1. Introduction
Alzheimer’s disease (AD) is the most common form of dementia, characterized by a progressive decline of cognitive functions [
1]. Recent research has seen significant advances in better understanding the cellular and molecular mechanisms of AD. This includes the general consensus that Aβ pathology that starts in the human neocortex 10–15 years before AD becomes symptomatic, and hippocampal tau pathology (which coincides with early stages of symptomatic AD in humans), together, underlie AD onset [
2]. One of the main objectives of AD research is to identify novel therapeutic approaches alleviating cognitive deficits. While lifestyle factors like physical exercise and a healthy diet might be able to delay the onset of AD, no treatment for AD has thus far been reported that can effectively counteract dementia symptoms after disease onset. Importantly, neuroinflammatory signals mediated by microglial cells and concomitant astrogliosis are major hallmarks of AD [
3,
4]. Accordingly, therapeutic approaches focusing on reduction in neuroinflammatory signaling of microglial cells are among the most promising intervention strategies to ameliorate AD deficits. Therefore, repurposing anti-inflammatory drugs for AD therapy that are already approved by the FDA for treatment of human subjects could provide an important benefit to rapidly develop an effective AD medication.
Fingolimod (FTY720) is a food and drug administration (FDA)-approved drug for the treatment of multiple sclerosis (MS) in humans since 2011 and has evolved in recent years as the first-line oral medication for relapsing MS in clinical practice [
5]. In target tissue, it can be converted to fingolimod phosphate, which is a potent modulator of sphingosine-1-phosphate receptors (S1PR
1–5); [
6,
7]. In the immune system, fingolimod-induced S1PR receptor modulation promotes the retention of T-lymphocytes within lymph nodes, thereby reducing the invasion of the CNS by lymphocytes where they promote inflammatory processes leading to demyelination [
7]. More recently, fingolimod was shown to increase brain-derived neurotrophic factor (BDNF) mRNA and protein levels in mouse models of different neurological or neurodegenerative diseases in the cortex, hippocampus, and striatum [
8,
9,
10,
11]. Deficits in BDNF signaling are discussed to contribute to cognitive dysfunctions in AD [
12,
13,
14], and enhanced signaling through the BDNF receptor tropomyosin related kinase B (TrkB) can ameliorate cognitive symptoms in AD model mice [
15,
16]. Given the anti-inflammatory effects of fingolimod and the potentially beneficial effects of this FDA-approved drug on BDNF/TrkB signaling, we set out to explore the capacity of fingolimod to be repurposed for the treatment of AD-like pathology in an amyloid precursor protein/presenilin-1 APP/PS1 Alzheimer mouse model. Importantly, we started with a one to two months long fingolimod treatment period at five to six months of age, when these mice had already entered the symptomatic phase of the disease. Our results reveal rescue of spine deficits, as well as of impaired LTP in this AD mouse model. These effects occurred independent from BDNF signaling but are consistent with the anti-inflammatory actions of fingolimod on microglia and astrocytes that we observed. Hence, our results reveal a high potential of fingolimod as an efficient drug for treatment regimes starting after AD symptoms have been diagnosed.
2. Results
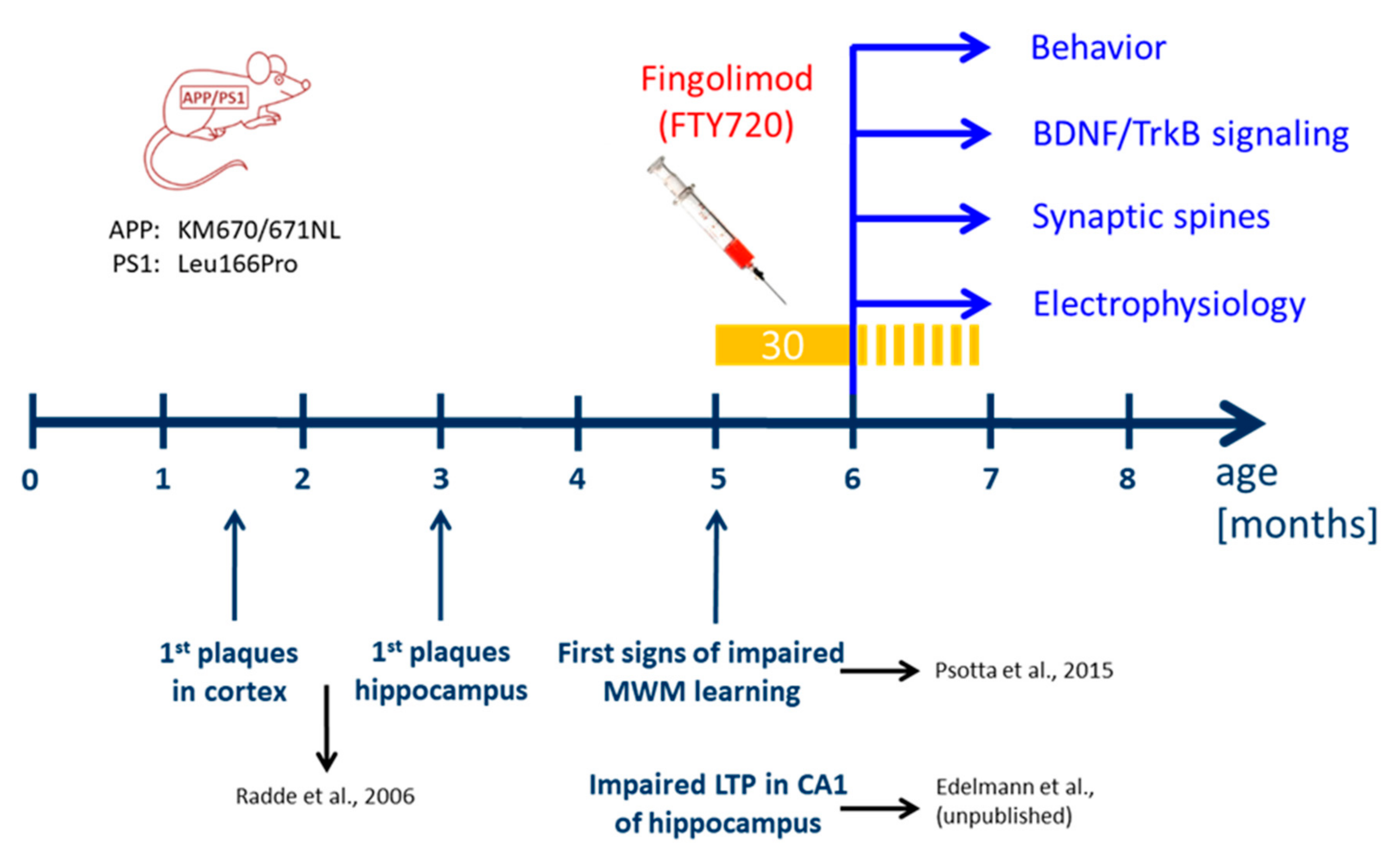
We used a previously established double transgenic APP/PS1 mouse model that starts to develop amyloid-β (Aβ) pathology in the neocortex and hippocampus at two and four months of age, respectively [
17]. Impaired LTP at Schaffer collateral CA1 synapses in acute slices ex vivo is observed at five months in these mice (Edelmann et al., data not shown) and becomes detectable with in vivo recordings beyond six months of age [
18]. Moreover, hippocampus-dependent learning deficits in this APP/PS1 mouse model become apparent five to eight months after birth (
Figure 1; [
17,
19,
20,
21]). To test for a possible amelioration of the AD phenotype at a symptomatic stage in our APP/PS1 mouse model, we started fingolimod (FTY720) treatment at five to six months of age, when AD-like symptoms are present. Male APP/PS1 mice and littermate controls were treated with intraperitoneal (i.p.) injections of fingolimod (FTY720; 1.0 mg/kg body weight) every second day, a dose that was previously reported to be effective against Aβ pathology [
10]. Spine density and Schaffer collateral LTP in CA1 hippocampal neurons, as well as hippocampus-dependent learning were tested at six to seven months (i.e., after four to eight weeks of fingolimod treatment).
2.1. Chronic Fingolimod (FTY720) Treatment of Five- to Six-Month-Old Amyloid Precursor Protein/Presenilin-1 (APP/PS1) Mice Rescues Spine Deficits
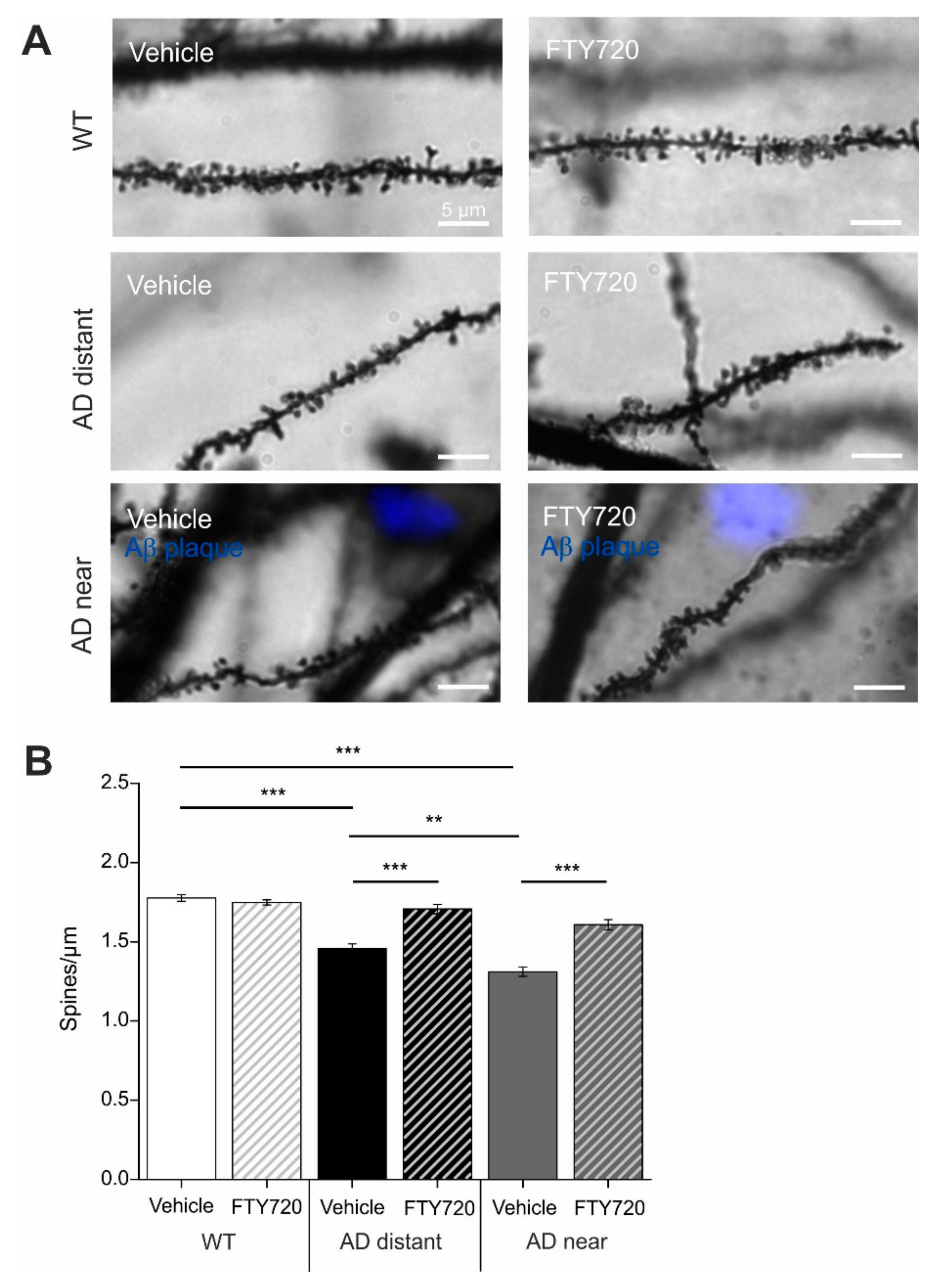
Golgi–Cox staining was used to determine spine densities of cornu ammonis-1 (CA1) pyramidal neurons in secondary apical dendrites located in stratum radiatum of the hippocampus. To directly relate spine numbers to Aβ pathology, we labelled Aβ plaques in six- to seven-month-old animals by i.p. injecting animals twice with the blue fluorescent dye Methoxy X-04 [
22,
23] one day before and two hours before sacrificing the mice for the Golgi staining procedure. In this set of experiments, dendritic stretches analyzed for spine density where subdivided into segments with a minimal distance of spines <50 µm (i.e., 3–44 µm) to the closest Aβ plaque (termed: near), and segments with 50–250 µm (i.e., 54–227 µm) minimal distance to Aβ plaques (termed: distant). Since plaque load is low in CA1 and stratum radiatum of the hippocampus at this AD stage (compare Figure 6A,B), there was no second plaque closer than 300 µm to the analyzed dendrite. Spine analysis in these two groups revealed significantly reduced spine densities in APP/PS1 animals that was more pronounced when spines were located near to Aβ plaques (wild type (WT): 1.78 ± 0.02; APP/PS1 near: 1.31 ± 0.03; APP/PS1 distant: 1.46 ± 0.03;
Figure 2).
Strikingly, one to two months’ treatment of mice with fingolimod (starting at five to six months of age) completely rescued spine densities of dendrites distant to plaques back to the level of WT littermate controls, and significantly ameliorated the spine deficits in dendrites near plaques (APP/PS1 + FTY720 near: 1.61 ± 0.03; APP/PS1 + FTY720 distant: 1.71 ± 0.03, post hoc Tukey’s test with p < 0.0001). Overall, two-way ANOVA analysis revealed a significant main effect of genotype x treatment interaction (F (2, 174) = 21.51 p < 0.0001). Of note, the identical fingolimod treatment of WT littermates did not change CA1 pyramidal neuron spine densities in these animals (WT vehicle: 1.78 ± 0.02; WT + FTY720: 1.75 ± 0.02).
An identical effect of spine rescue in APP/PS1 mice was also observed with a five times lower dose of fingolimod (0.2 mg/kg body weight) applied with the same treatment regime (data not shown).
These results demonstrate that chronic treatment of APP/PS1 mice with fingolimod starting with onset of disease symptoms at five to six months drastically reduced spine pathology in this Alzheimer mouse model.
2.2. Chronic Application of Fingolimod (FTY720) Rescues LTP Deficits at CA3-CA1 Synapses in APP/PS1 Mice
Extracellular field potentials were recorded to analyze long-term potentiation (LTP) in acute hippocampal slices from six- to seven-month-old APP/PS1 mice and WT littermate controls. The recording electrode was placed in stratum radiatum of the CA1 area, and LTP was elicited by high frequency tetanic stimulation of Schaffer collateral axons projecting from the CA3 area. Sixty minutes after LTP induction, the percentage of potentiation was determined in the four groups. The two-way ANOVA showed a significant main effect of genotype x treatment interaction (
F (1,31) = 19.59
p = 0.0001), but no main effects of genotype or treatment. Vehicle treated WT littermates showed robust potentiation of field excitatory postsynaptic potential (EPSP) slopes (WT vehicle: 133.4 ± 4.65%;
Figure 3A,B), whereas in hippocampal slices obtained from vehicle treated APP/PS1 mice LTP was significantly reduced (APP/PS1 vehicle: 107.6 ± 3.2%,
n = 8, significantly different from WT, Tukey comparison
p < 0.01;
Figure 3A,B).
Importantly, chronic treatment of AD mice with FTY720 completely rescued LTP when compared to vehicle-treated AD mice (AD FTY720: 135.9 ± 6.1%,
n = 8, significantly different from AD vehicle, Tukey’s
p < 0.01). Fingolimod treatment of WT littermates reduced LTP magnitude, however, this alteration did not reach significance when compared to WT vehicle (WT + FTY720: 116.8 ± 5.7%, WT vehicle compared to WT FTY720 mice
p > 0.05). While the latter result suggests a trend of fingolimod towards decreasing LTP magnitude in WT animals, this does not harm the complete LTP rescuing effect of the same treatment in APP/PS1 mice. Since fingolimod treatment did in either genotype not affect the extent of the typical synaptic fatigue during high frequency synaptic stimulation used for LTP induction, and also did not affect the degree of post-tetanic potentiation (PTP;
Supplementary Figure S1), these data suggest that fingolimod affects LTP expression mechanisms rather than LTP induction processes in our recordings.
Together, these electrophysiological data suggest that Schaffer collateral—CA1 LTP is completely rescued in APP/PS1 mice by the identical regime of fingolimod treatment that also rescues spine deficits.
2.3. Effect of Chronic Fingolimod (FTY720) Treatment on Spatial Memory in Six- to Seven-Month-Old APP/PS1 Mice
Hippocampus-dependent memory was tested in a Morris water maze spatial memory task (MWM). In the cue task training, we first tested the ability of the animals to find the platform when it was visible. Under these conditions, the animals from all four groups quickly learned to navigate to the platform and did not show any significant differences in neither the escape latencies nor the average speed, suggesting that vision and navigation properties were not different between the four groups (
Supplementary Figure S1).
In the subsequent test, the platform was submerged at a different position than previously, and cues were placed on the walls to encourage spatial navigation. Since it seemed as if the animals started on day 1 at slightly different levels, we analyzed their behavior in the single trials in more detail (
Supplementary Figure S1D), which did not reveal significant differences between the four groups. The performance across the four training days was analyzed using repeated measures (RM) ANOVA. Significant main effects were observed for the factors genotype (
F (1, 65) = 20.1
p < 0.001) and training day (
F (3, 63) = 16.25
p < 0.001), but not for the factor treatment (1, 65) = 0.9,
p = 0.34), and there were no significant interactions of these factors (
F’s ≤ 1.3,
p’s ≥ 0.3). These results indicate that there was no significant difference in training performance between the four groups.
Since AD vehicle animals seemed to show less improvement of performance during training sessions than the other three groups, we next analyzed the Δ(D1,D4)-scores (i.e., distance travelled to reach the platform on day 1 minus the distance travelled on day 4) for all animals in the four groups. (
Supplementary Figure S5). However, an ANOVA analysis did not reveal statistical significance for the factors genotype, treatment as well as for the interaction of these factors (
F’s ≤ 2.9,
p’s ≥ 0.09). Thus, we did not observe a significantly impaired performance in AD mice during the water maze training.
When the platform was removed and the animals were tested for long-term memory 24 h later, the RM ANOVA revealed a significant main effect of the factor quadrant (
F (1, 65) = 60.09,
p < 0.0001) with no main effects of the factors genotype and treatment as well as of the interactions of these factors (
F’s ≤ 1.4,
p’s ≥ 0.24). Tukey post hoc comparisons revealed that fingolimod (FTY720) treated APP/PS1 animals showed accurate discrimination between the target and the other quadrants. In detail, fingolimod treated APP/PS1 mice spent significantly more time in the target quadrant (T) than in the averaged three nontarget compartments (%time AD FTY720 target: 40.167 ± 4.105; nontarget: 19.94 ± 1.37,
p = 0.004,
Figure 3D). In contrast, vehicle-treated APP/PS1 mice failed to show a significant preference for the target quadrant (%time AD vehicle target: 34.13 ± 3.33; nontarget: 21.95 ± 1.10,
p = 0.30,
Figure 3D). As expected, both, vehicle and fingolimod treated WT littermates, occupied the target quadrant significantly longer than the nontarget compartments (
p’s < 0.001), as is also evident from the heat maps shown in
Figure 3E.
Taken together, since we did not observe a clear deficit in Morris water maze learning in the AD mice, we could not prove a beneficial effect of chronic FTY720 treatment for memory improvement. However, our statistical analysis revealed that chronic FTY720 treatment of AD mice resulted in a significant preference for the previous target quadrant, which was absent in AD vehicle mice. This might be taken as an indication of potential positive effects of FTY720 on rescuing impaired memory functions of AD mice.
2.4. Chronic Fingolimod (FTY720) Treatment Reduces Activation of Microglia and Astrogliosis in APP/PS1 Mice
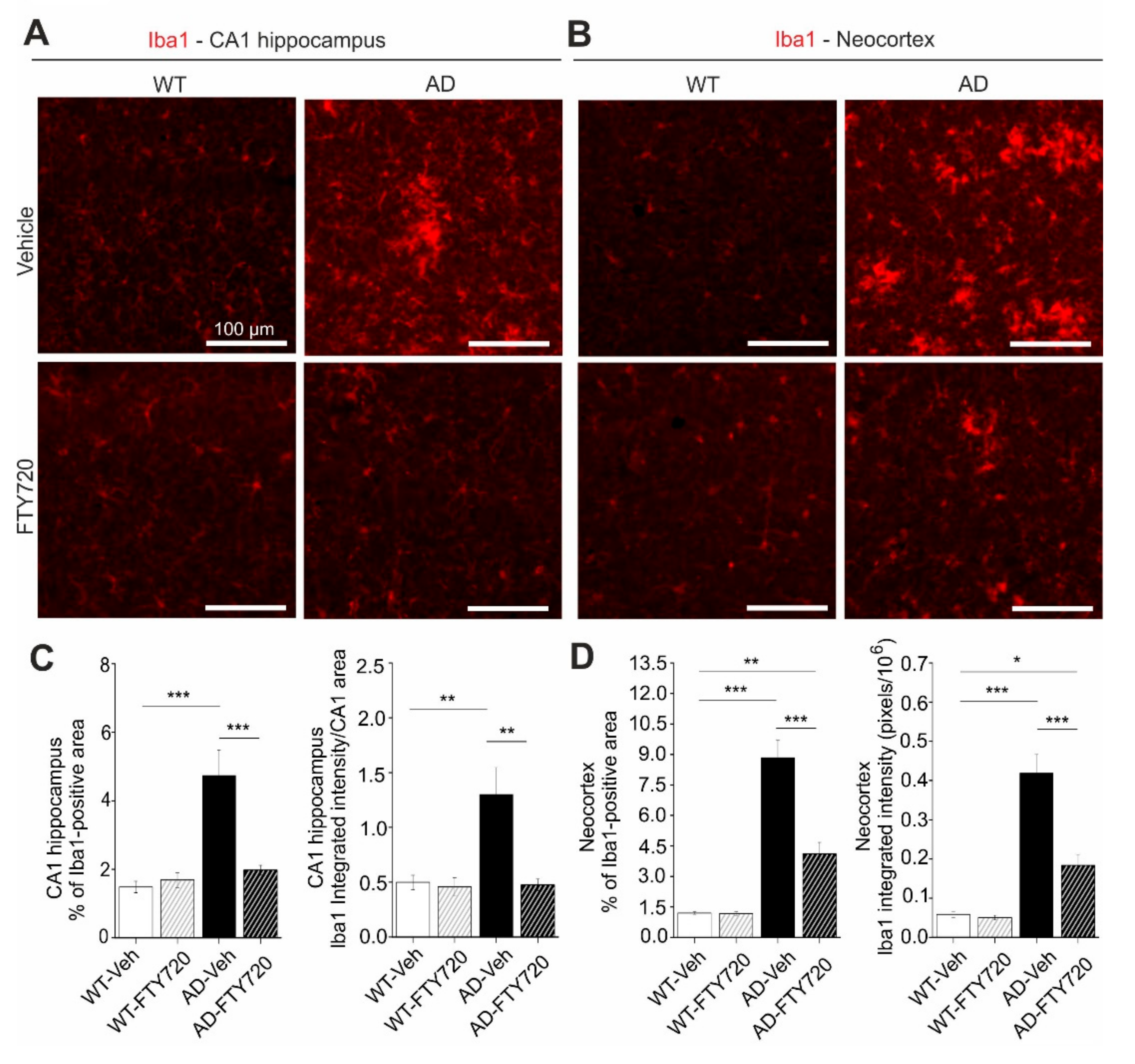
Since fingolimod was shown previously to affect microglia signaling in mouse models of AD [
24], we used anti-ionized calcium-binding adapter molecule 1 (Iba1) immunohiostological stainings to investigate microglia proliferation/growth, and the extent of microglia activation in brain slices from APP/PS1 and WT littermate mice (
Figure 4).
Here, two-way ANOVA analysis revealed a significant main effect of genotype x treatment interaction (
F (1, 21) = 14.46
p = 0.0010). In vehicle-treated APP/PS1 mice, the percentage of the hippocampal CA1 area covered by Iba1 positive microglia was increased threefold compared to WT littermates (AD vehicle: 4.73 ± 0.73%, WT vehicle: 1.48 ± 0.17%), reflecting growth and/or proliferation of microglial cells (
Figure 4C, left). Fingolimod (FTY720) treatment (for one to two months, starting at five to six months of age) reduced the area of Iba1-positive microglia back to control levels (AD + FTY720: 1.97 ± 0.14%, WT + FTY720: 1.69 ± 0.22%), indicating complete rescue from microgliosis. The integrated intensity of Iba1 staining divided by the analyzed CA1 area (i.e., normalized Iba1 intensity;
Figure 4C, right) is a measure of both the area covered by microglia and the intensity of Iba1 expression in these cells. Also here, a two-way ANOVA revealed a significant main effect of genotype x treatment interaction (
F (1, 21) = 8.565
p = 0.0081). The observed 2.5-fold increase in this normalized Iba1 intensity in vehicle treated APP/PS1 mice compared to WT littermates (WT vehicle: 0.50 ± 0.07, WT + FTY720: 0.46 ± 0.08, AD vehicle: 1.30 ± 0.25, AD + FTY720: 0.48 ± 0.05) was similar as compared to the increase in microglia area (threefold increase;
Figure 4C, left). Together these data suggest strongly increased microglial coverage of CA1 in untreated APP/PS1 mice, without an additional effect on the Iba1 expression level per microglial cell area. This finding is corroborated by the similar mean anti-Iba1 fluorescence intensity values observed for all four groups (compare
Supplementary Figure S2). Fingolimod treatment reduced also the value for normalized Iba1 intensity back to WT control levels, indicating a complete rescue. Together, both types of quantification suggest that fingolimod is a highly effective negative regulator of microglial coverage in the CA1 area of the hippocampus. Given the rescue of physiological functions of CA1 pyramidal neurons (spines and LTP) and the rescue of hippocampus-dependent learning (MWM) in response to the fingolimod treatment, it might be possible that the strong anti-inflammatory action of fingolimod on microglia may contribute to the rescue of synaptic and memory functions in APP/PS1 mice. Since cortical dysfunction contributes to reduced memory performance in AD, we also tested microglia activation in the neocortex of our APP/PS1 mouse model. We found that the area of Iba1 covered neocortex and the integrated Iba1 intensity in untreated APP/PS1 mice were both affected more severely than in the hippocampus (
Figure 4B,D). Importantly, the fingolimod treatment also dramatically reduced microglia associated neuro-inflammation in this brain area. Two-way ANOVA analysis showed a significant main effect of genotype x treatment interaction for the Iba1-covered area (
F (1, 21) = 22.22,
p = 0.0001), and also for integrated Iba1 intensity (
F (1, 21) = 17.42
p = 0.0004) in the neocortex.
Interaction of astrocytes and microglia regulate neuroinflammatory processes in neurodegeneration. In the light of the strong functional rescue of LTP and spine density by fingolimod observed in this study, we asked whether astrogliosis was reduced in parallel. Using glial fibrillary acidic protein (GFAP) immunohistochemistry (IHC) we found that the percentage of hippocampal CA1 area covered by astrocytes in vehicle injected APP/PS1 mice was increased roughly twofold (WT vehicle: 4.63 ± 0.33, AD vehicle: 8.01 ± 1.32) compared to vehicle injected WT littermate controls (
Figure 5). Strikingly, fingolimod treatment of APP/PS1 mice decreased astrocyte area back to WT control levels (WT + FTY720: 2.73 ± 0.28, AD + FTY720: 2.71 ± 0.37;
Figure 5C, left).
Likewise, the level of GFAP expression per CA1 area (i.e., normalized GFAP intensity) was regulated in the same way (WT vehicle: 1.51 ± 0.13, WT + FTY720: 0.86 ± 0.16, AD vehicle: 2.86 ± 0.47, AD + FTY720: 0.91 ± 0.15;
Figure 5C, right). Two-way ANOVA analysis showed a significant main effect of genotype x treatment interaction for GFAP-covered area (
F (1, 21) = 5.999
p = 0.0232), and also for normalized GFAP intensity (
F (1, 21) = 6.184
p = 0.0214). Together, this indicates reduced astrogliosis in the CA1 area of APP/PS1 mice in response to fingolimod treatment. Similar to the results for Iba1, the expression level of GFAP per astrocyte cell area was not different between the four groups (
Supplementary Figure S2), indicating that GFAP expression levels were not altered by the treatment. Of note, fingolimod showed a trend towards reduced astrogliosis also in the hippocampus of WT animals. Although this effect did not reach significance, this suggests that our regime of fingolimod treatment might regulate basal growth of astroglia under control conditions. Similar to Iba1 we also investigated changes in GFAP in the neocortex and found more dramatic astrogliosis in the neocortex than in CA1 of untreated APP/PS1 mice, which was effectively reduced by a factor of 2.5 upon fingolimod treatment (
Figure 5B,D). Two-way ANOVA analysis showed a significant main effect of genotype x treatment interaction for GFAP covered area (
F (1, 21) = 56.75
p < 0.0001), and also for integrated GFAP intensity (
F (1, 21) = 20.12
p = 0.0002) in the neocortex. Overall, astrogliosis and microgliosis in APP/PS1 mice are both significantly reduced by chronic fingolimod treatment and both processes together might contribute to the rescue of LTP, spine density, and memory formation in our fingolimod-treated APP/PS1 mice.
2.5. Chronic Fingolimod Treatment Reduces Plaque Formation and Amyloid-β (Aβ) Deposits in the Hippocampus of APP/PS1 Mice
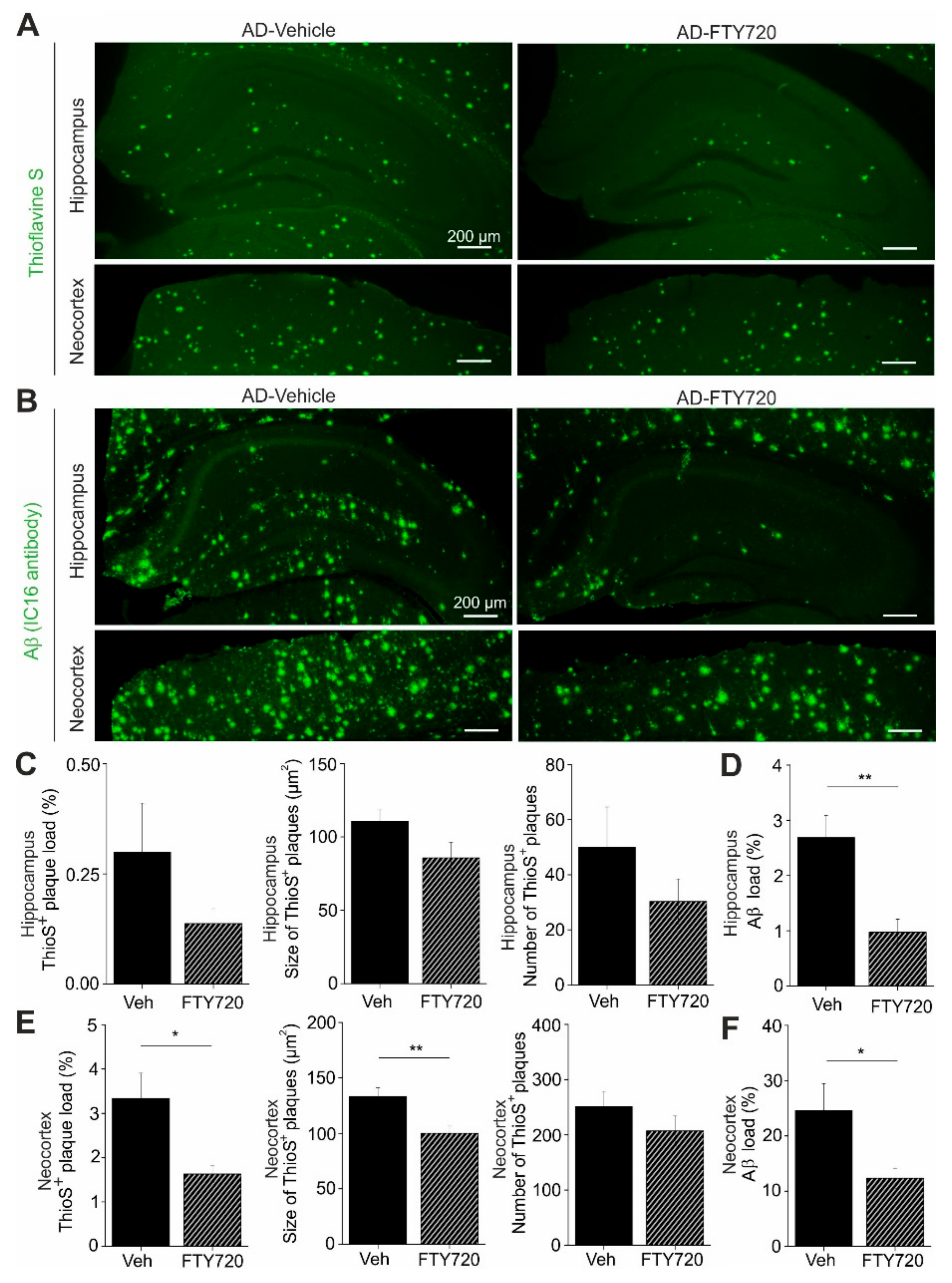
Next, we used Thioflavine S staining to investigate changes in the number and size of Aβ plaques in the hippocampus and in the neocortex of APP/PS1 mice (
Figure 6). We found that the percent area of the hippocampus that was covered by plaques was reduced roughly twofold by fingolimod treatment, although the effect was not reaching significance (AD vehicle: 0.30 ± 0.11, AD + FTY720: 0.14 ± 0.03;
Figure 6C). Smaller and also nonsignificant decreases upon fingolimod treatment were observed for the number of Aβ plaques per unit area, and for the average size of Aβ plaques (
Figure 6C). In the neocortex, plaque load and plaque size were both significantly reduced compared to vehicle treated controls, whereas plaque numbers remained unaffected (
Figure 6E). Overall, this analysis revealed a trend towards reduced Aβ plaque burden in the hippocampus and a significant reduction in the neocortex.
To get further insight into Aβ protein accumulation in treated and untreated animals, we performed, in slices from the identical individual mice as used in the Thioflavine S experiments, anti-Aβ immunohistological stainings, which detect in addition to Aβ plaques also less condensed Aβ protein deposits (including oligomers and fibrils;
Figure 6B). As expected, this analysis yielded higher values for the percent area covered by Aβ positive material (hippocampus: 2.69 ± 0.40%, neocortex: 24.8 ± 5.0%,
Figure 6D,F) than the Thioflavine S staining in vehicle treated APP/PS1 mice (
Figure 6C,E). Following treatment with fingolimod we observed a strong reduction in the percent area covered by Aβ positive structures in both brain areas (
Figure 6D,F).
Overall, these data suggest that the fingolimod induced rescue of functional plasticity and neuroinflammatory signaling in the hippocampus of APP/PS1 mice is accompanied by reduced accumulation of Aβ protein. This reduction of Aβ deposits under fingolimod treatment might be a secondary consequence of the reduced microgliosis we observe, since activated proinflammatory microglia was shown recently to contribute to Aβ plaque load via apoptosis-associated speck-like protein (ASC) specks formation [
25].
Of note, we did not observe any effect of fingolimod treatment on BDNF protein levels or TrkB signaling efficacy in APP/PS1 mice or WT littermates (compare
Supplementary Figure S3).
4. Materials and Methods
4.1. Animals
For all experiments five- to seven-month-old heterozygous APP/PS1 mice and their wild-type littermates were used. These APP/PS1 mice harbor a mutation in the APP gene (
KM670/671NL, “Swedish mutation”) as well as a mutated presenilin 1 (Leu166Pro mutation), both under control of the Thy1 promotor that drives neuron specific expression of the proteins [
17]. Both gene mutations are associated with early-onset familial AD (FAD) in humans. The animals were generated on a C57BL/6J genetic background and constantly backcrossed with C57BL/6J mice (Charles River, Sulzfeld, Germany). The animals were housed in groups of three to four animals and had constant access to food and water. They were maintained on a 12:12 h light dark cycle (lights on at 7 a.m.). All experiments were performed during the light period of the animals and were in accordance with the European Committees Council Directive (2010/63/EU) and were approved by the local animal care committees (APAFIS#6855-20 16091615385487 v5 and 42502-2-1383 UniMD).
4.2. Fingolimod (FTY720) Administration
Male WT and AD transgenic mice were treated with fingolimod (FTY720, Sigma-Aldrich, Lyon, France; Abcam, Cambridge, UK) at the age of 5–6 months (age range of animals at the start of treatment: 22–24 weeks). The drug was dissolved in 3% DMSO (Sigma-Aldrich, Lyon, France) in saline, and was administered every second day via i.p. injection at 1 mg/kg body weight. In another cohort of animals (spine measurements), we also tested the drug at a concentration of 0.2 mg/mL body weight. All animals were treated for one to two months according to this scheme until they were sacrificed for electrophysiological (treatment for four to six weeks), spine (four to five weeks), or immunohistological experiments (four to five weeks), or until completion of behavioral testing (four to six weeks). Electrophysiological experiments, spine analysis, and behavioral testing were done separately in distinct cohorts of animals. Control mice were treated identically with vehicle solution.
4.3. Combined Aβ Plaque Staining and Golgi–Cox Impregnation
All mice were injected twice (at a 24 h interval) i.p. with 75 µL of 10 mg/mL Methoxy-X04 (TOCRIS, Wiesbaden, Germany) in DMSO (Sigma-Aldrich, Taufkirchen, Germany) as described previously [
22,
40]. Two hours after the second injection, they were anesthetized and transcardially perfused with 0.9% saline followed by 4% PFA/PB (pH 7.4). The injection steps were performed according to Jahrling et al. [
40]. The brains were postfixed in 4% PFA/PB pH 7.4 for 24 h at 4 °C and transferred into Golgi–Cox solution in the dark and the solution was changed only once after seven days. After 14 days, the brains were placed into 25% sucrose in PBS at 4 °C for at least one to two days. Coronal sections of 100 μm thickness were cut using a Vibratome (Pelco Model 1000, The Vibratome Company, St. Louis, MO, USA). Sections were mounted onto gelatin-coated slides and after allowing them to dry, they were washed with distilled water for 2 min and then they were transferred into 20% ammonium hydroxide in distilled water (ammonium hydroxide solution, ACS reagent, 28.0–30.0% NH3 basis, Sigma-Aldrich, Taufkirchen, Germany). The sections were washed again with distilled water twice for 2 min each. The sections were dehydrated passing through ascending grades of ethanol 70%, 95% and 100% for 5 min each and cleaned in Xylol (Roth, Karlsruhe, Germany) twice for 10 min each. The sections were coverslipped with DePex medium (Serva) and after letting them dry for two days they stored at 4 °C ready for analysis. The composition for the Golgi–Cox solution was 5% potassium dichromate (Merck, Darmstadt, Germany), 5% mercuric chloride (Merck, Darmstadt, Germany) and 5% potassium chromate (Merck, Darmstadt, Germany; compare [
41]). All stock solutions as well as the final Golgi–Cox solution were prepared according to the protocol used by Bayram-Weston [
42].
4.4. Image Processing and Dendritic Spine Density Analysis
Spine density of secondary apical dendritic segments in stratum radiatum (SR) of CA1 pyramidal neurons was calculated as the number of spines per micrometer dendritic length. Ten dendritic segments (15 µm long) per animal, from 10 different neurons throughout intermediate and ventral hippocampus, were analyzed for all treated and untreated groups. Dendritic lengths were estimated using the NeuronJ plugin of NIH ImageJ software (
https://imagej.nih.gov/ij/). For AD mice, 10 dendritic segments which were at a distance more than 50 µm from the plaque border (AD distant) and 10 dendritic segments which were located within 50 µm from the plaque border (AD near) were analyzed per animal. This digital near/distant classification scheme was selected to facilitate quantification of the results. When plotting spine densities versus plaque distance for individual dendritic segments we did not observe a clear cut-off at a certain distance, but a decrease at distances <50 µm is evident (compare
Figure S4). Images for Golgi–Cox stained dendritic segments and the blue fluorescent methoxy-X04 stained Aβ plaques were captured with a ×40 magnification objective using a SPOT digital camera that was attached to a LEITZ DM R microscope (Leica, Wetzlar, Germany). The distance in z-direction of analyzed dendrites to the border of the Methoxy-X04 stained Aβ plaque was in all cases <5 µm. This deviation was minor compared to the calculated distances in the x-y space and was therefore neglected. Imaging for Golgi–Cox staining was performed in bright field mode while Methoxy-X04 fluorescence was captured with a filter cube (excitation filter: BP 360/40 nm, dichroic mirror: 400 nm, emission filter: LP 425; Leica, Wetzlar, Germany). The distance of the analyzed dendritic segments from the Aβ plaques was determined as the average distance of both segment end-points and the center of the segment to the respective plaque border in merged pictures using ImageJ software (
https://imagej.nih.gov/ij/). Dendritic segments from different neurons were traced and spine density was quantified as the number of spines per dendritic length in µm. The spines were counted manually using a 100× magnification oil immersion objective. Spine density analysis was conducted blindly to the treatment in WT and AD mice.
4.5. Preparation of Slices for IHC
Coronal slices were prepared from young adult male APP/PS1 mice or WT littermates at an age of six to seven months. Animals were anesthetized using intraperitoneal injection of ketamine hydrochloride/xylazine hydrochloride (Sigma-Aldrich, Taufkirchen, Germany) and then transcardially perfused with ice-cold 0.9% NaCl solution. After opening the cranium, the brains were gently removed and the right hemibrains were postfixed in 4% PFA/0.1 M phosphate-buffered saline (PBS) for 24 h at 4 °C and then cryoprotected in 25% sucrose (AppliChem, Darmstadt, Germany) in PBS. The left hemibrains were quickly placed in ice-cold artificial cerebrospinal fluid (ACSF) solution, anterior cortex, posterior cortex and hippocampus were dissected and all brain areas were snap-frozen for protein analysis. The fixed hemispheres were then cut to 40 µm thick coronal slices using a cryostat (Leica CM 3050, Wetzlar, Germany). Three free-floating sections 240 µm apart from each other (between Bregma levels −2.46 mm and −3.08 mm according to Franklin and Paxinos [
43], containing the hippocampus and cortex were selected for all histological fluorescent stainings. For imaging experiments, all sections were transferred to slides (Superfrost, Thermo Scientific, Dreieich, Germany), coverslipped with ImmunoMount mounting medium (Thermo Scientific, Dreieich, Germany).
4.6. Immunofluorescence for Total Aβ
Free-floating sections were permeabilized with PBS/0.1% Triton X-100 and incubated with 98% formic acid to perform antigen retrieval. This step was followed by washes with PBS. Sections were then blocked in 20% normal goat serum (Dianova, Hamburg, Germany) in PBS/Triton X-100 0.1% and incubated overnight at 4 °C with the mouse anti-Aβ IC16 antibody (1:400; kindly provided by Claus Pietrzik, Johannes-Gutenberg-University Mainz [
44] in 10% normal goat serum (Dianova, Hamburg, Germany) in PBS containing 0.1%Triton X-100. Sections were then washed with PBS/0.1%Triton X-100, incubated with an Alexa Fluor 488 labeled goat antimouse antibody (1:500, Thermo Scientific, Dreieich, Germany) in 10% normal goat serum (Dianova, Hamburg, Germany) in PBS/0.1% Triton X-100, and finally washed with PBS. Imaging was performed with ZEN 2010 software using a 5x objective in a confocal imaging system (LSM 780, Zeiss, Jena, Germany). Green fluorescence was excited using the 488 nm laser line from an Argon laser. For Aβ load quantification 16 bit images were converted to 8 bit gray-scale images and after defining the region of interest (total hippocampal or cortical area) they were thresholded within a linear range using the NIH ImageJ software (
https://imagej.nih.gov/ij/). The load was expressed as the percentage area covered by Aβ-positive staining (% Aβ load). The quantification performed blind to the treatment for WT and AD mice. The quantification was performed blind to the treatment for WT and AD mice.
4.7. Immunofluorescence for GFAP
For GFAP staining, antigen retrieval was performed with sodium citrate buffer (10 mM, pH 6.0) for 30 min at 80 °C followed by three washes with TBS. Sections were blocked in 5% normal goat serum (Dianova, Hamburg, Germany) in TBS with 0.4% Triton X-100 and incubated with a mouse anti-GFAP antibody (1:500, clone G-A-G, Sigma: G 3893) in blocking solution overnight at 4 °C. Sections were washed with TBS and incubated with an Alexa Fluor 488 labeled goat antimouse antibody (1:500, Thermo Scientific, Dreieich, Germany) in blocking solution and were finally washed with TBS. For the quantification of GFAP in the CA1 area of the hippocampus and neocortex images were captured with a 10× objective using a SPOT camera attached to a Leica LEITZ DM R microscope (Leica, Wetzlar, Germany) through an appropriate filter cube (excitation filter: BP 515–560 nm, dichroic mirror: 580 nm, emission filter: LP 590 nm, Leica, Wetzlar, Germany). Using NIH ImageJ software (
https://imagej.nih.gov/ij/) for quantification, the GFAP load was expressed as the percentage area covered by GFAP-positive staining, and the normalized integrated intensity (raw integrated density divided by the area of the analyzed CA1 area) or the integrated intensity (for identical sized neocortex pictures) were expressed in pixels (pixels/10
6). The quantification was performed blind to the treatment for WT and AD mice.
4.8. Immunofluorescence for Iba1
Free-floating sections were first treated with sodium citrate buffer (10 mM, pH 6.0) for 30 min at 80 °C for antigen retrieval and then washed three times with PBS. Sections were blocked in 10% FBS (Gibco, Dreieich, Germany) and 1% BSA (Sigma, Taufkirchen, Germany) in PBS containing 0.3% Triton X-100 and incubated overnight at 4 °C with a rabbit anti-Iba1 antibody (1:500, Wako, Richmond, VA, USA) in PBS containing 1% FBS (Gibco, Dreieich, Germany), 0.1% BSA (Sigma, Taufkirchen, Germany) and 0.3% Triton X-100. Sectionswere then washed with PBS and developed with an Alexa Fluor 555 labeled donkey antirabbit antibody (1:500, Thermo Scientific, Dreieich, Germany) in PBS containing 1% FBS, 1% BSA, and 0.3% Triton X-100. The sections were finally washed with PBS. Imaging and quantification were performed as described for GFAP immunohistochemistry. The quantification was performed blind to the treatment for WT and AD mice.
4.9. Thioflavine S Staining
Coronal free-floating sections were incubated for 9 min in 1% Thioflavine S (Sigma-Aldrich) aqueous solution and then treated with 80% ethanol two times for 3 min each, followed by a wash with 95% ethanol for 3 min. Sections were rinsed three times with distilled water. Thioflavine S imaging was performed with a confocal Zeiss laser-scanning-microscope using Zen software (LSM 780, Zeiss, Jena, Germany). A 5× magnification objective was used and green fluorescence was excited with a 488 nm Argon laser. Primary images were converted to 8 bit gray-scale and after defining the region of interest (total hippocampal or cortical area) they were thresholded using NIH ImageJ software (
https://imagej.nih.gov/ij/). Thioflavine S plaque load was expressed as the percentage area covered by Thioflavine S-positive staining (% ThioS). The ImageJ tool ‘Analyze particles’ was used to determine plaque number and size (plaque area in µm
2). The quantification was performed blind to the treatment for WT and AD mice.
4.10. Western Blotting
The hippocampal brain samples from WT and APP/PS1 mice chronically treated with fingolimod or vehicle were homogenized using an ultrasound sonicator in NP lysis buffer containing 2% orthovanadate and 4% protease inhibitor mix. The homogenized suspension was centrifuged (15,000× g, 10 min, +4 °C), and the resulting supernatant was used for analysis. Proteins were separated with SDS-PAGE and transferred to Polyvinylidenfluorid (PVDF) membrane. The membranes were incubated with antibodies against phosphorylated TrkA/TrkB at residues Y515, (C35G9, Cell Signaling Technology, Danvers, MA, USA #4619), Y706 (Cell Signaling Technology, #4621), Y816 (Cell Signaling Technology #4168), total TrkB (R&D Systems, Minneapolis, MN, USA, #AF1494), phospho-p70 S6 Kinase (Cell Signaling Technology #9205), total p70 S6 Kinase (Cell Signaling Technology #9202), and β-Actin (Sigma Aldrich, St. Louis, MO, USA, #A1978). Following incubation with primary antibodies, the membranes were incubated with horseradish peroxidase conjugated secondary antibodies, and the bands were visualized using chemiluminescent Western blotting substrate with a Fuji LAS3000 camera (Tamro Medlab, Vantaa, Finland). Bands obtained with antibodies against phosphorylated tyrosines in TrkB were normalized after reblotting against bands corresponding to total TrkB in the same lanes. Levels of total TrkB were normalized against beta-actin in the same lane.
4.11. BDNF Quantification
To assess the levels of hippocampal BDNF protein the BDNF Quantikine ELISA kit (R&D Systems, Wiesbaden, Germany) was used. Total hippocampi were dissected and processed according to the manufacturer’s instructions (compare [
45]). Protein levels were set in relation to the wet weight of the brain tissue sample.
4.12. Preparation and LTP Recordings of Acute Hippocampal Slices
Acute transverse hippocampal slices (350 µm thick for field recordings) were obtained from APP/PS1 and WT mice. Animals were first anaesthetized with isoflurane and killed in accordance with the European Communities Council Directive (80/609/EEC). Slices were cut on a vibratome (Microm HM600V, Thermo Scientific, Illkirch, France) in ice-cold dissecting solution containing (in mM): 234 sucrose, 2.5 KCl, 0.5 CaCl2, 10 MgCl2, 26 NaHCO3, 1.25 NaH2PO4 and 11 D-glucose, oxygenated with 95% O2 and 5% CO2, pH 7.4. Slices were first incubated, for 60 min at 37 °C, in artificial cerebrospinal fluid (ACSF) solution containing (in mM): 119 NaCl, 2.5 KCl, 1.25 NaH2PO4, 26 NaHCO3, 1.3 MgSO4, 2.5 CaCl2 and 11 D-glucose, oxygenated with 95% O2 and 5% CO2, pH 7.4. Slices were used after recovering for another 30 min at room temperature. For all experiments, slices were perfused with the oxygenated ACSF at 31 ± 1 °C. Field EPSPs were recorded in the stratum radiatum of the CA1 region using a glass electrode (RE) (filled with 1 M NaCl, 10 mM HEPES, pH 7.4) and the stimuli (30% of maximal fEPSP) were delivered to the Schaeffer Collateral pathway by a monopolar glass electrode (SE) (filled with ACSF). The recordings were performed using a Multiclamp 700B (Molecular Devices, San Jose, CA, USA) amplifier, under the control of pClamp10 software (Molecular Devices, San Jose, CA, USA).
LTP was induced using a high-frequency stimulation protocol (HFS) protocol with two pulses of 100 Hz spaced by 20 s inter-stimulus interval. For LTP analysis, the first third of the fEPSP slope was calculated in baseline condition (20 min prior to induction protocol delivery and for 45–60 min postinduction). The average baseline value was normalized to 100% and all values of the experiment were normalized to this baseline average (one minute bins). The recordings were performed blind to the treatment for WT and AD mice.
4.13. Morris Water Maze
Spatial learning and memory were tested with a Morris water maze paradigm. The apparatus consisted of a circular tank (Ø 90 cm) filled with water (temperature 23 ± 1 °C) made opaque with the addition of 100 mL of white opacifier (Viewpoint, Lyon, France). The test consisted of three phases: (1) cue task training (two days), (2) spatial learning training (four days) and (3) long-term reference memory-probe test (one day). Cued task was done prior to spatial learning training to detect visual and motor problems and to accustom the mice to the testing rule (find the platform to escape). A visible flag was placed on top of the platform and the maze was surrounded by opaque curtains. The mice were allowed to find the visible platform. The platform position was changed for the second cue task (day 2). The escape latency, average speed and distance travelled were recorded. For the spatial learning, the extra-maze cues were mounted on the side-walls. A new platform position was chosen and kept in the same position for the four training days. If the animal did not find the platform within the trial duration it was gently guided to it. For the cue task and spatial trainings, all animals performed four trials/day with a maximum trial duration of 90 s (+30 s on the platform at the end of each trial) and an intertrial interval of 10 min. Twenty-four hours after training completion, the platform was removed and a probe test was run for 60 s. All the trials were video recorded and tracked using ANYmaze software (Stoelting Europe, Dublin, Ireland). The distance travelled to reach the platform for each trial was analyzed. During the probe test, the distance travelled in each quadrant (target (previous position of the platform), left, right and opposite) was recorded and analyzed.
4.14. Statistical Analysis
Statistical analysis was performed by using GraphPad Prism 8.4.3 (GraphPad Software Inc., La Jolla, San Diego, CA, USA) software. Spine densities and quantification of IHC stainings were analyzed either by two-sided t-test comparisons or, in case of multiple comparisons, by one-way or two-way ANOVA (analysis of variance), as indicated. For LTP and behavioral analyses, two-way ANOVA or repeated measures (RM) two-way ANOVA was applied followed by Tukey’s post hoc comparisons when appropriate. All data were checked for normal distribution by using the Shapiro–Wilk test. If not otherwise indicated, all data followed the normal distribution. Statistical significance was determined as p < 0.05. All data are depicted as mean ± standard error of the mean (SEM).


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





