Scalable Microgravity Simulator Used for Long-Term Musculoskeletal Cells and Tissue Engineering

 , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
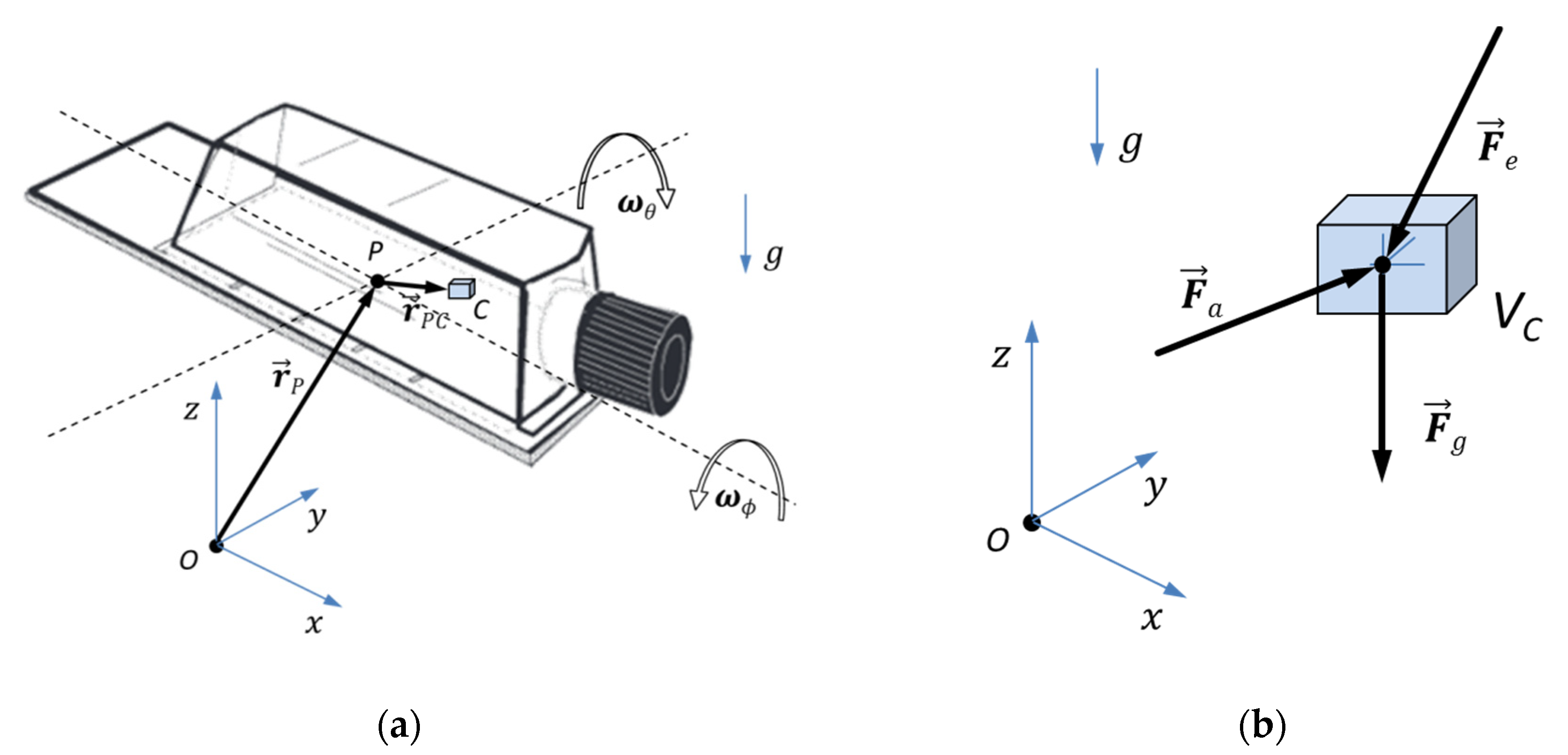
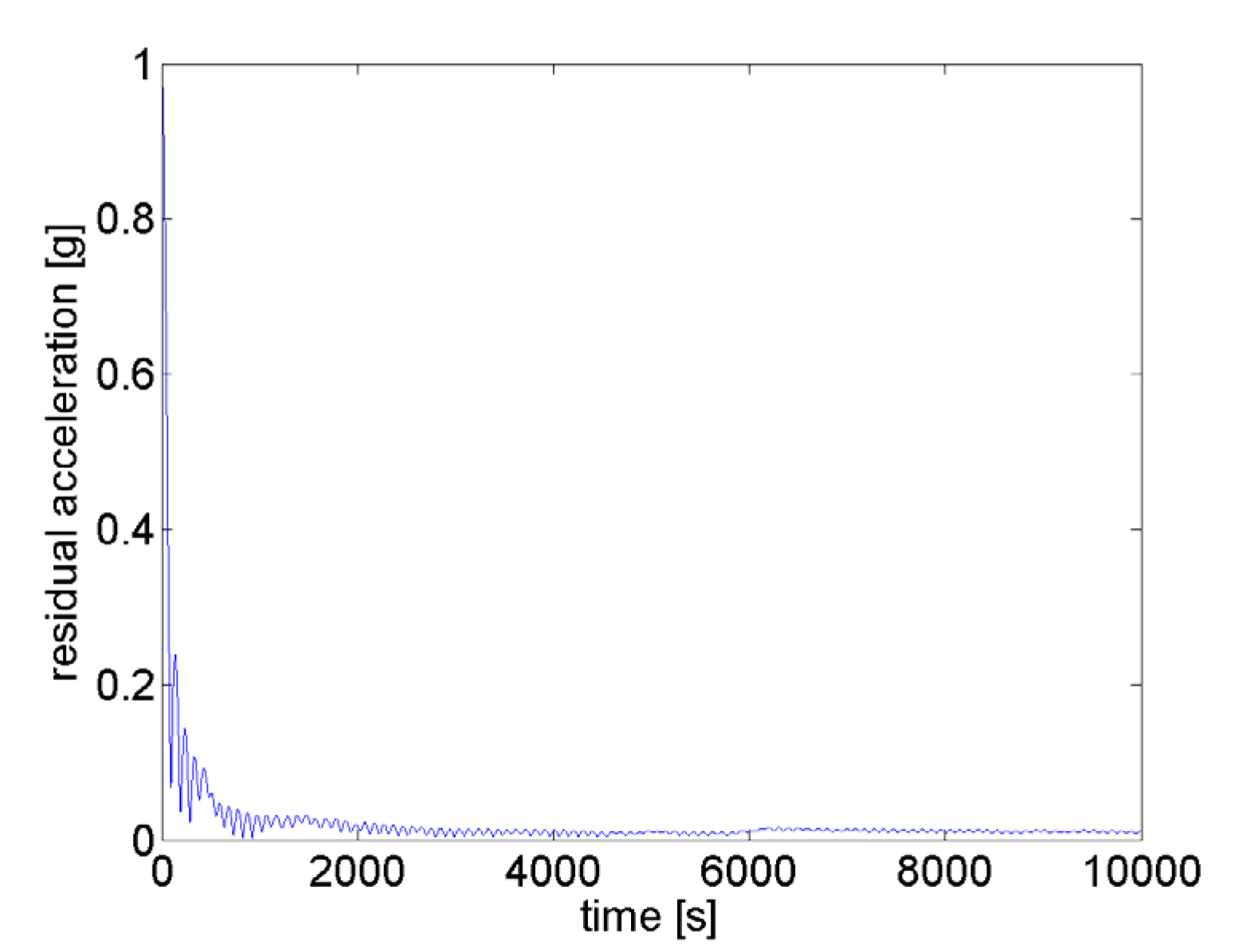
2.1. Description of the MGS
- Increased availability of a simulated microgravity environment by a simple desktop machine
- Down-sizing for the use in typical lab environments (e.g., fitting into CO2 cell incubators)
- Reproducible and optimized motion control (uniformity and nullification)
- Possibility to use and analyze several samples in parallel
- Scalable design (fulfilling the needs of users)
- Identical treatment of all samples (equality)
- Interface commonly used in cell cultures
- Simplified LabVIEW interface that allows an easy control of all the MGS functions.
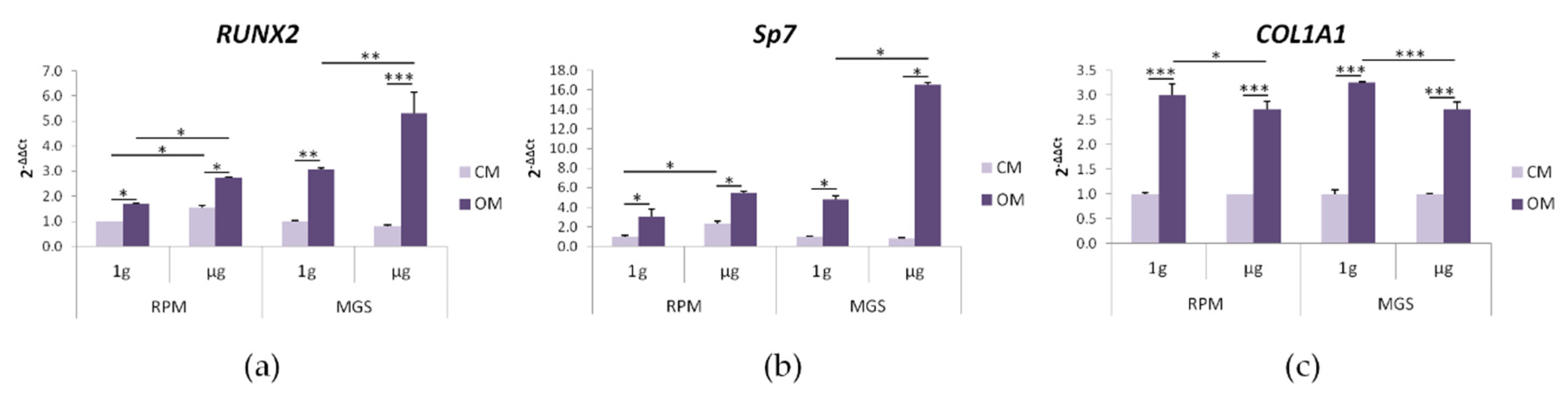
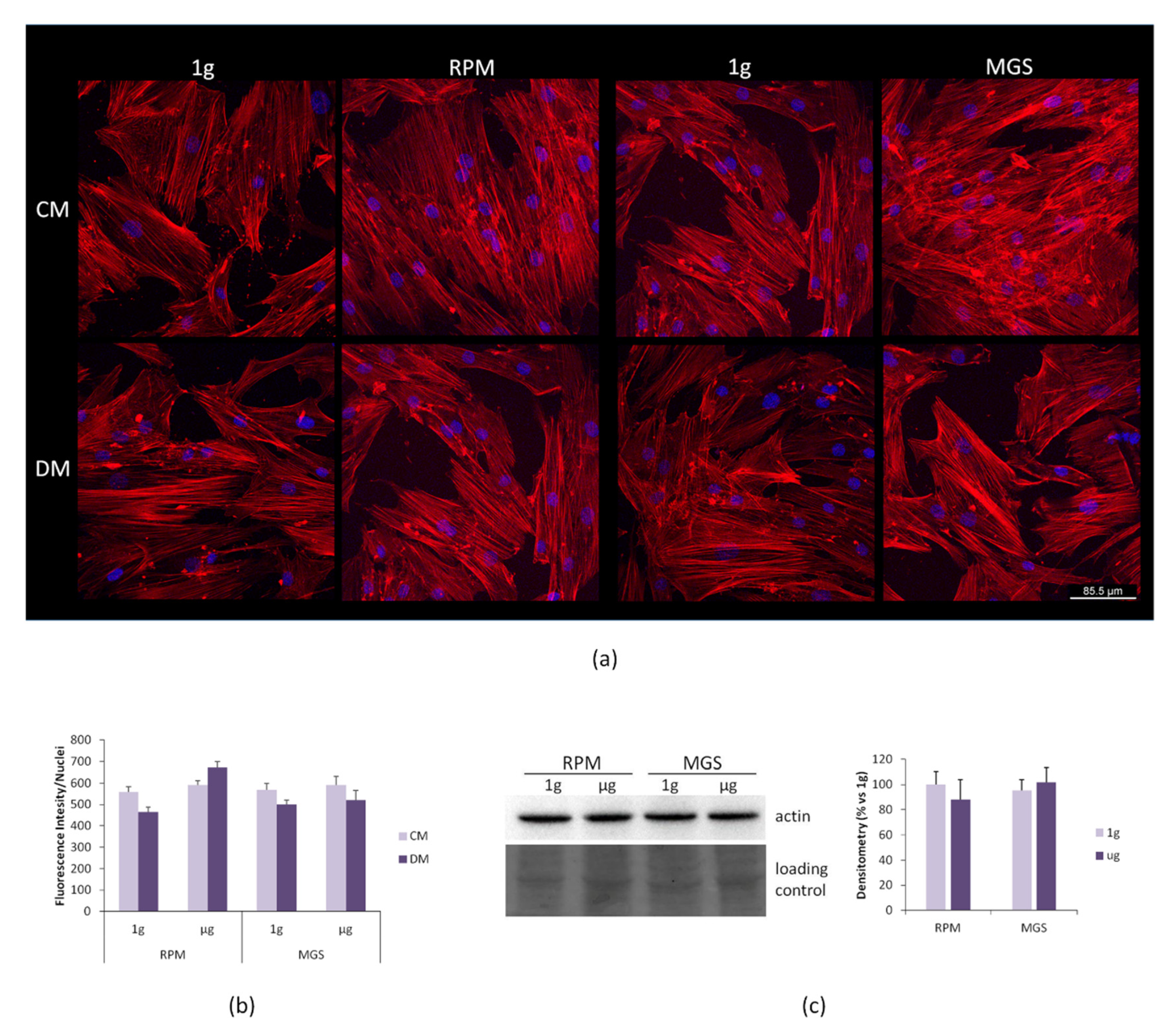
2.2. bMSC: Effects of Culture in MGS vs. RPM
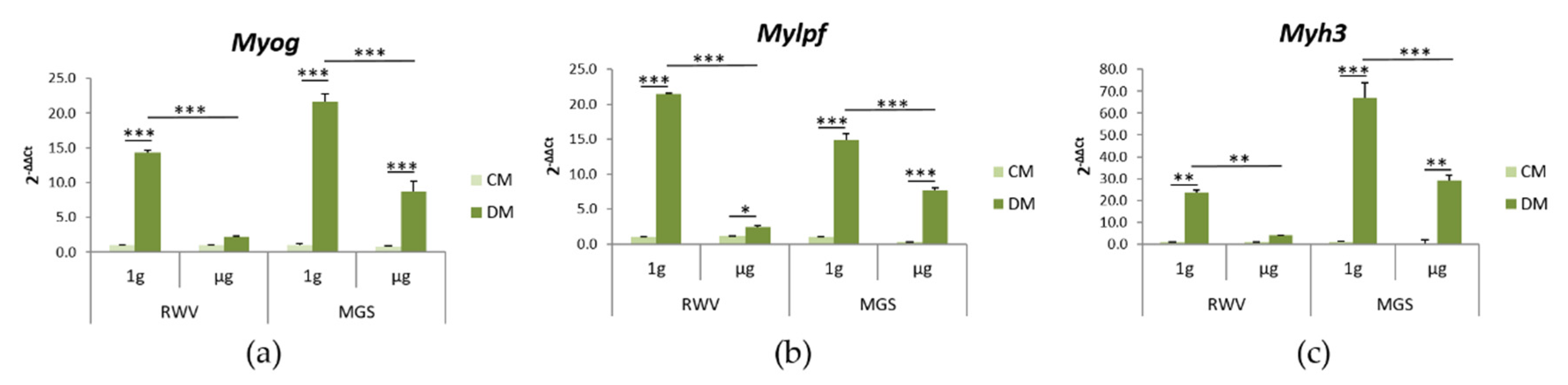
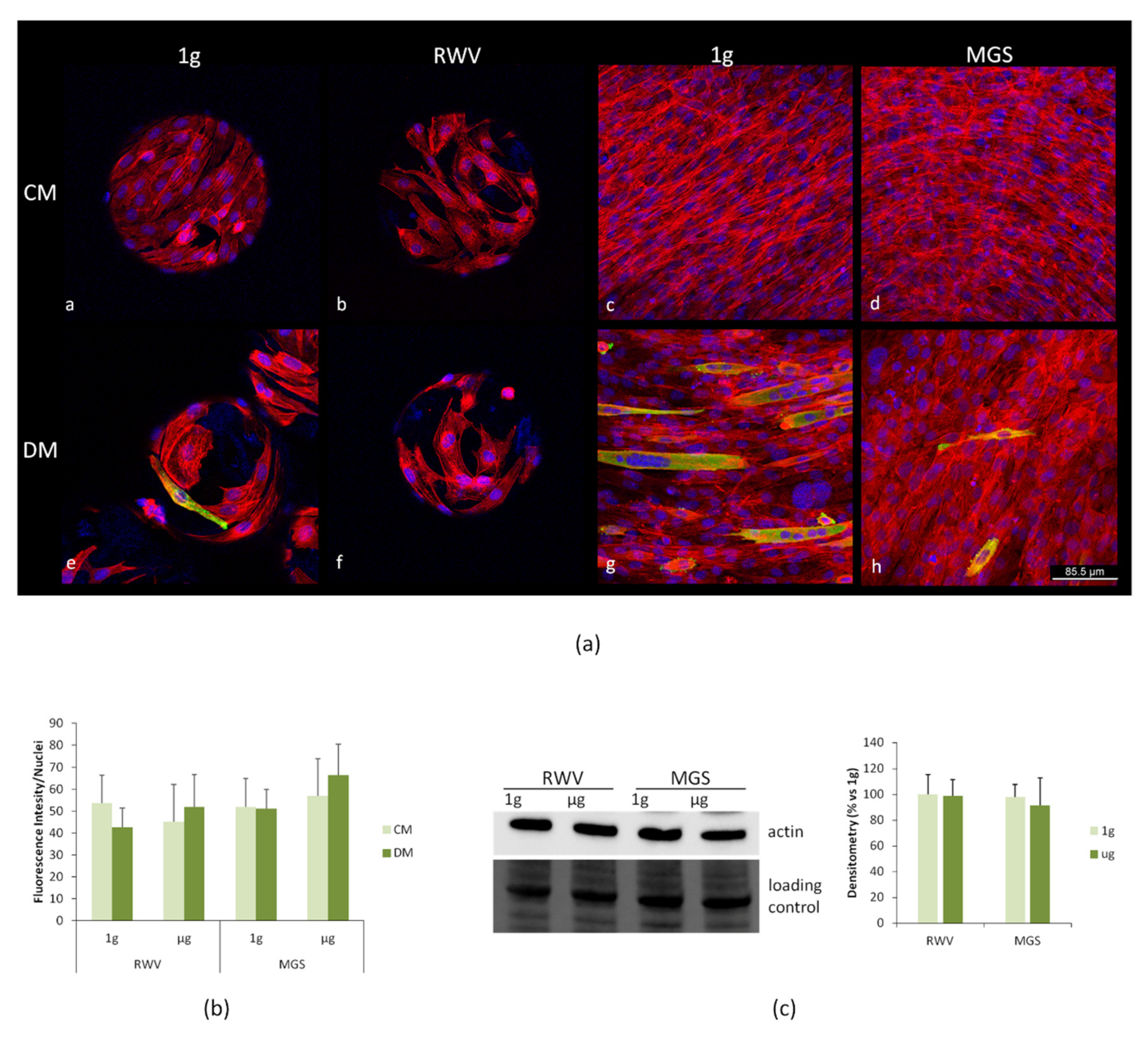
2.3. C2C12 Myoblasts: Effects of Culture in MGS vs. RWV
3. Discussion
4. Materials and Methods
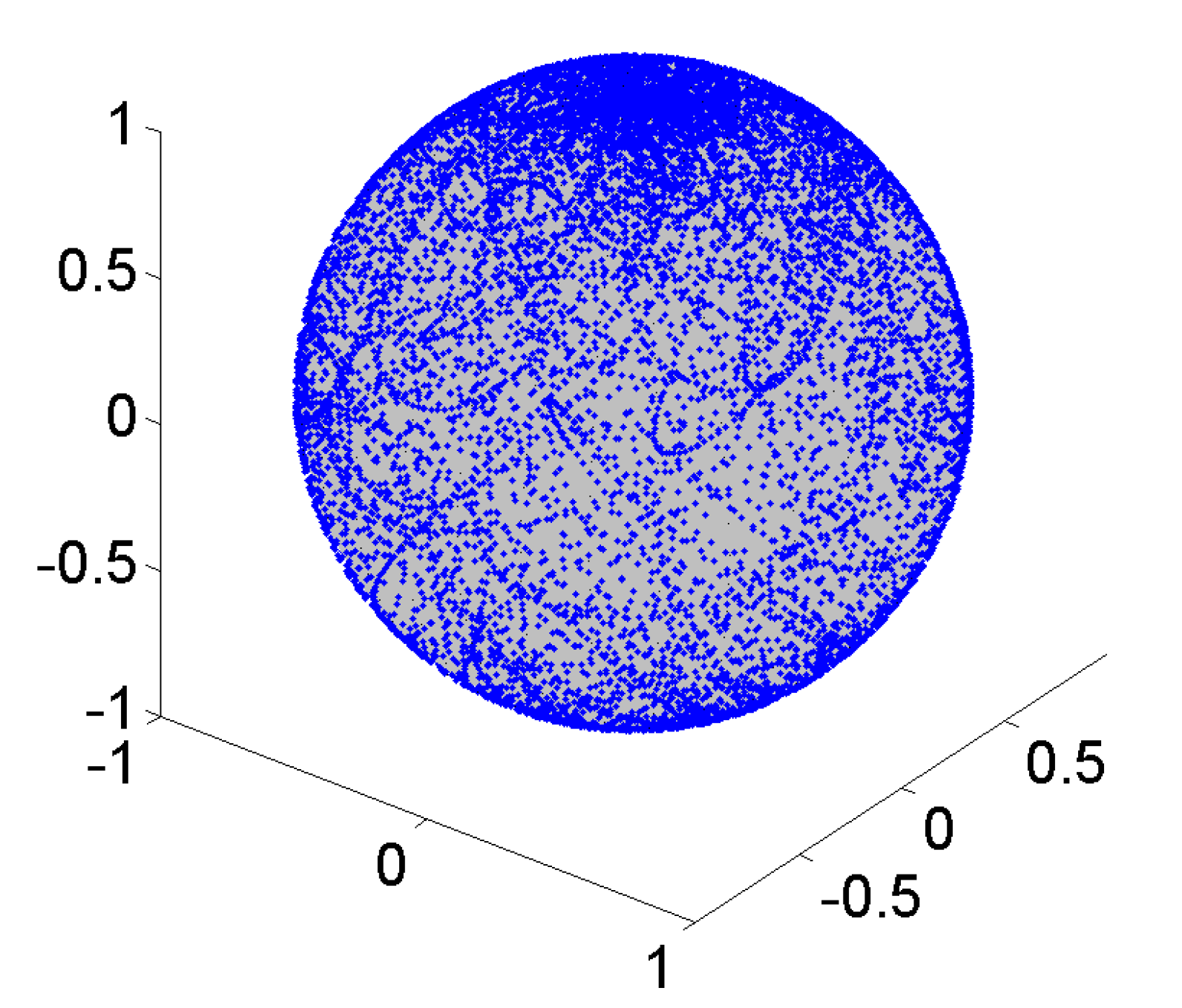
4.1. Description of Algorithm Used to Simulate Microgravity
- (1)
- a uniform path distribution with low uniformity factor F* for zero gravity application (for partial gravity, a controlled concentration of path lines is required)
- (2)
- in-advance (MATLAB) computer simulation of motion path, (not-randomly) control parameters, and statistical measures before the machine is utilized for biological tests.
4.2. Cell Culture
4.3. Real-Time PCR
4.4. Confocal Imaging
4.5. Western Blot
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| MGS | Microgravity simulator |
| RWV | Rotating Wall Vessel |
| RPM | Random Positioning Machine |
| CM | Culture Medium |
| OM | Osteogenic Medium |
| RUNX2 | Runt-related transcription factor 2 |
| COL1A1 | Collagen Type I Alpha 1 Chain |
| RT-PCR | Real Time-PCR |
| DM | Myogenic Medium |
| Myog | Myogenic Factor 4 |
| Mylpf | Myosin Light Chain 2 |
| Myh3 | Myosin Heavy Chain 3 |
| MHC | Myosin heavy chain |
| Akt | Protein Kinase B |
| FOXO1 | Forkhead Box O1 |
References
- Unsworth, B.R.; Lelkes, P.I. Growing tissues in microgravity. Nat. Med. 1998, 4, 901–907. [Google Scholar] [CrossRef]
- Mazzoleni, G.; Di Lorenzo, D.; Steimberg, N. Modelling tissues in 3D: The next future of pharmaco-toxicology and food research? Genes Nutr. 2009, 4, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Radtke, A.L.; Herbst-Kralovetz, M.M. Culturing and applications of rotating wall vessel bioreactor derived 3D epithelial cell models. J. Vis. Exp. 2012, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Grimm, D.; Egli, M.; Kruger, M.; Riwaldt, S.; Corydon, T.J.; Kopp, S.; Wehland, M.; Wise, P.; Infanger, M.; Mann, V.; et al. Tissue Engineering Under Microgravity Conditions-Use of Stem Cells and Specialized Cells. Stem Cells Dev. 2018, 27, 787–804. [Google Scholar] [CrossRef] [PubMed]
- Cazzaniga, A.; Locatelli, L.; Castiglioni, S.; Maier, J.A.M. The dynamic adaptation of primary human endothelial cells to simulated microgravity. FASEB J. 2019, 33, 5957–5966. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, L.; Cazzaniga, A.; De Palma, C.; Castiglioni, S.; Maier, J.A.M. Mitophagy contributes to endothelial adaptation to simulated microgravity. FASEB J. 2020, 34, 1833–1845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cazzaniga, A.; Castiglioni, S.; Maier, J.A.M. Conditioned media from microvascular endothelial cells cultured in simulated microgravity inhibit osteoblast activity. BioMed Res. Int. 2014, 2014, 857934. [Google Scholar] [CrossRef]
- Wuest, S.L.; Richard, S.; Walther, I.; Furrer, R.; Anderegg, R.; Sekler, J.; Egli, M. A Novel Microgravity Simulator Applicable for Three-Dimensional Cell Culturing. Microgravity Sci. Technol. 2014, 26, 77–88. [Google Scholar] [CrossRef]
- Van Loon, J.J.W.A. Some history and use of the random positioning machine, RPM, in gravity related research. Adv. Space Res. 2007, 39, 1161–1165. [Google Scholar] [CrossRef]
- Wuest, S.L.; Richard, S.; Kopp, S.; Grimm, D.; Egli, M. Simulated microgravity: Critical review on the use of random positioning machines for mammalian cell culture. BioMed Res. Int. 2015, 2015, 971474. [Google Scholar] [CrossRef] [Green Version]
- Maier, J.A.M.; Cialdai, F.; Monici, M.; Morbidelli, L. The impact of microgravity and hypergravity on endothelial cells. BioMed Res. Int. 2015, 2015, 434803. [Google Scholar] [CrossRef] [PubMed]
- Herranz, R.; Anken, R.; Boonstra, J.; Braun, M.; Christianen, P.C.M.; De Geest, M.; Hauslage, J.; Hilbig, R.; Hill, R.J.A.; Lebert, M.; et al. Ground-based facilities for simulation of microgravity: Organism-specific recommendations for their use, and recommended terminology. Astrobiology 2013, 13, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aleshcheva, G.; Bauer, J.; Hemmersbach, R.; Slumstrup, L.; Wehland, M.; Infanger, M.; Grimm, D. Scaffold-free Tissue Formation Under Real and Simulated Microgravity Conditions. Basic Clin. Pharmacol. Toxicol. 2016, 119, 26–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuest, S.L.; Stern, P.; Casartelli, E.; Egli, M. Fluid dynamics appearing during simulated microgravity using Random Positioning Machines. PLoS ONE 2017, 12, 1–19. [Google Scholar] [CrossRef]
- Sargenti, A.; Castiglioni, S.; Olivi, E.; Bianchi, F.; Cazzaniga, A.; Farruggia, G.; Cappadone, C.; Merolle, L.; Malucelli, E.; Ventura, C.; et al. Magnesium deprivation potentiates human mesenchymal stem cell transcriptional remodeling. Int. J. Mol. Sci. 2018, 19, 1410. [Google Scholar] [CrossRef] [Green Version]
- Cazzaniga, A.; Maier, J.A.M.; Castiglioni, S. Impact of simulated microgravity on human bone stem cells: New hints for space medicine. Biochem. Biophys. Res. Commun. 2016, 473, 181–186. [Google Scholar] [CrossRef]
- Di Prampero, P.E.; Narici, M.V. Muscles in microgravity: From fibres to human motion. J. Biomech. 2003, 36, 403–412. [Google Scholar] [CrossRef]
- Winnard, A.; Scott, J.; Waters, N.; Vance, M.; Caplan, N. Effect of Time on Human Muscle Outcomes During Simulated Microgravity Exposure Without Countermeasures—Systematic Review. Front. Physiol. 2019, 10, 1046. [Google Scholar] [CrossRef] [Green Version]
- Baek, M.O.; Ahn, C.B.; Cho, H.J.; Choi, J.Y.; Son, K.H.; Yoon, M.S. Simulated microgravity inhibits C2C12 myogenesis via phospholipase D2-induced Akt/FOXO1 regulation. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Calzia, D.; Ottaggio, L.; Cora, A.; Chiappori, G.; Cuccarolo, P.; Cappelli, E.; Izzotti, A.; Tavella, S.; Degan, P. Characterization of C2C12 cells in simulated microgravity: Possible use for myoblast regeneration. J. Cell. Physiol. 2020, 235, 3508–3518. [Google Scholar] [CrossRef]
- Faralli, H.; Dilworth, F.J. Turning on myogenin in muscle: A paradigm for understanding mechanisms of tissue-specific gene expression. Comp. Funct. Genom. 2012, 2012, 836374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiaffino, S.; Rossi, A.C.; Smerdu, V.; Leinwand, L.A.; Reggiani, C. Developmental myosins: Expression patterns and functional significance. Skelet. Muscle 2015, 5, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torgan, C.E.; Burge, S.S.; Collinsworth, A.M.; Truskey, G.A.; Kraus, W.E. Differentiation of mammalian skeletal muscle cells cultured on microcarrier beads in a rotating cell culture system. Med. Biol. Eng. Comput. 2000, 38, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.; Kuipers, A.; Mukai, C.; Thirsk, R. Acclimation during space flight: Effects on human physiology. Cmaj 2009, 180, 1317–1323. [Google Scholar] [CrossRef] [Green Version]
- Cappellesso, R.; Nicole, L.; Guido, A.; Pizzol, D. Spaceflight osteoporosis: Current state and future perspective. Endocr. Regul. 2015, 49, 231–239. [Google Scholar] [CrossRef]
- Teodori, L.; Costa, A.; Campanella, L.; Albertini, M.C. Skeletal muscle atrophy in simulated microgravity might be triggered by immune-related microRNAs. Front. Physiol. 2019, 10, 1–6. [Google Scholar] [CrossRef]
- Grimm, D.; Wehland, M.; Pietsch, J.; Aleshcheva, G.; Wise, P.; Van Loon, J.; Ulbrich, C.; Magnusson, N.E.; Infanger, M.; Bauer, J. Growing tissues in real and simulated microgravity: New methods for tissue engineering. Tissue Eng. Part B Rev. 2014, 20, 555–566. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, T.; Tanimoto, K.; Fukazawa, T.; Imura, T.; Kawahara, Y.; Yuge, L. Simulated microgravity attenuates myogenic differentiation via epigenetic regulations. Npj Microgravity 2018, 4, 1–8. [Google Scholar] [CrossRef]
- Yuge, L.; Kajiume, T.; Tahara, H.; Kawahara, Y.; Umeda, C.; Yoshimoto, R.; Wu, S.-L.; Yamaoka, K.; Asashima, M.; Kataoka, K.; et al. Microgravity potentiates stem cell proliferation while sustaining the capability of differentiation. Stem Cells Dev. 2006, 15, 921–929. [Google Scholar] [CrossRef]
- Sakai, S.; Mishima, H.; Ishii, T.; Akaogi, H.; Yoshioka, T.; Ohyabu, Y.; Chang, F.; Ochiai, N.; Uemura, T. Rotating three-dimensional dynamic culture of adult human bone marrow-derived cells for tissue engineering of hyaline cartilage. J. Orthop. Res. 2009, 27, 517–521. [Google Scholar] [CrossRef]
- Zayzafoon, M.; Gathings, W.E.; McDonald, J.M. Modeled microgravity inhibits osteogenic differentiation of human mesenchymal stem cells and increases adipogenesis. Endocrinology 2004, 145, 2421–2432. [Google Scholar] [CrossRef] [Green Version]
- Sheyn, D.; Pelled, G.; Netanely, D.; Domany, E.; Gazit, D. The effect of simulated microgravity on human mesenchymal stem cells cultured in an osteogenic differentiation system: A bioinformatics study. Tissue Eng. Part A 2010, 16, 3403–3412. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, P.; Wu, H.; Choi, J.U.; Rowan, A.E.; Zhang, H.; Poole, K.; Lauko, J.; Chou, J. Modeling the Impact of Microgravity at the Cellular Level: Implications for Human Disease. Front. Cell Dev. Biol. 2020, 8, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gershovich, P.M.; Gershovich, I.G.; Buravkova, L.B. Cytoskeleton structures and adhesion properties of human stromal precursors under conditions of simulated microgravity. Tsitologiia 2009, 51, 896–904. [Google Scholar] [CrossRef] [PubMed]
- Pache, C.; Kühn, J.; Westphal, K.; Toy, M.F.; Parent, J.M.; Büchi, O.; Franco-Obregón, A.; Depeursinge, C.; Egli, M. Digital holographic microscopy real-time monitoring of cytoarchitectural alterations during simulated microgravity. J. Biomed. Opt. 2010, 15, 26021. [Google Scholar] [CrossRef] [Green Version]
- Symbolic Math Toolbox, version I; The MathWorks: Natick, MA, USA, 2019.
- Evarist Giné, M. Invariant Tests for Uniformity on Compact Riemannian Manifolds Based on Sobolev Norms. In The Annals of Statistics; Institute of Mathematica: San Jose, CA, USA, 1975; Volume 3, pp. 1243–1266. [Google Scholar]
- Keilson, J.; Petrondas, D.; Sumita, U.; Wellner, J. Significance points for some tests of uniformity on the sphere. J. Stat. Comput. Simul. 1983, 17, 195–218. [Google Scholar] [CrossRef]
- Mardia, K.V.; Jupp, P.E. Statistics of Directional Data, 2nd ed.; J. Wiley: Chichester, UK, 2000; ISBN 0471953334. [Google Scholar]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cazzaniga, A.; Ille, F.; Wuest, S.; Haack, C.; Koller, A.; Giger-Lange, C.; Zocchi, M.; Egli, M.; Castiglioni, S.; Maier, J.A. Scalable Microgravity Simulator Used for Long-Term Musculoskeletal Cells and Tissue Engineering. Int. J. Mol. Sci. 2020, 21, 8908. https://doi.org/10.3390/ijms21238908
Cazzaniga A, Ille F, Wuest S, Haack C, Koller A, Giger-Lange C, Zocchi M, Egli M, Castiglioni S, Maier JA. Scalable Microgravity Simulator Used for Long-Term Musculoskeletal Cells and Tissue Engineering. International Journal of Molecular Sciences. 2020; 21(23):8908. https://doi.org/10.3390/ijms21238908
Chicago/Turabian StyleCazzaniga, Alessandra, Fabian Ille, Simon Wuest, Carsten Haack, Adrian Koller, Christina Giger-Lange, Monica Zocchi, Marcel Egli, Sara Castiglioni, and Jeanette A. Maier. 2020. "Scalable Microgravity Simulator Used for Long-Term Musculoskeletal Cells and Tissue Engineering" International Journal of Molecular Sciences 21, no. 23: 8908. https://doi.org/10.3390/ijms21238908