Are Metal Ions That Make up Orthodontic Alloys Cytotoxic, and Do They Induce Oxidative Stress in a Yeast Cell Model?

Abstract
:1. Introduction
2. Results
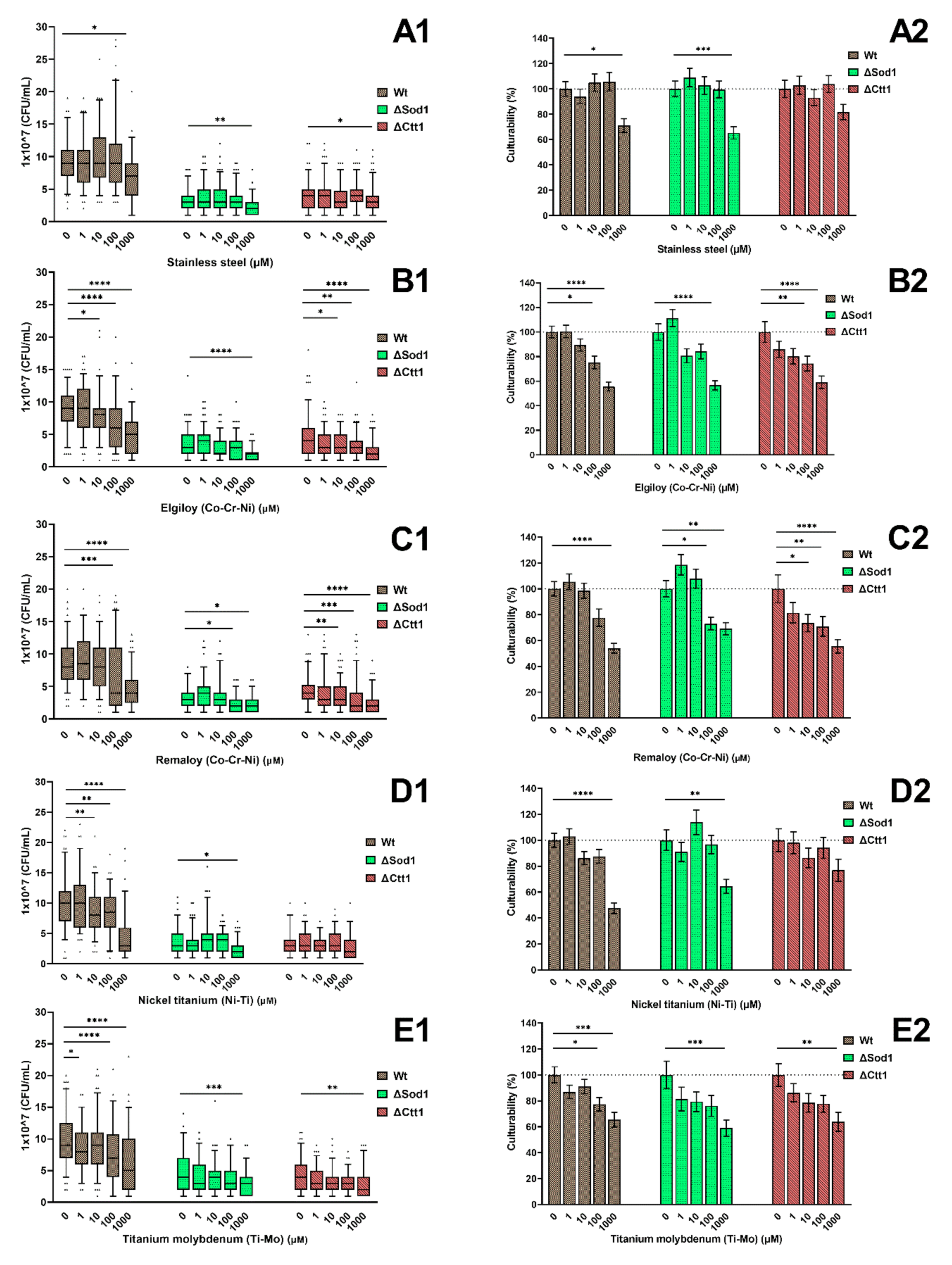
2.1. Cell Culturability
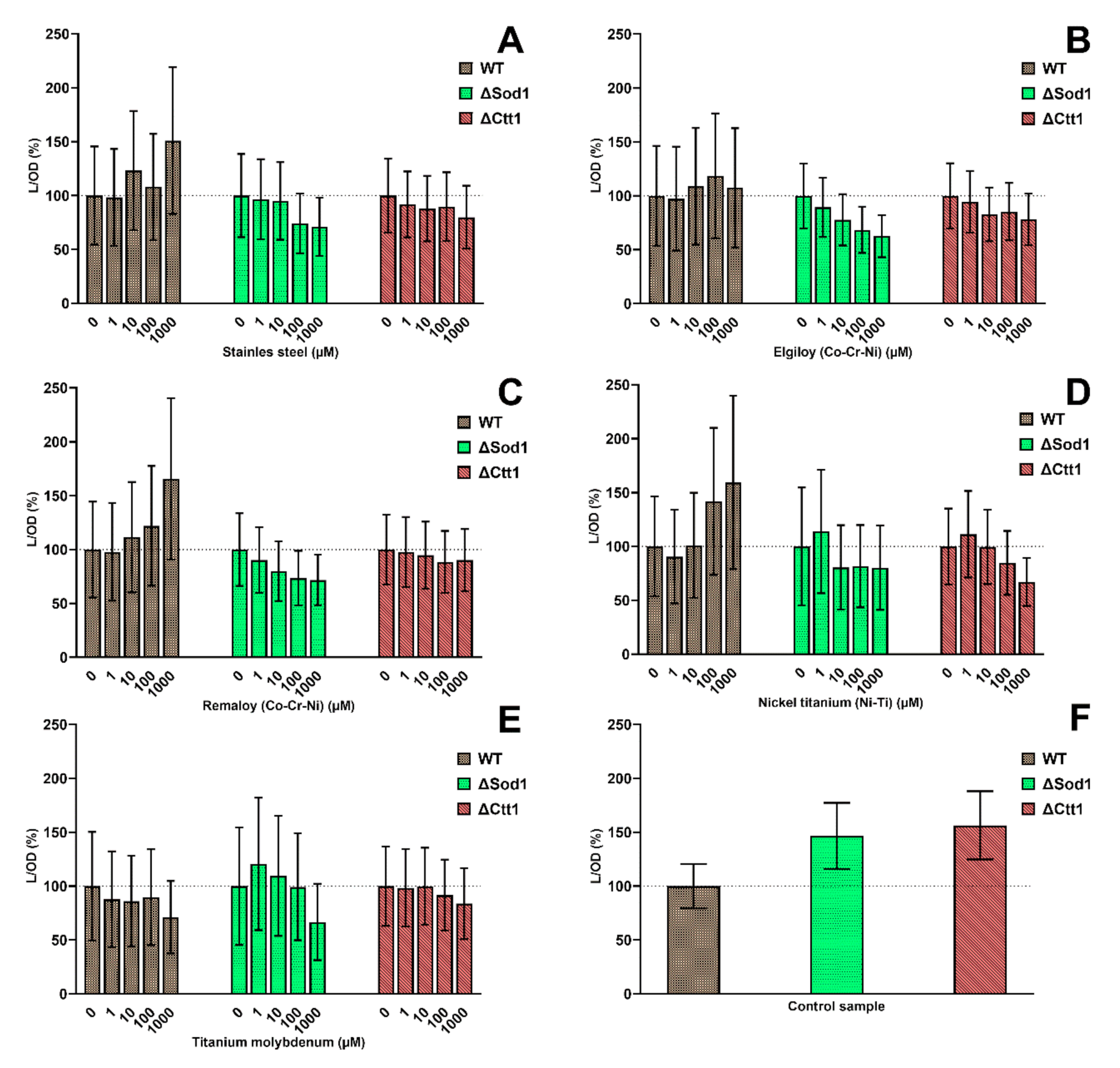
2.2. Cell Metabolic Activity
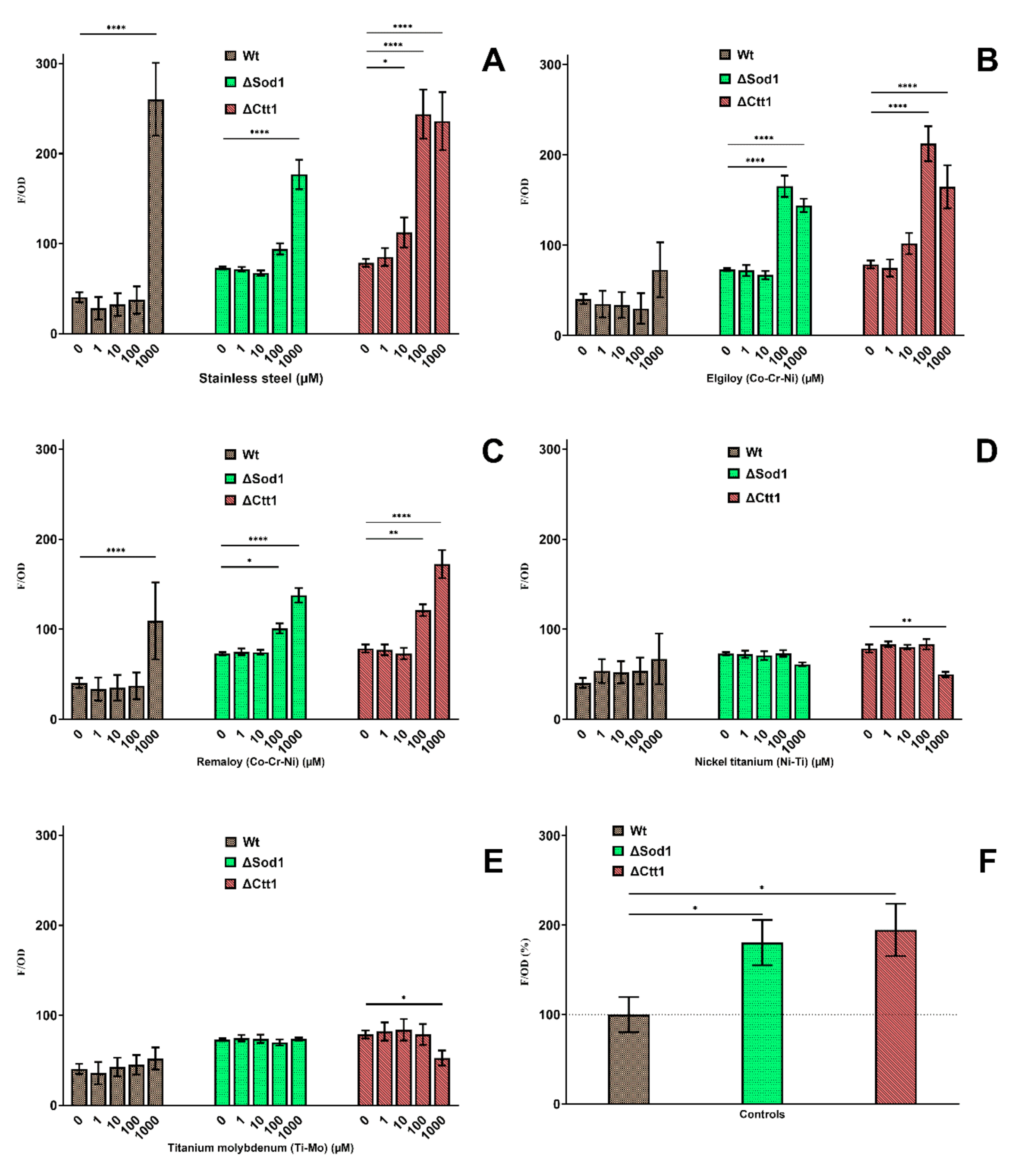
2.3. Intracellular Oxidation
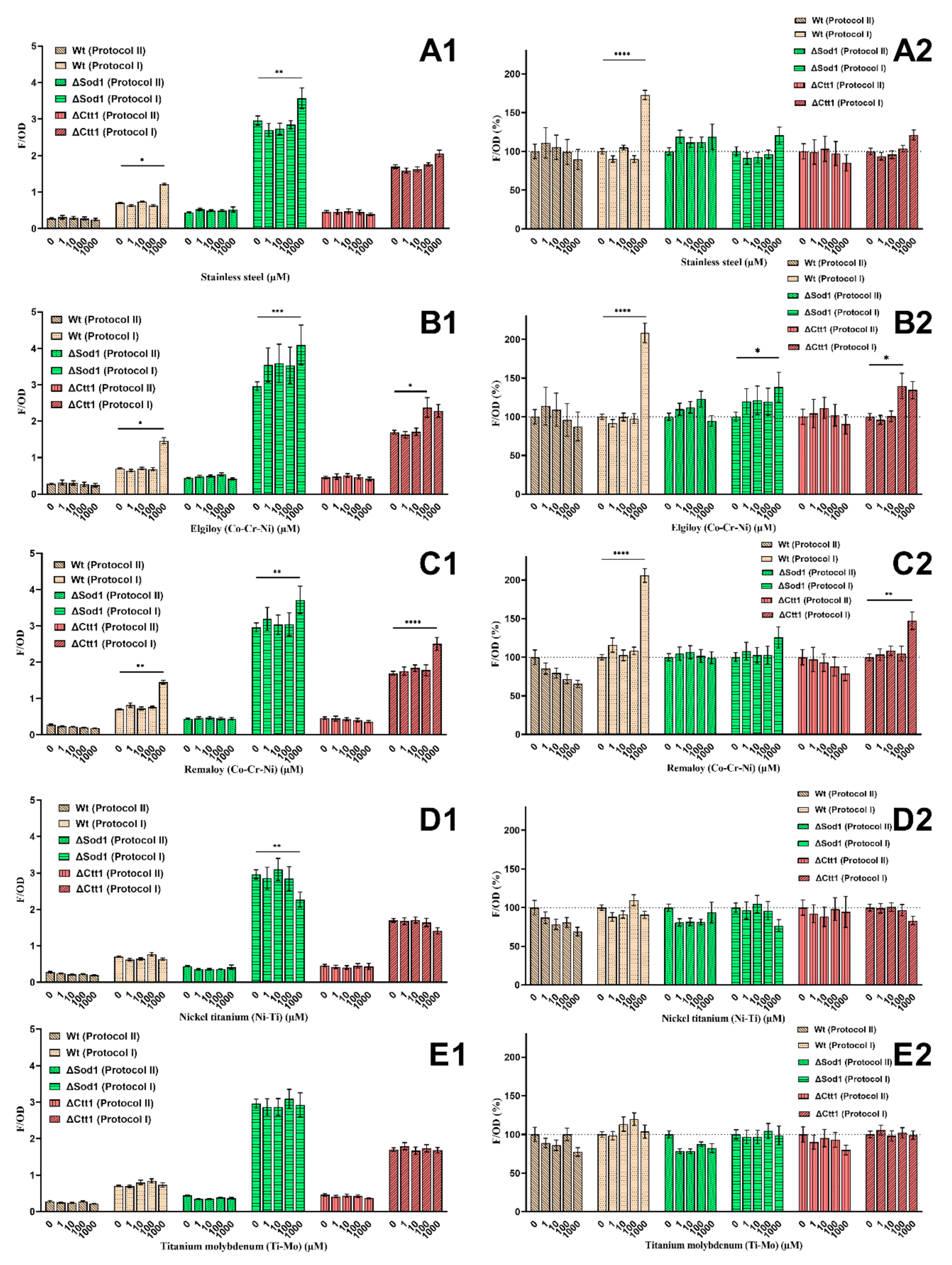
2.4. Oxidative Lipid Damage
3. Discussion
4. Materials and Methods
4.1. Preparation of Yeast Cultures
4.2. Preparation of Metal Ion Solutions
4.3. Metal Treatment of Yeast Strains
4.4. Cell Culturability
4.5. Cell Metabolic Activity
4.6. Intracellular Oxidation
4.7. Oxidative Lipid Damage
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
| CAT | Catalase |
| GPx | Glutathione peroxidase |
| L | Luminescence |
| OD | Optical density |
| H2DCF-DA | 2′,7′-dichlorofluorescein diacetate |
| MDA | Malondialdehyde |
| CFU | Colony forming units |
| PBS | Phosphate-Buffered Saline |
| Wt | Wild type |
| SS | Stainless steel |
| ELG | Elgiloy |
| REM | Remaloy |
| NiTi | Nickel-titanium |
| TiMo | Β-titanium |
| EDAX | Energy-dispersive X-ray spectroscopy |
References
- Wendl, B.; Wiltsche, H.; Lankmayr, E.; Winsauer, H.; Walter, A.; Muchitsch, A.; Jakse, N.; Wendl, M.; Wendl, T. Schwermetallfreisetzungsprofile aus kieferorthopädischen Bändern Brackets und Drähten: Eine in-vitro-Untersuchung. J. Orofac. Orthop. 2017, 78, 494–503. [Google Scholar] [CrossRef]
- Nascimento, G.; Santos, R.L.D.; Pithon, M.M.; De Souza Araújo, M.T.; Nojima, M.G.; Nojima, L.I. The effect of electric spot-welding on the mechanical properties of different orthodontic wire alloys leonard euler andrade. Mater. Res. 2012, 15, 409–414. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, M.-N.; Lou, T.; Retrouvey, J.-M.; Suri, S. Biomaterials used in orthodontics: Brackets, archwires, and clear aligners. In Advanced Dental Biomaterials; Elsevier: Amsterdam, The Netherlands, 2019; pp. 541–579. [Google Scholar]
- Sifakakis, I.; Eliades, T. Adverse reactions to orthodontic materials. Aust. Dent. J. 2017, 62, 20–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucchese, A.; Carinci, F.; Brunelli, G.; Monguzzi, R. An in vitro study of resistance to corrosion in brazed and laser-welded orthodontic appliances. Eur. J. Inflamm. 2011, 9, 67–72. [Google Scholar]
- Muguruma, T.; Iijima, M.; Mizoguchi, I. Corrosion of laser-welded stainless steel orthodontic wires. Orthod. Waves 2018, 77, 18–23. [Google Scholar] [CrossRef]
- Mikulewicz, M.; Chojnacka, K.; Wołowiec, P. Release of metal ions from fixed orthodontic appliance An in vitro study in continuous flow system. Angle Orthod. 2014, 84, 140–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahmasbi, S.; Ghorbani, M.; Sheikh, T.; Yaghoubnejad, Y. Galvanic Corrosion and Ion Release from Different Orthodontic Brackets and Wires in Acidic Artificial Saliva. J. Dent. Sch. Shahid Beheshti Univ. Med. Sci. 2019, 32, 37–44. [Google Scholar] [CrossRef]
- Reimann, S.; Rewari, A.; Keilig, L.; Widu, F.; Jäger, A.; Bourauel, C. Materialtechnische Untersuchungen kieferorthopädischer Brackets nach Wiederaufbereitung. J. Orofac. Orthop. 2012, 73, 454–466. [Google Scholar] [CrossRef]
- Mikulewicz, M.; Chojnacka, K. Trace Metal release from orthodontic appliances by in vivo studies: A systematic literature review. Biol. Trace Elem. Res. 2010, 137, 127–138. [Google Scholar] [CrossRef]
- Mikulewicz, M.; Chojnacka, K. Release of metal ions from orthodontic appliances by in vitro studies: A systematic literature review. Biol. Trace Elem. Res. 2011, 139, 241–256. [Google Scholar] [CrossRef]
- Bandeira, A.M.; Ferreira Martinez, E.; Ana Dias Demasi, P. Evaluation of toxicity and response to oxidative stress generated by orthodontic bands in human gingival fibroblasts. Angle Orthod. 2020, 90, 285–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanti Das, T.; Wati, M.R.; Fatima-Shad, K. Oxidative Stress Gated by Fenton and Haber Weiss Reactions and Its Association with Alzheimer’s Disease. Arch. Neurosci. 2014, 2, e20078. [Google Scholar] [CrossRef] [Green Version]
- Nuran Ercal, B.S.P.; Hande Gurer-Orhan, B.S.P.; Nukhet Aykin-Burns, B.S.P.; Ercal, N.; Gurer-Orhan, H.; Aykin-Burns, N. Toxic metals and oxidative stress. Part I: Mechanisms involved in metal induced oxidative damage. Curr. Top. Med. Chem. 2001, 1, 529–539. [Google Scholar] [CrossRef]
- Toy, E.; Yuksel, S.; Ozturk, F.; Karatas, O.H.; Yalcin, M. Evaluation of the genotoxicity and cytotoxicity in the buccal epithelial cells of patients undergoing orthodontic treatment with three light-cured bonding composites by using micronucleus testing. Korean J. Orthod. 2014, 44, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Galagan, J.E.; Henn, M.R.; Ma, L.J.; Cuomo, C.A.; Birren, B. Genomics of the fungal kingdom: Insights into eukaryotic biology. Genome Res. 2005, 15, 1620–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, V.; Moradas-Ferreira, P. Oxidative stress and signal transduction in Saccharomyces cerevisiae: Insights into ageing, apoptosis and diseases. Mol. Aspects Med. 2001, 22, 217–246. [Google Scholar] [CrossRef]
- Khurana, V.; Lindquist, S. Modelling neurodegeneration in Saccharomyces cerevisiae: Why cook with baker’s yeast? Nat. Rev. Neurosci. 2010, 11, 436–449. [Google Scholar] [CrossRef]
- Sigler, K.; Chaloupka, J.; Brozmanová, J.; Stadler, N.; Höfer, M. Oxidative stress in microorganisms—I. Microbial vs. higher cells—Damage and defenses in relation to cell aging and death. Folia Microbiol. Praha 1999, 44, 587–624. [Google Scholar] [CrossRef]
- Moradas-Ferreira, P.; Costa, V.; Piper, P.; Mager, W. The molecular defences against reactive oxygen species in yeast. Mol. Microbiol. 1996, 19, 651–658. [Google Scholar] [CrossRef]
- Traini, T.; Danza, M.; Zollino, I.; Altavilla, R.; Lucchese, A.; Sollazzo, V.; Trapella, G.; Brunelli, G.; Carinci, F. Histomorphic-metric evaluation of an immediately loaded implant retrieved from human mandible after 2 years. Int. J. Immunopathol. Pharmacol. 2011, 24, 31–36. [Google Scholar] [CrossRef]
- Cal, E.; Cetintas, V.; Boyacioglu, H.; Güneri, P. Cytotoxicity of Dental Implants: The Effects of Ultrastructural Elements. Int. J. Oral Maxillofac. Implants 2017, 32, 1281–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandar, S.; Kotian, R.; Madhyastha, P.; Kabekkodu, S.; Rao, P. In vitro evaluation of cytotoxicity and corrosion behavior of commercially pure titanium and Ti-6Al-4V alloy for dental implants. J. Indian Prosthodont. Soc. 2017, 17, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Cândea Ciurea, A.; Şurlin, P.; Stratul, Ş.I.; Soancă, A.; Roman, A.; Moldovan, M.; Tudoran, B.L.; Pall, E. Evaluation of the biocompatibility of resin composite-based dental materials with gingival mesenchymal stromal cells. Microsc. Res. Tech. 2019, 82, 1768–1778. [Google Scholar] [CrossRef]
- Jiang, R.D.; Lin, H.; Zheng, G.; Zhang, X.M.; Du, Q.; Yang, M. In vitro dentin barrier cytotoxicity testing of some dental restorative materials. J. Dent. 2017, 58, 28–33. [Google Scholar] [CrossRef]
- Caldas, I.P.; Alves, G.G.; Barbosa, I.B.; Scelza, P.; De Noronha, F.; Scelza, M.Z. In vitro cytotoxicity of dental adhesives: A systematic review. Dent. Mater. 2019, 35, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Hafez, H.S.; Selim, E.M.N.; Kamel Eid, F.H.; Tawfik, W.A.; Al-Ashkar, E.A.; Mostafa, Y.A. Cytotoxicity, genotoxicity, and metal release in patients with fixed orthodontic appliances: A longitudinal in-vivo study. Am. J. Orthod. Dentofac. Orthop. 2011, 140, 298–308. [Google Scholar] [CrossRef]
- Ortiz, A.J.; Fernández, E.; Vicente, A.; Calvo, J.L.; Ortiz, C. Metallic ions released from stainless steel, nickel-free, and titanium orthodontic alloys: Toxicity and DNA damage. Am. J. Orthod. Dentofac. Orthop. 2011, 140, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Ortega, E.; Jos, A.; Cameán, A.M.; Pato-Mourelo, J.; Segura-Egea, J.J. In vitro evaluation of cytotoxicity and genotoxicity of a commercial titanium alloy for dental implantology. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2010, 702, 17–23. [Google Scholar] [CrossRef]
- Bueno, R.C.; Basting, R.T. In vitro study of human osteoblast proliferation and morphology on orthodontic mini-implants. Angle Orthod. 2015, 85, 920–926. [Google Scholar] [CrossRef] [Green Version]
- Limberger, K.M.; Westphalen, G.H.; Menezes, L.M.; Medina-Silva, R. Cytotoxicity of orthodontic materials assessed by survival tests in Saccharomyces cerevisiae. Dent. Mater. 2011, 27, e81–e86. [Google Scholar] [CrossRef]
- Gonçalves, T.S.; De Menezes, L.M.; Trindade, C.; Machado, M.D.S.; Thomas, P.; Fenech, M.; Henriques, J.A.P. Cytotoxicity and genotoxicity of orthodontic bands with or without silver soldered joints. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2014, 762, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Arndt, M.; Brück, A.; Scully, T.; Jäger, A.; Bourauel, C. Nickel ion release from orthodontic NiTi wires under simulation of realistic in-situ conditions. J. Mater. Sci. 2005, 40, 3659–3667. [Google Scholar] [CrossRef]
- Kusy, R.P. Orthodontic Biomaterials: From the Past to the Present. Angle Orthod. 2002, 72, 501–512. [Google Scholar] [CrossRef]
- Nayak, R.S.; Khanna, B.; Pasha, A.; Vinay, K.; Narayan, A.; Chaitra, K. Evaluation of Nickel and Chromium Ion Release During Fixed Orthodontic Treatment Using Inductively Coupled Plasma-Mass Spectrometer: An In Vivo Study. J. Int. oral Heal. JIOH 2015, 7, 14–20. [Google Scholar]
- Chaturvedi, T.; Upadhayay, S. An overview of orthodontic material degradation in oral cavity. Indian J. Dent. Res. 2010, 21, 275. [Google Scholar] [CrossRef] [PubMed]
- Hurt, A.J. Digital technology in the orthodontic laboratory. Am. J. Orthod. Dentofac. Orthop. 2012, 141, 245–247. [Google Scholar] [CrossRef]
- Freitas, M.P.M.; Oshima, H.M.S.; Menezes, L.M. Release of toxic ions from silver solder used in orthodontics: An in-situ evaluation. Am. J. Orthod. Dentofac. Orthop. 2011, 140, 177–181. [Google Scholar] [CrossRef]
- Quadras, D.; Nayak, U.; Kumari, N.; Priyadarshini, H.; Gowda, S.; Fernandes, B. In vivo study on the release of nickel, chromium, and zinc in saliva and serum from patients treated with fixed orthodontic appliances. Dent. Res. J. Isfahan. 2019, 16, 209–215. [Google Scholar] [CrossRef]
- Amini, F.; Jafari, A.; Amini, P.; Sepasi, S. Metal ion release from fixed orthodontic appliances-An in vivo study. Eur. J. Orthod. 2012, 34, 126–130. [Google Scholar] [CrossRef] [Green Version]
- Kovac, V.; Poljsak, B.; Perinetti, G.; Primozic, J.; Reis, F.S. Systemic Level of Oxidative Stress during Orthodontic Treatment with Fixed Appliances. Biomed Res. Int. 2019, 2019. [Google Scholar] [CrossRef]
- Martín-Cameán, A.; Jos, Á.; Mellado-García, P.; Iglesias-Linares, A.; Solano, E.; Cameán, A.M. In vitro and in vivo evidence of the cytotoxic and genotoxic effects of metal ions released by orthodontic appliances: A review. Environ. Toxicol. Pharmacol. 2015, 40, 86–113. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, V.; Subba Reddy, V. Biodegradation of nickel and chromium from space maintainers: An in vitro study. J. Indian Soc. Pedod. Prev. Dent. 2010, 28, 6. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Applications of Toxicogenomic Technologies to Predictive Toxicology and Risk Assessment; National Academies Press: Washington, DC, USA, 2007. [Google Scholar]
- Zhao, Z. Iron and oxidizing species in oxidative stress and Alzheimer’s disease. AGING Med. 2019, 2, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef]
- Farrugia, G.; Balzan, R.; Madeo, F.; Breitenbach, M. Oxidative stress and programmed cell death in yeast. Front. Oncol. 2012, 2, 64. [Google Scholar] [CrossRef] [Green Version]
- El Medawar, L.; Rocher, P.; Hornez, J.C.; Traisnel, M.; Breme, J.; Hildebrand, H.F. Electrochemical and cytocompatibility assessment of NiTiNOL memory shape alloy for orthodontic use. In Proceedings of the Biomolecular Engineering; Elsevier: Amsterdam, The Netherlands, 2002; Volume 19, pp. 153–160. [Google Scholar]
- Issa, Y.; Brunton, P.; Waters, C.M.; Watts, D.C. Cytotoxicity of metal ions to human oligodendroglial cells and human gingival fibroblasts assessed by mitochondrial dehydrogenase activity. Dent. Mater. 2008, 24, 281–287. [Google Scholar] [CrossRef]
- Terpilowska, S.; Siwicki, A.K. Interactions between chromium(III) and iron(III), molybdenum(III) or nickel(II): Cytotoxicity, genotoxicity and mutagenicity studies. Chemosphere 2018, 201, 780–789. [Google Scholar] [CrossRef]
- Orrenius, S. Mechanisms of Oxidative Cell Damage. In Free Radicals: From Basic Science to Medicine; Birkhäuser Basel: Basel, Switzerland, 1993; pp. 47–64. [Google Scholar]
- Chen, A.; Zeng, G.; Chen, G.; Liu, L.; Shang, C.; Hu, X.; Lu, L.; Chen, M.; Zhou, Y.; Zhang, Q. Plasma membrane behavior, oxidative damage, and defense mechanism in Phanerochaete chrysosporium under cadmium stress. Process. Biochem. 2014, 49, 589–598. [Google Scholar] [CrossRef]
- Huang, M.L.-H.; Chiang, S.; Kalinowski, D.S.; Bae, D.-H.; Sahni, S.; Richardson, D.R. The Role of the Antioxidant Response in Mitochondrial Dysfunction in Degenerative Diseases: Cross-Talk between Antioxidant Defense, Autophagy, and Apoptosis. Oxid. Med. Cell. Longev. 2019, 2019, 6392763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhova, A.V.; Tkachenko, A.G. ATP/ADP alteration as a sign of the oxidative stress development in Escherichia coli cells under antibiotic treatment. FEMS Microbiol. Lett. 2014, 353, 69–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, C.M.; Rinna, A.; Forman, H.J.; Ventura, A.L.M.; Persechini, P.M.; Ojcius, D.M. ATP activates a reactive oxygen species-dependent oxidative stress response and secretion of proinflammatory cytokines in macrophages. J. Biol. Chem. 2007, 282, 2871–2879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Roper, M.G. Measurement of DCF fluorescence as a measure of reactive oxygen species in murine islets of Langerhans. Anal. Methods 2014, 6, 3019–3024. [Google Scholar] [CrossRef] [Green Version]
- Diaz, G.; Liu, S.; Isola, R.; Diana, A.; Falchi, A.M. Mitochondrial localization of reactive oxygen species by dihydrofluorescein probes. Histochem. Cell Biol. 2003, 120, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Wang, Y.F.; Lin, Y.H.; Yen, S.F. Nickel-induced oxidative stress and effect of antioxidants in human lymphocytes. Arch. Toxicol. 2003, 77, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Gholinejad, Z.; Khadem Ansari, M.H.; Rasmi, Y. Titanium dioxide nanoparticles induce endothelial cell apoptosis via cell membrane oxidative damage and p38, PI3K/Akt, NF-κB signaling pathways modulation. J. Trace Elem. Med. Biol. 2019, 54, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.A.; Saquib, Q.; Ahamed, M.; Farshori, N.N.; Ahmad, J.; Wahab, R.; Khan, S.T.; Alhadlaq, H.A.; Musarrat, J.; Al-Khedhairy, A.A.; et al. Molybdenum nanoparticles-induced cytotoxicity, oxidative stress, G2/M arrest, and DNA damage in mouse skin fibroblast cells (L929). Colloids Surf. B Biointerfaces 2015, 125, 73–81. [Google Scholar] [CrossRef]
- Terpilowska, S.; Siwicki, A.K. Pro- and antioxidant activity of chromium(III), iron(III), molybdenum(III) or nickel(II) and their mixtures. Chem. Biol. Interact. 2019, 298, 43–51. [Google Scholar] [CrossRef]
- Caicedo, M.; Jacobs, J.J.; Reddy, A.; Hallab, N.J. Analysis of metal ion-induced DNA damage, apoptosis, and necrosis in human (Jurkat) T-cells demonstrates Ni2+ and V3+ are more toxic than other metals: Al 3+, Be 2+, Co 2+, Cr 3+, Cu. J. Biomed. Mater. Res. Part A 2008, 86A, 905–913. [Google Scholar] [CrossRef]
- Dentaurum Materials for Orthodontic Products. Available online: https://www.dentaurum.de/files/KFO-Werkstoffliste-20.pdf (accessed on 27 August 2020).
- Jakubowski, W.; Bartosz, G. Estimation of oxidative stress in Saccharomyces cerevisae with fluorescent probes. Int. J. Biochem. Cell Biol. 1997, 29, 1297–1301. [Google Scholar] [CrossRef]
- Tetz, L.M.; Kamau, P.W.; Cheng, A.A.; Meeker, J.D.; Loch-Caruso, R. Troubleshooting the dichlorofluorescein assay to avoid artifacts in measurement of toxicant-stimulated cellular production of reactive oxidant species. J. Pharmacol. Toxicol. Methods 2013, 67, 56–60. [Google Scholar] [CrossRef] [Green Version]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Source |
|---|---|---|
| ATCC 204508 (Wt) | MATa; SUC2; mal; mel; gal2; CUP1; flo1; flo8-1; hap1 | American Type Culture Collection, Manassas, Virginia, United States |
| Y06913 (ΔSod1) | BY4741; MATa; ura3Δ0; leu2Δ0; his3Δ1; met15Δ0; YJR104c::kanMX4 | EUROSCARF, Oberursel, Germany |
| Y04718 (ΔCtt1) | BY4741; MATa; ura3Δ0; leu2Δ0; his3Δ1; met15Δ0; YGR088w::kanMX4 | EUROSCARF, Oberursel, Germany |
| Metal Composition (w/v) | ||||||
|---|---|---|---|---|---|---|
| Orthodontic Alloy | Fe | Ni | Cr | Co | Ti | Mo |
| Stainless steel (SS) | 72% | 10% | 18% | |||
| Cobalt-chromium (Elgiloy—ELG) | 18% | 15% | 20% | 40% | 7% | |
| Cobalt-chromium (Remaloy—REM) | 5% | 21% | 20% | 50% | 4% | |
| Nickel-titanium (NiTi) | 55% | 45% | ||||
| β-titanium (TiMo) | 78% | 12% | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovač, V.; Poljšak, B.; Primožič, J.; Jamnik, P. Are Metal Ions That Make up Orthodontic Alloys Cytotoxic, and Do They Induce Oxidative Stress in a Yeast Cell Model? Int. J. Mol. Sci. 2020, 21, 7993. https://doi.org/10.3390/ijms21217993
Kovač V, Poljšak B, Primožič J, Jamnik P. Are Metal Ions That Make up Orthodontic Alloys Cytotoxic, and Do They Induce Oxidative Stress in a Yeast Cell Model? International Journal of Molecular Sciences. 2020; 21(21):7993. https://doi.org/10.3390/ijms21217993
Chicago/Turabian StyleKovač, Vito, Borut Poljšak, Jasmina Primožič, and Polona Jamnik. 2020. "Are Metal Ions That Make up Orthodontic Alloys Cytotoxic, and Do They Induce Oxidative Stress in a Yeast Cell Model?" International Journal of Molecular Sciences 21, no. 21: 7993. https://doi.org/10.3390/ijms21217993