Three-Dimensional Nuclear Telomere Profiling as a Biomarker for Recurrence in Oligodendrogliomas: A Pilot Study

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Clinical Surrogates and Pathological Diagnosis
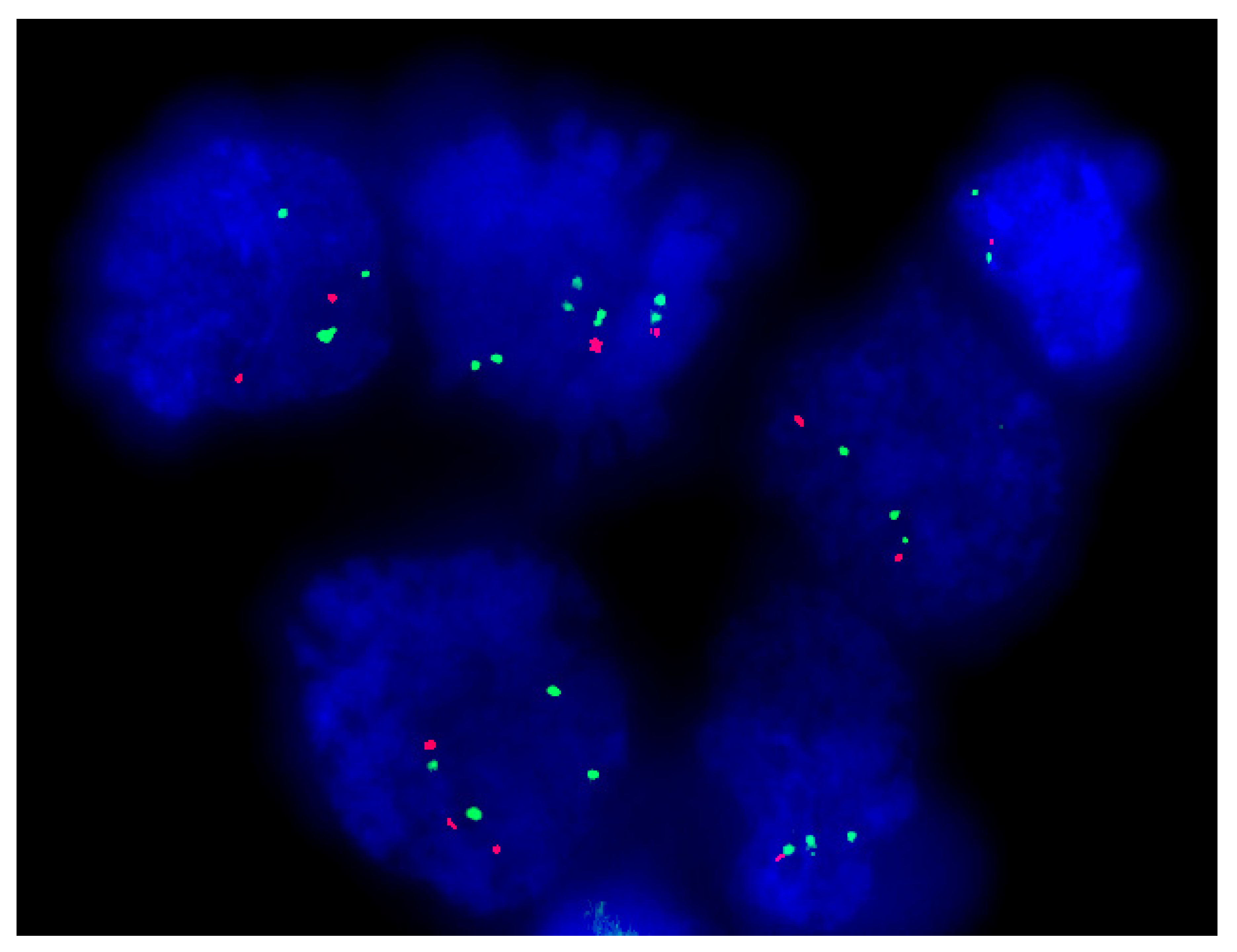
2.2. Molecular Cytogenetic Analysis: FISH for 1p/19q
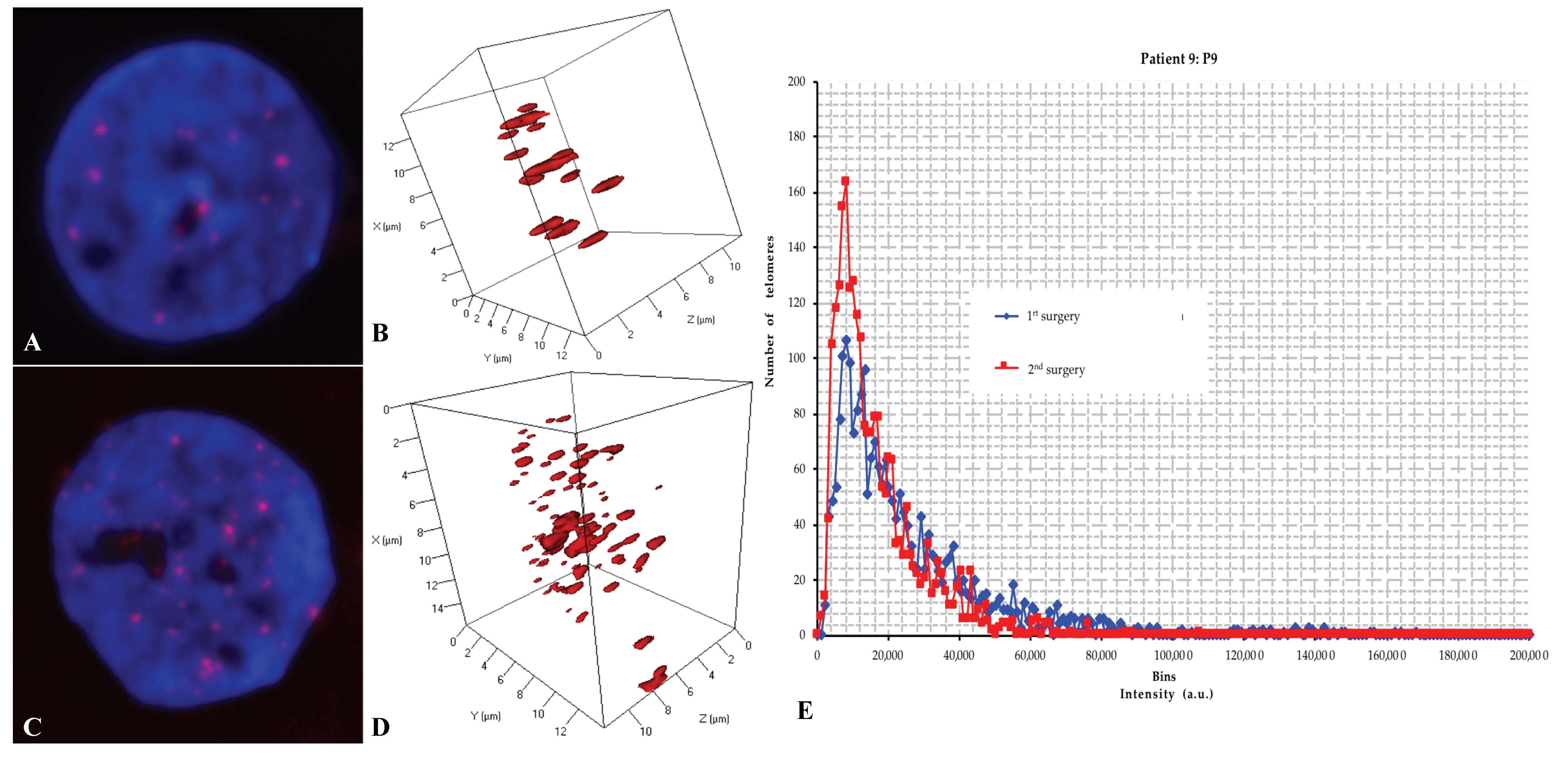
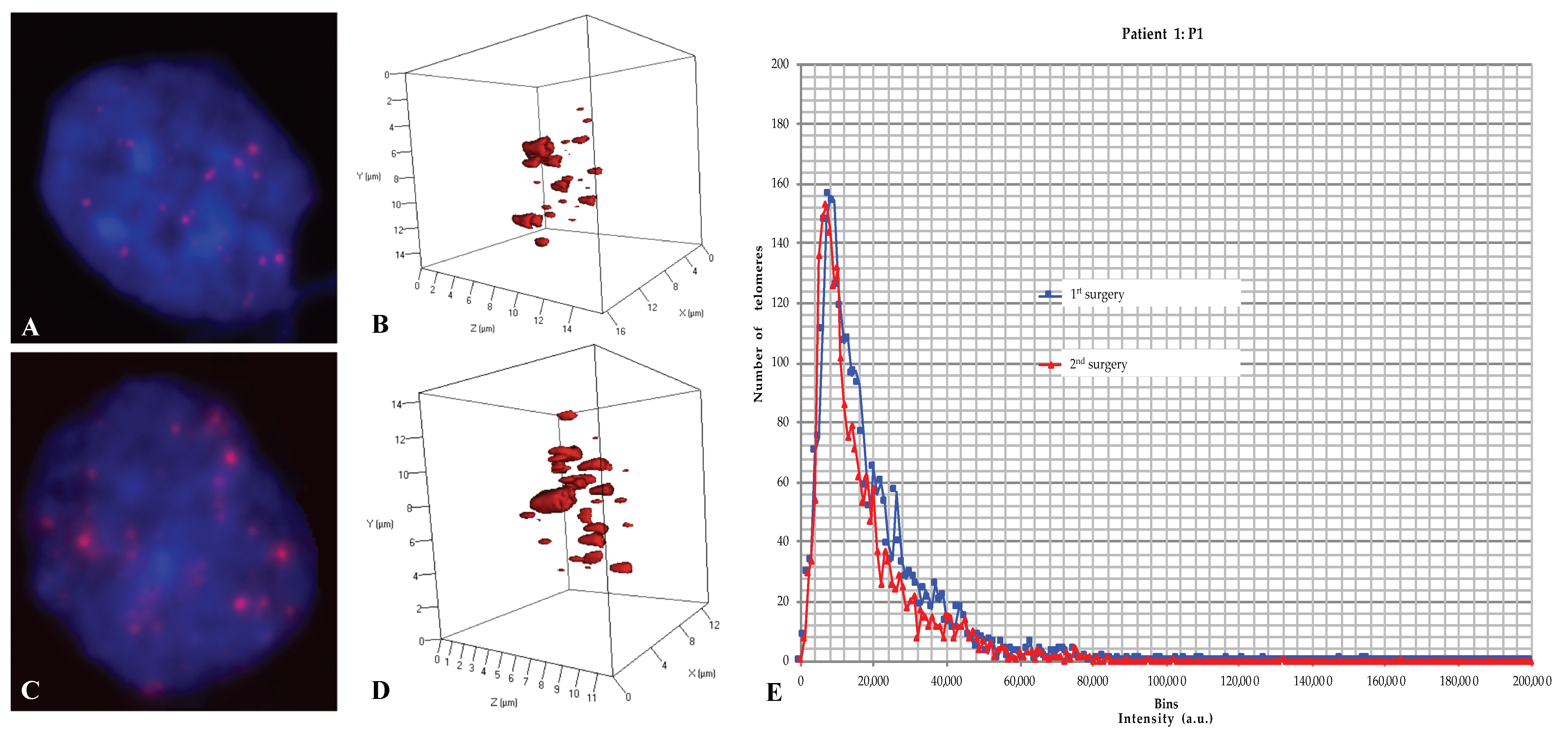
2.3. Three-Dimensional Nuclear Telomere Architecture Analyses
2.4. Comparative Analyses of the 3D Telomere Profile between the Two Groups
2.4.1. The Number of Signals, i.e., the Number of Telomeres
2.4.2. The signal Intensity, i.e., the Telomere Length
2.4.3. The Number of Telomere Aggregates
2.4.4. Nuclear Volume i.e., Volume of Each Cell
2.4.5. The a/c Ratio and Telomere Distributions per Nuclear Volume
2.5. Clinical Significance of Both Groups
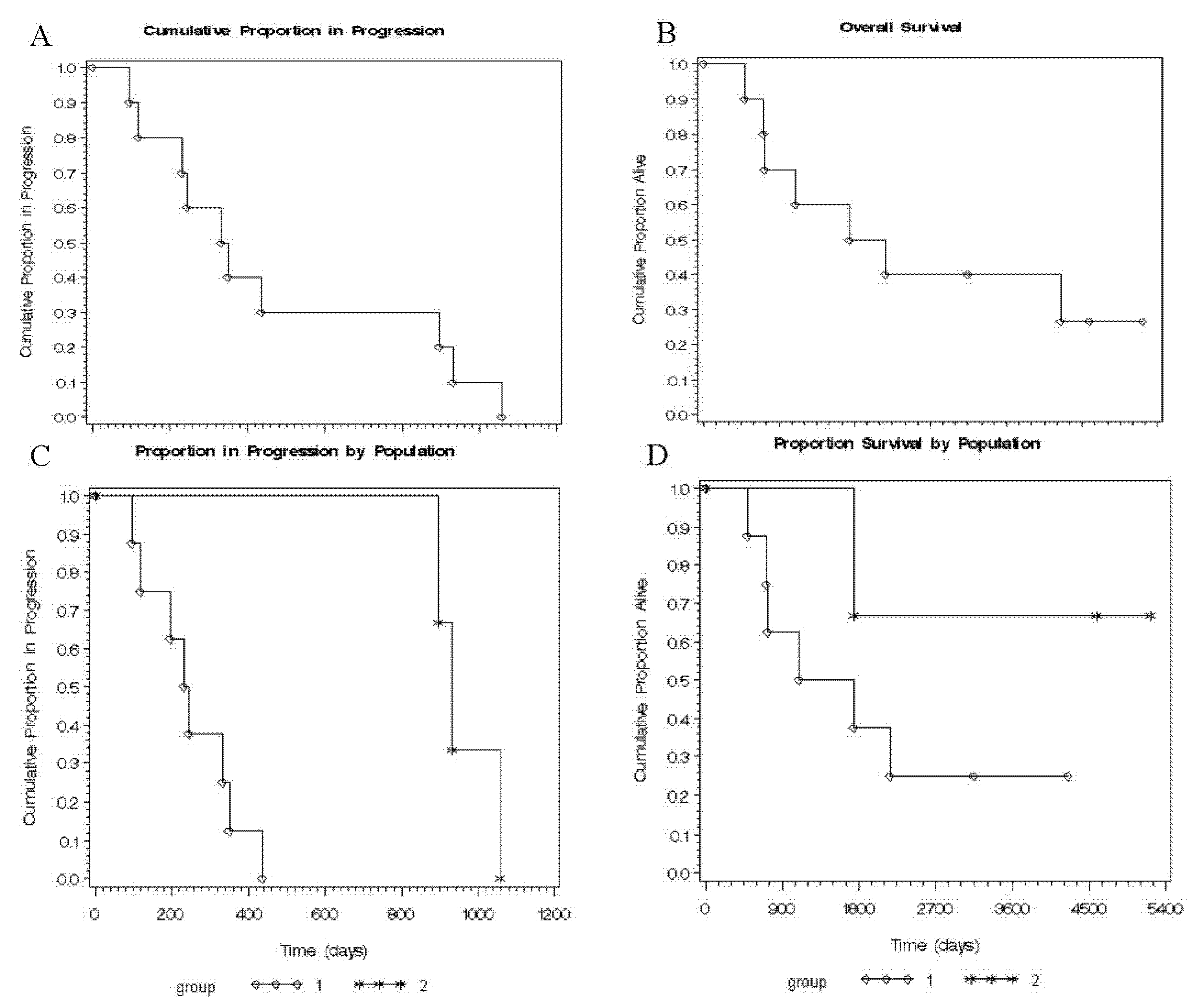
2.6. Time to Progression (TTP) and Overall Survival (OS) in Both Groups
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Samples
4.3. Histopathological Diagnosis
4.4. Molecular Cytogenetic Analysis
4.4.1. FISH Analysis
4.4.2. Image Acquisition for FISH with 1p/19q Probes
4.5. Quantitative—Fluorescent in Situ Hybridization (Q-FISH) Protocol for 3D Analysis
4.6. Image Acquisition and 3D Image Analysis Using TeloView®
4.7. Data Presentation and Statistical Analysis
4.8. Statistical Analyses and Overall Survival
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 3D | three-dimensional |
| AML | acute myeloid leukemia |
| CHUS | Centre Hospitalier Universitaire de Sherbrooke |
| CIN | chromosomal instability |
| DAPI | 4′,6-diamidino-2-phenylindole |
| FISH | fluorescence in situ hybridization |
| GBM | Glioblastoma |
| MDS | myelodysplastic syndromes |
| OS | overall survival |
| PNA | peptide nuclei acid |
| Q-FISH | Quantitative-Fluorescent in situ hybridization |
| RT | room temperature |
| TA | telomere aggregates |
| TP | touch preparation |
| TTP | time to progression |
| WHO | World Health Organization |
References
- Heidenreich, B.; Rachakonda, P.S.; Hosen, I.; Volz, F.; Hemminki, K.; Weyerbrock, A.; Kumar, R. TERT promoter mutations and telomere length in adult malignant gliomas and recurrences. Oncotarget 2015, 6, 10617–10633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K.; Burger, P.C.; Jouvet, A.; Scheithauer, B.W.; Kleihues, P. The 2007 WHO classification of tumours of the central nervous system. Acta Neuropathol. 2007, 114, 97–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadji, M.; Fortin, D.; Tsanaclis, A.M.; Drouin, R. Is the 1p/19q deletion a diagnostic marker of oligodendrogliomas? Cancer Genet. Cytogenet. 2009, 194, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Wood, M.D.; Halfpenny, A.M.; Moore, S.R. Applications of molecular neuro-oncology—A review of diffuse glioma integrated diagnosis and emerging molecular entities. Diagn. Pathol. 2019, 14, 29. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Lv, X.; Lu, C.; Ye, X.; Chen, X.; Fu, J.; Luo, C.; Zhao, Y. Prognostic factors of patients with Gliomas—An analysis on 335 patients with Glioblastoma and other forms of Gliomas. BMC Cancer 2020, 20, 35. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Kim, S.H.; Ahn, S.S.; Choi, H.J.; Yoon, H.I.; Cho, J.H.; Roh, T.H.; Kang, S.G.; Chang, J.H.; Suh, C.O. Extent of resection and molecular pathologic subtype are potent prognostic factors of adult WHO grade II glioma. Sci. Rep. 2020, 10, 2086. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Christopher, P.W.; Weiderpass, E.; Bernard, W.S. World Cancer Report Cancer Research for Cancer Prevention; International Agency for Research on Cancer: Lyon, France, 2020; ISBN 9789283204299. [Google Scholar]
- Perry, A.; Wesseling, P. Histologic classification of gliomas. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2016; Volume 134, pp. 71–95. ISBN 9780128029978. [Google Scholar]
- Wesseling, P.; van den Bent, M.; Perry, A. Oligodendroglioma: Pathology, molecular mechanisms and markers. Acta Neuropathol. 2015, 129, 809–827. [Google Scholar] [CrossRef] [Green Version]
- Jooma, R.; Waqas, M.; Khan, I. Diffuse low-grade glioma—Changing concepts in diagnosis and management: A review. Asian J. Neurosurg. 2019, 14, 356–363. [Google Scholar] [CrossRef] [Green Version]
- Rorke, L.B. Pathologic diagnosis as the gold standard. Cancer 1997, 79, 665–667. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.X.; Shi, Z.; Aibaidula, A.; Chen, H.; Tang, Q.; Li, K.K.W.; Chung, N.Y.F.; Chan, D.T.M.; Poon, W.S.; Mao, Y.; et al. Not all 1p/19q non-codeleted oligodendroglial tumors are astrocytic. Oncotarget 2016, 7, 64615–64630. [Google Scholar] [CrossRef]
- Burger, P.C. What is an Oligodendroglioma? Brain Pathol. 2006, 12, 257–259. [Google Scholar] [CrossRef] [PubMed]
- Bigner, S.H.; Rasheed, B.K.A.; Wiltshire, R.; McLendon, R.E. Morphologic and molecular genetic aspects of oligodendroglial neoplasms. Neuro Oncol. 2004, 1, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Hatanpaa, K.J.; Burger, P.C.; Eshleman, J.R.; Murphy, K.M.; Berg, K.D. Molecular diagnosis of oligodendroglioma in paraffin sections. Lab. Investig. 2003, 83, 419–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Bent, M.J.; Reni, M.; Gatta, G.; Vecht, C. Oligodendroglioma. Crit. Rev. Oncol. Hematol. 2008, 66, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Giannini, C.; Burger, P.C.; Berkey, B.A.; Cairncross, J.G.; Jenkins, R.B.; Mehta, M.; Curran, W.J.; Aldape, K. Anaplastic oligodendroglial tumors: Refining the correlation among histopathology, 1p 19q deletion and clinical outcome in Intergroup Radiation Therapy Oncology Group Trial 9402. Brain Pathol. 2008, 18, 360–369. [Google Scholar] [CrossRef]
- Cairncross, J.G.; Ueki, K.; Zlatescu, M.C.; Lisle, D.K.; Finkelstein, D.M.; Hammond, R.R.; Silver, J.S.; Stark, P.C.; Macdonald, D.R.; Ino, Y.; et al. Specific genetic predictors of chemotherapeutic response and survival in patients with anaplastic oligodendrogliomas. J. Natl. Cancer Inst. 1998, 90, 1473–1479. [Google Scholar] [CrossRef]
- Jenkins, R.B.; Blair, H.; Flynn, H.; Passe, S.; Law, M.; Ballman, K.V.; Aldape, K.; Giannini, C.; Buckner, J.C. A t(1;19)(q10;p10) mediates the combined deletions of 1p and 19q in human oligodendrogliomas. Cancer Res. 2006, 66, 9852–9861. [Google Scholar] [CrossRef] [Green Version]
- Griffin, C.A.; Burger, P.; Morsberger, L.; Yonescu, R.; Swierczynski, S.; Weingart, J.D.; Murphy, K.M. Identification of der(1;19)(q10;p10) in five oligodendrogliomas suggests mechanism of concurrent 1p and 19q loss. J. Neuropathol. Exp. Neurol. 2006, 65, 988–994. [Google Scholar] [CrossRef]
- Razis, E.; Kotoula, V.; Koliou, G.A.; Papadopoulou, K.; Vrettou, E.; Giannoulatou, E.; Tikas, I.; Labropoulos, S.V.; Rigakos, G.; Papaemmanoyil, S.; et al. Is There an Independent Role of TERT and NF1 in High Grade Gliomas? Transl. Oncol. 2020, 13, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Wesseling, P.; Capper, D. WHO 2016 Classification of gliomas. Neuropathol. Appl. Neurobiol. 2018, 44, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, C.; Azevedo, A.; Esteves, S.; Marques, A.R.; Martins, C.; Costa, I.; Mafra, M.; Bravo Marques, J.M.; Roque, L.; Pojo, M. Clinical insights gained by refining the 2016 WHO classification of diffuse gliomas with: EGFR amplification, TERT mutations, PTEN deletion and MGMT methylation. BMC Cancer 2019, 19, 968. [Google Scholar] [CrossRef] [Green Version]
- Walsh, K.M.; Codd, V.; Smirnov, I.V.; Rice, T.; Decker, P.A.; Hansen, H.M.; Kollmeyer, T.; Kosel, M.L.; Molinaro, A.M.; McCoy, L.S.; et al. Variants near TERT and TERC influencing telomere length are associated with high-grade glioma risk. Nat. Genet. 2014, 46, 731–735. [Google Scholar] [CrossRef] [Green Version]
- Walsh, K.M.; Wiencke, J.K.; Lachance, D.H.; Wiemels, J.L.; Molinaro, A.M.; Eckel-Passow, J.E.; Jenkins, R.B.; Wrensch, M.R. Telomere maintenance and the etiology of adult glioma. Neuro Oncol. 2015, 17, 1445–1452. [Google Scholar] [CrossRef] [Green Version]
- Boveri, T. On multipolar mitosis as a means of analysis of the cell nucleus. Found. Exp. Embryol. 1902, 74–97. [Google Scholar]
- Boveri, T. Concerning the origin of malignant tumours by Theodor Boveri. Translated and annotated by Henry Harris. J. Cell Sci. 2008, 121, 1–84. [Google Scholar] [CrossRef]
- Boveri, T. Zur Frage der Entstehung Maligner Tumoren [Origin of malignant tumors]. Gustav Fish. Jena 1914. [Google Scholar] [CrossRef]
- Callén, E.; Surrallés, J. Telomere dysfunction in genome instability syndromes. Mutat. Res. Rev. Mutat. Res. 2004, 567, 85–104. [Google Scholar] [CrossRef]
- de Lange, T.; Shiue, L.; Myers, R.M.; Cox, D.R.; Naylor, S.L.; Killery, A.M.; Varmus, H.E. Structure and variability of human chromosome ends. Mol. Cell. Biol. 1990, 10, 518–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harley, C.B.; Futcher, A.B.; Greider, C.W. Telomeres shorten during ageing of human fibroblasts. Nature 1990, 345, 458–460. [Google Scholar] [CrossRef] [PubMed]
- DePinho, R.A.; Polyak, K. Cancer chromosomes in crisis. Nat. Genet. 2004, 36, 932–934. [Google Scholar] [CrossRef] [PubMed]
- Cheung, A.L.M.; Deng, W. Telomere dysfunction, genome instability and cancer. Front. Biosci. 2008, 13, 2075–2090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernal, A.; Tusell, L. Telomeres: Implications for cancer development. Int. J. Mol. Sci. 2018, 19, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lansdorp, P.M. Telomeres and disease. EMBO J. 2009, 28, 2532–2540. [Google Scholar] [CrossRef]
- Chuang, T.C.Y.; Moshir, S.; Garini, Y.; Chuang, A.Y.C.; Young, I.T.; Vermolen, B.; van den Doel, R.; Mougey, V.; Perrin, M.; Braun, M.; et al. The three-dimensional organization of telomeres in the nucleous of mammalian cells. BMC Biol. 2004, 2, 12. [Google Scholar] [CrossRef] [Green Version]
- De Vos, W.H.; Hoebe, R.A.; Joss, G.H.; Haffmans, W.; Baatout, S.; Van Oostveldt, P.; Manders, E.M.M. Controlled light exposure microscopy reveals dynamic telomere microterritories throughout the cell cycle. Cytom. Part A 2009, 75, 428–439. [Google Scholar] [CrossRef]
- Gadji, M.; Fortin, D.; Tsanaclis, A.M.; Garini, Y.; Katzir, N.; Wienburg, Y.; Yan, J.; Klewes, L.; Klonisch, T.; Drouin, R.; et al. Three-dimensional nuclear telomere architecture is associated with differential time to progression and overall survival in glioblastoma patients. Neoplasia 2010, 12, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Vermolen, B.J.; Garini, Y.; Mai, S.; Mougey, V.; Fest, T.; Chuang, T.C.Y.; Chuang, A.Y.C.; Wark, L.; Young, I.T. Characterizing the three-dimensional organization of telomeres. Cytom. Part A 2005, 67, 144–150. [Google Scholar] [CrossRef]
- Louis, S.F.; Vermolen, B.J.; Garini, Y.; Young, I.T.; Guffei, A.; Lichtensztejn, Z.; Kuttler, F.; Chuang, T.C.Y.; Moshir, S.; Mougey, V.; et al. c-Myc induces chromosomal rearrangements through telomere and chromosome remodeling in the interphase nucleus. Proc. Natl. Acad. Sci. USA 2005, 102, 9613–9618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadji, M.; Awe, J.A.; Rodrigues, P.; Kumar, R.; Houston, D.S.; Klewes, L.; Diéye, T.N.; Rego, E.M.; Passetto, R.F.; De Oliveira, F.M.; et al. Profiling three-dimensional nuclear telomeric architecture of myelodysplastic syndromes and acute myeloid leukemia defines patient subgroups. Clin. Cancer Res. 2012, 18, 3293–3304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakimoto, H.; Aoyagi, M.; Nakayama, T.; Nagashima, G.; Yamamoto, S.; Tamaki, M.; Hirakawa, K. Prognostic significance of Ki-67 labeling indices obtained using MIB-1 monoclonal antibody in patients with supratentorial astrocytomas. Cancer 1996, 77, 373–380. [Google Scholar] [CrossRef]
- Coons, S.W.; Johnson, P.C.; Pearl, D.K. The prognostic significance of Ki-67 labeling indices for oligodendrogliomas. Neurosurgery 1997, 41, 878–885. [Google Scholar] [CrossRef]
- Bailey, O.T. Genesis of the percival bailey-cushing classification of gliomas. Pediatr. Neurosurg. 1985, 12, 261–265. [Google Scholar] [CrossRef]
- Ferguson, S.; Lesniak, M.S. Percival Bailey and the classification of brain tumors. Neurosurg. Focus 2005, 18, E7. [Google Scholar] [CrossRef]
- Schiffer, D. Brain Tumor Pathology: Current Diagnostic Hotspots and Pitfalls; Springer: Dordrecht, The Netherlands, 2006; ISBN 9781402039980. [Google Scholar]
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K. WHO Classification Of Tumours Of The Central Nervous System (IARC WHO Classification Of Tumours); International Agency for Research on Cancer, World Health Organization: Genewa, Switzerland, 2016. [Google Scholar]
- Wen, P.Y.; Huse, J.T. 2016 World Health Organization Classification of Central Nervous System Tumors. Contin. Lifelong Learn. Neurol. 2017, 23, 1531–1547. [Google Scholar] [CrossRef]
- Gupta, A.; Dwivedi, T. A simplified overview of World Health Organization classification update of central nervous system tumors 2016. J. Neurosci. Rural Pract. 2017, 8, 629–641. [Google Scholar] [CrossRef]
- Pekmezci, M.; Rice, T.; Molinaro, A.M.; Walsh, K.M.; Decker, P.A.; Hansen, H.; Sicotte, H.; Kollmeyer, T.M.; McCoy, L.S.; Sarkar, G.; et al. Adult infiltrating gliomas with WHO 2016 integrated diagnosis: Additional prognostic roles of ATRX and TERT. Acta Neuropathol. 2017, 133, 1001–1016. [Google Scholar] [CrossRef]
- Gadji, M.; Vallente, R.; Klewes, L.; Righolt, C.; Wark, L.; Kongruttanachok, N.; Knecht, H.; Mai, S. Nuclear Remodeling as a Mechanism for Genomic Instability in Cancer. In Advances in Cancer Research; Intratumor Diversity and Clonal Evolution in Cancer; Gisselsson, D., Ed.; Academic Press: Oxford, UK, 2011; Volume 112, pp. 77–126. [Google Scholar]
- Guffei, A.; Lichtensztejn, Z.; Gonçlves, A.; Silva, S.; Louis, S.F.; Caporali, A.; Mai, S. c-Myc—Dependent Formation of Robertsonian Translocation Chromosomes in Mouse Cells. Neoplasia 2007, 9, 578-IN1. [Google Scholar] [CrossRef] [Green Version]
- Mai, S.; Garini, Y. The significance of telomeric aggregates in the interphase nuclei of tumor cells. J. Cell. Biochem. 2006, 97, 904–915. [Google Scholar] [CrossRef] [PubMed]
- Guijon, F.B.; Greulich-Bode, K.M.; Paraskevas, M.; Baker, P.; Mai, S. Premalignant cervical lesions are characterized by dihydrofolate reductase gene amplification and c-Myc overexpression: Possible biomarkers. J. Low. Genit. Tract Dis. 2007, 11, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Knecht, H.; Sawan, B.; Lichtensztejn, D.; Lemieux, B.; Wellinger, R.J.; Mai, S. The 3D nuclear organization of telomeres marks the transition from Hodgkin to Reed-Sternberg cells. Leukemia 2009, 23, 565–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knecht, H.; Kongruttanachok, N.; Sawan, B.; Brossard, J.; Prévost, S.; Turcotte, E.; Lichtensztejn, Z.; Lichtensztejn, D.; Mai, S. Three-dimensional telomere signatures of Hodgkin- and Reed-Sternberg cells at diagnosis identify patients with poor response to conventional chemotherapy. Transl. Oncol. 2012, 5, 269–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadji, M.; Crous-Tsanaclis, A.-M.; Mathieu, D.; Mai, S.; Fortin, D.; Drouin, R. A new der(1;7)(q10;p10) leading to a singular 1p loss in a case of glioblastoma with oligodendroglioma component. Neuropathology 2014, 34, 170–178. [Google Scholar] [CrossRef]
- Clark, K.H.; Villano, J.L.; Nikiforova, M.N.; Hamilton, R.L.; Horbinski, C. 1p/19q testing has no significance in the workup of glioblastomas. Neuropathol. Appl. Neurobiol. 2013, 39, 706–717. [Google Scholar] [CrossRef] [Green Version]
- Ballester, L.Y.; Huse, J.T.; Tang, G.; Fuller, G.N. Molecular classification of adult diffuse gliomas: Conflicting IDH1/IDH2, ATRX, and 1p/19q results. Hum. Pathol. 2017, 69, 15–22. [Google Scholar] [CrossRef]
- Vogazianou, A.P.; Chan, R.; Bäcklund, L.M.; Pearson, D.M.; Liu, L.; Langford, C.F.; Gregory, S.G.; Collins, V.P.; Ichimura, K. Distinct patterns of 1p and 19q alterations identify subtypes of human gliomas that have different prognoses. Neuro Oncol. 2010, 12, 664–678. [Google Scholar] [CrossRef] [Green Version]
- Vega, J.E.V.; Brat, D.J. Incorporating Advances in Molecular Pathology into Brain Tumor Diagnostics. Adv. Anat. Pathol. 2018, 25, 143–171. [Google Scholar] [CrossRef]
- Smith, J.S.; Jenkins, R.B. Genetic alterations in adult diffuse glioma: Occurrence, significance, and prognostic implications. Front. Biosci. 2000, 5, D213–D231. [Google Scholar] [CrossRef] [Green Version]
- Fallon, K.B.; Palmer, C.A.; Roth, K.A.; Nabors, L.B.; Wang, W.; Carpenter, M.; Banerjee, R.; Forsyth, P.; Rich, K.; Perry, A. Prognostic Value of 1p, 19q, 9p, 10q, and EGFR-FISH Analyses in Recurrent Oligodendrogliomas. J. Neuropathol. Exp. Neurol. 2004, 63, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Snuderl, M.; Eichler, A.F.; Ligon, K.L.; Vu, Q.U.; Silver, M.; Betensky, R.A.; Ligon, A.H.; Wen, P.Y.; Louis, D.N.; Iafrate, A.J. Polysomy for chromosomes 1 and 19 predicts earlier recurrence in anaplastic oligodendrogliomas with concurrent 1p/19q loss. Clin. Cancer Res. 2009, 15, 6430–6437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bralten, L.B.; Nouwens, S.; Kockx, C.; Erdem, L.; Hoogenraad, C.C.; Kros, J.M.; Moorhouse, M.J.; Smitt, P.A.S.; van der Spek, P.; van Ijcken, W.; et al. Absence of common somatic alterations in genes on 1p and 19q in oligodendrogliomas. PLoS ONE 2011, 6, e22000. [Google Scholar] [CrossRef] [Green Version]
- Bettegowda, C.; Agrawal, N.; Jiao, Y.; Sausen, M.; Wood, L.D.; Hruban, R.H.; Rodriguez, F.J.; Cahill, D.P.; McLendon, R.; Riggins, G.; et al. Mutations in CIC and FUBP1 contribute to human oligodendroglioma. Science 2011, 333, 453–455. [Google Scholar] [CrossRef] [Green Version]
- Yip, S.; Butterfield, Y.S.; Morozova, O.; Chittaranjan, S.; Blough, M.D.; An, J.; Birol, I.; Chesnelong, C.; Chiu, R.; Chuah, E.; et al. Concurrent CIC mutations, IDH mutations, and 1p/19q loss distinguish oligodendrogliomas from other cancers. J. Pathol. 2012, 226, 7–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, Z.; Li, Y.; Fan, X.; Zhang, C.; Wang, Y.; Jiang, T.; Liu, X. Prognostic value of a microRNA signature as a novel biomarker in patients with lower-grade gliomas. J. Neuro Oncol. 2018, 137, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Wang, C.; Hou, Z.; Wang, G.; Lv, J.; Wang, H.; Yang, J.; Zhang, Z.; Zhang, H. Serum microRNA-376 family as diagnostic and prognostic markers in human gliomas. Cancer Biomark Sect. A Dis. Markers. 2017, 19, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Vuong, H.G.; Altibi, A.M.A.; Duong, U.N.P.; Ngo, H.T.T.; Pham, T.Q.; Fung, K.M.; Hassell, L. BRAF Mutation is Associated with an Improved Survival in Glioma—A Systematic Review and Meta-analysis. Mol. Neurobiol. 2018, 55, 3718–3724. [Google Scholar] [CrossRef]
- Chen, W.; Yu, Q.; Chen, B.; Lu, X.; Li, Q. The prognostic value of a seven-microRNA classifier as a novel biomarker for the prediction and detection of recurrence in glioma patients. Oncotarget 2016, 7, 53392–53413. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, L.H.; Schuster, D.; Herz, H. Generalized approach for accelerated maximum likelihood based image restoration applied to three-dimensional fluorescence microscopy. J. Microsc. 2001, 204, 99–107. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID and Exponent 1 or 0 | Age | Sex | KI67 | TTP (day) | OS (day) | Surgery | Diagnostic and Clinical Evolution | 1p/19q Status | |
|---|---|---|---|---|---|---|---|---|---|
| 1 P1 | 40 | F | 5% | 894 | 5212 | 1 | Oligodendroglioma Mid aggressive | 1p-/19q- | |
| 2 | Oligodendroglioma | 1p-/19q- | |||||||
| 1 P2 | 40 | F | ++ | 350 | 2172 | 1 | Oligodendroglioma Mid aggressive | 1p-/19q- | |
| 2 | Oligodendroglioma | 1p-/19q- | |||||||
| 1 P3 | 42 | M | 15% | 95 | 721 | 1 | Oligodendroglioma aggressive | 1p- | |
| 2 | Oligodendroglioma | 1p- | |||||||
| 1 P4 | P4a | 29 | M | ++ | 930 | 1748 | 1 | Oligodendroglioma indolent | 1p-/19q- Polysomic |
| 2 | Oligodendroglioma | 1p-/19q- Polysomic | |||||||
| P4b | ++ | 197 | 3 | Oligodendroglioma | 19q- | ||||
| 4 | Oligoastrocytoma | 19q- | |||||||
| 1 P5 | 33 | F | >40% | 335 | 4242 | 1 | Oligodendroglioma Mid aggressive | 1p-/19q- Polysomic | |
| 2 | Oligodendroglioma | 1p-/19q- Polysomic | |||||||
| 0 P6 | 55 | F | 5% | 1058 | 4582 | 1 | Oligodendroglioma indolent | 1p-/19q- | |
| 2 | Oligodendroglioma | 1p-/19q- | |||||||
| 0 P7 | 37 | M | >25% | 437 | 3141 | 1 | Oligodendroglioma indolent | 1p-/19q- | |
| 2 | Anaplastic oligodendroglioma | 1p-/19q- | |||||||
| 1 P8 | 47 | F | Hetero-geneous till 60% | 246 | 500 | 1 | Anaplastic oligodendroglioma Invasive | 19q- Polysomic | |
| 2 | Anaplastic oligodendroglioma | 19q- Polysomic | |||||||
| 1 P9 | 79 | M | >30% | 232 | 1103 | 1 | Anaplastic oligodendroglioma Aggressive | 1p-/19q- Polysomic | |
| 2 | Anaplastic oligodendroglioma | 1p- Polysomic | |||||||
| 1 P10 | 46 | F | Nd | 119 | 735 | 1 | Oligodendroglioma aggressive | 1p- Polysomic | |
| 2 | Oligodendroglioma | 1p- Polysomic | |||||||
| Group | Level of Surgery | Total Number of Signals | Total Number of Aggregates | Total Intensity | Average Intensity of All Signals | Nuclear Volume | a/c Ratio | Telomere per Nuclear Volume | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | Std Dev | Mean | Std Dev | Mean | Std Dev | Mean | Std Dev | Mean | Std Dev | Mean | Std Dev | Mean | Std Dev | ||
| 1 | 1a | 29.8444816 | 15.5864981 | 3.23411371 | 2.64075267 | 569,912.151 | 281,821.009 | 20,387.6865 | 8092.16645 | 954,885.37 | 527,813.807 | 9.527 | 5.8 | 0.03802044 | 0.02309804 |
| 2a | 33.5899160 | 18.0550011 | 3.79327731 | 2.90812548 | 544,668.245 | 290,813.157 | 16,772.3811 | 4445.41517 | 1,131,519.38 | 961,970.627 | 12.749 | 45.4 | 0.27817647 | 4.15627785 | |
| p value 1a-2a | <0.0001 | <0.0001 | 0.0804 | <0.0001 | <0.0001 | 1.0000 | 0.1053 | ||||||||
| 2 | 1b | 29.5312500 | 19.7783065 | 3.28125000 | 3.27593066 | 427,011.450 | 190,557.547 | 16,499.5065 | 5437.76924 | 745,605.69 | 446,145.342 | 827,766.671 | 10,470,359.1 | 0.06224833 | 0.06443754 |
| 2b | 32.4500000 | 17.6534779 | 3.72500000 | 2.89425803 | 528,013.631 | 219,867.712 | 18,502.6929 | 7327.63749 | 865,677.43 | 470,170.711 | 7.255 | 4.4 | 0.04492112 | 0.02969461 | |
| p value 1b-2b | 0.0365 | 0.0929 | 0.0002 | 0.0020 | 0.0799 | 0.0299 | 0.9524 | ||||||||
| Group 1 versus group 2 with the different surgeries | p value 1a-1b | 0.0518 | 0.0803 | <0.0001 | <0.0001 | <0.0001 | 0.0540 | 0.6117 | |||||||
| p value 1a-2b | <0.0001 | 0.0002 | 0.0236 | <0.0001 | <0.0001 | 0.5573 | 0.5642 | ||||||||
| p value 2a-1b | 0.2070 | 0.4434 | <0.0001 | 0.0017 | <0.0001 | 0.0542 | 0.1398 | ||||||||
| p value 2a -2b | 0.2471 | 0.2394 | 0.2244 | 0.6714 | <0.0001 | 0.5577 | 0.1222 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gadji, M.; Mathur, S.; Bélanger, B.; Jangamreddy, J.R.; Lamoureux, J.; Tsanaclis, A.M.C.; Fortin, D.; Drouin, R.; Mai, S. Three-Dimensional Nuclear Telomere Profiling as a Biomarker for Recurrence in Oligodendrogliomas: A Pilot Study. Int. J. Mol. Sci. 2020, 21, 8539. https://doi.org/10.3390/ijms21228539
Gadji M, Mathur S, Bélanger B, Jangamreddy JR, Lamoureux J, Tsanaclis AMC, Fortin D, Drouin R, Mai S. Three-Dimensional Nuclear Telomere Profiling as a Biomarker for Recurrence in Oligodendrogliomas: A Pilot Study. International Journal of Molecular Sciences. 2020; 21(22):8539. https://doi.org/10.3390/ijms21228539
Chicago/Turabian StyleGadji, Macoura, Shubha Mathur, Brigitte Bélanger, Jaganmohan Reddy Jangamreddy, Josée Lamoureux, Ana Maria Crous Tsanaclis, David Fortin, Régen Drouin, and Sabine Mai. 2020. "Three-Dimensional Nuclear Telomere Profiling as a Biomarker for Recurrence in Oligodendrogliomas: A Pilot Study" International Journal of Molecular Sciences 21, no. 22: 8539. https://doi.org/10.3390/ijms21228539