Cabs1 Maintains Structural Integrity of Mouse Sperm Flagella during Epididymal Transit of Sperm

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
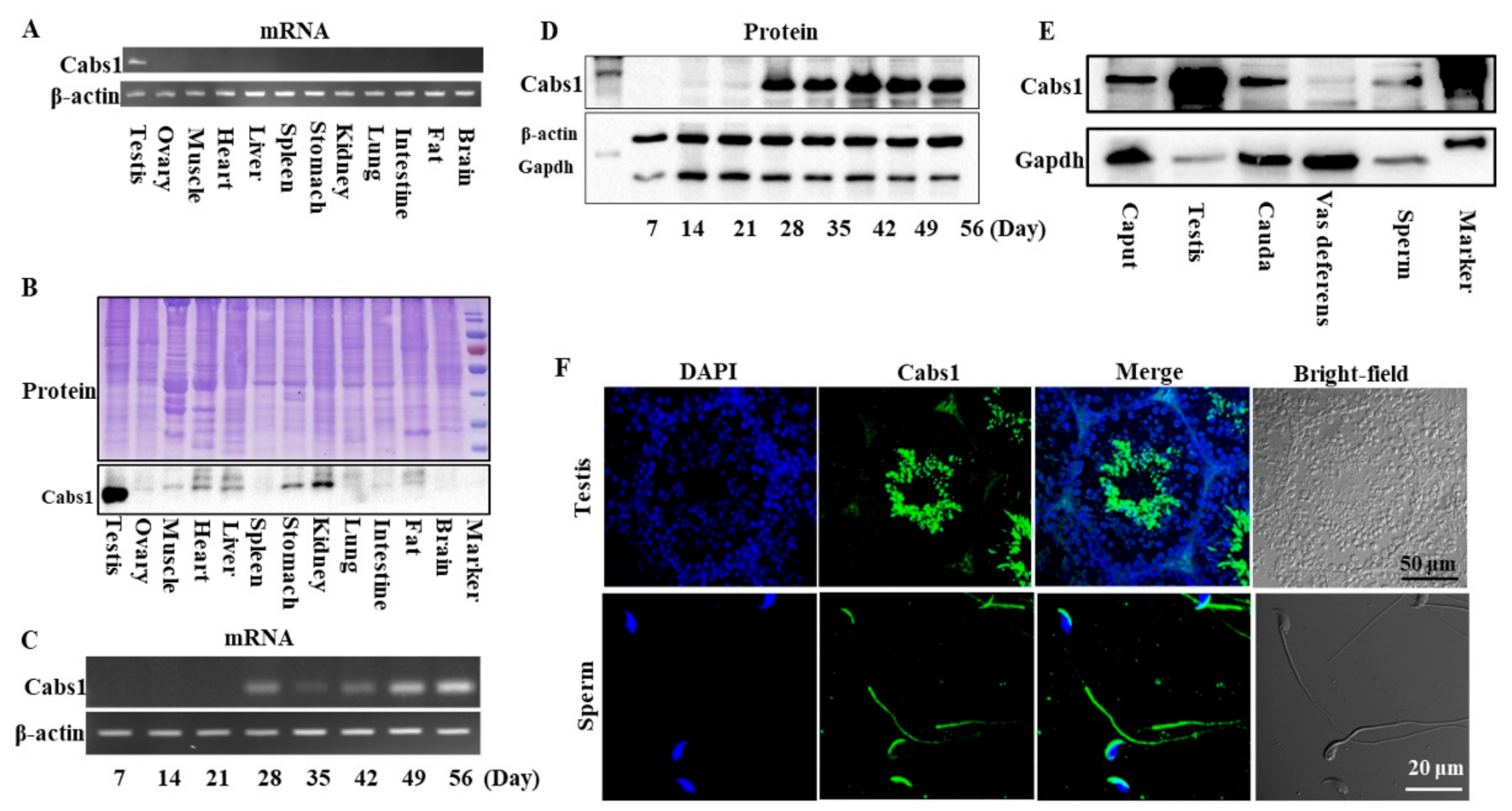
2.1. Temporal and Spatial Expression Profiles of Cabs1 and AntiCabs1
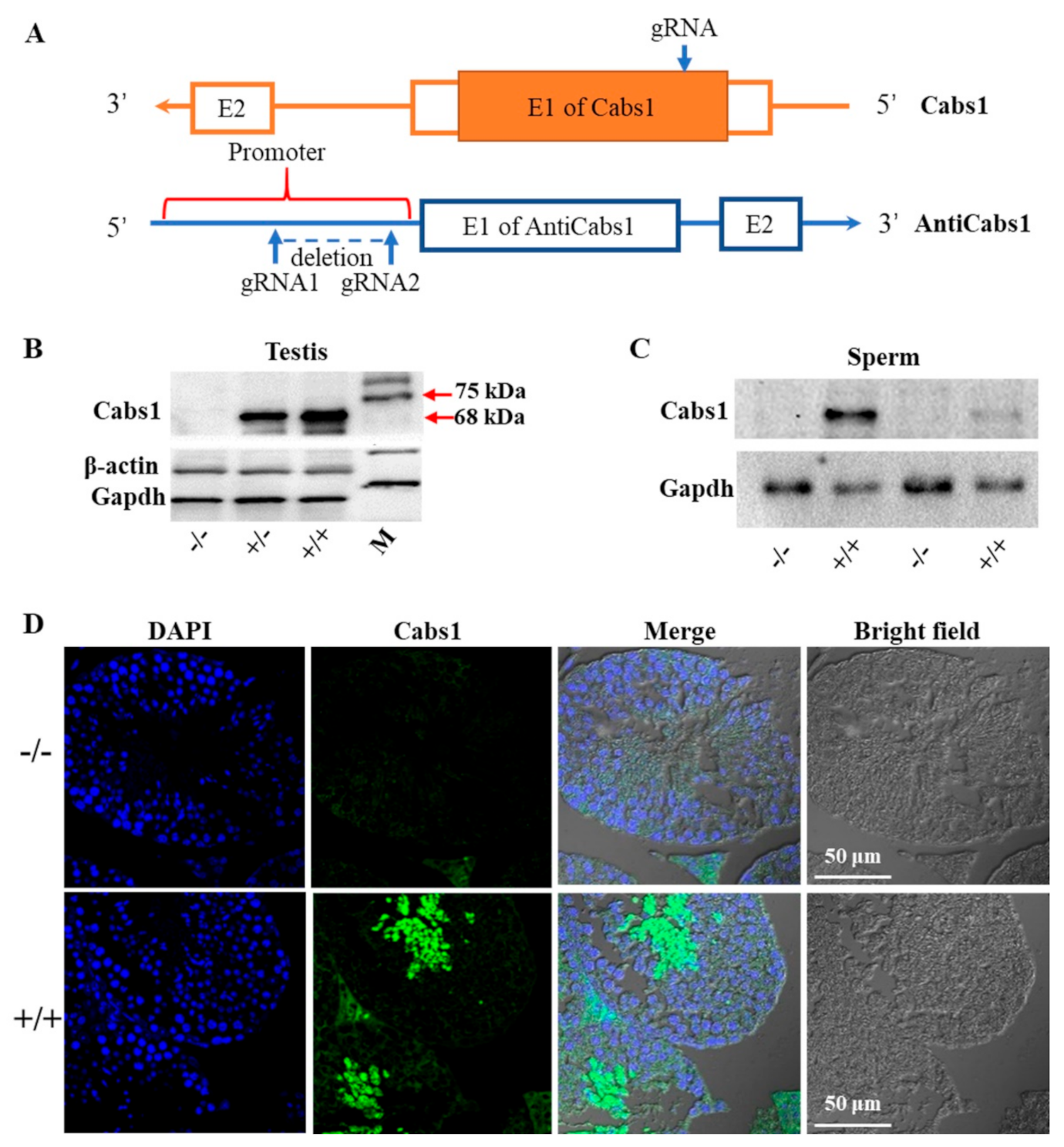
2.2. Generation of Cabs1 and AntiCabs1 KO Mice
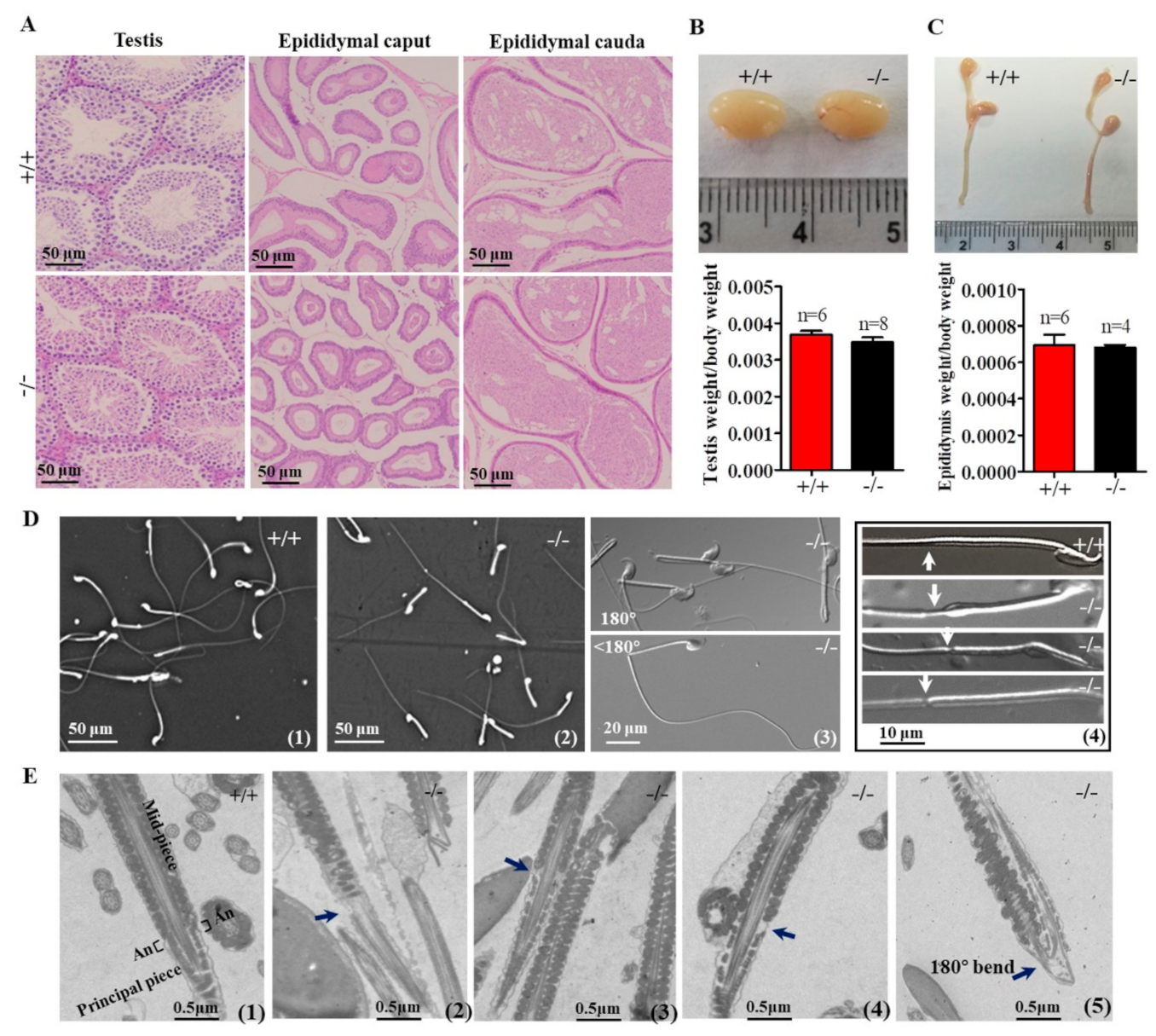
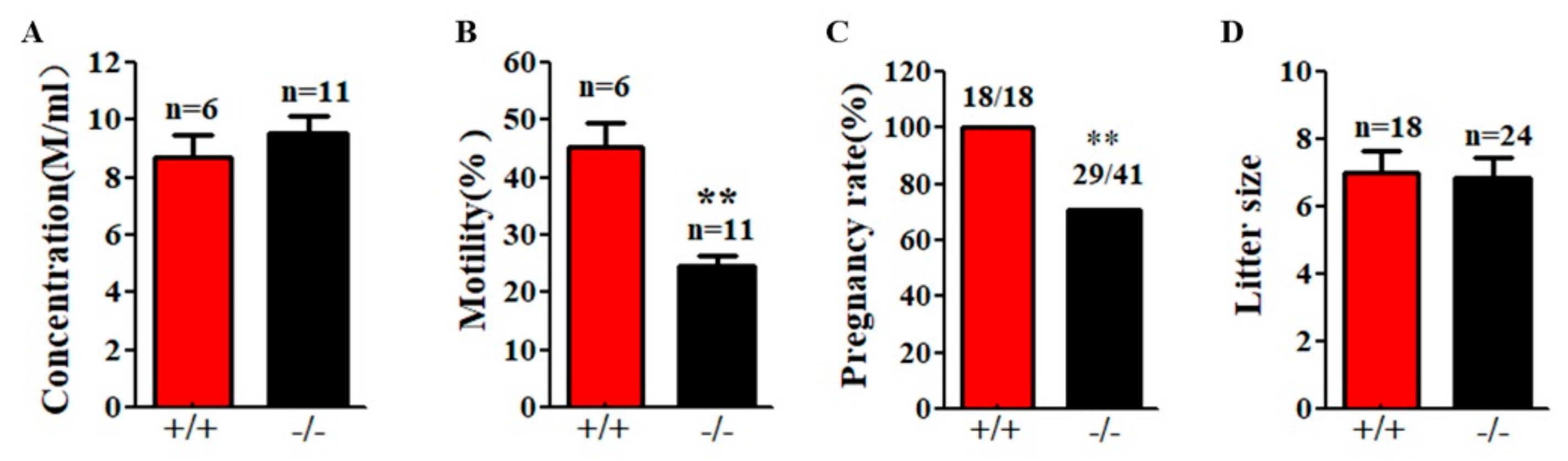
2.3. Genetic Loss of Cabs1 Caused Sperm Malformation
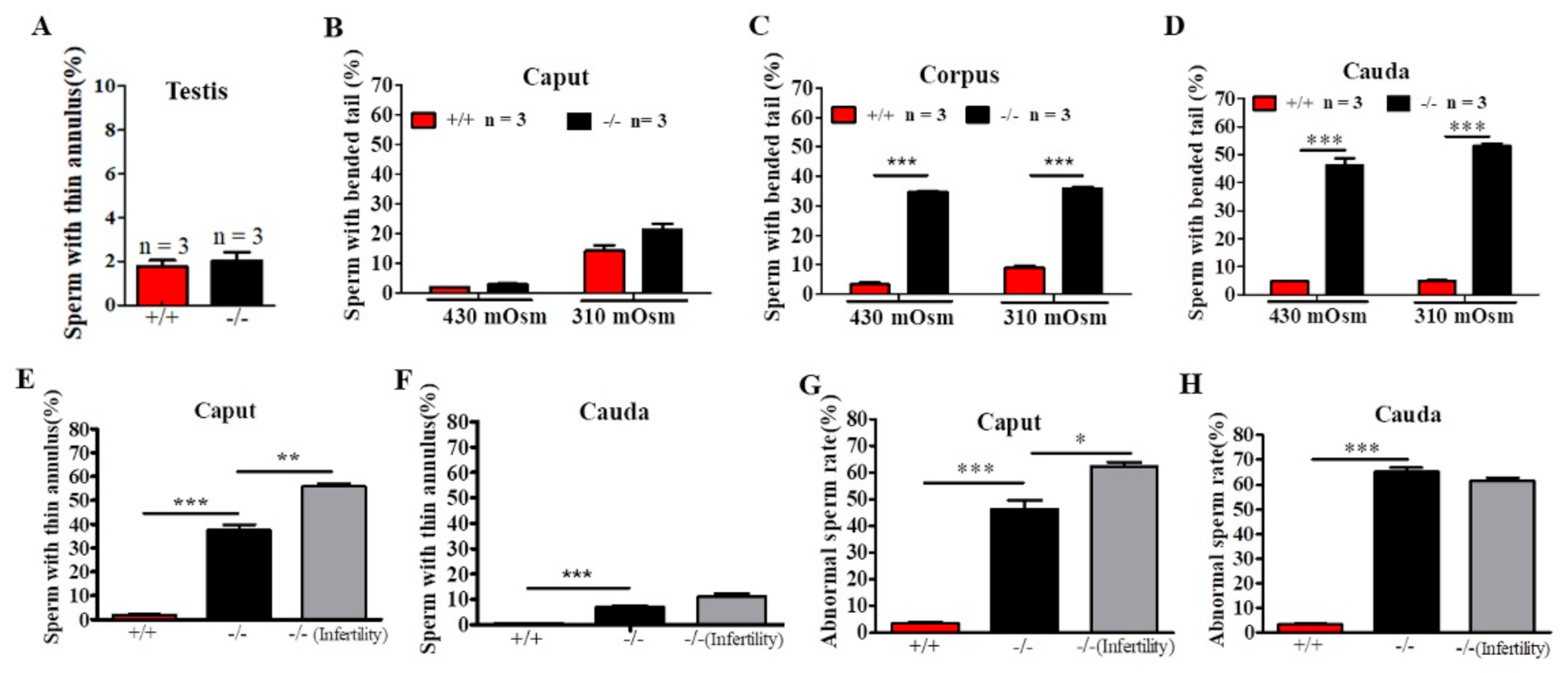
2.4. Defects in Sperm Flagella Occurred in the Epididymis of Cabs1−/− Mice
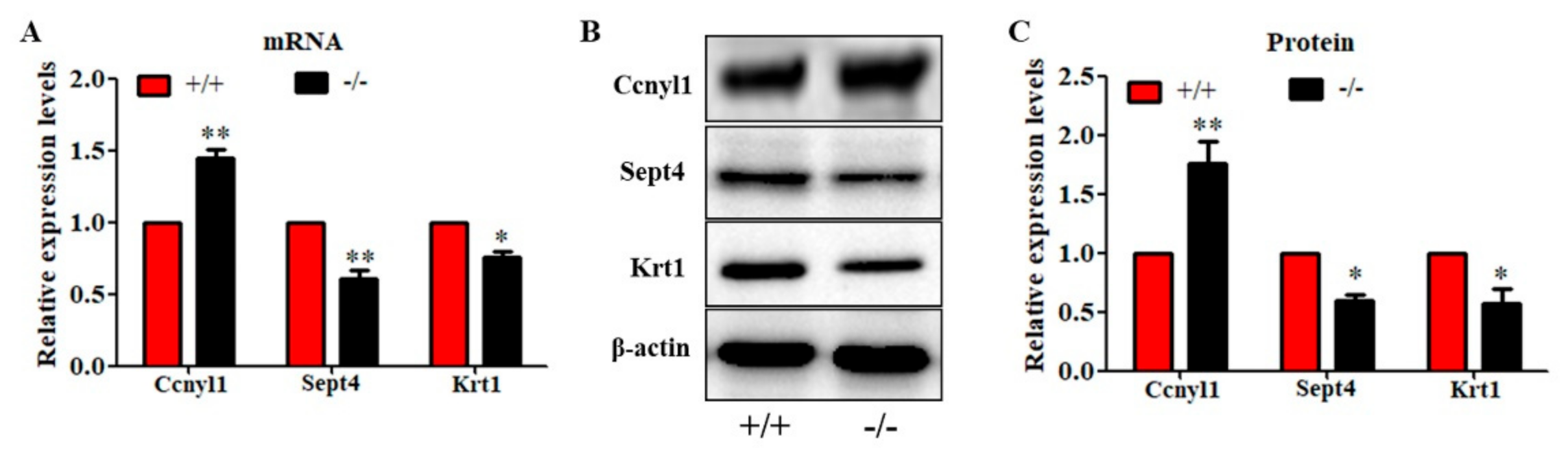
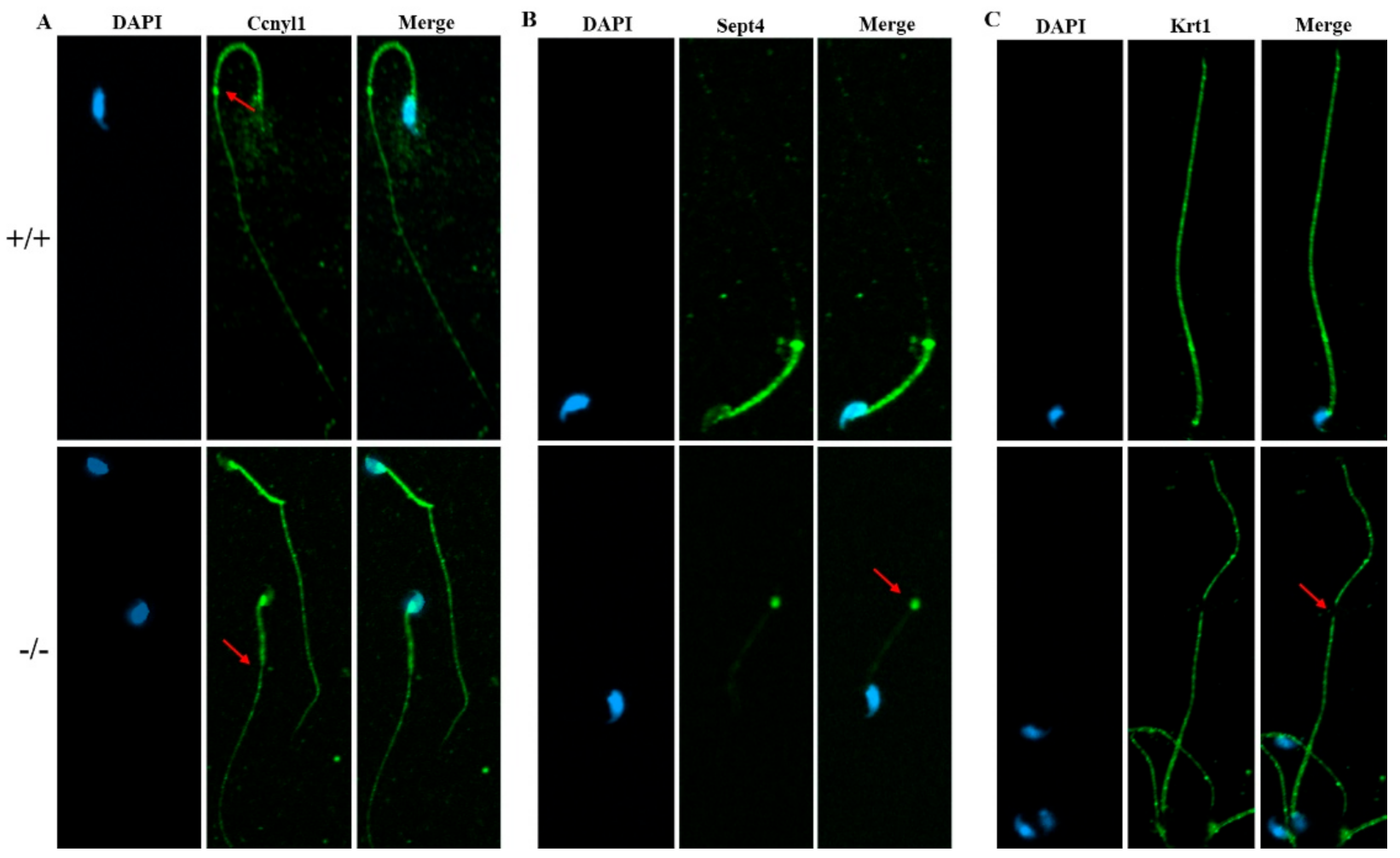
2.5. Deletion of Cabs1 Causes Abnormal Expression of Flagella-Related Proteins
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Histological Analysis
4.3. Sperm Morphology and Motility Analysis
4.4. RT-PCR and Real-Time Quantitative PCR (qPCR)
4.5. Western Blot and Immunofluorescence Staining
4.6. Transmission Electron Microscopy (TEM)
4.7. Cell Culture and Transfection
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CRISPR | Clustered Regularly Interspaced Short Palindromic Repeats |
| DMEM | Dulbecco’s modified Eagle’s medium |
| HS | High-Salt Solution |
| EGFP | Enhanced Green Fluorescent Protein |
| RIPA | Radio Immunoprecipitation Assay |
References
- Kamiński, P.; Baszyński, J.; Jerzak, I.; Kavanagh, B.P.; Nowacka-Chiari, E.; Polanin, M.; Szymański, M.; Woźniak, A.; Kozera, W. External and genetic conditions determining male infertility. Int. J. Mol. Sci. 2020, 21, 5274. [Google Scholar]
- Wong, W.Y.; Zielhuis, G.A.; Thomas, C.M.; Merkus, H.M.; Steegers-Theunissen, R.P. New evidence of the influence of exogenous and endogenous factors on sperm count in man. Eur. J. Obstet. Gynecol. Reprod. 2003, 110, 49–54. [Google Scholar]
- Nantel, F.; Monaco, L.; Foulkes, N.S.; Masquilier, D.; LeMeur, M.; Henriksén, K.; Dierich, A.; Parvinen, M.; Sassone-Corsi, P. Spermiogenesis deficiency and germ-cell apoptosis in CREM-mutant mice. Nature 1996, 380, 159–162. [Google Scholar] [PubMed]
- Wu, J.Y.; Ribar, T.J.; Cummings, D.E.; Burton, K.A.; McKnight, G.S.; Means, A.R. Spermiogenesis and exchange of basic nuclear proteins are impaired in male germ cells lacking Camk4. Nat. Genet. 2000, 25, 448–452. [Google Scholar]
- Yan, W. Male infertility caused by spermiogenic defects: Lessons from gene knockouts. Mol. Cell. Endocrinol. 2009, 306, 24–32. [Google Scholar]
- Escalier, D. Knockout mouse models of sperm flagellum anomalies. Hum. Reprod. Update 2006, 12, 449–461. [Google Scholar]
- Kawashima, A.; Osman, B.A.; Takashima, M.; Kikuchi, A.; Kohchi, S.; Satoh, E.; Tamba, M.; Matsuda, M.; Okamura, N. CABS1 is a novel calcium-binding protein specifically expressed in elongate spermatids of mice. Biol. Reprod. 2009, 80, 1293–1304. [Google Scholar]
- Shawki, H.H.; Kigoshi, T.; Katoh, Y.; Matsuda, M.; Ugboma, C.M.; Takahashi, S.; Oishi, H.; Kawashima, A. Identification, localization, and functional analysis of the homologues of mouse CABS1 protein in porcine testis. Exp. Anim. 2016, 63, 253–265. [Google Scholar]
- Ritz, T.; Rosenfield, D.; Laurent, C.D.S.; Trueba, A.F.; Werchan, C.A.; Vogel, P.D.; Auchus, R.J.; Reyes-Serratos, E.; Befus, A.D. A novel biomarker associated with distress in humans: Calcium-binding protein, spermatid-specific 1 (CABS1). Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 312, 1004–1016. [Google Scholar]
- Calvel, P.; Kervarrec, C.; Lavigne, R.; Vallet-Erdtmann, V.; Guerrois, M.; Rolland, A.D.; Chalmel, F.; Jégou, B.; Pineau, C. CLPH, a novel casein kinase 2-phosphorylated disordered protein, is specifically associated with postmeiotic germ cells in rat spermatogenesis. J. Proteome Res. 2009, 8, 2953–2965. [Google Scholar]
- Xu, X.; Toselli, P.A.; Russell, L.D.; Seldin, D.C. Globozoospermia in mice lacking the casein kinase II α’ catalytic subunit. Nat. Genet. 1999, 23, 118–121. [Google Scholar] [PubMed]
- Marcet-Palacios, M.; Reyes-Serratos, E.; Gonshor, A.; Buck, R.; Lacy, P.; Befus, A. Structural and posttranslational analysis of human calcium-binding protein, spermatid-associated 1. J. Cell. Biochem. 2020, 121, 4945–4958. [Google Scholar] [CrossRef]
- Sun, J.; Lu, Y.; Nozawa, K.; Xu, Z.; Morohoshi, A.; Castaneda, J.M.; Noda, T.; Miyata, H.; Abbasi, F.; Shawki, H.H.; et al. CRISPR/Cas9-based genome editing in mice uncovers 13 testis-or epididymis-enriched genes individually dispensable for male reproduction. Biol. Reprod. 2020, 103, 183–194. [Google Scholar] [PubMed]
- Bhat, S.A.; Ahmad, S.M.; Mumtaz, P.T.; Malik, A.A.; Dar, M.A.; Urwat, U.; Shah, R.A.; Ganai, N.A. Long non-coding RNAs: Mechanism of action and functional utility. Non Coding RNA Res. 2016, 1, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Yan, G.-Y. Novel human lncRNA–disease association inference based on lncRNA expression profiles. Bioinformatics 2013, 29, 2617–2624. [Google Scholar]
- Necsulea, A.; Soumillon, M.; Warnefors, M.; Liechti, A.; Daish, T.; Zeller, U.; Baker, J.C.; Grützner, F.; Kaessmann, H. The evolution of lncRNA repertoires and expression patterns in tetrapods. Nature 2014, 505, 635–640. [Google Scholar] [CrossRef]
- Lin, X.; Han, M.; Cheng, L.; Chen, J.; Zhang, Z.; Shen, T.; Wang, M.; Wen, B.; Ni, T.; Han, C. Expression dynamics, relationships, and transcriptional regulations of diverse transcripts in mouse spermatogenic cells. RNA Biol. 2016, 13, 1011–1024. [Google Scholar] [CrossRef] [Green Version]
- Wen, K.; Yang, L.; Xiong, T.; Di, C.; Ma, D.; Wu, M.; Xue, Z.; Zhang, X.; Long, L.; Zhang, W. Critical roles of long noncoding RNAs in Drosophila spermatogenesis. Genome Res. 2016, 26, 1233–1244. [Google Scholar]
- Dai, Y.-B.; Lin, Y.; Song, N.; Sun, F. LncRNA4667 is dispensable for spermatogenesis and fertility in mice. Reprod. Dev. Med. 2019, 3, 18–23. [Google Scholar]
- Goudarzi, M.; Berg, K.; Pieper, L.M.; Schier, A.F. Individual long non-coding RNAs have no overt functions in zebrafish embryogenesis, viability and fertility. Elife 2019, 8, e40815. [Google Scholar]
- Li, C.; Shen, C.; Shang, X.; Tang, L.; Xiong, W.; Ge, H.; Zhang, H.; Lu, S.; Shen, Y.; Wang, J. Two novel testis-specific long noncoding RNAs produced by 1700121C10Rik are dispensable for male fertility in mice. J. Reprod. Dev. 2019, 66, 57–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil, N.; Ulitsky, I. Regulation of gene expression by cis-acting long non-coding RNAs. Nat. Rev. Genet. 2020, 21, 102–117. [Google Scholar] [CrossRef] [PubMed]
- Matzuk, M.M.; Lamb, D.J. Genetic dissection of mammalian fertility pathways. Nat. Med. 2002, 8, 33–40. [Google Scholar] [CrossRef]
- Befus, D.; Ritz, T.; Laurent, C.S.; Laurent, K.S. Calcium Binding Protein, Spermatid Specific 1, as a Biomarker for Diagnosis or Treatment of Stress. U.S. Patent Application No. 16/084,617, 21 September 2019. [Google Scholar]
- St. Laurent, C.D.; St. Laurent, K.E.; Mathison, R.D.; Befus, A.D. Calcium-binding protein, spermatid-specific 1 is expressed in human salivary glands and contains an anti-inflammatory motif. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, 569–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engreitz, J.M.; Haines, J.E.; Perez, E.M.; Munson, G.; Chen, J.; Kane, M.; McDonel, P.E.; Guttman, M.; Lander, E.S. Local regulation of gene expression by lncRNA promoters, transcription and splicing. Nature 2016, 539, 452–455. [Google Scholar] [CrossRef] [PubMed]
- Goyal, A.; Myacheva, K.; Groß, M.; Klingenberg, M.; Duran Arqué, B.; Diederichs, S. Challenges of CRISPR/Cas9 applications for long non-coding RNA genes. Nucleic Acids Res. 2017, 45, e12. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Liu, Y.; Yang, Z.; Liu, F.; Yang, Y.; Han, X.; Geng, T.; Gao, B.; Song, C.; Cui, H. Efficient knockout of lncRNA promoter region by CRISPR/Cas9 system in zebrafish (Danio rerio). J. Agr. Biotechnol. 2016, 24, 649–656. [Google Scholar]
- Lishko, P.V.; Mannowetz, N. CatSper: A unique calcium channel of the sperm flagellum. Curr. Opin. Physiol. 2018, 2, 109–113. [Google Scholar] [CrossRef]
- Zhu, Y.; Lin, Y.; He, Y.; Wang, H.; Chen, S.; Li, Z.; Song, N.; Sun, F. Deletion of lncRNA5512 has no effect on spermatogenesis and reproduction in mice. Reprod. Fertil. Dev. 2020, 32, 706–713. [Google Scholar]
- Wei, S.; Chen, H.; Dzakah, E.E.; Yu, B.; Wang, X.; Fu, T.; Li, J.; Liu, L.; Fang, S.; Liu, W. Systematic evaluation of C. elegans lincRNAs with CRISPR knockout mutants. Genome Biol. 2019, 20, 1–19. [Google Scholar]
- Sahlu, B.W.; Zhao, S.; Wang, X.; Umer, S.; Zou, H.; Huang, J.; Zhu, H. Long noncoding RNAs: New insights in modulating mammalian spermatogenesis. J. Anim. Sci. Biotechnol. 2020, 11, 1–12. [Google Scholar]
- Zeng, X.-H.; Yang, C.; Kim, S.T.; Lingle, C.J.; Xia, X.-M. Deletion of the Slo3 gene abolishes alkalization-activated K+ current in mouse spermatozoa. Proc. Natl. Acad. Sci. USA 2011, 108, 5879–5884. [Google Scholar] [PubMed] [Green Version]
- Chen, Q.; Peng, H.; Lei, L.; Zhang, Y.; Kuang, H.; Cao, Y.; Shi, Q.-x.; Ma, T.; Duan, E. Aquaporin3 is a sperm water channel essential for postcopulatory sperm osmoadaptation and migration. Cell. Res. 2011, 21, 922–933. [Google Scholar] [PubMed]
- Joseph, A.; Shur, B.D.; Ko, C.; Chambon, P.; Hess, R.A. Epididymal hypo-osmolality induces abnormal sperm morphology and function in the estrogen receptor alpha knockout mouse. Biol. Reprod. 2010, 82, 958–967. [Google Scholar] [PubMed] [Green Version]
- Gervasi, M.G.; Visconti, P.E. Molecular changes and signaling events occurring in spermatozoa during epididymal maturation. Andrology 2017, 5, 204–218. [Google Scholar]
- Zhou, W.; De Iuliis, G.N.; Dun, M.D.; Nixon, B. Characteristics of the epididymal luminal environment responsible for sperm maturation and storage. Front. Endocrinol. (Lausanne) 2018, 9, 59. [Google Scholar] [CrossRef] [Green Version]
- Skerget, S.; Rosenow, M.A.; Petritis, K.; Karr, T.L. Sperm proteome maturation in the mouse epididymis. PLoS ONE 2015, 10, e0140650. [Google Scholar] [CrossRef]
- Nixon, B.; De Iuliis, G.N.; Hart, H.M.; Zhou, W.; Mathe, A.; Bernstein, I.R.; Anderson, A.L.; Stanger, S.J.; Skerrett-Byrne, D.A.; Jamaluddin, M.F.B. Proteomic profiling of mouse epididymosomes reveals their contributions to post-testicular sperm maturation. Mol. Cell. Proteom. 2019, 18, 91–108. [Google Scholar] [CrossRef] [Green Version]
- Kissel, H.; Georgescu, M.-M.; Larisch, S.; Manova, K.; Hunnicutt, G.R.; Steller, H. The Sept4 septin locus is required for sperm terminal differentiation in mice. Dev. Cell. 2005, 8, 353–364. [Google Scholar]
- Ihara, M.; Kinoshita, A.; Yamada, S.; Tanaka, H.; Tanigaki, A.; Kitano, A.; Goto, M.; Okubo, K.; Nishiyama, H.; Ogawa, O. Cortical organization by the septin cytoskeleton is essential for structural and mechanical integrity of mammalian spermatozoa. Dev. Cell. 2005, 8, 343–352. [Google Scholar]
- Touré, A.; Lhuillier, P.; Gossen, J.A.; Kuil, C.W.; Lhôte, D.; Jégou, B.; Escalier, D.; Gacon, G. The testis anion transporter 1 (Slc26a8) is required for sperm terminal differentiation and male fertility in the mouse. Hum. Mol. Genet. 2007, 16, 1783–1793. [Google Scholar] [CrossRef] [PubMed]
- Olson, G.E.; Winfrey, V.P.; NagDas, S.K.; Hill, K.E.; Burk, R.F. Selenoprotein P is required for mouse sperm development. Biol. Reprod. 2005, 73, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Zi, Z.; Zhang, Z.; Li, Q.; An, W.; Zeng, L.; Gao, D.; Yang, Y.; Zhu, X.; Zeng, R.; Shum, W.W. CCNYL1, but not CCNY, cooperates with CDK16 to regulate spermatogenesis in mouse. PLoS Genet. 2015, 11, e1005485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gervasi, M.G.; Xu, X.; Carbajal-Gonzalez, B.; Buffone, M.G.; Visconti, P.E.; Krapf, D. The actin cytoskeleton of the mouse sperm flagellum is organized in a helical structure. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashemitabar, M.; Sabbagh, S.; Orazizadeh, M.; Ghadiri, A.; Bahmanzadeh, M. A proteomic analysis on human sperm tail: Comparison between normozoospermia and asthenozoospermia. J. Assist. Reprod. Genet. 2015, 32, 853–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Zhou, W.; Zhang, P.; Gao, F.; Zhao, X.; Shum, W.W.; Zeng, X. Cabs1 Maintains Structural Integrity of Mouse Sperm Flagella during Epididymal Transit of Sperm. Int. J. Mol. Sci. 2021, 22, 652. https://doi.org/10.3390/ijms22020652
Zhang X, Zhou W, Zhang P, Gao F, Zhao X, Shum WW, Zeng X. Cabs1 Maintains Structural Integrity of Mouse Sperm Flagella during Epididymal Transit of Sperm. International Journal of Molecular Sciences. 2021; 22(2):652. https://doi.org/10.3390/ijms22020652
Chicago/Turabian StyleZhang, Xiaoning, Wenwen Zhou, Peng Zhang, Fengxin Gao, Xiuling Zhao, Winnie Waichi Shum, and Xuhui Zeng. 2021. "Cabs1 Maintains Structural Integrity of Mouse Sperm Flagella during Epididymal Transit of Sperm" International Journal of Molecular Sciences 22, no. 2: 652. https://doi.org/10.3390/ijms22020652