Mitochondrial Membranes of Human SH-SY5Y Neuroblastoma Cells Express Serotonin 5-HT7 Receptor

 ,
,  , , , , , and
, , , , , and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. 5-HT7Rs Are Located in Cytosolic and Mitochondrial Fractions of SH-SY5Y Cells
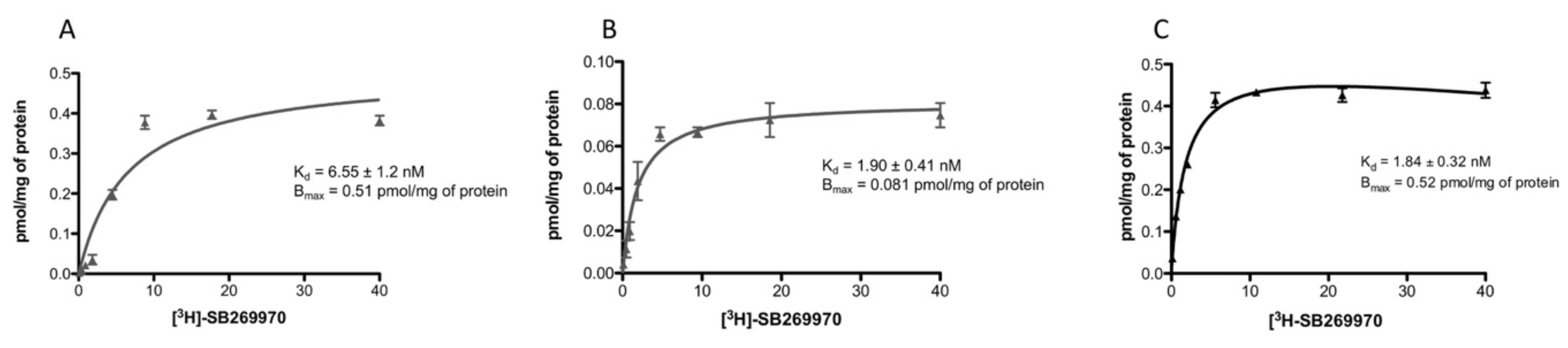
2.2. Saturation-Binding Assay
2.3. Administration of SB-269970 (but Not LP-211) to Mitochondria Weakly Influences Mitochondrial Respiratory Chain (MRC) Cytochrome c Oxidase Activity
3. Discussion
4. Materials and Methods
4.1. Drugs
4.2. Cell Culture
4.3. Mitochondrial-Enriched Fraction
4.4. Western Blot Analysis
4.5. Cytochrome c Oxidase Activity Measurements
4.6. SH-SY5Y Membrane Preparation for Saturation-Binding Assay
4.7. Saturation-Binding Assay
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 5-HT | 5-hydroxytryptamine |
| cAMP | cyclic Adenosine MonoPhosphate |
| CNS | Central Nervous System |
| GPCR | G-protein-coupled receptor |
| MCR | Mitochondrial respiratory chain |
References
- Sriram, K.; Insel, P.A. G Protein-Coupled receptors as targets for approved drugs: How many targets and how many Drugs? Mol. Pharmacol. 2018, 93, 251–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jong, Y.I.; Harmon, S.K.; O’Malley, K.L. Intracellular GPCRs play key roles in synaptic plasticity. ACS Chem. Neurosci. 2018, 9, 2162–2172. [Google Scholar] [CrossRef] [PubMed]
- Belous, A.; Wakata, A.; Knox, C.D.; Nicoud, I.B.; Pierce, J.; Anderson, C.D.; Pinson, C.W.; Chari, R.S. Mitochondrial P2Y-Like receptors link cytosolic adenosine nucleotides to mitochondrial calcium uptake. J. Cell. Biochem. 2004, 92, 1062–1073. [Google Scholar] [CrossRef] [PubMed]
- Abadir, P.M.; Walston, J.D.; Carey, R.M. Subcellular characteristics of functional intracellular renin-angiotensin systems. Peptides 2012, 38, 437–445. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zhang, H.; Xu, H.; Guo, D.; Shi, H.; Li, Y.; Zhang, W.; Gu, Y. 5HTR3 and 5-HTR4 located on the mitochondrial membrane and functionally regulated mitochondrial functions. Sci. Rep. 2016, 6, 37336. [Google Scholar] [CrossRef] [Green Version]
- Suofu, Y.L.W.; Jean-Alphonse, F.G.; Jia, J.; Khattar, N.K.; Li, J.; Baranov, S.V.; Leronni, D.; Mihalik, A.C.; He, Y.; Cecon, E.; et al. Dual role of mitochondria in producing melatonin and driving GPCR signaling to block cytochrome c release. Proc. Natl. Acad. Sci. USA 2017, 114, E7997–E8006. [Google Scholar] [CrossRef] [Green Version]
- Modica, M.N.; Lacivita, E.; Intagliata, S.; Salerno, L.; Romeo, G.; Pittalà, V.; Leopoldo, M. Structure-activity relationships and therapeutic potentials of 5-HT7 receptor ligands: An update. J. Med. Chem. 2018, 61, 8475–8503. [Google Scholar] [CrossRef]
- Crispino, M.; Volpicelli, F.; Perrone-Capano, C. Role of the serotonin receptor 7 in brain plasticity: From development to disease. Int. J. Mol. Sci. 2020, 21, 505. [Google Scholar] [CrossRef] [Green Version]
- Costa, L.; Sardone, L.M.; Bonaccorso, C.M.; D’Antoni, S.; Spatuzza, M.; Gulisano, W.; Tropea, M.R.; Puzzo, D.; Leopoldo, M.; Lacivita, E.; et al. Activation of Serotonin 5-HT7 Receptors Modulates Hippocampal Synaptic Plasticity by Stimulation of Adenylate Cyclases and Rescues Learning and Behavior in a Mouse Model of Fragile X Syndrome. Front. Mol. Neurosci. 2018, 11, 353. [Google Scholar] [CrossRef] [Green Version]
- Hedlund, P.B.; Leopoldo, M.; Caccia, S.; Sarkisyan, G.; Fracasso, C.; Martelli, G.; Lacivita, E.; Berardi, F.; Perrone, R. LP-211 is a brain penetrant selective agonist for the serotonin 5-HT7 receptor. Neurosci. Lett. 2010, 481, 12–16. [Google Scholar] [CrossRef] [Green Version]
- Valenti, D.; de Bari, L.; Vigli, D.; Lacivita, E.; Leopoldo, M.; Laviola, G.; Vacca, R.A.; De Filippis, B. Stimulation of the brain serotonin receptor 7 rescues mitochondrial dysfunction in female mice from two models of Rett syndrome. Neuropharmacology 2017, 121, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Vigli, D.; Rusconi, L.; Valenti, D.; La Montanara, P.; Cosentino, L.; Lacivita, E.; Leopoldo, M.; Amendola, E.; Gross, C.; Landsberger, N.; et al. Rescue of prepulse inhibition deficit and brain mitochondrial dysfunction by pharmacological stimulation of the central serotonin receptor 7 in a mouse model of CDKL5 Deficiency Disorder. Neuropharmacology 2019, 144, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Rangaraju, V.; Lewis, T.L., Jr.; Hirabayashi, Y.; Bergami, M.; Motori, E.; Cartoni, R.; Kwon, S.K.; Courchet, J. Pleiotropic mitochondria: The influence of mitochondria on neuronal development and disease. J. Neurosci. 2019, 39, 8200–8208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yusuf, M.; Leung, K.; Morris, K.J.; Volpi, E.V. Comprehensive cytogenomic profile of the in vitro neuronal model SH-SY5Y. Neurogenetics 2013, 14, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Samarajeewa, A.; Goldemann, L.; Vasefi, M.S.; Ahmed, N.; Gondora, N.; Khanderia, C.; Mielke, J.G.; Beazely, M.A. 5-HT7 receptor activation promotes an increase in TrkB receptor expression and phosphorylation. Front. Behav. Neurosci. 2014, 8, 391. [Google Scholar] [CrossRef] [Green Version]
- Vasefi, M.S.; Kruk, J.S.; Liu, H.; Heikkila, J.J.; Beazely, M.A. Activation of 5-HT7 receptors increases neuronal platelet-derived growth factor β receptor expression. Neurosci. Lett. 2012, 511, 65–69. [Google Scholar] [CrossRef]
- Yuksel, T.N.; Yayla, M.; Halici, Z.; Cadirci, E.; Polat, B.; Kose, D. Protective effect of 5-HT7 receptor activation against glutamate-induced neurotoxicity in human neuroblastoma SH-SY5Y cells via antioxidative and antiapoptotic pathways. Neurotoxicol. Teratol. 2019, 72, 22–28. [Google Scholar] [CrossRef]
- Lacivita, E.; Niso, M.; Stama, M.L.; Arzuaga, A.; Altamura, C.; Costa, L.; Desaphy, J.F.; Ragozzino, M.E.; Ciranna, L.; Leopoldo, M. Privileged scaffold-based design to identify a novel drug-like 5-HT7 receptor-preferring agonist to target Fragile X syndrome. Eur. J. Med. Chem. 2020, 199, 112395. [Google Scholar] [CrossRef]
- Heidmann, D.E.; Metcalf, M.A.; Kohen, R.; Hamblin, M.W. Four 5-hydroxytryptamine7 (5-HT7) receptor isoforms in human and rat produced by alternative splicing: Species differences due to altered intron-exon organization. J. Neurochem. 1997, 68, 1372–1381. [Google Scholar] [CrossRef]
- Lovenberg, T.; Baron, B.; de Lecea, L.; Miller, J.; Prosser, R.; Rea, M.; Foye, P.; Racke, M.; Slone, A.; Siegel, B.; et al. A novel adenylyl cyclase-activating serotonin receptor (5-HT7) implicated in the regulation of mammalian circadian rhythms. Neuron 1993, 11, 449–458. [Google Scholar] [CrossRef]
- Gorinski, N.; Ponimaskin, E. Palmitoylation of serotonin receptors. Biochem. Soc. Trans. 2013, 41, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahé, C.; Bernhard, M.; Bobirnac, I.; Keser, C.; Loetscher, E.; Feuerbach, D.; Dev, K.K.; Schoeffter, P. Functional expression of the serotonin 5-HT7 receptor in human glioblastoma cell lines. Br. J. Pharmacol. 2004, 143, 404–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanibunda, S.E.; Deb, S.; Maniyadath, B.; Tiwari, P.; Ghai, U.; Gupta, S.; Figueiredo, D.; Weisstaub, N.; Gingrich, J.A.; Vaidya, A.D.B.; et al. Serotonin regulates mitochondrial biogenesis and function in rodent cortical neurons via the 5-HT2A receptor and SIRT1-PGC-1α axis. Proc. Natl. Acad. Sci. USA 2019, 116, 11028–11037. [Google Scholar] [CrossRef] [Green Version]
- Lovell, P.J.; Bromidge, S.M.; Dabbs, S.; Duckworth, D.M.; Forbes, I.T.; Jennings, A.J.; King, F.D.; Middlemiss, D.N.; Rahman, S.K.; Saunders, D.V.; et al. A novel, potent, and selective 5-HT7 antagonist: (R)-3-(2-(2-(4-methylpiperidin-1-yl)ethyl)pyrrolidine-1-sulfonyl) phen ol (SB-269970). J. Med. Chem. 2000, 43, 342–345. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.R.; Atkinson, P.J.; Ho, M.; Bromidge, S.M.; Lovell, P.J.; Villani, A.J.; Hagan, J.J.; Middlemiss, D.N.; Price, G.W. [3H]-SB-269970--A selective antagonist radioligand for 5-HT7 receptors. Br. J. Pharmacol. 2000, 130, 409–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hevner, R.F.; Wong-Riley, M.T. Brain cytochrome oxidase: Purification, antibody production, and immunohistochemical/histochemical correlations in the CNS. J. Neurosci. 1989, 9, 3884–3898. [Google Scholar] [CrossRef]
- Hüttemann, M.; Lee, I.; Grossman, L.I.; Doan, J.W.; Sanderson, T.H. Phosphorylation of mammalian cytochrome c and cytochrome c oxidase in the regulation of cell destiny: Respiration, apoptosis, and human disease. Adv. Exp. Med. Biol. 2012, 748, 237–264. [Google Scholar] [CrossRef] [Green Version]
- Hebert-Chatelain, E.; Desprez, T.; Serrat, R.; Bellocchio, L.; Soria-Gomez, E.; Busquets-Garcia, A.; Pagano Zottola, A.C.; Delamarre, A.; Cannich, A.; Vincent, P.; et al. A cannabinoid link between mitochondria and memory. Nature 2016, 539, 555–559. [Google Scholar] [CrossRef]
- Spinazzi, M.; Casarin, A.; Pertegato, V.; Salviati, L.; Angelini, C. Assessment of mitochondrial respiratory chain enzymatic activities on tissues and cultured cells. Nat. Protoc. 2012, 7, 1235–1246. [Google Scholar] [CrossRef]
- Valsecchi, F.; Ramos-Espiritu, L.S.; Buck, J.; Levin, L.R.; Manfredi, G. cAMP and mitochondria. Physiology (Bethesda) 2013, 28, 199–209. [Google Scholar] [CrossRef]
- Palacino, J.; Sagi, D.; Goldberg, M.; Krauss, S.; Motz, C.; Wacker, M.; Klose, J.; Shen, J. Mitochondrial dysfunction and oxidative damage in parkin-deficient mice. J. Biol. Chem. 2004, 279, 18614–18622. [Google Scholar] [CrossRef] [Green Version]
- Colabufo, N.A.; Berardi, F.; Contino, M.; Niso, M.; Abate, C.; Perrone, R.; Tortorella, V. Antiproliferative and cytotoxic effects of some sigma2 agonists and sigma1 antagonists in tumour cell lines. Naunyn Schmiedebergs Arch. Pharmacol. 2004, 370, 106–113. [Google Scholar] [CrossRef]
- Volpicelli, F.; Speranza, L.; di Porzio, U.; Crispino, M.; Perrone-Capano, C. The serotonin receptor 7 and the structural plasticity of brain circuits. Front. Behav. Neurosci. 2014, 8, 318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.; Hirabayashi, Y.; Kwon, S.K.; Lewis, T.L., Jr.; Polleux, F. Emerging roles of mitochondria in synaptic transmission and neurodegeneration. Curr. Opin. Physiol. 2018, 3, 82–93. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tempio, A.; Niso, M.; Laera, L.; Trisolini, L.; Favia, M.; Ciranna, L.; Marzulli, D.; Petrosillo, G.; Pierri, C.L.; Lacivita, E.; et al. Mitochondrial Membranes of Human SH-SY5Y Neuroblastoma Cells Express Serotonin 5-HT7 Receptor. Int. J. Mol. Sci. 2020, 21, 9629. https://doi.org/10.3390/ijms21249629
Tempio A, Niso M, Laera L, Trisolini L, Favia M, Ciranna L, Marzulli D, Petrosillo G, Pierri CL, Lacivita E, et al. Mitochondrial Membranes of Human SH-SY5Y Neuroblastoma Cells Express Serotonin 5-HT7 Receptor. International Journal of Molecular Sciences. 2020; 21(24):9629. https://doi.org/10.3390/ijms21249629
Chicago/Turabian StyleTempio, Alessandra, Mauro Niso, Luna Laera, Lucia Trisolini, Maria Favia, Lucia Ciranna, Domenico Marzulli, Giuseppe Petrosillo, Ciro Leonardo Pierri, Enza Lacivita, and et al. 2020. "Mitochondrial Membranes of Human SH-SY5Y Neuroblastoma Cells Express Serotonin 5-HT7 Receptor" International Journal of Molecular Sciences 21, no. 24: 9629. https://doi.org/10.3390/ijms21249629