinal center B cell migration. J. Leukoc. Biol. 2016, 100, 913–918. [Google Scholar] [CrossRef]
- Wilson, R.P.; McGettigan, S.E.; Dang, V.D.; Kumar, A.; Cancro, M.P.; Nikbakht, N.; Stohl, W.; Debes, G.F. IgM Plasma Cells Reside in Healthy Skin and Accumulate with Chronic Inflammation. J. Invest. Dermatol. 2019, 139, 2477–2487. [Google Scholar] [CrossRef]
- Debes, G.F.; McGettigan, S.E. Skin-Associated B Cells in Health and Inflammation. J. Immunol. 2019, 202, 1659–1666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar, V.A.; Arranz-Trullén, J.; Prats-Ejarque, G.; Torrent, M.; Andreu, D.; Pulido, D.; Boix, E. Insight into the antifungal mechanism of action of human RNase N-terminus derived peptides. Int. J. Mol. Sci. 2019, 20, 4558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertelsen, T.; Ljungberg, C.; Boye Kjellerup, R.; Iversen, L.; Johansen, C. IL-17F regulates psoriasis-associated genes through IκBζ. Exp. Dermatol. 2017, 26, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Bertelsen, T.; Iversen, L.; Johansen, C. The human IL-17A/F heterodimer regulates psoriasis-associated genes through IκBζ. Exp. Dermatol. 2018, 27, 1048–1052. [Google Scholar] [CrossRef] [PubMed]
- Gao, N.; Yu, F.S.X. Chitinase 3-like 1 promotes candida albicans killing and preserves corneal structure and function by controlling host antifungal responses. Infect. Immun. 2015, 83, 4154–4164. [Google Scholar] [CrossRef] [Green Version]
- Cederlund, A.; Agerberth, B.; Bergman, P. Specificity in killing pathogens is mediated by distinct repertoires of human neutrophil peptides. J. Innate Immun. 2010, 2, 508–521. [Google Scholar] [CrossRef]
- Nalmpantis, D.; Gatou, A.; Fragkioudakis, I.; Margariti, A.; Skoura, L.; Sakellari, D. Azurocidin in gingival crevicular fluid as a potential biomarker of chronic periodontitis. J. Periodontal Res. 2020, 55, 209–214. [Google Scholar] [CrossRef]
- Günther, C.; Bello-Fernandez, C.; Kopp, T.; Kund, J.; Carballido-Perrig, N.; Hinteregger, S.; Fassl, S.; Schwärzler, C.; Lametschwandtner, G.; Stingl, G.; et al. CCL18 Is Expressed in Atopic Dermatitis and Mediates Skin Homing of Human Memory T Cells. J. Immunol. 2005, 174, 1723–1728. [Google Scholar] [CrossRef] [Green Version]
- Fujita, H.; Shemer, A.; Suárez-Fariñas, M.; Johnson-Huang, L.M.; Tintle, S.; Cardinale, I.; Fuentes-Duculan, J.; Novitskaya, I.; Carucci, J.A.; Krueger, J.G.; et al. Lesional dendritic cells in patients with chronic atopic dermatitis and psoriasis exhibit parallel ability to activate T-cell subsets. J. Allergy Clin. Immunol. 2011, 128, 574–582.e1-12. [Google Scholar] [CrossRef]
- Kim, H.O.; Cho, S.I.; Chung, B.Y.; Ahn, H.K.; Park, C.W.; Lee, C.H. Expression of CCL1 and CCL18 in atopic dermatitis and psoriasis. Clin. Exp. Dermatol. 2012, 37, 521–526. [Google Scholar] [CrossRef]
- Davanian, H.; Stranneheim, H.; Båge, T.; Lagervall, M.; Jansson, L.; Lundeberg, J.; Yucel-Lindberg, T. Gene Expression Profiles in Paired Gingival Biopsies from Periodontitis-Affected and Healthy Tissues Revealed by Massively Parallel Sequencing. PLoS ONE 2012, 7, e46440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De-La-Torre, J.; Quindós, G.; Marcos-Arias, C.; Marichalar-Mendia, X.; Gainza, M.L.; Eraso, E.; Acha-Sagredo, A.; Aguirre-Urizar, J.M. Oral Candida colonization in patients with chronic periodontitis. Is there any relationship? Rev. Iberoam. Micol. 2018, 35, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Vieira Colombo, A.P.; Magalhães, C.B.; Hartenbach, F.A.R.R.; Martins do Souto, R.; Maciel da Silva-Boghossian, C. Periodontal-disease-associated biofilm: A reservoir for pathogens of medical importance. Microb. Pathog. 2015, 94, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gu, H.; Xie, S.; Su, Y. Periodontitis in patients with psoriasis: A systematic review and meta-analysis. Oral Dis. 2020. [Google Scholar] [CrossRef]
- Scheffold, A.; Bacher, P.; LeibundGut-Landmann, S. T cell immunity to commensal fungi. Curr. Opin. Microbiol. 2020, 58, 116–123. [Google Scholar] [CrossRef]
- Santamaria Babi, L.F.; Perez Soler, M.T.; Hauser, C.; Blaser, K. Skin-homing T cells in human cutaneous allergic inflammation. Immunol. Res. 1995, 14, 317–324. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Plaque | Guttate | p Value | ||
|---|---|---|---|---|
| Number of patients | 31 | 21 | NA | |
| Age (mean ± SD) | 46.65 (11.97) | 32.65 (10.92) | *** | |
| PASI (mean ± SD) | 13.74 (5.99) | 7.23 (2.91) | *** | |
| HLA-Cw*6 % (n) | Positive | 32.26 (10) | 85.71 (18) | NA |
| Negative | 61.29 (19) | 14.29 (3) | NA | |
| UK | 6.45 (2) | - | NA | |
| Anti-CA IgA | Anti-CA IgG | |||||||
|---|---|---|---|---|---|---|---|---|
| Coculture Condition | Cytokine | Spearman ρ | p Value | Spearman ρ | p Value | |||
| Plaque psoriasis (n = 31) | CLA+T/EPI | IL-17F | 0.3735 | 0.0385 | * | −0.0842 | 0.6525 | ns |
| IL-17A | 0.4798 | 0.0063 | ** | 0.0349 | 0.8521 | ns | ||
| IL-9 | 0.2363 | 0.2006 | ns | 0.1250 | 0.5028 | ns | ||
| IFN-γ | 0.1872 | 0.3219 | ns | 0.0116 | 0.9515 | ns | ||
| CLA−T/EPI | IL-17F | 0.5028 | 0.0039 | ** | −0.0267 | 0.8864 | ns | |
| IL-17A | 0.4714 | 0.0074 | ** | 0.0625 | 0.7383 | ns | ||
| IL-9 | 0.2558 | 0.1724 | ns | 0.1183 | 0.5264 | ns | ||
| IFN-γ | −0.0073 | 0.9690 | ns | −0.0589 | 0.7573 | ns | ||
| Healthy controls (n = 12) | CLA+T/EPI | IL-17F | −0.4496 | 0.1681 | ns | 0.0458 | 0.9015 | ns |
| IL-17A | −0.4307 | 0.1622 | ns | −0.0392 | 0.9061 | ns | ||
| IL-9 | −0.5691 | 0.0591 | ns | 0.0734 | 0.8288 | ns | ||
| IFN-γ | −0.2203 | 0.5015 | ns | −0.3885 | 0.2227 | ns | ||
| CLA−T/EPI | IL-17F | 0.2197 | 0.5192 | ns | 0.2197 | 0.5192 | ns | |
| IL-17A | −0.1706 | 0.5948 | ns | 0.2538 | 0.4246 | ns | ||
| IL-9 | −0.2527 | 0.4394 | ns | 0.0161 | 0.9697 | ns | ||
| IFN-γ | −0.0275 | 0.9394 | ns | 0.4773 | 0.1258 | ns | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Jesús-Gil, C.; Sans-de San Nicolàs, L.; Ruiz-Romeu, E.; Ferran, M.; Soria-Martínez, L.; García-Jiménez, I.; Chiriac, A.; Casanova-Seuma, J.M.; Fernández-Armenteros, J.M.; Owens, S.; et al. Interplay between Humoral and CLA+ T Cell Response against Candida albicans in Psoriasis. Int. J. Mol. Sci. 2021, 22, 1519. https://doi.org/10.3390/ijms22041519
de Jesús-Gil C, Sans-de San Nicolàs L, Ruiz-Romeu E, Ferran M, Soria-Martínez L, García-Jiménez I, Chiriac A, Casanova-Seuma JM, Fernández-Armenteros JM, Owens S, et al. Interplay between Humoral and CLA+ T Cell Response against Candida albicans in Psoriasis. International Journal of Molecular Sciences. 2021; 22(4):1519. https://doi.org/10.3390/ijms22041519
Chicago/Turabian Stylede Jesús-Gil, Carmen, Lídia Sans-de San Nicolàs, Ester Ruiz-Romeu, Marta Ferran, Laura Soria-Martínez, Irene García-Jiménez, Anca Chiriac, Josep Manel Casanova-Seuma, Josep Manel Fernández-Armenteros, Sherry Owens, and et al. 2021. "Interplay between Humoral and CLA+ T Cell Response against Candida albicans in Psoriasis" International Journal of Molecular Sciences 22, no. 4: 1519. https://doi.org/10.3390/ijms22041519