Docosahexaenoic Acid-Loaded Polylactic Acid Core-Shell Nanofiber Membranes for Regenerative Medicine after Spinal Cord Injury: In Vitro and In Vivo Study

 ,
,
Abstract
:1. Introduction
2. Results
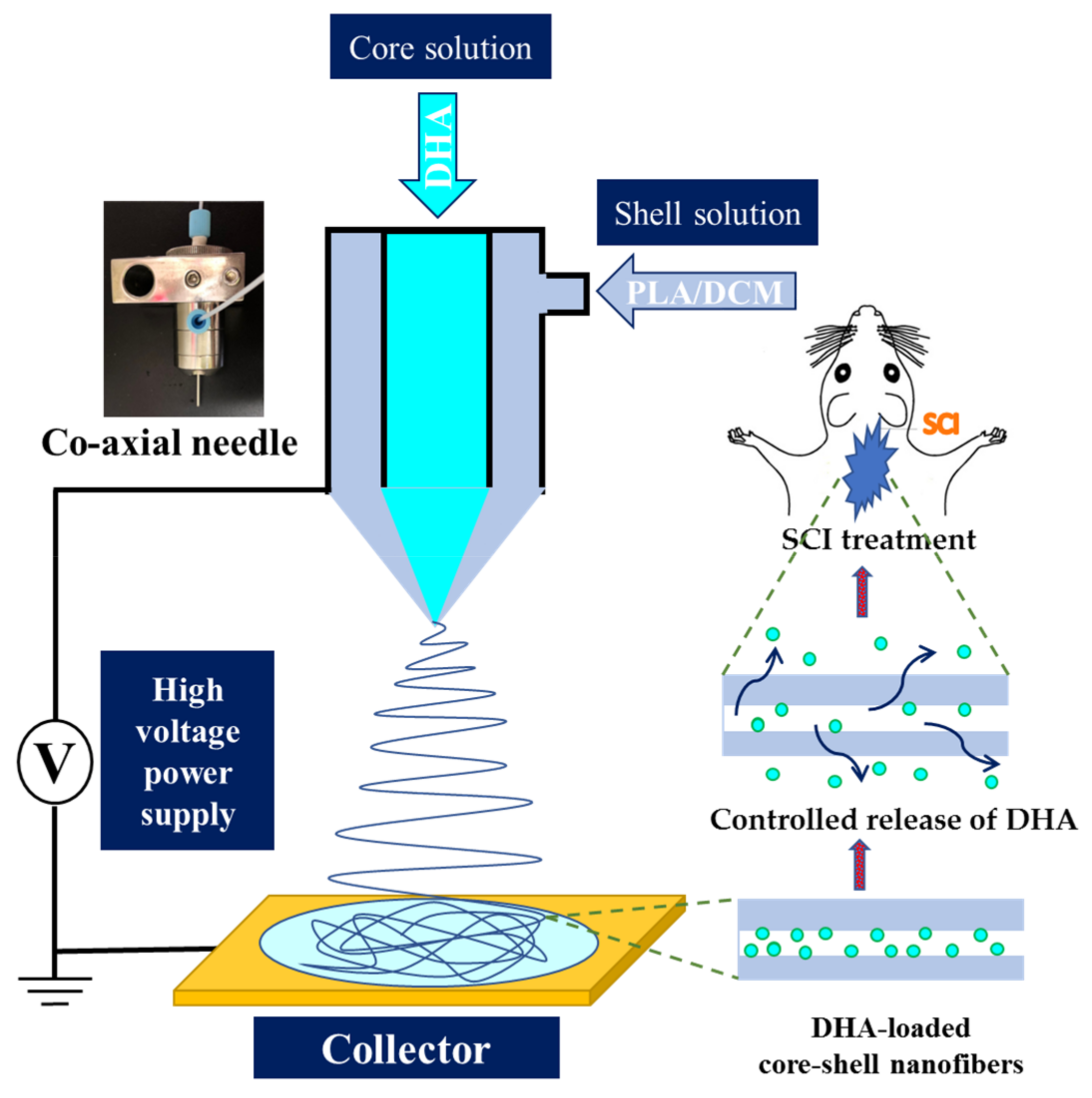
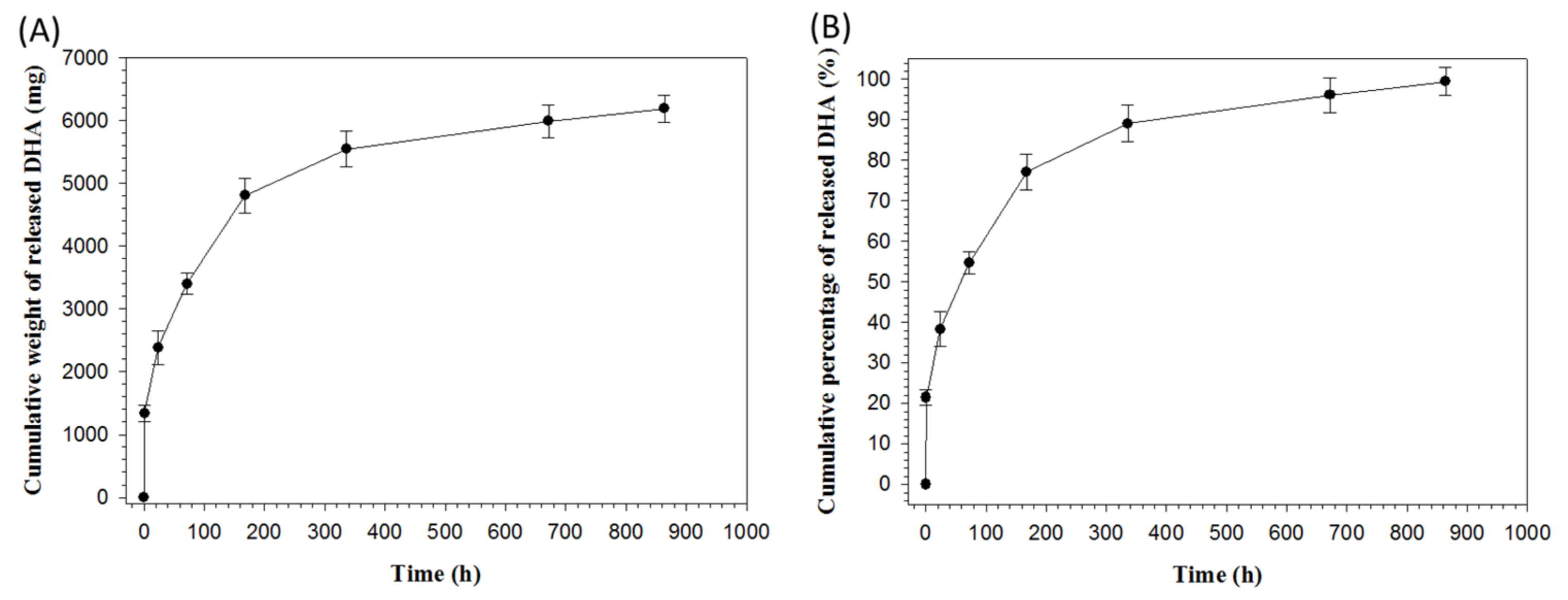
2.1. Preparation and Characterization of Core-Shell Nanofiber Membrane (CSNM)
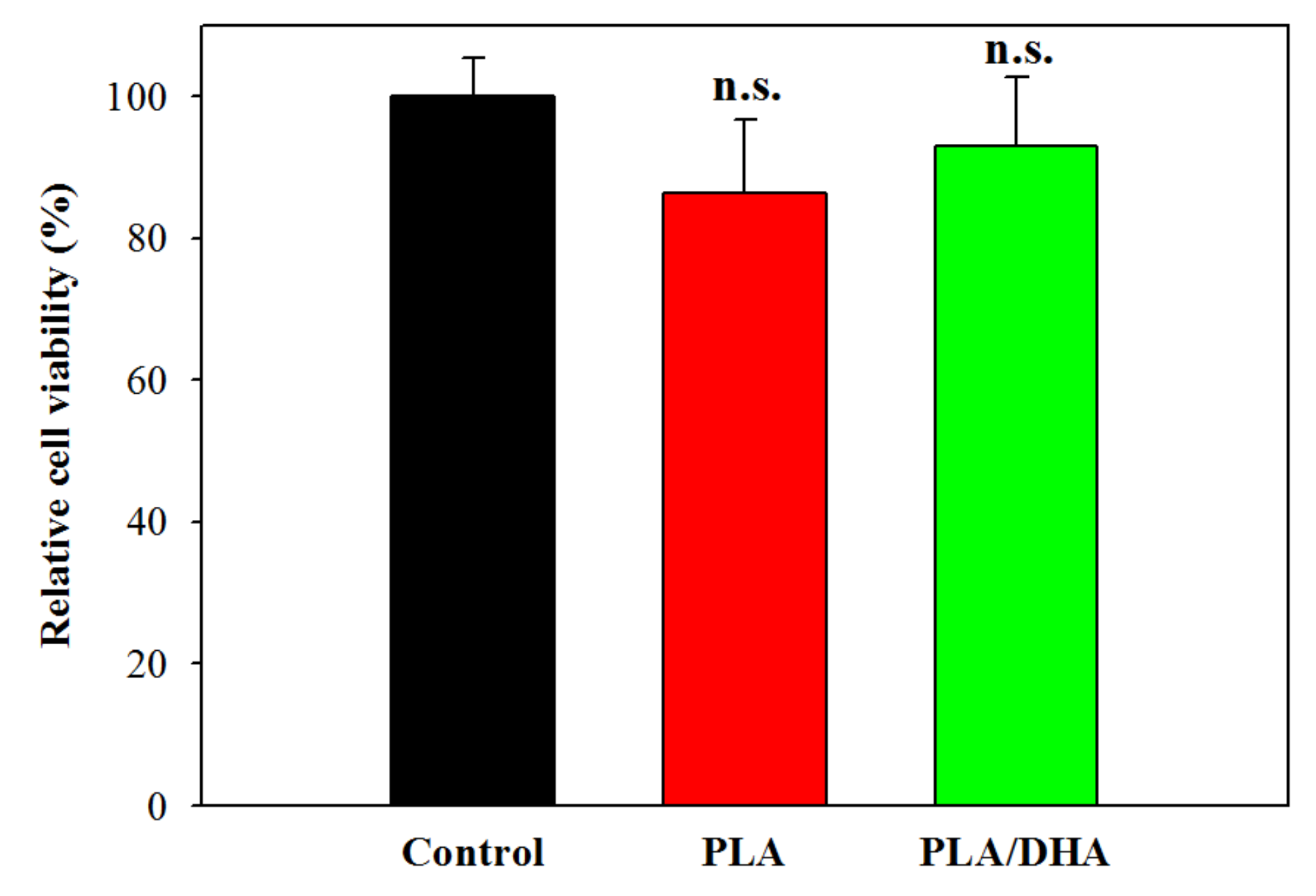
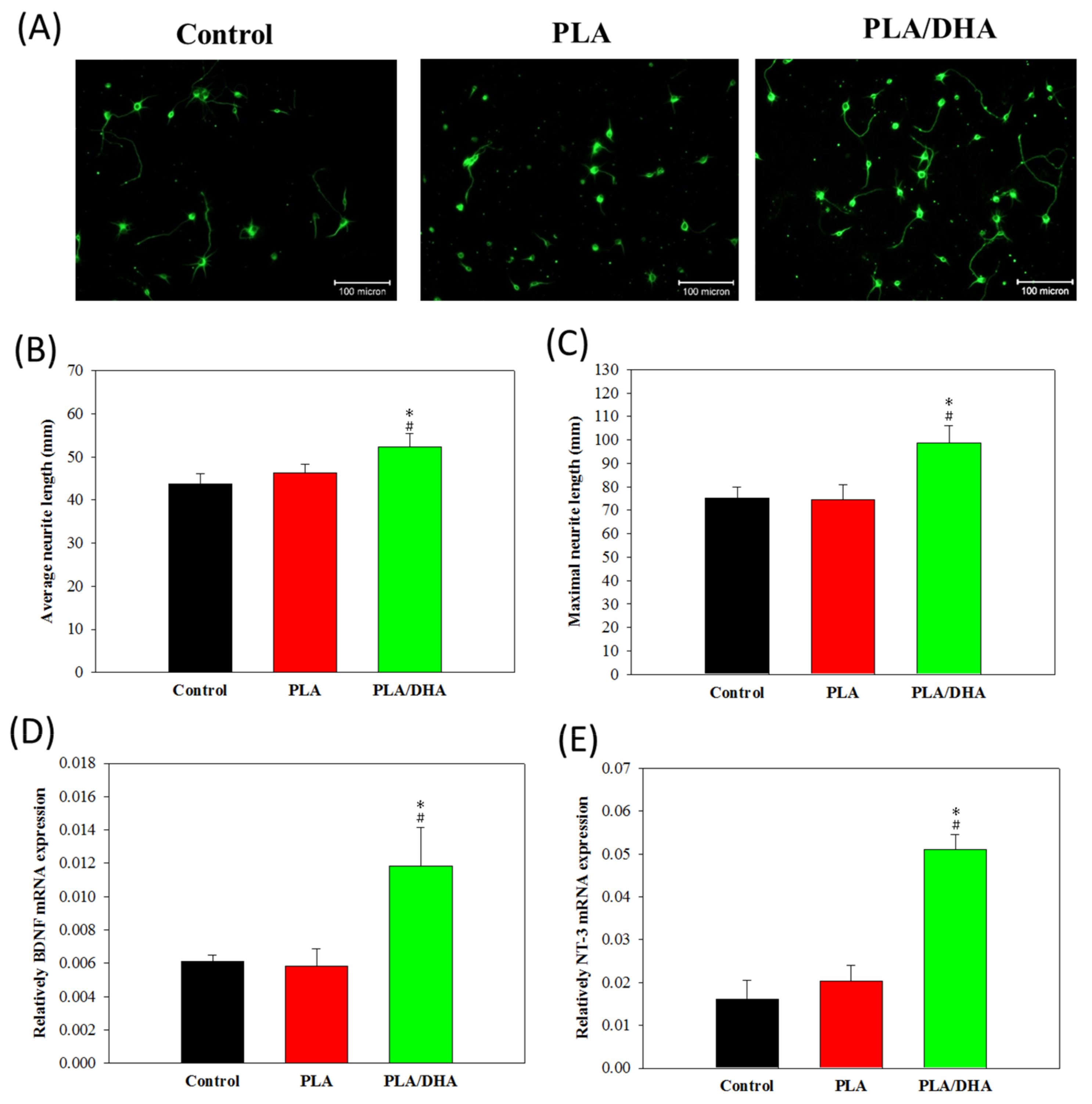
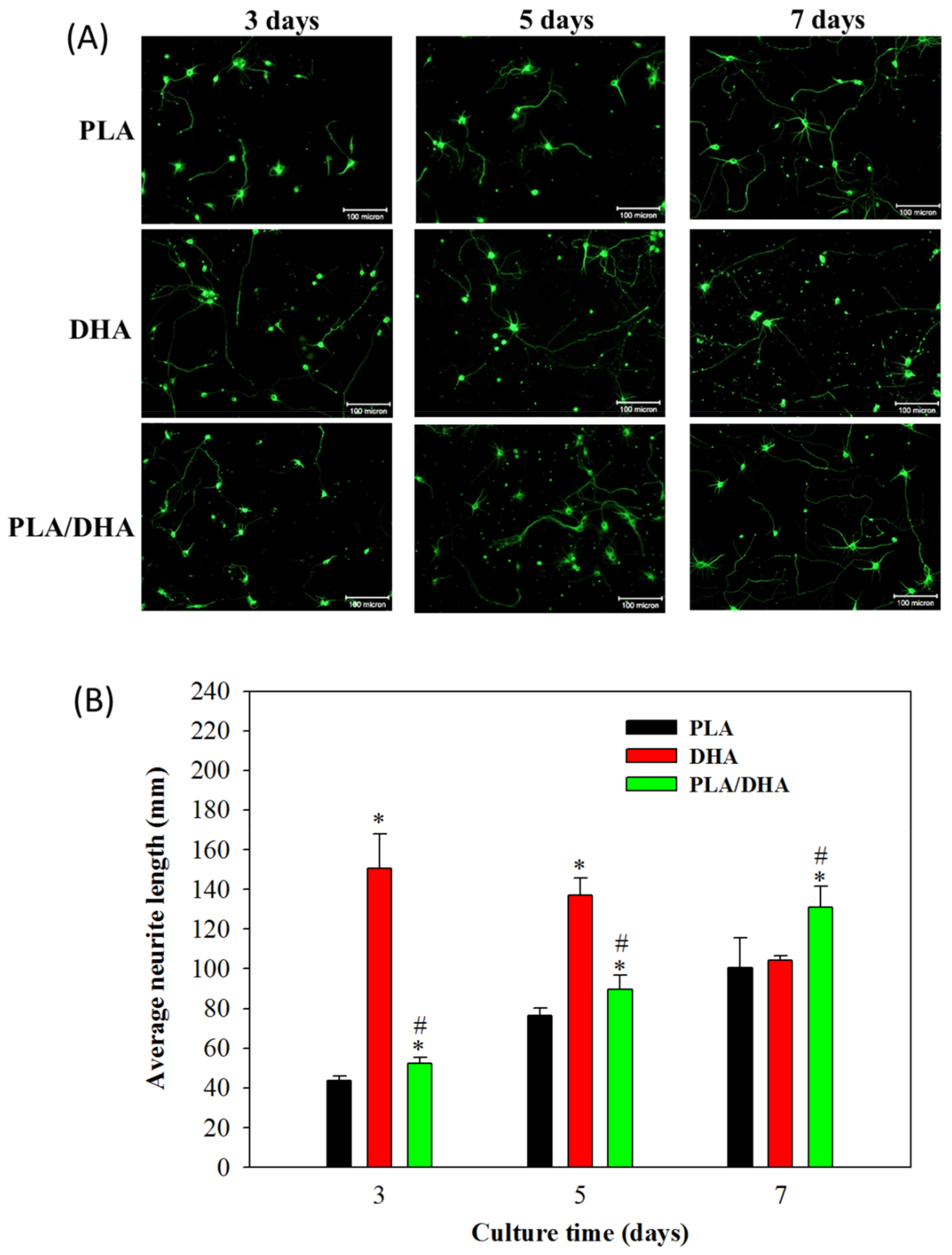
2.2. In Vitro Cell Culture
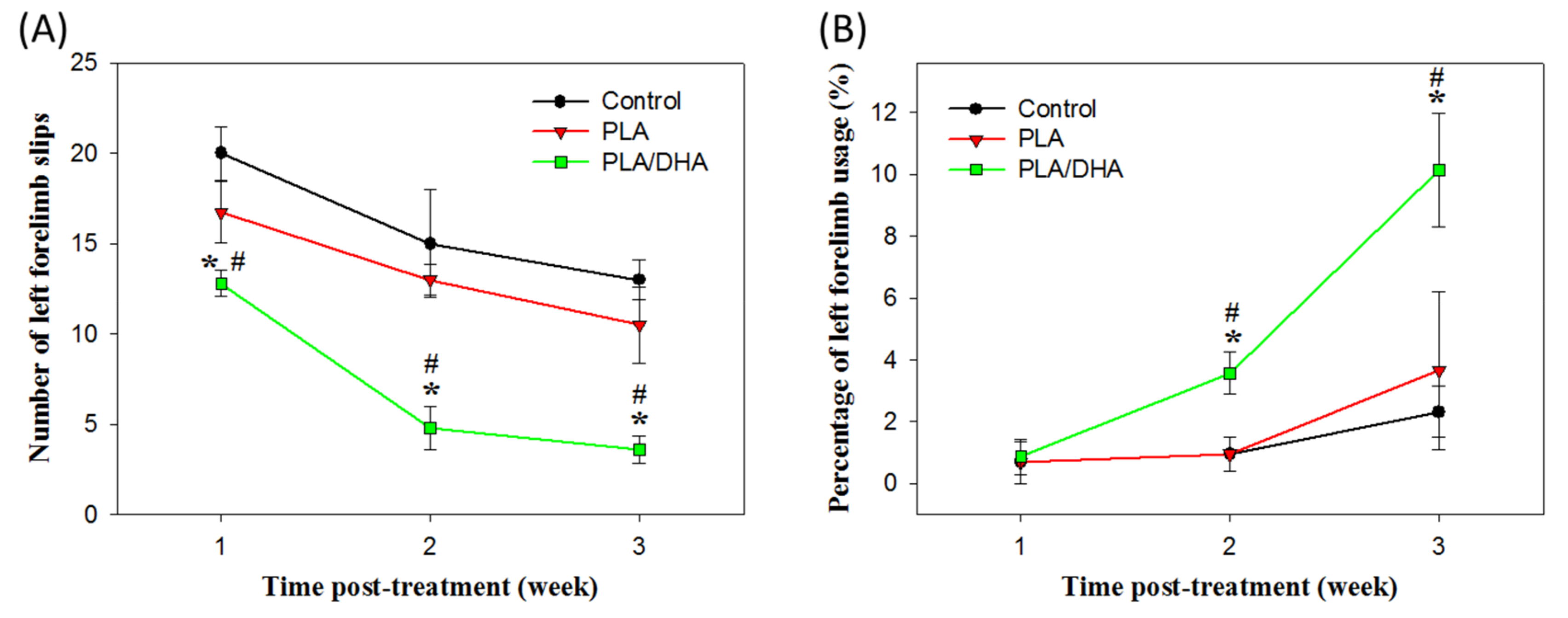
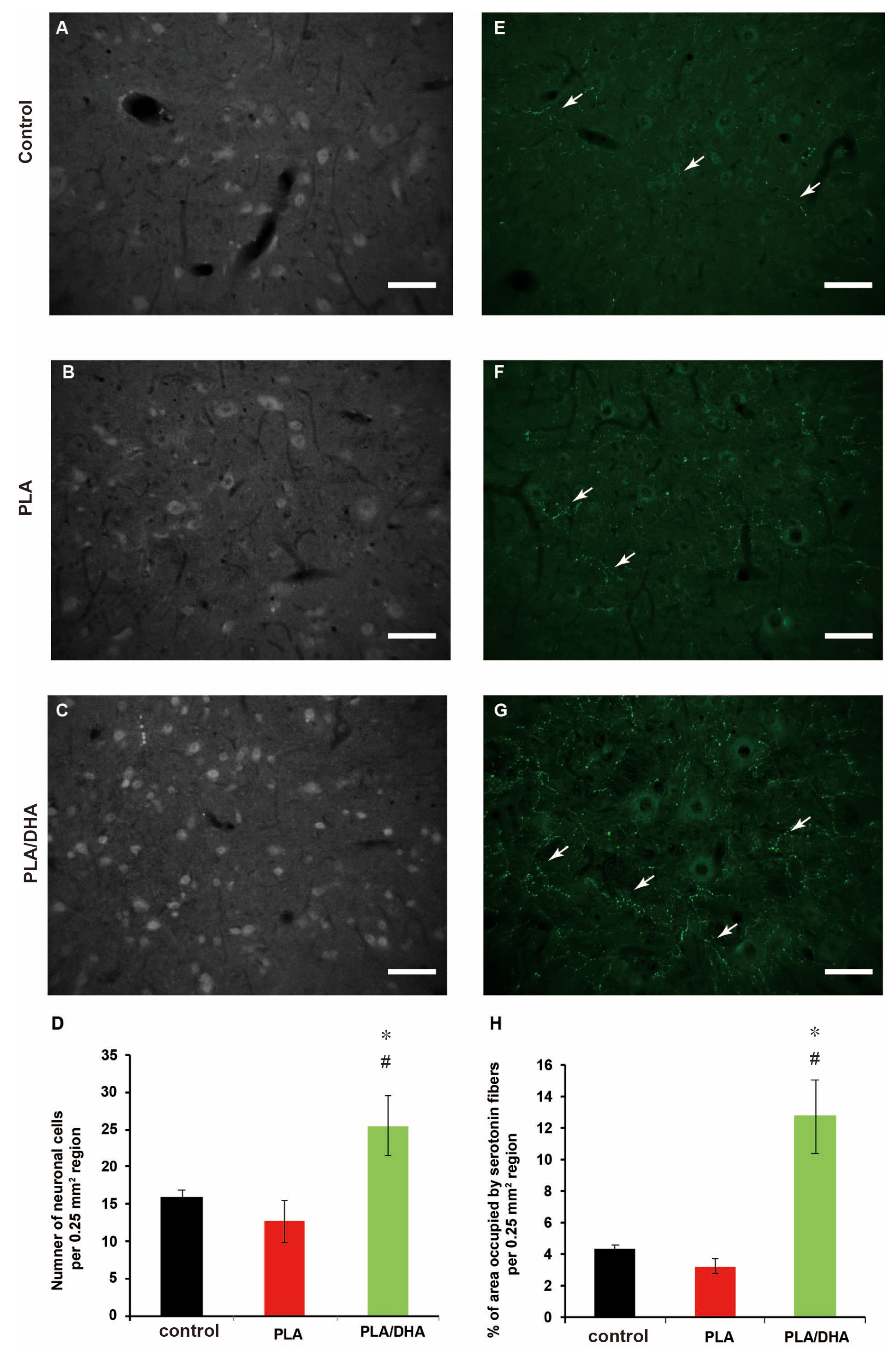
2.3. In Vivo Study
3. Discussion
4. Materials and Methods
4.1. Preparation of Core-Shell Nanofiber Membrane (CSNM)
4.2. Characterization of Nanofiber Membrane
4.3. Primary Cortical Cell Culture
4.4. Cytotoxicity Evaluation
4.5. Neurite Outgrowth Evaluation
4.6. qRT-PCR to Detect BDNF and NT-3 mRNA Expression
4.7. Animal Surgery
4.8. Grid Exploration Test
4.9. Cylinder Test
4.10. Histological Analysis
4.11. Image Analysis of Histological Sections
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schwab, M.E. Repairing the injured spinal cord. Science 2002, 295, 1029–1031. [Google Scholar] [CrossRef] [Green Version]
- Thuret, S.; Moon, L.D.; Gage, F.H. Therapeutic interventions after spinal cord injury. Nat. Rev. Neurosci. 2006, 7, 628–643. [Google Scholar] [CrossRef]
- Pardridge, W.M. Drug transport across the blood-brain barrier. J. Cereb. Blood Flow Metab. 2012, 32, 1959–1972. [Google Scholar] [CrossRef] [PubMed]
- Tsintou, M.; Dalamagkas, K.; Seifalian, A.M. Advances in regenerative therapies for spinal cord injury: A biomaterials approach. Neural Regen. Res. 2015, 10, 726–742. [Google Scholar] [CrossRef] [PubMed]
- Straley, K.S.; Foo, C.W.; Heilshorn, S.C. Biomaterial design strategies for the treatment of spinal cord injuries. J. Neurotrauma 2010, 27, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, M.; Eslamifar, M.; Khezri, K.; Dizaj, S.M. Applications of nanotechnology in drug delivery to the central nervous system. Biomed. Pharmacother. 2019, 111, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Salem, N., Jr.; Hullin, F.; Yoffe, A.M.; Karanian, J.W.; Kim, H.Y. Fatty acid and phospholipid species composition of rat tissues after a fish oil diet. Adv. Prostaglandinthromboxaneand Leukot. Res. 1989, 19, 618–622. [Google Scholar]
- Horrocks, L.A.; Farooqui, A.A. Docosahexaenoic acid in the diet: Its importance in maintenance and restoration of neural membrane function. Prostaglandinsleukotrienesand Essent. Fat. Acids 2004, 70, 361–372. [Google Scholar] [CrossRef]
- Michael-Titus, A.T. Omega-3 fatty acids: Their neuroprotective and regenerative potential in traumatic neurological injury. Clin. Lipidol. 2009, 4, 343–353. [Google Scholar] [CrossRef]
- Noaghiul, S.; Hibbeln, J.R. Cross-national comparisons of seafood consumption and rates of bipolar disorders. Am. J. Psychiatry 2003, 160, 2222–2227. [Google Scholar] [CrossRef]
- Conquer, J.A.; Tierney, M.C.; Zecevic, J.; Bettger, W.J.; Fisher, R.H. Fatty acid analysis of blood plasma of patients with Alzheimer’s disease, other types of dementia, and cognitive impairment. Lipids 2000, 35, 1305–1312. [Google Scholar] [PubMed]
- Lang-Lazdunski, L.; Blondeau, N.; Jarretou, G.; Lazdunski, M.; Heurteaux, C. Linolenic acid prevents neuronal cell death and paraplegia after transient spinal cord ischemia in rats. J. Vasc. Surg. 2003, 38, 564–575. [Google Scholar] [PubMed] [Green Version]
- Calon, F.; Cole, G. Neuroprotective action of omega-3 polyunsaturated fatty acids against neurodegenerative diseases: Evidence from animal studies. Prostaglandinsleukotrienesand Essent. Fat. Acids 2007, 77, 287–293. [Google Scholar] [CrossRef]
- Morris, M.C.; Evans, D.A.; Bienias, J.L.; Tangney, C.C.; Bennett, D.A.; Wilson, R.S.; Aggarwal, N.; Schneider, J. Consumption of fish and n-3 fatty acids and risk of incident Alzheimer disease. Arch. Neurol. 2003, 60, 940–946. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.L.; King, V.R.; Curran, O.E.; Dyall, S.C.; Ward, R.E.; Lal, N.; Priestley, J.V.; Michael-Titus, A.T. A combination of intravenous and dietary docosahexaenoic acid significantly improves outcome after spinal cord injury. Brain 2007, 130, 3004–3019. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, J.D.; Cordero, K.; Llan, M.S.; De Leon, M. Dietary omega-3 polyunsaturated fatty acids improve the neurolipidome and restore the DHA status while promoting functional recovery after experimental spinal cord injury. J. Neurotrauma 2013, 30, 853–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, P.J.; Noronha, J.; DeGeorge, J.J.; Freed, L.M.; Nariai, T.; Rapoport, S.I. A quantitative method for measuring regional in vivo fatty-acid incorporation into and turnover within brain phospholipids: Review and critical analysis. Brain Res. Brain Res. Rev. 1992, 17, 187–214. [Google Scholar] [CrossRef]
- Rapoport, S.I. In vivo fatty acid incorporation into brain phosholipids in relation to plasma availability, signal transduction and membrane remodeling. J. Mol. Neurosci. 2001, 16, 243–261, discussion 279–284. [Google Scholar] [CrossRef]
- Rapoport, S.I. In vivo approaches to quantifying and imaging brain arachidonic and docosahexaenoic acid metabolism. J. Pediatrics 2003, 143, S26–S34. [Google Scholar]
- Moomand, K.; Lim, L.-T. Oxidative stability of encapsulated fish oil in electrospun zein fibres. Food Res. Int. 2014, 62, 523–532. [Google Scholar] [CrossRef]
- Yang, H.; Wen, P.; Feng, K.; Zong, M.H.; Lou, W.Y.; Wu, H. Encapsulation of fish oil in a coaxial electrospun nanofibrous mat and its properties. Rsc Adv. 2017, 7, 14939–14946. [Google Scholar] [CrossRef] [Green Version]
- Song, W.; Yu, X.; Markel, D.C.; Shi, T.; Ren, W. Coaxial PCL/PVA electrospun nanofibers: Osseointegration enhancer and controlled drug release device. Biofabrication 2013, 5, 035006. [Google Scholar] [CrossRef] [PubMed]
- Mickova, A.; Buzgo, M.; Benada, O.; Rampichova, M.; Fisar, Z.; Filova, E.; Tesarova, M.; Lukas, D.; Amler, E. Core/shell nanofibers with embedded liposomes as a drug delivery system. Biomacromolecules 2012, 13, 952–962. [Google Scholar] [CrossRef] [PubMed]
- Zupančič, Š. Core-shell nanofibers as drug delivery systems. Acta Pharm. 2019, 69, 131–153. [Google Scholar] [CrossRef] [Green Version]
- Faccendini, A.; Vigani, B.; Rossi, S.; Sandri, G.; Bonferoni, M.C.; Caramella, C.M.; Ferrari, F. Nanofiber Scaffolds as Drug Delivery Systems to Bridge Spinal Cord Injury. Pharmaceuticals 2017, 10, 63. [Google Scholar] [CrossRef] [Green Version]
- Armentano, I.; Dottori, M.; Fortunati, E.; Mattioli, S.; Kenny, J. Biodegradable polymer matrix nanocomposites for tissue engineering: A review. Polym. Degrad. Stab. 2010, 95, 2126–2146. [Google Scholar] [CrossRef]
- Tian, H.; Tang, Z.; Zhuang, X.; Chen, X.; Jing, X. Biodegradable synthetic polymers: Preparation, functionalization and biomedical application. Prog. Polym. Sci. 2012, 37, 237–280. [Google Scholar] [CrossRef]
- Shalumon, K.T.; Anjana, J.; Mony, U.; Jayakumar, R.; Chen, J.-P. Process study, development and degradation behavior of different size scale electrospun poly(caprolactone) and poly(lactic acid) fibers. J. Polym. Res. 2018, 25, 82. [Google Scholar] [CrossRef]
- Sandrow, H.R.; Shumsky, J.S.; Amin, A.; Houle, J.D. Aspiration of a cervical spinal contusion injury in preparation for delayed peripheral nerve grafting does not impair forelimb behavior or axon regeneration. Exp. Neurol. 2008, 210, 489–500. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.H.; Yip, P.K.; Adams, L.; Davies, M.; Lee, J.W.; Michael, G.J.; Priestley, J.V.; Michael-Titus, A.T. A Single Bolus of Docosahexaenoic Acid Promotes Neuroplastic Changes in the Innervation of Spinal Cord Interneurons and Motor Neurons and Improves Functional Recovery after Spinal Cord Injury. J. Meuroscience 2015, 35, 12733–12752. [Google Scholar] [CrossRef] [Green Version]
- Schallert, T.; Fleming, S.M.; Leasure, J.L.; Tillerson, J.L.; Bland, S.T. CNS plasticity and assessment of forelimb sensorimotor outcome in unilateral rat models of stroke, cortical ablation, parkinsonism and spinal cord injury. Neuropharmacology 2000, 39, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Corey, J.M.; Gertz, C.C.; Wang, B.S.; Birrell, L.K.; Johnson, S.L.; Martin, D.C.; Feldman, E.L. The design of electrospun PLLA nanofiber scaffolds compatible with serum-free growth of primary motor and sensory neurons. Acta Biomater. 2008, 4, 863–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Xie, Y.Y.; Wang, B. Role and prospects of regenerative biomaterials in the repair of spinal cord injury. Neural Regen. Res. 2019, 14, 1352–1363. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.N.; Huang, W.; Hall, J.C.; Michael-Titus, A.T.; Priestley, J.V. Improved outcome after spinal cord compression injury in mice treated with docosahexaenoic acid. Exp. Neurol. 2013, 239, 13–27. [Google Scholar] [CrossRef]
- King, V.R.; Huang, W.L.; Dyall, S.C.; Curran, O.E.; Priestley, J.V.; Michael-Titus, A.T. Omega-3 fatty acids improve recovery, whereas omega-6 fatty acids worsen outcome, after spinal cord injury in the adult rat. J. Neurosci. 2006, 26, 4672–4680. [Google Scholar] [CrossRef] [Green Version]
- Calderon, F.; Kim, H.Y. Docosahexaenoic acid promotes neurite growth in hippocampal neurons. J. Neurochem. 2004, 90, 979–988. [Google Scholar] [CrossRef]
- Cao, D.; Xue, R.; Xu, J.; Liu, Z. Effects of docosahexaenoic acid on the survival and neurite outgrowth of rat cortical neurons in primary cultures. J. Nutr. Biochem. 2005, 16, 538–546. [Google Scholar] [CrossRef]
- Robson, L.G.; Dyall, S.; Sidloff, D.; Michael-Titus, A.T. Omega-3 polyunsaturated fatty acids increase the neurite outgrowth of rat sensory neurones throughout development and in aged animals. Neurobiol. Aging 2010, 31, 678–687. [Google Scholar] [CrossRef]
- Wurtman, R.J.; Ulus, I.H.; Cansev, M.; Watkins, C.J.; Wang, L.; Marzloff, G. Synaptic proteins and phospholipids are increased in gerbil brain by administering uridine plus docosahexaenoic acid orally. Brain Res. 2006, 1088, 83–92. [Google Scholar] [CrossRef]
- Wu, A.; Ying, Z.; Gomez-Pinilla, F. Docosahexaenoic acid dietary supplementation enhances the effects of exercise on synaptic plasticity and cognition. Neuroscience 2008, 155, 751–759. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.; Tuszynski, M.H. Growth factors and combinatorial therapies for CNS regeneration. Exp. Neurol. 2008, 209, 313–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fouad, K.; Bennett, D.J.; Vavrek, R.; Blesch, A. Long-term viral brain-derived neurotrophic factor delivery promotes spasticity in rats with a cervical spinal cord hemisection. Front. Neurol. 2013, 4, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Shine, H.D. Neurotrophic factors expressed in both cortex and spinal cord induce axonal plasticity after spinal cord injury. J. Neurosci. Res. 2003, 74, 221–226. [Google Scholar] [CrossRef]
- Tanabe, Y.; Hashimoto, M.; Sugioka, K.; Maruyama, M.; Fujii, Y.; Hagiwara, R.; Hara, T.; Hossain, S.M.; Shido, O. Improvement of spatial cognition with dietary docosahexaenoic acid is associated with an increase in Fos expression in rat CA1 hippocampus. Clin. Exp. Pharmacol. Physiol. 2004, 31, 700–703. [Google Scholar] [CrossRef]
- Cao, D.; Kevala, K.; Kim, J.; Moon, H.S.; Jun, S.B.; Lovinger, D.; Kim, H.Y. Docosahexaenoic acid promotes hippocampal neuronal development and synaptic function. J. Neurochem. 2009, 111, 510–521. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, H.S.; Agrawal, R.; Sharma, S.; Huo, Y.X.; Ying, Z.; Gomez-Pinilla, F. Omega-3 fatty acid deficiency during brain maturation reduces neuronal and behavioral plasticity in adulthood. PloS ONE 2011, 6, e28451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Ruan, X.Z.; Powis, S.H.; Fernando, R.; Mon, W.Y.; Wheeler, D.C.; Moorhead, J.F.; Varghese, Z. EPA and DHA reduce LPS-induced inflammation responses in HK-2 cells: Evidence for a PPAR-gamma-dependent mechanism. Kidney Int. 2005, 67, 867–874. [Google Scholar] [CrossRef] [Green Version]
- Dyall, S.C.; Michael, G.J.; Michael-Titus, A.T. Omega-3 fatty acids reverse age-related decreases in nuclear receptors and increase neurogenesis in old rats. J. Neurosci. Res. 2010, 88, 2091–2102. [Google Scholar] [CrossRef]
- Meng, B.; Zhang, Q.; Huang, C.; Zhang, H.T.; Tang, T.; Yang, H.L. Effects of a single dose of methylprednisolone versus three doses of rosiglitazone on nerve growth factor levels after spinal cord injury. J. Int. Med Res. 2011, 39, 805–814. [Google Scholar]
- D’Angelo, B.; Benedetti, E.; Di Loreto, S.; Cristiano, L.; Laurenti, G.; Ceru, M.P.; Cimini, A. Signal transduction pathways involved in PPARbeta/delta-induced neuronal differentiation. J. Cell. Physiol. 2011, 226, 2170–2180. [Google Scholar] [CrossRef]
- Wu, A.; Ying, Z.; Gomez-Pinilla, F. Dietary omega-3 fatty acids normalize BDNF levels, reduce oxidative damage, and counteract learning disability after traumatic brain injury in rats. J. Neurotrauma 2004, 21, 1457–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, A.; Ying, Z.; Gomez-Pinilla, F. The salutary effects of DHA dietary supplementation on cognition, neuroplasticity, and membrane homeostasis after brain trauma. J. Neurotrauma 2011, 28, 2113–2122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blondeau, N.; Nguemeni, C.; Debruyne, D.N.; Piens, M.; Wu, X.; Pan, H.; Hu, X.; Gandin, C.; Lipsky, R.H.; Plumier, J.C.; et al. Subchronic alpha-linolenic acid treatment enhances brain plasticity and exerts an antidepressant effect: A versatile potential therapy for stroke. Neuropsychopharmacology 2009, 34, 2548–2559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, R.E.; Huang, W.; Curran, O.E.; Priestley, J.V.; Michael-Titus, A.T. Docosahexaenoic acid prevents white matter damage after spinal cord injury. J. Neurotrauma 2010, 27, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- Saruhashi, Y.; Matsusue, Y.; Fujimiya, M. The recovery of 5-HT transporter and 5-HT immunoreactivity in injured rat spinal cord. Arch. Orthop. Trauma Surg. 2009, 129, 1279–1285. [Google Scholar] [CrossRef]
- Chen, C.-H.; Chen, S.-H.; Shalumon, K.T.; Chen, J.-P. Dual functional core–sheath electrospun hyaluronic acid/polycaprolactone nanofibrous membranes embedded with silver nanoparticles for prevention of peritendinous adhesion. Acta Biomater. 2015, 26, 225–235. [Google Scholar] [CrossRef]
- Lee, R.K.; Wurtman, R.J.; Cox, A.J.; Nitsch, R.M. Amyloid precursor protein processing is stimulated by metabotropic glutamate receptors. Proc. Natl. Acad. Sci. USA 1995, 92, 8083–8087. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Zhou, J.; Xu, Y. Study of the in vitro cytotoxicity testing of medical devices. Biomed. Rep. 2015, 3, 617–620. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.H.; Yip, P.K.; Priestley, J.V.; Michael-Titus, A.T. a single dose of docosahexaenoic acid increases the functional recovery promoted by rehabilitation after cervical spinal cord injury in the rat. J. Neurotrauma 2017, 34, 1766–1777. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Primer (5′→3′) | Reverse Primer (5′→3′) |
|---|---|---|
| NT-31 | CGTGGTGGCGAACAGAACAT | GGCCGATGACTTGTCGGTC |
| BDNF2 | CTACGAGACCAAGTGCAATCC | AATCGCCAGCCAATTCTCTTT |
| GAPDH3 | GCAAGTTCAAGGCACA | CATTTGATGTTAGCGGGAT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.-H.; Huang, Y.-C.; Kuo, C.-Y.; Kuo, C.-Y.; Chin, C.-Y.; Yip, P.K.; Chen, J.-P. Docosahexaenoic Acid-Loaded Polylactic Acid Core-Shell Nanofiber Membranes for Regenerative Medicine after Spinal Cord Injury: In Vitro and In Vivo Study. Int. J. Mol. Sci. 2020, 21, 7031. https://doi.org/10.3390/ijms21197031
Liu Z-H, Huang Y-C, Kuo C-Y, Kuo C-Y, Chin C-Y, Yip PK, Chen J-P. Docosahexaenoic Acid-Loaded Polylactic Acid Core-Shell Nanofiber Membranes for Regenerative Medicine after Spinal Cord Injury: In Vitro and In Vivo Study. International Journal of Molecular Sciences. 2020; 21(19):7031. https://doi.org/10.3390/ijms21197031
Chicago/Turabian StyleLiu, Zhuo-Hao, Yin-Cheng Huang, Chang-Yi Kuo, Chao-Ying Kuo, Chieh-Yu Chin, Ping K. Yip, and Jyh-Ping Chen. 2020. "Docosahexaenoic Acid-Loaded Polylactic Acid Core-Shell Nanofiber Membranes for Regenerative Medicine after Spinal Cord Injury: In Vitro and In Vivo Study" International Journal of Molecular Sciences 21, no. 19: 7031. https://doi.org/10.3390/ijms21197031