Offensive Behavior, Striatal Glutamate Metabolites, and Limbic–Hypothalamic–Pituitary–Adrenal Responses to Stress in Chronic Anxiety

 , , , , , ,
, , , , , , 
Abstract
:1. Introduction
2. Results
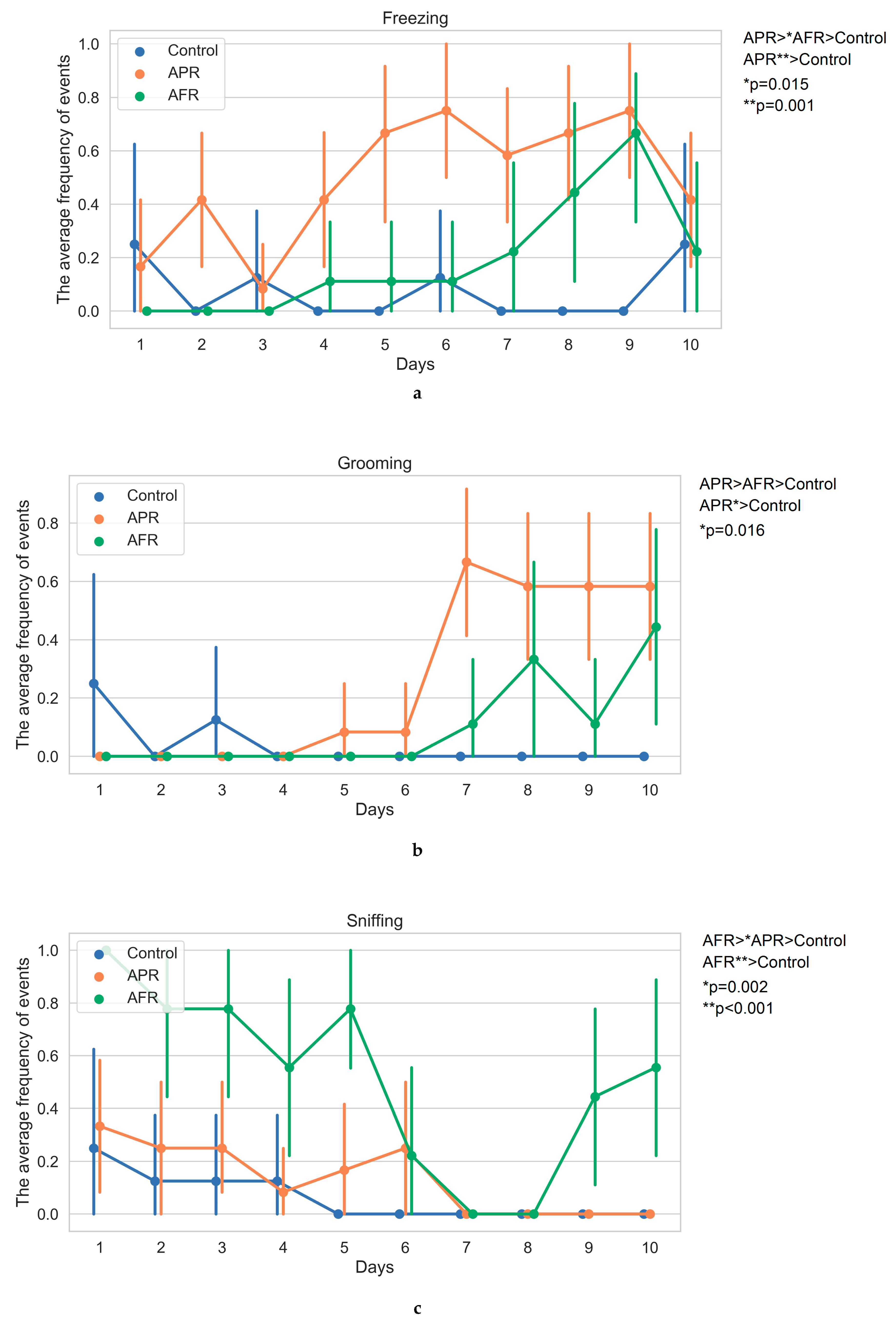
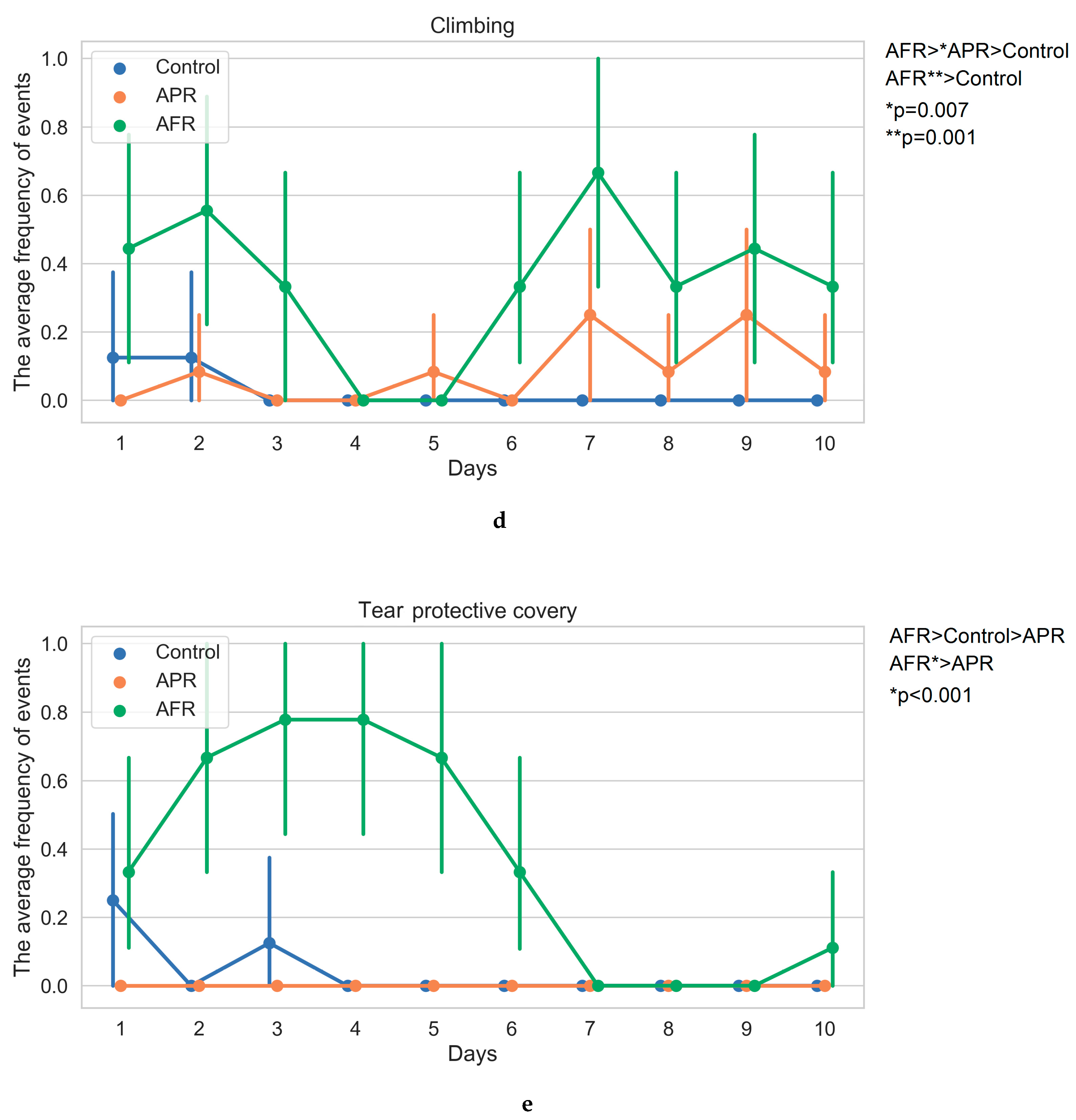
2.1. Behavior on Exposure to Stimuli (Kruskal–Wallis One-Way/Bonferroni Post Hoc Tests)
2.2. AFR and APR Rat Behavior in Elevated Plus Maze
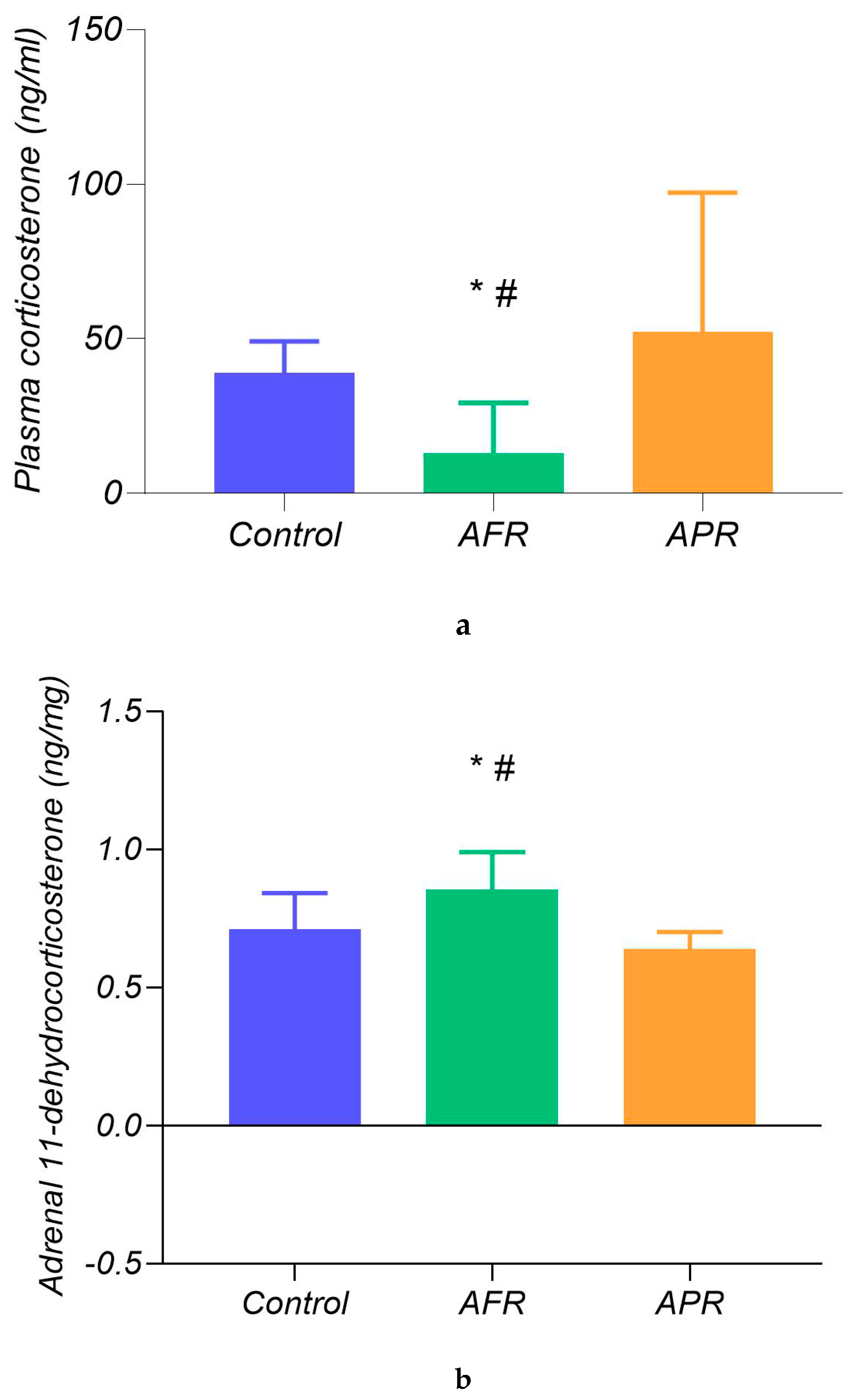
2.3. Biochemical Differences in Our Classified Behavioral Subtypes
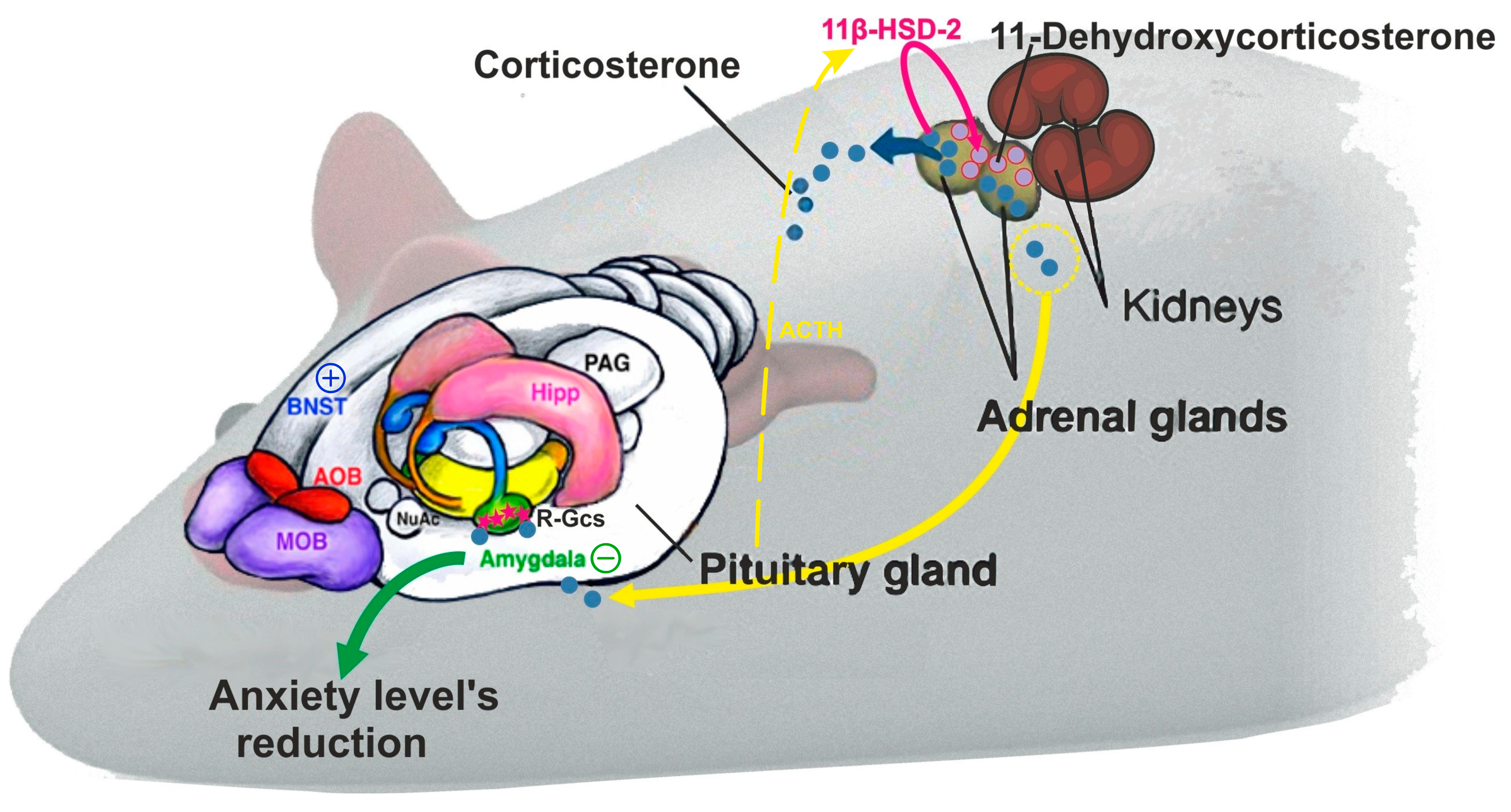
3. Discussion
4. Materials and Methods
4.1. Experimental Procedure
- Days 1–10: PSS;
- Days 11–22: rest;
- Day 23: elevated plus maze test;
- Day 27: striatum metabolite measurement by MRS;
- Day 28: euthanasia, harvesting of blood and organs.
4.2. Behavioral Activity
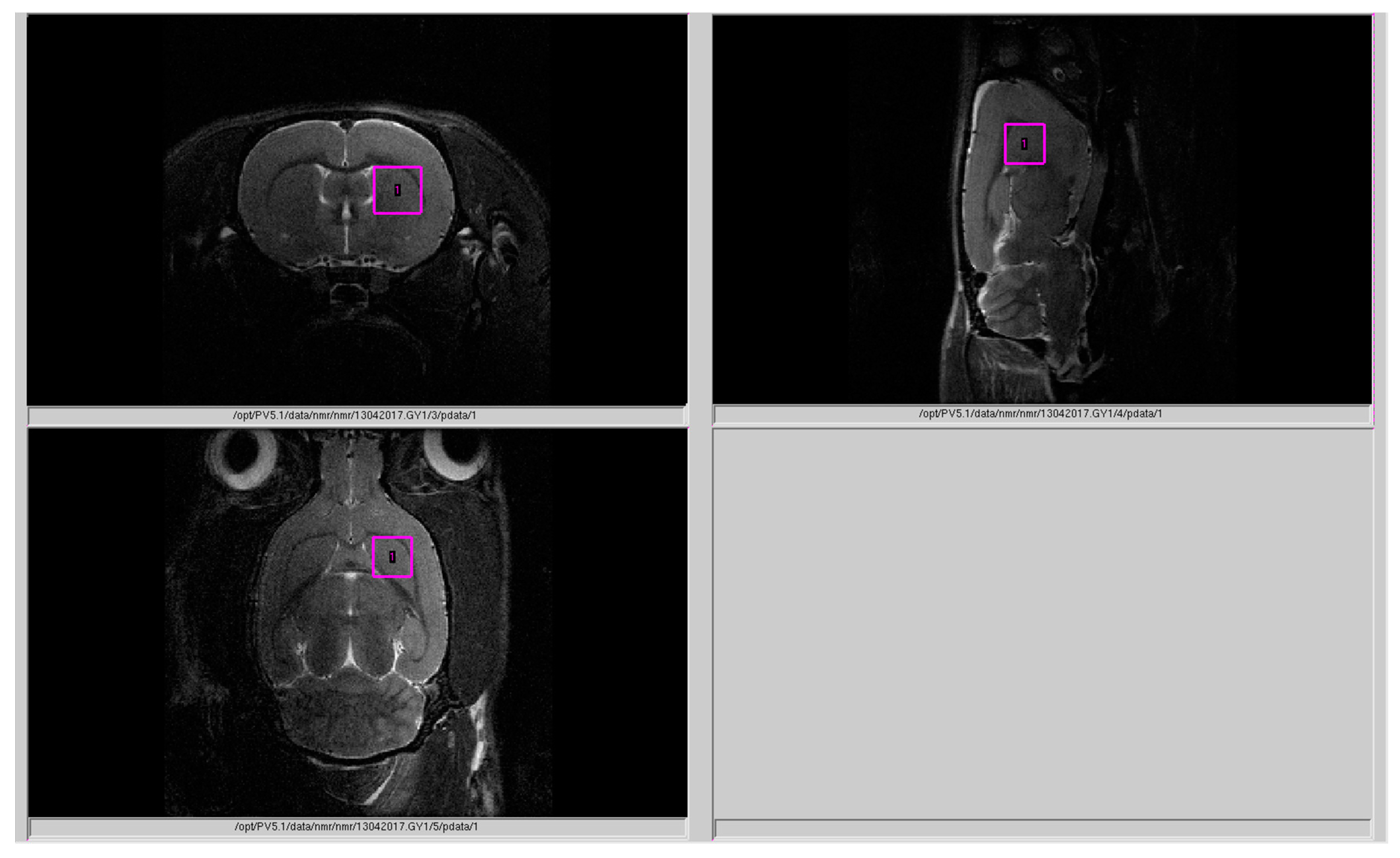
4.3. Magnetic Resonance Spectroscopy (MRS)
4.4. Processing of 1H Spectra
4.5. Hormonal Measurements
4.6. Data Analyses
5. Conclusions
6. Limitations of the Study
Author Contributions
Funding
Conflicts of Interest
References
- Bowen, M.T.; Dass, S.A.H.; Booth, J.; Suraev, A.; Vyas, A.; McGregor, I.S. Active coping toward predatory stress is associated with lower corticosterone and progesterone plasma levels and decreased methylation in the medial amygdala vasopressin system. Horm. Behav. 2014. [Google Scholar] [CrossRef] [PubMed]
- Schwabe, L.; Dalm, S.; Schächinger, H.; Oitzl, M.S. Chronic stress modulates the use of spatial and stimulus-response learning strategies in mice and man. Neurobiol. Learn. Mem. 2008, 90, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, E.; Perry, S.W.; Licinio, J.; Wong, M.L.; Dremencov, E.; Zavjalov, E.L.; Shevelev, O.B.; Khotskin, N.V.; Koncevaya, G.V.; Khotshkina, A.S.; et al. From allostatic load to allostatic state—An endogenous sympathetic strategy to deal with chronic anxiety and stress? Front. Behav. Neurosci. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, S.E.; Girgenti, M.J.; Davis, M.T.; Pietrzak, R.H.; DellaGioia, N.; Nabulsi, N.; Matuskey, D.; Southwick, S.; Duman, R.S.; Carson, R.E.; et al. Altered metabotropic glutamate receptor 5 markers in PTSD: In vivo and postmortem evidence. Proc. Natl. Acad. Sci. USA 2017, 114, 8390–8395. [Google Scholar] [CrossRef] [Green Version]
- Korte, S.M.; Meijer, O.C.; de Kloet, E.R.; Buwalda, B.; Keijser, J.; Sluyter, F.; van Oortmerssen, G.; Bohus, B. Enhanced 5-HT1A receptor expression in forebrain regions of aggressive house mice. Brain Res. 1996, 736, 338–343. [Google Scholar] [CrossRef]
- McEwen, B.S. Neurobiological and Systemic Effects of Chronic Stress. Chronic Stress 2017, 1, 2470547017692328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, M.J. Individual differences in response to stress. Am. J. Community Psychol. 1973, 1, 113–137. [Google Scholar] [CrossRef]
- Ebner, K.; Singewald, N. Individual differences in stress susceptibility and stress inhibitory mechanisms. Curr. Opin. Behav. Sci. 2017, 14, 54–64. [Google Scholar] [CrossRef]
- Dopfel, D.; Perez, P.D.; Verbitsky, A.; Bravo-Rivera, H.; Ma, Y.; Quirk, G.J.; Zhang, N. Individual variability in behavior and functional networks predicts vulnerability using an animal model of PTSD. Nat. Commun. 2019, 10, 2372. [Google Scholar] [CrossRef]
- Yehuda, R.; Flory, J.D.; Pratchett, L.C.; Buxbaum, J.; Ising, M.; Holsboer, F. Putative biological mechanisms for the association between early life adversity and the subsequent development of PTSD. Psychopharmacology (Berl) 2010, 212, 405–417. [Google Scholar] [CrossRef]
- Yehuda, R.; LeDoux, J. Response variation following trauma: A translational neuroscience approach to understanding PTSD. Neuron 2007, 56, 19–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koolhaas, J.M. Coping style and immunity in animals: Making sense of individual variation. Brain Behav. Immun. 2008, 22, 662–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feder, A.; Nestler, E.J.; Charney, D.S. Psychobiology and molecular genetics of resilience. Nat. Rev. Neurosci. 2009, 10, 446–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapp, H.E.; Ahmed, S.; Moore, C.L.; Hunter, R.G. Toxic stress history and hypothalamic-pituitary-adrenal axis function in a social stress task: Genetic and epigenetic factors. Neurotoxicol. Teratol. 2019, 71, 41–49. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhu, G.; Hariri, A.R.; Enoch, M.-A.; Scott, D.; Sinha, R.; Virkkunen, M.; Mash, D.C.; Lipsky, R.H.; Hu, X.-Z.; et al. Genetic variation in human NPY expression affects stress response and emotion. Nature 2008, 452, 997–1001. [Google Scholar] [CrossRef] [Green Version]
- Korte, S.M.; Bouws, G.A.H.; Koolhaas, J.M.; Bohus, B. Neuroendocrine and behavioral responses during conditioned active and passive behavior in the defensive burying/probe avoidance paradigm: Effects of ipsapirone. Physiol. Behav. 1992, 52, 355–361. [Google Scholar] [CrossRef] [Green Version]
- Gregus, A.; Wintink, A.J.; Davis, A.C.; Kalynchuk, L.E. Effect of repeated corticosterone injections and restraint stress on anxiety and depression-like behavior in male rats. Behav. Brain Res. 2005, 156, 105–114. [Google Scholar] [CrossRef]
- Popoli, M.; Yan, Z.; McEwen, B.S.; Sanacora, G. The stressed synapse: The impact of stress and glucocorticoids on glutamate transmission. Nat. Rev. Neurosci. 2011, 13, 22–37. [Google Scholar] [CrossRef] [Green Version]
- Dunn, A.J.; Swiergiel, A.H. The role of corticotropin-releasing factor and noradrenaline in stress-related responses, and the inter-relationships between the two systems. Eur. J. Pharmacol. 2008, 583, 186–193. [Google Scholar] [CrossRef] [Green Version]
- Cuadra, G.; Zurita, A.; Lacerra, C.; Molina, V. Chronic stress sensitizes frontal cortex dopamine release in response to a subsequent novel stressor: Reversal by naloxone. Brain Res. Bull. 1999, 48, 303–308. [Google Scholar] [CrossRef]
- Gardner, K.L.; Hale, M.W.; Lightman, S.L.; Plotsky, P.M.; Lowry, C.A. Adverse early life experience and social stress during adulthood interact to increase serotonin transporter mRNA expression. Brain Res. 2009, 1305, 47–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tottenham, N.; Galván, A. Stress and the adolescent brain: Amygdala-prefrontal cortex circuitry and ventral striatum as developmental targets. Neurosci. Biobehav. Rev. 2016, 70, 217–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina, A.C.; Charles, J.R.; Espinoza-González, V.; Sánchez-Resendis, O.; Prado-Alcalá, R.A.; Roozendaal, B.; Quirarte, G.L. Glucocorticoid administration into the dorsal striatum corrected facilitates memory consolidation of inhibitory avoidance training but not of the context or footshock components. Learn. Mem. 2007, 14, 673–677. [Google Scholar] [CrossRef] [Green Version]
- Quirarte, G.L.; de La Teja, I.S.L.; Casillas, M.; Serafín, N.; Prado-Alcalá, R.A.; Roozendaal, B. Corticosterone infused into the dorsal striatum selectively enhances memory consolidation of cued water-maze training. Learn. Mem. 2009, 16, 586–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwabe, L.; Wolf, O.T. Stress modulates the engagement of multiple memory systems in classification learning. J. Neurosci. 2012, 32, 11042–11049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siller-Pérez, C.; Fuentes-Ibañez, A.; Sotelo-Barrera, E.L.; Serafín, N.; Prado-Alcalá, R.A.; Campolongo, P.; Roozendaal, B.; Quirarte, G.L. Glucocorticoid interactions with the dorsal striatal endocannabinoid system in regulating inhibitory avoidance memory. Psychoneuroendocrinology 2019, 99, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Ono, Y.; Lin, H.-C.; Tzen, K.-Y.; Chen, H.-H.; Yang, P.-F.; Lai, W.-S.; Chen, J.-H.; Onozuka, M.; Yen, C.-T. Active coping with stress suppresses glucose metabolism in the rat hypothalamus. Stress 2012, 15, 207–217. [Google Scholar] [CrossRef]
- Edwards, C.R.W.; Burt, D.; Mcintyre, M.A.; de Kloet, E.R.; Stewart, P.M.; Brett, L.; Sutanto, W.S.; Monder, C. Localisation of 11β-Hydroxysteroid Dehydrogenase—Tissue Specific Protector of the Mineralocorticoid Receptor. Lancet 1988, 332, 986–989. [Google Scholar] [CrossRef]
- Funder, J.W.; Pearce, P.T.; Smith, R.; Smith, A.I. Mineralocorticoid action: Target tissue specificity is enzyme, not receptor, mediated. Science 1988, 242, 583–585. [Google Scholar] [CrossRef]
- Goodman, J.; Leong, K.-C.; Packard, M.G. Emotional modulation of multiple memory systems: Implications for the neurobiology of post-traumatic stress disorder. Rev. Neurosci. 2012, 23, 627–643. [Google Scholar] [CrossRef]
- Schwabe, L.; Tegenthoff, M.; Höffken, O.; Wolf, O.T. Mineralocorticoid receptor blockade prevents stress-induced modulation of multiple memory systems in the human brain. Biol. Psychiatry 2013, 74, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.; Lacadie, C.M.; Constable, R.T.; Seo, D. Dynamic neural activity during stress signals resilient coping. Proc. Natl. Acad. Sci. USA 2016, 113, 8837–8842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolova, Y.S.; Bogdan, R.; Brigidi, B.D.; Hariri, A.R. Ventral striatum reactivity to reward and recent life stress interact to predict positive affect. Biol. Psychiatry 2012, 72, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Corral-Frías, N.S.; Nikolova, Y.S.; Michalski, L.J.; Baranger, D.A.A.; Hariri, A.R.; Bogdan, R. Stress-related anhedonia is associated with ventral striatum reactivity to reward and transdiagnostic psychiatric symptomatology. Psychol. Med. 2015, 45, 2605–2617. [Google Scholar] [CrossRef] [Green Version]
- Fareri, D.S.; Tottenham, N. Effects of early life stress on amygdala and striatal development. Dev. Cogn. Neurosci. 2016, 19, 233–247. [Google Scholar] [CrossRef] [Green Version]
- Heller, A.S.; Johnstone, T.; Light, S.N.; Peterson, M.J.; Kolden, G.G.; Kalin, N.H.; Davidson, R.J. Relationships between changes in sustained fronto-striatal connectivity and positive affect in major depression resulting from antidepressant treatment. Am. J. Psychiatry 2013, 170, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Gatzke-Kopp, L.M.; Beauchaine, T.P.; Shannon, K.E.; Chipman, J.; Fleming, A.P.; Crowell, S.E.; Liang, O.; Johnson, L.C.; Aylward, E. Neurological correlates of reward responding in adolescents with and without externalizing behavior disorders. J. Abnorm. Psychol. 2009, 118, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Taghzouti, K.; Lamarque, S.; Kharouby, M.; Simon, H. Interindividual differences in active and passive behaviors in the forced-swimming test: Implications for animal models of psychopathology. Biol. Psychiatry 1999, 45, 750–758. [Google Scholar] [CrossRef]
- Jama, A.; Cecchi, M.; Calvo, N.; Watson, S.J.; Akil, H. Inter-individual differences in novelty-seeking behavior in rats predict differential responses to desipramine in the forced swim test. Psychopharmacology 2008, 198, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Flores-Serrano, A.G.; Vila-Luna, M.L.; Álvarez-Cervera, F.J.; Heredia-López, F.J.; Góngora-Alfaro, J.L.; Pineda, J.C. Clinical doses of citalopram or reboxetine differentially modulate passive and active behaviors of female Wistar rats with high or low immobility time in the forced swimming test. Pharmacol. Biochem. Behav. 2013, 110, 89–97. [Google Scholar] [CrossRef]
- Shin, L.M.; Liberzon, I. The neurocircuitry of fear, stress, and anxiety disorders. Neuropsychopharmacology 2010, 35, 169–191. [Google Scholar] [CrossRef]
- Kalin, N.H.; Shelton, S.E.; Rickman, M.; Davidson, R.J. Individual differences in freezing and cortisol in infant and mother rhesus monkeys. Behav. Neurosci. 1998, 112, 251–254. [Google Scholar] [CrossRef] [PubMed]
- Núñez, J.F.; Ferré, P.; Escorihuela, R.M.; Tobeña, A.; Fernández-Teruel, A. Effects of Postnatal Handling of Rats on Emotional, HPA-Axis, and Prolactin Reactivity to Novelty and Conflict. Physiol. Behav. 1996, 60, 1355–1359. [Google Scholar] [CrossRef]
- Takahashi, L.K.; Rubin, W.W. Corticosteroid induction of threat-induced behavioral inhibition in preweanling rats. Behav. Neurosci. 1993, 107, 860–866. [Google Scholar] [CrossRef] [PubMed]
- Avinun, R.; Nevo, A.; Knodt, A.R.; Elliott, M.L.; Radtke, S.R.; Brigidi, B.D.; Hariri, A.R. Reward-Related Ventral Striatum Activity Buffers against the Experience of Depressive Symptoms Associated with Sleep Disturbances. J. Neurosci. 2017, 37, 9724–9729. [Google Scholar] [CrossRef] [PubMed]
- Shimojo, M.; Whorwood, C.B.; Stewart, P.M. 11 beta-Hydroxysteroid dehydrogenase in the rat adrenal. J. Mol. Endocrinol. 1996, 17, 121–130. [Google Scholar] [CrossRef]
- Kotelevtsev, Y.; Holmes, M.C.; Burchell, A.; Houston, P.M.; Schmoll, D.; Jamieson, P.; Best, R.; Brown, R.; Edwards, C.R.; Seckl, J.R.; et al. 11beta-hydroxysteroid dehydrogenase type 1 knockout mice show attenuated glucocorticoid-inducible responses and resist hyperglycemia on obesity or stress. Proc. Natl. Acad. Sci. USA 1997, 94, 14924–14929. [Google Scholar] [CrossRef] [Green Version]
- Pérez, J.H.; Swanson, R.E.; Lau, H.J.; Cheah, J.; Bishop, V.R.; Snell, K.R.S.; Reid, A.M.A.; Meddle, S.L.; Wingfield, J.C.; Krause, J.S. Tissue-specific expression of 11β-HSD and its effects on plasma corticosterone during the stress response. J. Exp. Biol. 2020, 223, jeb209346. [Google Scholar] [CrossRef]
- Lecybyl, R.; Jagodzinski, P.P.; Krozowski, Z.S.; Trzeciak, W.H. Regulation of 11beta-hydroxysteroid dehydrogenase type II in rat adrenocortical cells. Endocr. Res. 1998, 24, 643–647. [Google Scholar] [CrossRef]
- Estanislau, C. Cues to the usefulness of grooming behavior in the evaluation of anxiety in the elevated plus-maze. Psychol. Neurosci. 2012, 5, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Le Moëne, O.; Ågmo, A. The neuroendocrinology of sexual attraction. Front. Neuroendocrinol. 2018, 51, 46–67. [Google Scholar] [CrossRef] [PubMed]
- Manukhina, E.B.; Tseilikman, V.E.; Tseilikman, O.B.; Komelkova, M.V.; Kondashevskaya, M.V.; Goryacheva, A.V.; Lapshin, M.S.; Platkovskii, P.O.; Alliluev, A.V.; Downey, H.F. Intermittent hypoxia improves behavioral and adrenal gland dysfunction induced by posttraumatic stress disorder in rats. J. Appl. Physiol. 2018, 125, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Kulikov, V.A.; Khotskin, N.V.; Nikitin, S.V.; Lankin, V.S.; Kulikov, A.V.; Trapezov, O.V. Application of 3-D imaging sensor for tracking minipigs in the open field test. J. Neurosci. Methods 2014, 235, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Serova, L.I.; Laukova, M.; Alaluf, L.G.; Pucillo, L.; Sabban, E.L. Intranasal neuropeptide Y reverses anxiety and depressive-like behavior impaired by single prolonged stress PTSD model. Eur. Neuropsychopharmacol. 2014, 24, 142–147. [Google Scholar] [CrossRef]
- Lapiz-Bluhm, M.D.S.; Bondi, C.O.; Doyen, J.; Rodriguez, G.A.; Bédard-Arana, T.; Morilak, D.A. Behavioural assays to model cognitive and affective dimensions of depression and anxiety in rats. J. Neuroendocrinol. 2008, 20, 1115–1137. [Google Scholar] [CrossRef] [Green Version]
- Gruetter, R. Automatic, localized in vivo adjustment of all first- and second-order shim coils. Magn. Reson. Med. 1993, 29, 804–811. [Google Scholar] [CrossRef]
- Tkac, I.; Starcuk, Z.; Choi, I.Y.; Gruetter, R. In vivo 1H NMR spectroscopy of rat brain at 1 ms echo time. Magn. Reson. Med. 1999, 41, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Provencher, S.W. Estimation of metabolite concentrations from localized in vivo proton NMR spectra. Magn. Reson. Med. 1993, 30, 672–679. [Google Scholar] [CrossRef]
- Moshkin, M.P.; Akulov, A.E.; Petrovski, D.V.; Saik, O.V.; Petrovskiy, E.D.; Savelov, A.A.; Koptyug, I.V. Proton magnetic resonance spectroscopy of brain metabolic shifts induced by acute administration of 2-deoxy-d-glucose and lipopolysaccharides. NMR Biomed. 2014, 27, 399–405. [Google Scholar] [CrossRef]
- Selyatitskaya, V.G.; Cherkasova, O.P.; Pankina, T.V.; Palchikova, N.A. Functional state of adrenocortical system in rats with manifest alloxan-induced diabetes mellitus. Bull. Exp. Biol. Med. 2008, 146, 708–710. [Google Scholar] [CrossRef]
- Baĭkova, L.A.; Fedorov, V.I.; Cherkasova, O.P. Analiz kortikosteroidov plazmy krovi metodom mikrokolonochnoĭ zhidkostnoĭ khromatografii. Lab. Delo 1989, 5, 57–60. [Google Scholar]
- Baram, G.I.; Grachev, M.A.; Komarova, N.I.; Perelroyzen, M.P.; Bolvanov, Y.A.; Kuzmin, S.V.; Kargaltsev, V.V.; Kuper, E.A. Micro-column liquid chromatography with multi-wave-length photometric detection. J. Chromatogr. A 1983, 264, 69–90. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | APR | AFR | |
|---|---|---|---|
| Central square time | 0.10 ± 0.02 | 0.12 ± 0.02 | 0.19 ± 0.01 *,# |
| % Closed arms time | 0.89 ± 0.03 | 0.8 ± 0.03 | 0.62 ± 0.03 *,# |
| % Open arms time | 0.11 ± 0.01 | 0.08 ± 0.01 | 0.19 ± 0.03 *,# |
| Entries into open arms | 1.32 ± 0.24 | 1.15 ± 0.024 | 4.42 ± 0.45 *,# |
| Entries into closed arms | 4.55 ± 0.78 | 8.68 ± 1.25 | 5.43 ± 0.35 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ullmann, E.; Chrousos, G.; Perry, S.W.; Wong, M.-L.; Licinio, J.; Bornstein, S.R.; Tseilikman, O.; Komelkova, M.; Lapshin, M.S.; Vasilyeva, M.; et al. Offensive Behavior, Striatal Glutamate Metabolites, and Limbic–Hypothalamic–Pituitary–Adrenal Responses to Stress in Chronic Anxiety. Int. J. Mol. Sci. 2020, 21, 7440. https://doi.org/10.3390/ijms21207440
Ullmann E, Chrousos G, Perry SW, Wong M-L, Licinio J, Bornstein SR, Tseilikman O, Komelkova M, Lapshin MS, Vasilyeva M, et al. Offensive Behavior, Striatal Glutamate Metabolites, and Limbic–Hypothalamic–Pituitary–Adrenal Responses to Stress in Chronic Anxiety. International Journal of Molecular Sciences. 2020; 21(20):7440. https://doi.org/10.3390/ijms21207440
Chicago/Turabian StyleUllmann, Enrico, George Chrousos, Seth W. Perry, Ma-Li Wong, Julio Licinio, Stefan R. Bornstein, Olga Tseilikman, Maria Komelkova, Maxim S. Lapshin, Maryia Vasilyeva, and et al. 2020. "Offensive Behavior, Striatal Glutamate Metabolites, and Limbic–Hypothalamic–Pituitary–Adrenal Responses to Stress in Chronic Anxiety" International Journal of Molecular Sciences 21, no. 20: 7440. https://doi.org/10.3390/ijms21207440