Short-Term Mild Temperature-Stress-Induced Alterations in the C. elegans Phosphoproteome


State Key Laboratory of Genetic Engineering, Department of Biochemistry, School of Life Sciences, Fudan University, Shanghai 200438, China
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(17), 6409; https://doi.org/10.3390/ijms21176409
Submission received: 29 July 2020
/
Revised: 28 August 2020
/
Accepted: 31 August 2020
/
Published: 3 September 2020
(This article belongs to the Section Molecular Toxicology)
Abstract
:Exposure to mild early-life stresses can slow down aging, and protein phosphorylation might be an essential regulator in this process. However, the mechanisms of phosphorylation-based signaling networks during mild early-life stress remain elusive. Herein, we systematically analyzed the phosphoproteomes of Caenorhabditis elegans, which were treated with three mild temperatures (15 °C, 20 °C, and 25 °C) in two different short-term groups (10 min and 60 min). By utilizing an iTRAQ-based quantitative phosphoproteomic approach, 18,187 phosphosites from 3330 phosphoproteins were detected in this study. Volcano plots illustrated that the phosphorylation abundance of 374 proteins and 347 proteins, were significantly changed at 15 °C and 25 °C, respectively. Gene ontology, KEGG pathway and protein-protein interaction network analyses revealed that these phosphoproteins were primarily associated with metabolism, translation, development, and lifespan determination. A motif analysis of kinase substrates suggested that MAPK, CK, and CAMK were most likely involved in the adaption processes. Moreover, 16 and 14 aging-regulated proteins were found to undergo phosphorylation modifications under the mild stresses of 15 °C and 25 °C, respectively, indicating that these proteins might be important for maintaining long-term health. Further lifespan experiments confirmed that the candidate phosphoproteins, e.g., EGL-27 and XNP-1 modulated longevity at 15 °C, 20 °C, and 25 °C, and they showed increased tolerance to thermal and oxidative stresses. In conclusion, our findings offered data that supports understanding of the phosphorylation mechanisms involved in mild early-life stresses in C. elegans. Data are available via ProteomeXchange with identifier PXD021081.
1. Introduction
Environmental stresses, such as starvation and temperature, have an important effect on animal aging process [1,2,3,4]. Exposure to severe stresses often causes tissue damage and ultimately shortens the lifespan, while mild stresses usually promote defenses against damage and therefore promote longevity [5,6,7]. More importantly, studies in animals have shown that mild early-life stress, such as heat stress, could significantly extend the lifespan of animals [8,9], suggesting that stress in early life could be beneficial for health promotion throughout the animal’s lifespan. However, the exact biological mechanisms behind this phenomenon are largely unknown.
Temperature is a key regulator of the aging process in both heterothermic and homoiothermal animals [10,11], including Caenorhabditis elegans, a widely used model animal for aging studies [11,12]. The limiting factors for aging-related research are mainly the long natural lifespan, genetic heterogeneity, and few offspring of most model organisms [13]. C. elegans is an excellent aging model thanks to its short lifespan (3–4 weeks) and large numbers of offspring (200–300 animals) [14]. Notably, dozens of aging-related genes are also evolutionarily conserved from C. elegans to humans [11]. To date, many aging-related milestone discoveries have been first reported in C. elegans [15,16,17]. For example, insulin/IGF-1 signaling (e.g., daf-2, daf-16, akt-2, daf-18, and dao-5), germ line signaling (e.g., daf-9, daf-12, daf-36, and glp-1), and mTOR signaling (e.g., let-363 and daf-15) were uncovered via genetic perturbations in these animals [18]. Since then, plenty of genes have been identified to regulate longevity by influencing responses to a variety of stresses and nutrient alterations [19]. Various genes related to oxidative stress (clk-1) [20], mild heat stress (tax2/tax4) [21], hermetic heat stress (hsf-1) [22], and starvation (pha-4) [23] are associated with aging.
Nematodes have been cultured under standard laboratory conditions between 15 and 25 °C. The average lifespan of nematodes cultured at 25 °C (high temperature) is considerably shorter than that of nematodes cultured at 15 °C (low temperature) or 20 °C (normal temperature) [12]. However, severe short-term heat stress (36 °C) will not shorten the life of nematodes but will rather prolong life [24]. Notably, in the larval stage and early adult stage, suitable mild heat stress (25 °C), under laboratory conditions, can extend the lifespan of the nematode as well [8,9]. In addition, under mild heat stress (25 °C), heat-sensitive activated tax-2/tax-4 ion channels are distributed in ASJ neurons, and then insulin-like peptides are secreted to inhibit the activity of DAF-16/FOXO, resulting in a shortened lifespan [21]. Moreover, epigenetic regulators (cbp-1 and swsn-1) and a stress-activated protein kinase (pmk-1) enhance pathogen resistance, promoting longevity following early hermetic heat stress [9]. Overall, these results further indicate that temperature changes occurring in the early stage of life can significantly influence life at later stages, suggesting that proteins’ responses to early temperature changes might be potentially related to the regulation of the life cycle.
Protein phosphorylation, the most widespread type of post-translational modification, is essential for protein function [25,26] and may play an important role in the regulation of late-life activities of nematodes at high temperatures [27,28]. Moreover, phosphorylation is a key regulator that mediates signal conversion in response to external environmental signals for a short time, indicating it might also mediate mild temperature-stress responses in early life. Many studies have achieved the identification and analysis of only a limited number of phosphoproteins, due to the low mounts and technical limitations involved [29,30]. To date, there are no reports of a quantitative atlas of protein phosphorylation driven by different temperatures in early life in nematodes adapting to changing environments. Recently, owing to advancements in proteomics approaches and phosphopeptide enrichment approaches, it has become possible to identify and localize thousands of phosphorylation sites in a single experiment [31].
In this study, the iTRAQ (isobaric tags for relative and absolute quantification)-based proteomic approach [31] was introduced to systematically investigate the phosphoproteomic alterations of L4 nematodes by treating with 15 °C, 20 °C and 25 °C, respectively. A total of 18,187 phosphorylation sites from 3330 proteins were identified. The functional annotations were analyzed to better understand differentially regulated phosphoproteins (DRPs) and major events during early adaption processes. A Motif-X analysis of kinase substrates revealed that MAPK, CK2, and CAMK2 are most likely involved in the adaption processes. We carried out functional validation experiments on four candidates, EGL-27, XNP-1, PAR-1, and SAX-2. It was revealed that for egl-27- and xnp-1-deficient nematodes, the lifespan was significantly shorter and the stress tolerance was lower than control groups, whereas for par-1- and sax-2-deficient nematodes, the lifespan did not significantly change but the stress resistance was lower.
2. Results
2.1. Comparative Phosphoproteomic Analysis of C. elegans under Short-Term Mild Temperature Stress
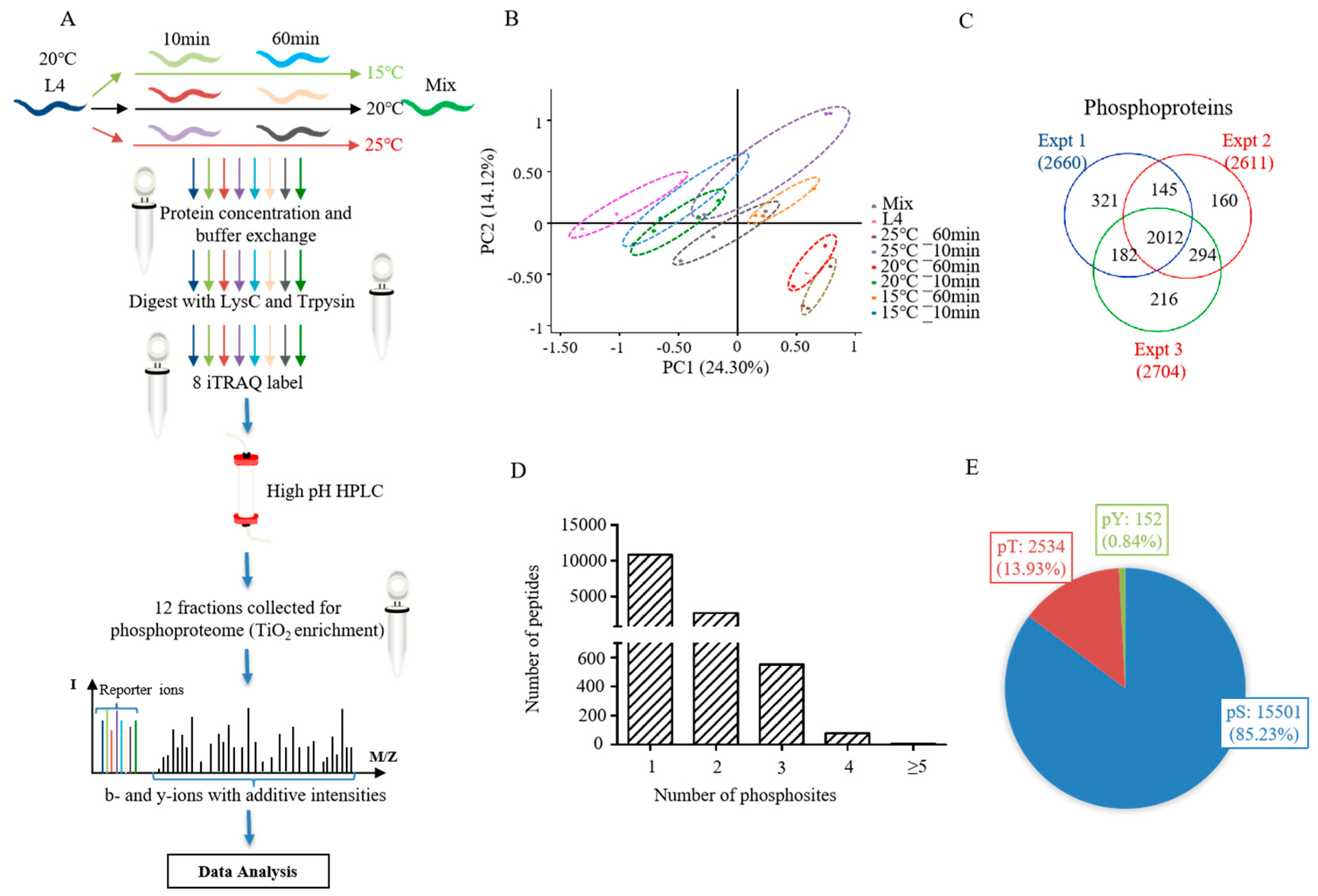
The phosphoproteomes of L4 nematodes subjected to different temperatures for different amounts of time were systematically studied by labeling with iTRAQ 8-plex reagents. This included L4 larvals, L4 larvals treated for 10 min or 60 min at 15 °C, 20 °C and 25 °C (six samples), and a mixture of all samples. The labeled peptides were fractionated by HPLC, combined, and then enriched by TiO2-affinity chromatography. A total of 12 fractions for each biological replicate were subjected to LC-MS/MS analysis (Figure 1A).
The phosphoproteomic data of all samples were analyzed to evaluate the reliability of experiments using principal component analysis (PCA) (Figure 1B and Supplementary Table S1), which showed similar tendencies of the three biological replicates, indicating high repeatability. From our phosphoproteomic analysis, there were 18,187 phosphosites identified from 3330 phosphoproteins (Supplementary Table S1). Of the three biological replicates, the phosphorylated proteins identified at least twice accounted for 78.4% of all proteins (Figure 1C). In total, 10,822 (76.6%), 2676 (18.9%), and 637 (4.5%) phosphopeptides were identified in one, two, and at least three phosphosites, respectively (Figure 1D). The percentage of phosphosites on phosphorylated serine (pSer), threonine (pThr), and tyrosine (pTyr) were 85.23% (11,501/18,187), 13.93% (2534/18,187), and 0.84% (152/18,187), respectively (Figure 1E).
2.2. Identification and Functional Classification of DRPs in C. elegans Responding to Different Temperature
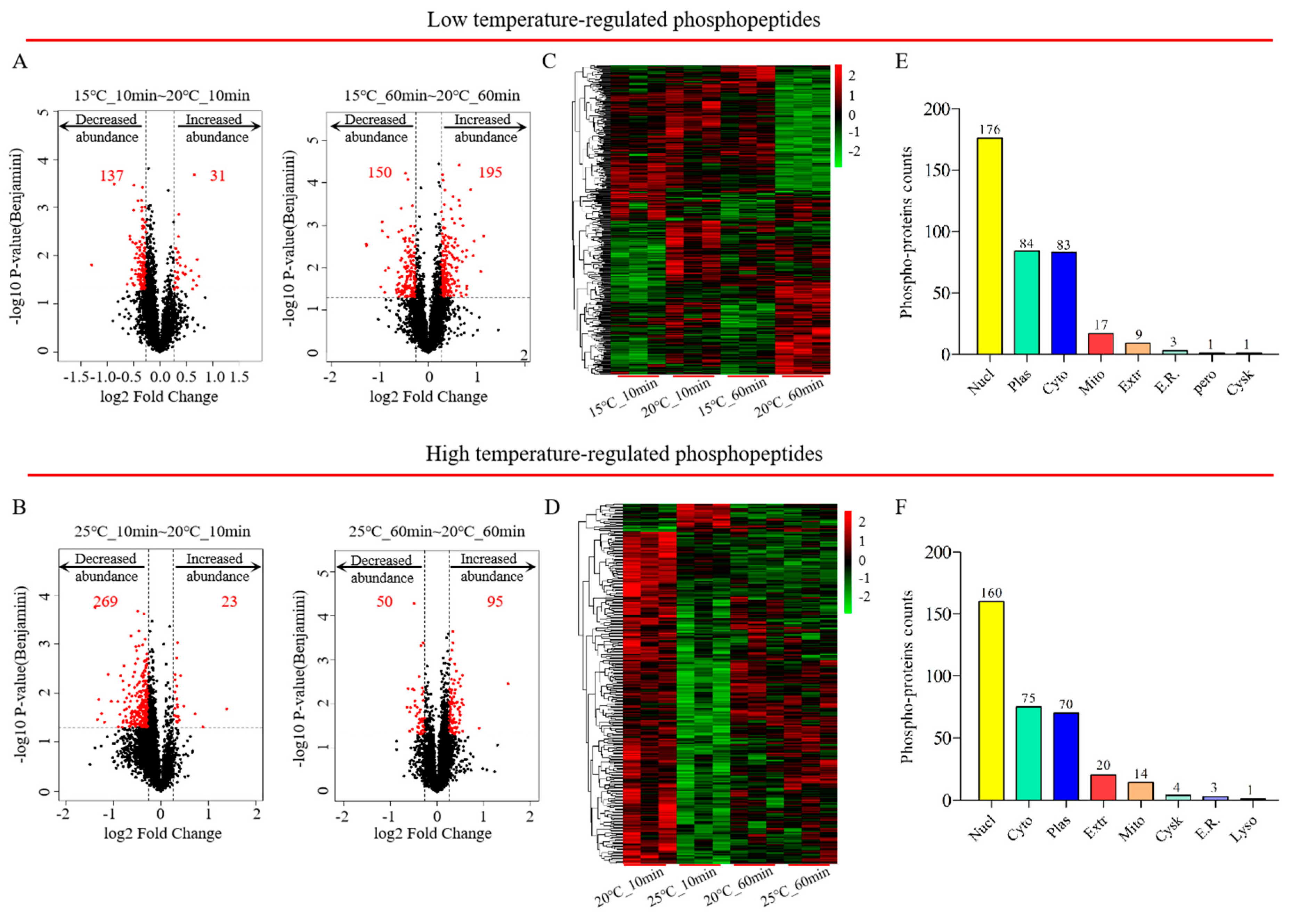
To further study the exact biological mechanisms of phosphoproteomes in nematodes exposed to various temperatures over a short period of time, differentially regulated phosphopeptides were graphically shown in volcano plots based on the criteria (fold change ratios ≥ 1.2 or ≤ 0.83 and p-value < 0.05). According to the results of the volcano plots (Figure 2A,B and Supplementary Table S2), a total of 500 differentially regulated phosphopeptides were acquired from 374 DRPs in C. elegans at 15 °C (217 up-regulated; 283 down-regulated), whilst 428 phosphopeptides were statistically significantly associated with 347 DRPs (115 up-regulated; 313 down-regulated). The significantly altered phosphorylation status of differentially regulated phosphopeptides induced by exposure to various temperatures for 10 min or 60 min was visualized using a heatmap (Figure 2C,D and Supplementary Table S2). The subcellular localization of differential phosphoproteins was intracellular for more than 90% of proteins, of which about 50% of phosphoproteins were located in the nucleus, indicating a rapid response to temperature changes and the regulation of genetic information involved in many biological processes (Figure 2E,F).
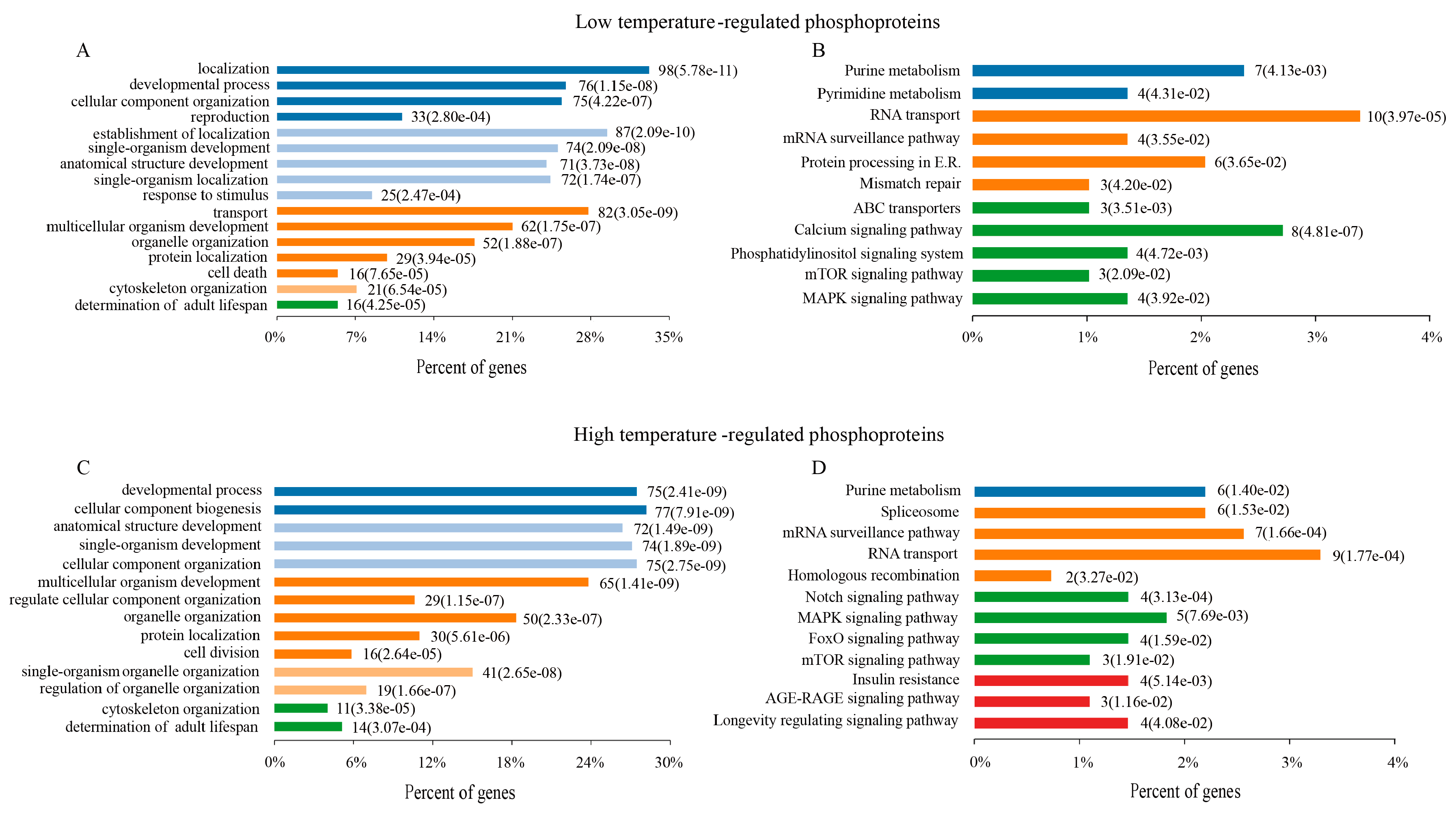
To better understand the physiological functions of DRPs, all of DRPs (10 min and 60 min) were assessed in terms of biological processes (BPs) at 15 °C and 25 °C using gene ontology analysis, respectively. Numerous BPs were found to be significantly altered in C. elegans at 15 °C, including development and reproduction, response to stimuli, determination of the adult lifespan, cell death, protein localization, and so on (Figure 3A and Supplementary Table S3). Similar to the BP results, the KEGG pathway enrichment results of these DRPs in C. elegans at 15 °C were related to metabolism, RNA transport, proteins proceeding in endoplasmic reticulum, and the longevity pathway (mTOR and MAPK signaling pathway) (Figure 3B and Supplementary Table S3).
In the organisms responding to 25 °C, these DRPs were principally related to many biological processes, including development, determination of adult lifespan, cell division, cellular component and organelle organization (Figure 3C and Supplementary Table S3). The KEGG pathway enrichment results of these DRPs in C. elegans mainly indicated the longevity pathway, spliceosome, RNA transport, Notch pathway, mTOR and MAPK signaling pathway (Figure 3D and Supplementary Table S3).
To further explore the functions of DRPs between 10 min and 60 min, a Venn analysis and KEGG pathway enrichment analysis of these DRPs were conducted at 15 °C and 25 °C (Supplementary Figure S1 and Supplementary Table S3). The comparison found that at 15 °C, the functions of 49 DRPs existed in 10 min and 60 min, related to calcium signaling pathway using KEGG pathway enrichment. In terms of different time points, 97 DRPs were found exclusively following 10 min at 15 °C, while 228 DRPs were found exclusively following treatment for 60 min. The KEGG pathway enrichment results revealed that 97 DRPs were related to RNA transport and degradation, calcium signaling pathway, and the longevity pathway (phosphatidylinositol, mTOR, and MAPK signaling pathway), and the 228 DRPs identified at 60 min were mainly involved in purine metabolism, RNA transport, mismatch repair, protein proceeding in endoplasmic reticulum, and the calcium signaling pathway.
Of the DRPs identified after 10 min and 60 min DRPs at 25 °C, 38 were present after both durations, and these related to taurine and hypotaurine metabolism, and non-homologous end joining. 216 DRPs identified at 10 min mainly participated in purine metabolism, RNA transport, protein proceeding in endoplasmic reticulum, and the longevity pathway (mTOR and MAPK signaling pathway). 93 DRPs identified at 60 min mainly related to spliceosome, RNA transport, notch signaling pathway, and AGE-RAGE signaling pathway.
Taken together, the BP and KEGG enrichment analysis results were similar for low- and high-temperature samples, indicating that these physiological functions are driven by extensive phosphorylation alterations following temperature changes.
2.3. Protein Protein Interaction Networks of DRPs
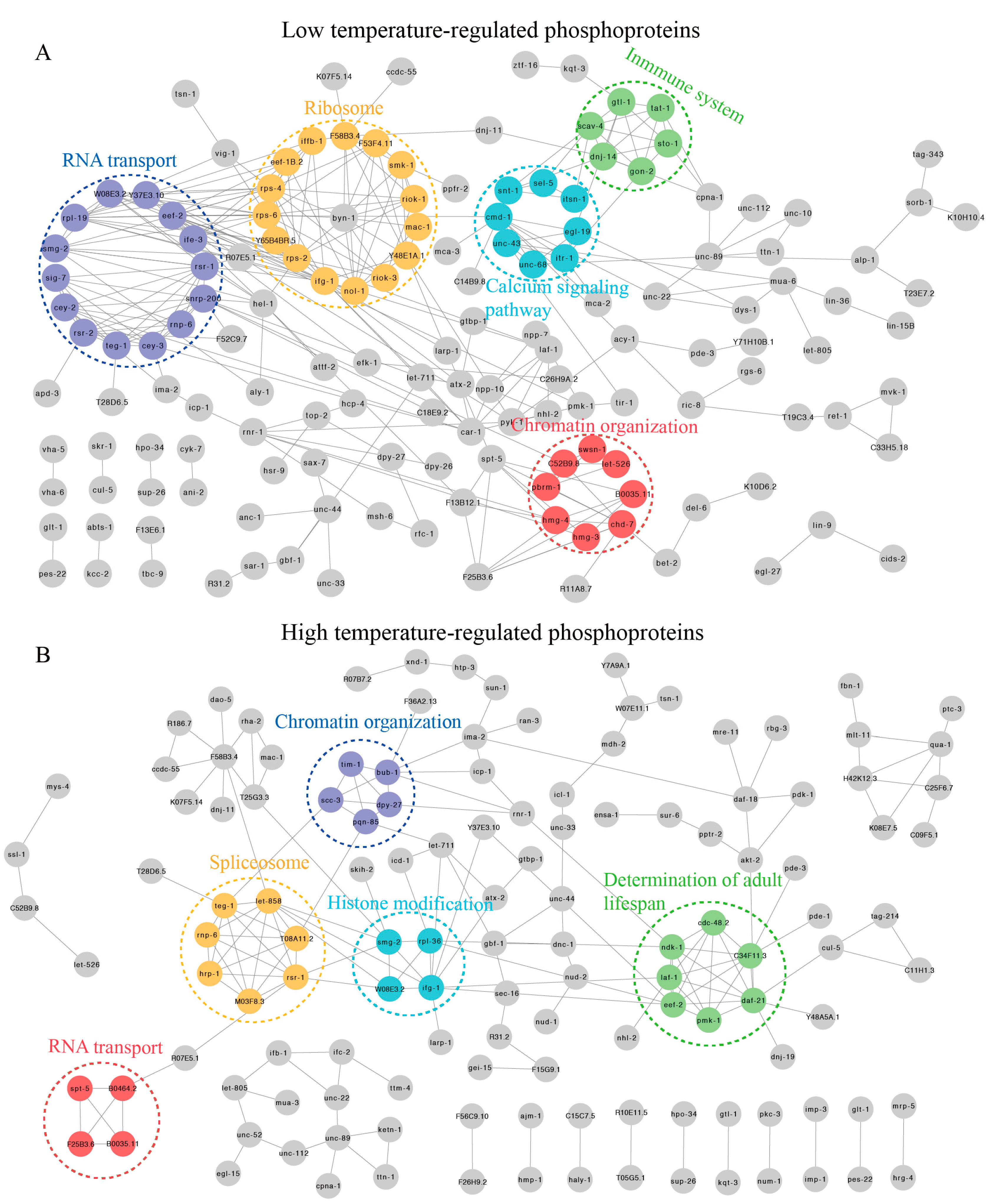
To provide insight into the possible cellular mechanisms, comprehensive protein–protein interactions (PPI) were used to depict numerous biological aspects of complex regulatory network and signaling pathways [32]. Hence, we used STRING11.0 to construct PPI networks for DRPs at 15 °C. As a result, we discovered that these DRPs might play important roles in translation, RNA transport, chromatin organization, immune system, and calcium signaling pathway (Figure 4A and Supplementary Table S4). In contrast, the highly clustered DRPs at 25 °C were related to spliceosome, the longevity pathway, chromatin organization, and RNA transport (Figure 4B and Supplementary Table S4). These results were similar to those found in the biological processes and KEGG enrichment analyses at 15 °C and 25 °C, further indicating that these processes and pathways may play important roles in early short-term temperature-stress-induced alterations.
2.4. Phosphorylation Motifs Analysis in C. elegans Following Short-Term Temperature Stress
Protein phosphorylation is regulated in vivo by protein kinases, which can be predicted using the sequences before and after the phosphosites [29]. To discover the kinases likely responding to low or high temperatures, the motif of protein phosphorylation was further derived from our phosphoproteomic data using the Motif-X algorithm.
As for the upregulated peptides in C. elegans responding to 15 °C, the conserved motif was occupied by a sequence of Arg-ser (the small letters represent the phosphorylated residue). Based on previous studies [33,34], we discovered that calcium/calmodulin kinase II (CaMK2) recognized the “...Rxxs......” sequence (Figure 5A). For the downregulated peptides, the enriched motifs were dominated by the sequences ser-Asp/Glu, Arg-ser, and ser-Pro (Figure 5B). The-e are two motifs recognized by casein kinase II (CK2): “......sxxD...” and “......sD.E...” sequences. These kinases—MAPK, mTOR or CDK—recognize the motifs of the “…...sP.....” sequence. Collectively, our results suggested that CAMK2 was specifically influenced by C. elegans in response to a temperature of 15 °C, while CK2 and MAPK, mTOR, or CDK were suppressed, indicating that these protein kinases might be essential regulators at low temperatures.
With regard to the upregulated peptides in C. elegans following exposure to a temperature of 25 °C, the enriched motif was occupied by the sequence of Arg-ser (Figure 5C). We found that the “...Rxxs......” sequence is recognized by CAMK2 kinases. As for the downregulated peptides, the conserved motifs were dominated by the sequence of ser-Glu, Arg-ser, and ser-Pro (Figure 5D). The motifs were recognized by CK2 kinase were “......sxE...”, “......sD.E...”. In addition, the motif recognized by CAMK2 kinase and the three kinases MAPK, mTOR, and CDK, including “...Rxxs......” and “…...sP.....”. These results demonstrate that CAMK2 was specifically influenced in the response of C. elegans to 25 °C temperatures, while CK2 and MAPK, mTOR and CDK were suppressed, suggesting that these kinases might be involved in the regulation under higher temperatures.
2.5. Interaction Analysis of the Kinase-Substrate in C. elegans Induced by Short-Term Temperature Stress
To further explore kinase substrate relationships, a comprehensive kinase substrate phosphorylation network (KSPN) was constructed for the differentially regulated phosphopeptides using iGPS 1.0 software [35].
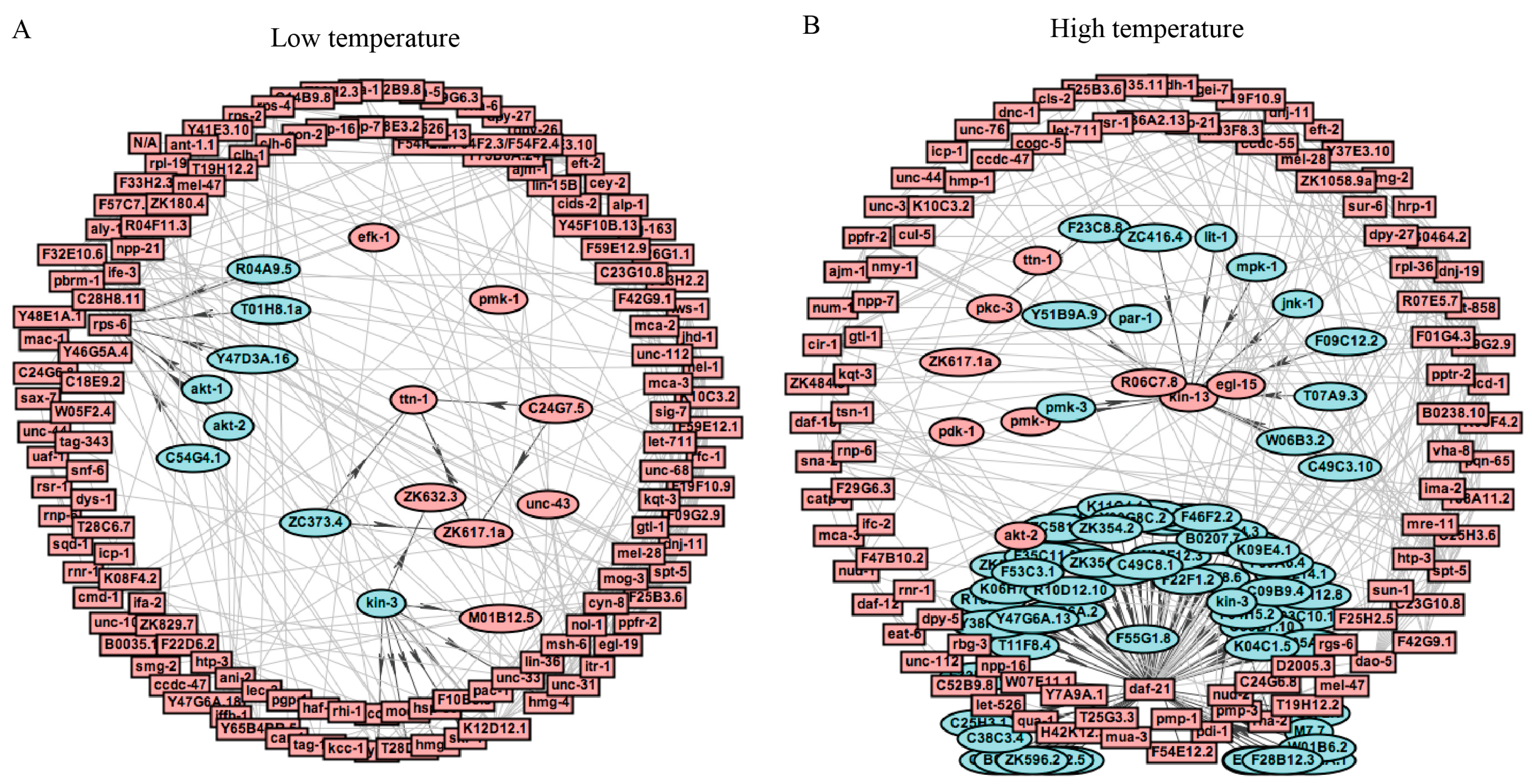
Of the kinases with enriched substrates, 11 kinases were found to have enriched substrate sites and possible high activity in C. elegans at 15 °C (Figure 6A and Supplementary Table S5). Similarly, to the results for phosphorylation motifs, the CAMK kinase family, which consists of the kinase TTN-1, ZK617.1a, ZC373.4, and C24G7.5; was identified. The CK2 kinase family member KIN-3 was predicted, and the Ser/Thr protein AGC kinase family, AKT-1, AKT-2, T01H8.1a, C54G4.1, Y47D3A.16, R04A9.5, were identified, indicating that these kinases might be key regulators of low temperature adaptation in C. elegans.
With regard to phosphopeptides significantly altered in C. elegans at 25 °C, we identified many kinase families, including CAMK, CK1, CK2, and MAPK. The CAMK kinases PAR-1 and F23C8.8, the CK2 kinase KIN-3, and the PLK kinase F55G1.8 were identified (Figure 6B and Supplementary Table S5). Interestingly, the substrates DAF-12 and KIN-13 were found to be regulated by 58 different CK1 kinases and 11 different MAPK kinases, respectively. In-depth studies of this possible kinase-substrate could help to elucidate the effects of short-term temperature-stress on C. elegans.
2.6. Temperature Responsive Phosphoproteins Associated with the Lifespan of C. elegans
To elucidate the important roles of the temperature responsive phosphoproteins in the regulation of long-term health and lifespan, we focused on the aging-related proteins. As shown in the results of the BP enrichment analysis of differentially phosphorylated proteins responding to low or high temperature in Table 1 and Table 2, lots of aging-related proteins undergo phosphorylation modifications. Interestingly, the aging-related phosphoproteins PMK-1, CNNM-1, NPP-7, and PDE-3 were found at 15 °C and 25 °C, indicating that these proteins might be vital in both stages of the lifespan. A previous study found the serine/threonine kinase PMK-1 [36], which is related to many environmental stresses, in intestine and nerve cells at moderate temperature, while it was shown to translocate into the nucleus at temperature above 32 °C, implicating that the function of PMK-1 is related to temperature changes. Double-knockout CNNM-1 with CNNM-3 resulted in reduced Mg2+ levels in other tissues, leading to a shortened lifespan [37].
As for other DRPs, multiple aging-related proteins have been reported to respond to temperature and regulate corresponding life activities. For example, the LMD-3 mutant shortens the lifespan of nematodes at low and high temperatures [38]. The TIR-1 mutant is temperature-sensitive for the AWCON phenotype, with the percentages of AWCON animals being 25%, 58%, and 81% at 15 °C, 20 °C and 25 °C, respectively [39]. NACT-1 modulates broad-spectrum stress resistance against factors such as multiple metals, oxidation, and heat [40]. Moreover, the longevity of worms is regulated by the DAF-12 steroid signaling pathway at high temperatures [41]. More importantly, AKT-2, DAF-18, and DAO-5 are key members in insulin signaling pathway regulating aging process in organisms [42].
These results indicate that early transient temperature changes could affect the longevity signal pathway, which would involve the regulation of physiological activities in response to temperature changes.
2.7. Temperature Responsive Phosphoproteins Could Regulate the Lifespan of C. elegans
The physiological functions of early-life DRPs were further explored in relation to the lifespan of C. elegans. It was reported previously that a few tissues are associated with aging pathways, e.g., the insulin-like growth factor (e.g., daf-2, daf-16, pha-4, and skn-1) pathway in neurons and the germ line signaling pathway [15,16,17,18]. Germ line elimination increases the lifespan of C. elegans, and many genes (e.g., glp-1, daf-9, and daf-36) involved in the reproductive system play important roles in longevity [18]. In addition, many conserved genes (e.g., trpa-1, crh-1, and tax-2/tax-4) expressed in the nervous system may sense temperature changes and regulate the aging process [8,21,43]. These results indicate that genes expressed in the reproductive and nervous systems might be essential regulators of the lifespan. Therefore, we focused on the candidate DRPs in the above tissues. Egg-laying defective protein 27 (EGL-27) was found to be expressed in the neurons and gonads and was associated with a shortened lifespan under normal temperatures [44]. Sensory axon guidance (SAX-2) was found to be expressed in head neurons and involved in neuron development [45]. Transcriptional regulator ATRX homolog (XNP-1) was found to be expressed in the germ line and it influences vulval development [46]. Serine/threonine-protein kinase (PAR-1) was found to be critical for vulval and neuronal morphogenesis [47,48].
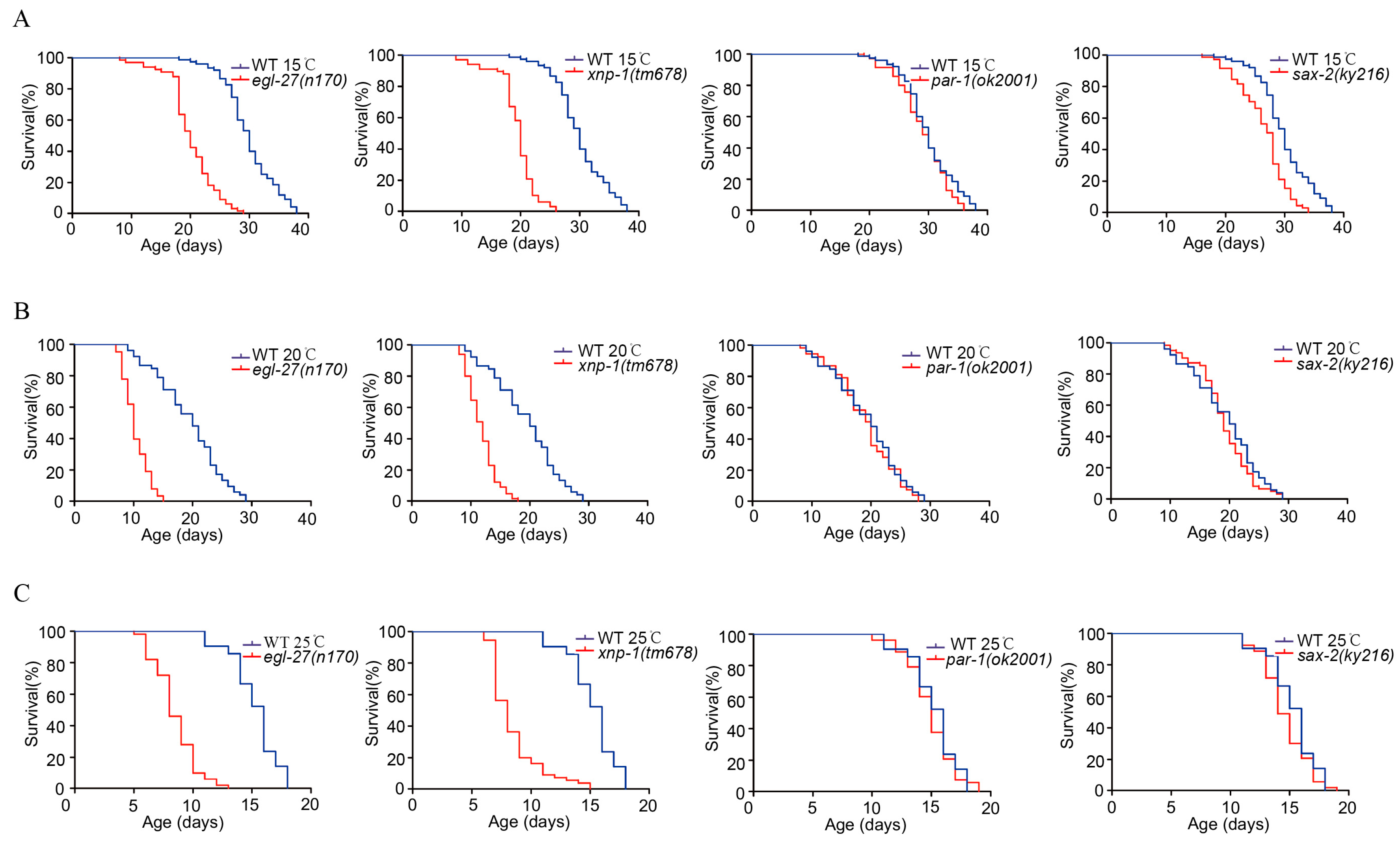
Based on these conditions, the candidate phosphoproteins (EGL-27, XNP-1, PAR-1, and SAX-2) were further chosen to confirm their effects on the longevity of C. elegans. The lifespans of EGL-27, XNP-1, PAR-1, and SAX-2 mutants were detected at 15 °C, 20°C and 25 °C (Figure 7). As shown in Figure 7, EGL-27 and XNP-1 mutants significantly shorten the lifespan compared with the wild-type nematodes (p < 0.01), while PAR-1 and SAX-2 are not significant compared with wild-type (p > 0.05), indicating the critical functions of EGL-27 and XNP-1 in regulating the lifespans of these worms.
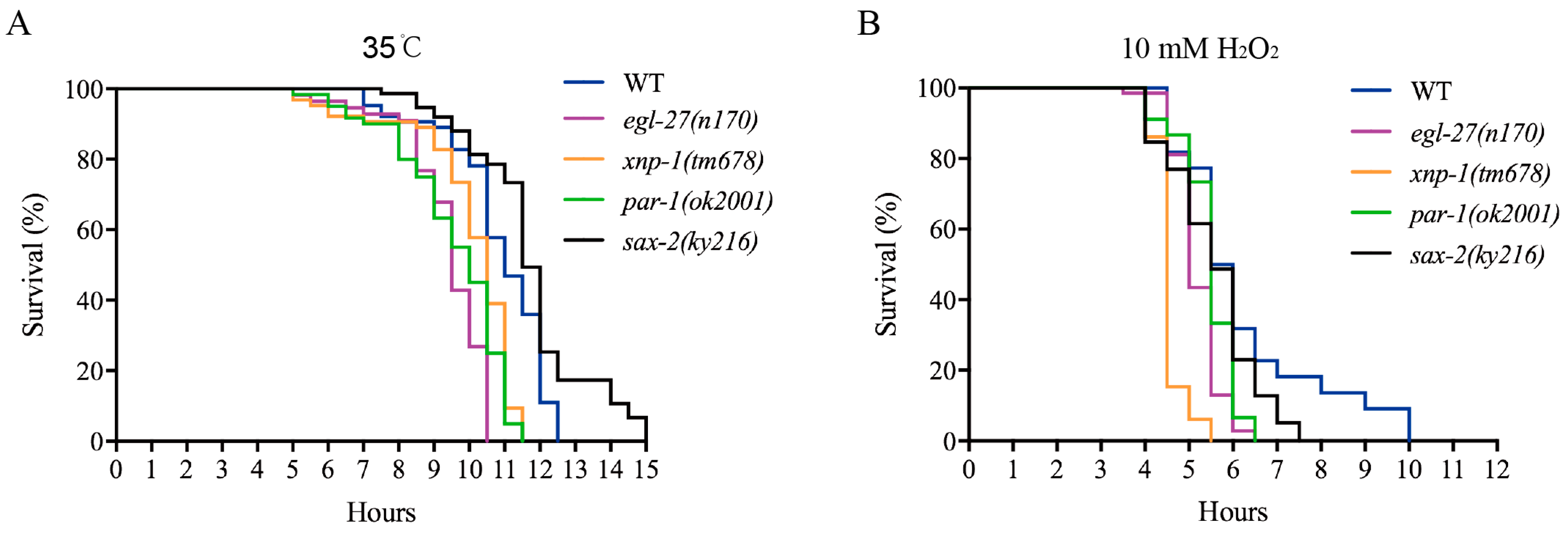
Lifespan extension has a certain correlation with resistance to different stresses, e.g.; thermal and oxidative stresses [49,50]. Survival curve analysis is typically used to evaluate the impacts of various stresses on organisms. Under thermal stress (35 °C) or hydrogen peroxide induced oxidative stress, the survival curves of EGL-27, XNP-1, and PAR-1 mutants were found to be shorter than those of wild-type strains (p < 0.01) but not that of SAX-2 mutants (Figure 8A,B). Taken together, EGL-27, XNP-1, and PAR-1 confer resistance to various stresses, improving survival rates.
3. Discussion
Many studies have studied the effects of mild stresses on extending the lifespan [8,9]. However, very little work has investigated how short-term mild stresses could have long-term beneficial effects. Herein, we conducted a systematic phosphoproteome analysis to identify and quantify the transient phosphoproteins of nematodes in response to different short-term temperature treatments. Functional experiments further confirmed that the responses of the early proteins EGL-27, XNP-1, and GTBP-1 to temperature, could regulate the lifespan of C. elegans at 15 °C, 20 °C and 25 °C.
As mentioned above, a total of 18,187 phosphorylation sites from 3330 proteins were recognized and the phosphosite distribution of pSer, pThr, and pTyr was consistent with that of other organisms [29], suggesting that protein phosphorylation is a highly conserved modification. A volcano plot revealed 374 DRPs in C. elegans at 15 °C, whilst there were 347 DRPs at 25 °C. More than 90% differentially expressed phosphoproteins were located in cells, of which about 50% were in the nucleus. Using BP and KEGG analysis, these phosphoproteins were found to be mainly involved in spliceosome and RNA transport. This suggests that the rapid response to external temperature changes involves dynamically changing phosphorylation modifications that regulate genetic information in many biological processes. In addition, other processes, including development, reproduction, and determination of the adult lifespan, were found to be enriched at 15 °C and 25 °C. Our results are compatible with the reported temperature regulation of physiological activities in organisms [41,51,52].
Notably, differentially regulated proteins responding to low temperature or high temperatures in the early nematode life stages are involved in the regulation of several conserved longevity pathways. PMK-1 is one kind of MAPK that affects the cascade reaction of MAPKs and could regulate meiosis and mitosis in cells during the aging process [53]. Moreover, the essential regulators of the insulin-signaling pathway, AKT-2, DAF-18, and DAO-5, can regulate the aging process [11]. In addition, DAF-12, an important component of reproductive system signaling, may regulate the process of high-temperature aging [41]. RPL-19 and RPS-6, which are related to the ribosome protein synthesis machine, can regulate proteomic synthesis, thus affecting the aging process [54]. Furthermore, it has also been reported that proteins that respond to temperature changes, such as LMD-3, can regulate the lifespan of C. elegans [38]. These results offer phosphoproteome-wide datasets that can deepen our understanding of early thermal processes.
The motif-substrate and kinase-substrate phosphorylation network results showed that CK, CAMK, AGC, MAPK, mTOR, and CDK are involved in early-life of nematodes’ responses to short-term temperature change. The kinase CK2 can regulate various biological processes, including gene expression and DNA repair, and can retard DNA damage and ensure genome integrity [55], thus delaying aging [54]. In addition, the reduction of protein synthesis improves protein balance, and thus extends longevity [54]. Several lines of evidence indicated that the early thermal-stress kinase CK2 might be an important regulator of long-term health benefits in nematodes. Moreover, MAPK, mTOR, and CDK are associated with multiple biological processes, e.g., immunity and stem-cell regeneration, accompanying the aging process [56,57]. Failure to defend against harmful factors may be due to a decline in immunity and could accelerate aging [54]. Furthermore, CAMK2 can impact cell growth and proliferation by regulating the calmodulin-induced reaction cascade [58]. It has been reported that the growth and proliferation of stem cells is significantly reduced during the aging process [59]. More importantly, these kinases might be essential regulators in the later life [28]. In brief, these kinases may play essential roles in the regulation of phosphorylation events throughout the lifespan.
The phosphoprotein EGL-27 (egg-laying defective protein 27) is a transcription factor that promoted stress survival and delayed aging at 20 °C [44]. We found that EGL-27 also modulated the lifespan of nematodes at both 15 °C and 25 °C. In addition, the phosphoprotein XNP-1 (transcriptional regulator ATRX homolog) mutant is temperature-sensitive, with 38% embryonic lethality at 25 °C [46]. The XNP-1 mutation significantly shortened the lifespans in nematodes at 15 °C, 20 °C and 25 °C and was associated with resistance to multiple stresses. These results add our understanding of the early temperature-sensitive phosphoproteins’ functions in later life.
In conclusion, our phosphoproteomic study systematically explored the effects of phosphorylation evens in response to early-life temperature changes. The functions of EGL-27 and XNP-1 modulated the lifespan upon exposure to different temperature. Nevertheless, further studies are needed to explore the mechanisms associated with these phosphoproteins. Our results offer phosphoproteome-wide data that will aid in the elucidation of short-term stress effects on long-term health benefits.
4. Materials and Methods
4.1. Worm Culture
The worms were cultured in accordance with the method presented in a previous study [60]. wild-type N2, MT170 egl-27 (n170) II., IG256 xnp-1 (tm678) I., RB1625 par-1 (ok2001) V., and CX3385 sax-2 (ky216) III. strains were purchased from the Caenorhabditis Genetics Center (CGC) (University of Minnesota, Minneapolis, MN, USA). Strains were placed onto agar plates with E. coli OP50 as food.
4.2. Phosphoproteomic Sample Preparation
To achieve age-synchronized L1 larval cells, eggs were originally obtained from gravid worms in accordance with a previous study [61]. The L1 larval cells were cultivated on NGM plates at 20 °C. The L4 nematodes were derived from age-synchronized L1 larval cells. In the late L4 stage, nematodes were separated into three groups and treated at 15 °C, 20 °C and 25 °C, respectively. The late-stage L4 nematodes growing at 20 °C were treated at 15 °C, 20 °C and 25 °C for 10 min and 60 min, and then immediately frozen to liquid nitrogen, followed by storage at −80 °C.
4.3. Protein Sample Preparation, Enzymatic Digestion, and iTRAQ Labeling
Briefly, the samples were resuspended in lysis buffer (4 M Gnd·HCl, 100 mM TEAB, 1% protease and phosphatase inhibitor cocktail). The samples were sonicated for 6 min on ice in 5 s bursts with pauses of 2 s. The samples were centrifuged for 20 min at 20,000× g and 4 °C. The supernatant was collected, and the protein concentration was measured with the Bradford assay. The samples were reduced, alkylated, and digested by Lys-C (MS grade, Wako Chemicals, Osaka, Japan) and trypsin (MS grade, Promega, Madison, WI, USA), as previously described [62]. Eight-plex iTRAQ reagents (AB Sciex, Foster City, CA, USA) were used to label digestive peptides based on the manufacturer’s instructions. The peptides (L4, 15 °C_10 min, 20 °C _10 min, 25 °C_10 min, 15 °C_60 min, 20 °C _60 min, 5 °C_60 min, and the mixture of the seven samples) were marked by 113, 114, 115, 116, 117, 118, 119, and 121-reagents, respectively.
4.4. Peptide Fractionation with HPLC and TiO2 Affinity Chromatography Enrichment
The labeled peptides were pooled with the same amount of each and then fractionated using a Dionex UltiMate-3000 HPLC system (Thermo Fisher Scientific, MA, USA). A high-pH reverse-phase separation was employed as the first dimensional separation. The peptides were passed through a C18 analytical column (2.6 µm, 100 mm × 3 mm (Phenomenex, Torrance, CA, USA)) through Solvent A (10 mM NH4Cl, pH 10.0) and Solvent B (10 mM NH4Cl, 80% ACN). Three hundred microgram samples of the generated peptides were loaded onto the column using Mobile Phase A, and the flow rate was set as follows: 0.1 mL/min for 10 min, 0.1–0.2 mL/min for 1 min and 0.2 mL/min for 34 min. The separation gradient was set using Mobile Phase B as follows: 0% for 11 min, 0–40% in 25 min, 40–100% in 2 min, 100% for 2 min, 100–0% in 2 min, and 0% for 2 min. Fractions were collected from the 6th min with 1 min intervals over the following 40 min. Forty fractions were collected and mixed into 12 fractions. The mixed fractions were lyophilized and enriched followed by TiO2-affinity chromatography in accordance with previous study [63].
4.5. Mass Spectrometry Analysis
The LC-MS/MS assay was conducted in a nanoflow EASY-nLC 1000 system (Thermo Fisher Scientific, Odense, Denmark). The peptides were separated using an Acclaim PepMap100 C18 Column (5 μm, 100 μm i.d. × 2 cm, 100 Å). The separated peptides were examined using an Acclaim PepMap RSLC C18 column (2 μm, 100 Å, 75 μm i.d. × 25 cm; Thermo Fisher Scientific, Sunnyvale, CA, USA). The LC was coupled with a LTQ Orbitrap Elite mass spectrometer (Thermo Fisher Scientific, Bremen, Germany).
The mobile phases contained Solvent A (0.1% formic acid) and Solvent B (0.1% formic acid in ACN). The separated peptides were set using Solvent B as follows: 2–5% B in 11 min, 5–28% in 98 min, 28–35% in 5 min, 35–39% in 2 min, 98% for 13 min at a flow rate of 200 nL/min. The data collection method was fixed to acquire a 1 MS scan with a resolution of 60,000 (m/z 400–1600, automatic gain control (AGC) target of 1e6, and a maximum ion injection time (IIT) of 100 ms), and 15 MS/MS scans of the strongest ion were detected by HCD (normalized collision energy (NCE) of 35; isolation width of 2 m/z; resolution of 15,000; AGC target of 5e4 and maximum IIT of 100 ms). Repeated peptides were dynamically excluded of 60 s. The precursor mass tolerance was set at 10 ppm.
4.6. Database Searching and Analysis
The uniprot worms database (20161228, 17,392 sequences) was searched for raw data using Proteome Discover (Version 1.4, Thermo Fisher Scientific, Bremen, Germany) followed by scoring with the Mascot server (Version 2.3, Matrix Science, London, UK). The searched parameters were set as follows: 10 ppm for the precursor ion mass deviation and 0.05 Da for the allowed fragment mass deviation; up to two miscleavage sites; cysteine carbamidomethylation, iTRAQ 8-plex on lysine and the N-terminal set as fixed modifications; methionine oxidation; and STY (serine, threonine and tyrosine) phosphorylation as variable modifications. The Target Decoy PSM verification program incorporated in the Proteome Discoverer was uitilized to verify the search results using only matches with FDR ≤ 0.01. Functional classifications of phosphoproteins were analyzed using the David database, as previously described [64]. Subcellular locations were predicted through Wolf PSORT [65]. Protein–protein interactions were detected by STRING [32]. The motifs of protein phosphorylation were derived from our phosphoproteomic data using the Motif-X algorithm [66]. The kinase-substrate phosphorylation network (KSPN) was constructed by submitting differentially expressed phosphopeptides by using iGPS 1.0 software (the CUCKOO workgroup, DICP, Dalian, China) [35].
4.7. Lifespan and Stress Assays
The lifespans of nematodes were monitored at 15 °C, 20 °C and 25 °C. Herein, one-day-old worms were considered to be at the first day of adulthood. Nematodes were recorded as dead in the absence of pharyngeal pumping and no response to gentle contact with a platinum wire. Worms that died of causes other than aging, such as sticking to the plate walls, bursting in the vulval region, or forming worm bags, were excluded from our analysis. With regard to each lifespan assay, 50–80 nematodes were cultured on NGM plates. All assays were carried out in triplicate with 50–80 worms in each replicate. To monitor the effects of temperature on longevity, L4-stage strains were cultured at 20 °C, and shifted to 15 °C, 20 °C and 25 °C.
For stress assays, One-day-old strains were placed onto agar plates at 35 °C and then the lifespans were recorded. One-day-old strains were transferred into S-basal buffer containing 10 mM of hydrogen peroxide and then the lifespans were recorded. All assays were carried out in triplicate using 40–80 worms in each replicate. The log-rank (Kaplan–Meier) test was used to calculate p-values. The survival curves were drafted using GraphPad Prism 8 (GraphPad Software Inc., San Diego, CA, USA).
Supplementary Materials
The Supplementary Materials are available online at https://www.mdpi.com/1422-0067/21/17/6409/s1.
Author Contributions
Conceptualization, J.H. and X.Z.; experiments, J.H.; writing—original draft preparation, J.H.; writing—review and editing, J.H.; Z.W., and X.Z.; project administration, X.Z.; funding acquisition, X.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (No. 31870822), Fudan University.
Acknowledgments
We thank Lin Huang (Proteomics Platform at the State Key Laboratory of Genetic Engineering) for the kind help with MS analysis. We thank Zhiyong Shao for his kind help and suggestions on experiments.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| BP | biologic process |
| CaMK II | calcium/calmodulin kinase II |
| CDK | cyclin-dependent kinase |
| CK | casein kinase |
| DRP | differentially regulated phosphoprotein |
| FDR | false discovery rate |
| HPLC | high performance liquid chromatography |
| iTRAQ | isobaric tag for relative and absolute quantitation |
| KEGG | Kyoto encyclopedia of genes and genomes |
| LC-MS/MS | liquid chromatography-tandem mass spectrometry |
| MAPK | mitogen-activated protein kinase |
| mTOR | mammalian target of rapamycin |
| PCA | principal component analysis |
| PPI | protein-protein interactions |
References
- Klosin, A.; Casas, E.; Hidalgo-Carcedo, C.; Vavouri, T.; Lehner, B. Transgenerational transmission of environmental information in C. elegans. Science 2017, 356, 320–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rechavi, O.; Houri-Ze’evi, L.; Anava, S.; Goh, W.S.S.; Kerk, S.Y.; Hannon, G.J.; Hobert, O. Starvation-induced transgenerational inheritance of small RNAs in C. elegans. Cell 2014, 158, 277–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schott, D.; Yanai, I.; Hunter, C.P. Natural RNA interference directs a heritable response to the environment. Sci. Rep. 2014, 4, 7387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, J.Z.; Kalinava, N.; Chen, E.; Huang, A.; Trinh, T.; Gu, S.G. A transgenerational role of the germline nuclear RNAi pathway in repressing heat stress-induced transcriptional activation in C. elegans. Epigenet. Chromatin 2016, 9, 3. [Google Scholar] [CrossRef] [Green Version]
- Cypser, J.R.; Johnson, T.E. Multiple stressors in Caenorhabditis elegans induce stress hormesis and extended longevity. J. Gerontol. A Biol. Sci. Med. Sci. 2002, 57, B109–B114. [Google Scholar] [CrossRef] [Green Version]
- Lithgow, G.J.; White, T.M.; Melov, S.; Johnson, T.E. Thermotolerance and extended life-span conferred by single-gene mutations and induced by thermal stress. Proc. Natl. Acad. Sci. USA 1995, 92, 7540–7544. [Google Scholar] [CrossRef] [Green Version]
- Dillin, A.; Hsu, A.L.; Arantes-Oliveira, N.; Lehrer-Graiwer, J.; Hsin, H.; Fraser, A.G.; Kamath, R.S.; Ahringer, J.; Kenyon, C. Rates of behavior and aging specified by mitochondrial function during development. Science 2002, 298, 2398–2401. [Google Scholar] [CrossRef]
- Zhang, B.; Xiao, R.; Ronan, E.A.; He, Y.; Hsu, A.L.; Liu, J.; Xu, X.Z. Environmental Temperature Differentially Modulates C. elegans Longevity through a Thermosensitive TRP Channel. Cell Rep. 2015, 11, 1414–1424. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; He, B.; Deng, J.; Pang, S.; Tang, H. Histone acetylation promotes long-lasting defense responses and longevity following early life heat stress. PLoS Genet. 2019, 15, e1008122. [Google Scholar] [CrossRef] [Green Version]
- Fontana, L.; Partridge, L.; Longo, V.D. Extending healthy life span--from yeast to humans. Science 2010, 328, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Kenyon, C.J. The genetics of ageing. Nature 2010, 464, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Zhang, B.; Dong, Y.; Gong, J.; Xu, T.; Liu, J.; Xu, X.Z. A genetic program promotes C. elegans longevity at cold temperatures via a thermosensitive TRP channel. Cell 2013, 152, 806–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, S.J.; Scheibye-Knudsen, M.; Longo, D.L.; De Cabo, R. Animal models of aging research: Implications for human aging and age-related diseases. Annu. Rev. Anim. Biosci. 2015, 3, 283–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narayan, V.; Ly, T.; Pourkarimi, E.; Murillo, A.B.; Gartner, A.; Lamond, A.I.; Kenyon, C. Deep proteome analysis identifies age-related processes in C. elegans. Cell Syst. 2016, 3, 144–159. [Google Scholar] [CrossRef] [Green Version]
- Kenyon, C.; Chang, J.; Gensch, E.; Rudner, A.; Tabtiang, R. A C. elegans mutant that lives twice as long as wild type. Nature 1993, 366, 461–464. [Google Scholar] [CrossRef]
- Kimura, K.D.; Tissenbaum, H.A.; Liu, Y.; Ruvkun, G. daf-2, an insulin receptor-like gene that regulates longevity and diapause in Caenorhabditis elegans. Science 1997, 277, 942–946. [Google Scholar] [CrossRef] [PubMed]
- Vellai, T.; Takacs-Vellai, K.; Zhang, Y.; Kovacs, A.L.; Orosz, L.; Müller, F. Genetics: Influence of TOR kinase on lifespan in C. elegans. Nature 2003, 426, 620. [Google Scholar] [CrossRef] [PubMed]
- Uno, M.; Nishida, E. Lifespan-regulating genes in C. elegans. NPJ Aging Mech. Dis. 2016, 2, 16010. [Google Scholar] [CrossRef]
- Kishimoto, S.; Uno, M.; Nishida, E. Molecular mechanisms regulating lifespan and environmental stress responses. Inflamm. Regen. 2018, 38, 22. [Google Scholar] [CrossRef] [Green Version]
- Murakami, S.; Johnson, T.E. A genetic pathway conferring life extension and resistance to UV stress in Caenorhabditis elegans. Genetics 1996, 143, 1207–1218. [Google Scholar]
- Lin, C.J.; Neve, I.A.A.; Wang, M.C. Neuronal regulation of longevity by staying cool. Genes Dev. 2018, 32, 197–198. [Google Scholar] [CrossRef]
- Hsu, A.L.; Murphy, C.T.; Kenyon, C. Regulation of aging and age-related disease by DAF-16 and heat-shock factor. Science 2003, 300, 1142–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panowski, S.H.; Wolff, S.; Aguilaniu, H.; Durieux, J.; Dillin, A. PHA-4/Foxa mediates diet-restriction-induced longevity of C. elegans. Nature 2007, 447, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Kumsta, C.; Chang, J.T.; Schmalz, J.; Hansen, M. Hormetic heat stress and HSF-1 induce autophagy to improve survival and proteostasis in C. elegans. Nat. Commun. 2017, 8, 14337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shumyantseva, V.V.; Suprun, E.V.; Bulko, T.V.; Archakov, A.I. Electrochemical methods for detection of post-translational modifications of proteins. Biosens. Bioelectron. 2014, 61, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Seet, B.T.; Dikic, I.; Zhou, M.M.; Pawson, T. Reading protein modifications with interaction domains. Nat. Rev. Mol. Cell Biol. 2006, 7, 473–483. [Google Scholar] [CrossRef]
- Rapacciuolo, A.; Suvarna, S.; Barki-Harrington, L.; Luttrell, L.M.; Cong, M.; Lefkowitz, R.J.; Rockman, H.A. Protein kinase A and G protein-coupled receptor kinase phosphorylation mediates beta-1 adrenergic receptor endocytosis through different pathways. J. Biol. Chem. 2003, 278, 35403–35411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Wu, Z.; Wang, J.; Zhang, X. Quantitative phosphoproteomics reveals GTBP-1 regulating C. elegans lifespan at different environmental temperatures. Biochem. Biophys. Res. Commun. 2018, 503, 1962–1967. [Google Scholar] [CrossRef]
- Johnson, S.A.; Hunter, T. Kinomics: Methods for deciphering the kinome. Nat. Methods 2005, 2, 17–25. [Google Scholar] [CrossRef]
- Park, S.J.; Ahmad, F.; Philp, A.; Baar, K.; Williams, T.; Luo, H.; Ke, H.; Rehmann, H.; Taussig, R.; Brown, A.L.; et al. Resveratrol ameliorates aging-related metabolic phenotypes by inhibiting cAMP phosphodiesterases. Cell 2012, 148, 421–433. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, N.; Masuda, T.; Shinoda, K.; Nakamura, A.; Tomita, M.; Ishihama, Y. Phosphopeptide enrichment by aliphatic hydroxy acid-modified metal oxide chromatography for nano-LC-MS/MS in proteomics applications. Mol. Cell Proteom. 2007, 6, 1103–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Bian, Y.; Cheng, K.; Gu, L.F.; Ye, M.; Zou, H.; Sun, S.S.; He, J.X. A large-scale protein phosphorylation analysis reveals novel phosphorylation motifs and phosphoregulatory networks in Arabidopsis. J. Proteom. 2013, 78, 486–498. [Google Scholar] [CrossRef] [PubMed]
- Nakayasu, E.S.; Tempel, R.; Cambronne, X.A.; Petyuk, V.A.; Jones, M.B.; Gritsenko, M.A.; Monroe, M.E.; Yang, F.; Smith, R.D.; Adkins, J.N.; et al. Comparative phosphoproteomics reveals components of host cell invasion and post-transcriptional regulation during Francisella infection. Mol. Cell Proteom. 2013, 12, 3297–3309. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Ye, M.; Liu, Z.; Cheng, H.; Jiang, X.; Han, G.; Songyang, Z.; Tan, Y.; Wang, H.; Ren, J.; et al. Systematic analysis of protein phosphorylation networks from phosphoproteomic data. Mol. Cell Proteom. 2012, 11, 1070–1083. [Google Scholar] [CrossRef] [Green Version]
- Mertenskotter, A.; Keshet, A.; Gerke, P.; Paul, R.J. 6shows heat-induced nuclear translocation, supports chaperone expression, and affects the heat tolerance of Caenorhabditis elegans. Cell Stress Chaperones 2013, 18, 293–306. [Google Scholar] [CrossRef] [Green Version]
- Ishii, T.; Funato, Y.; Hashizume, O.; Yamazaki, D.; Hirata, Y.; Nishiwaki, K.; Kono, N.; Arai, H.; Miki, H. Mg2+ Extrusion from Intestinal Epithelia by CNNM Proteins Is Essential for Gonadogenesis via AMPK-TORC1 Signaling in Caenorhabditis elegans. PLoS Genet. 2016, 12, e1006276. [Google Scholar] [CrossRef] [Green Version]
- Sanada, Y.; Asai, S.; Ikemoto, A.; Moriwaki, T.; Nakamura, N.; Miyaji, M.; Zhang-Akiyama, Q.M. Oxidation resistance 1 is essential for protection against oxidative stress and participates in the regulation of aging in Caenorhabditis elegans. Free Radic. Res. 2014, 48, 919–928. [Google Scholar] [CrossRef]
- Chuang, C.F.; Bargmann, C.I. A Toll-interleukin 1 repeat protein at the synapse specifies asymmetric odorant receptor expression via ASK1 MAPKKK signaling. Genes Dev. 2005, 19, 270–281. [Google Scholar] [CrossRef] [Green Version]
- Warnhoff, K.; Murphy, J.T.; Kumar, S.; Schneider, D.L.; Peterson, M.; Hsu, S.; Guthrie, J.; Robertson, J.D.; Kornfeld, K. The DAF-16 FOXO transcription factor regulates natc-1 to modulate stress resistance in Caenorhabditis elegans, linking insulin/IGF-1 signaling to protein N-terminal acetylation. PLoS Genet. 2014, 10, e1004703. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.J.; Kenyon, C. Regulation of the longevity response to temperature by thermosensory neurons in Caenorhabditis elegans. Curr. Biol. 2009, 19, 715–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbieri, M.; Bonafe, M.; Franceschi, C.; Paolisso, G. Insulin/IGF-I-signaling pathway: An evolutionarily conserved mechanism of longevity from yeast to humans. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E1064–E1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.C.; Chen, H.J.; Tseng, W.C.; Hsu, J.M.; Huang, T.T.; Chen, C.H.; Pan, C.L. A C. elegans thermosensory circuit regulates longevity through crh-1/CREB-dependent flp-6 neuropeptide signaling. Dev. Cell 2016, 39, 209–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Kim, S.K. The GATA transcription factor egl-27 delays aging by promoting stress resistance in Caenorhabditis elegans. PLoS Genet. 2012, 8, e1003108. [Google Scholar] [CrossRef] [Green Version]
- Gallegos, M.E.; Bargmann, C.I. Mechanosensory neurite termination and tiling depend on SAX-2 and the SAX-1 kinase. Neuron 2004, 44, 239–249. [Google Scholar] [CrossRef] [Green Version]
- Bender, A.M.; Wells, O.; Fay, D.S. lin-35/Rb and xnp-1/ATR-X function redundantly to control somatic gonad development in C. elegans. Dev. Biol. 2004, 273, 335–349. [Google Scholar] [CrossRef] [Green Version]
- Hurd, D.D.; Kemphues, K.J. PAR-1 is required for morphogenesis of the Caenorhabditis elegans vulva. Dev. Biol. 2003, 253, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.S.; Hung, W.; Narbonne, P.; Roy, R.; Zhen, M. C. elegans STRADalpha and SAD cooperatively regulate neuronal polarity and synaptic organization. Development 2010, 137, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Hirota, K.; Shigekawa, C.; Araoi, S.; Sha, L.; Inagawa, T.; Kanou, A.; Kako, K.; Daitoku, H.; Fukamizu, A. Simultaneous ablation of prmt-1 and prmt-5 abolishes asymmetric and symmetric arginine dimethylations in Caenorhabditis elegans. J. Biochem. 2017, 161, 521–527. [Google Scholar] [CrossRef]
- Vilchez, D.; Morantte, I.; Liu, Z.; Douglas, P.M.; Merkwirth, C.; Rodrigues, A.P.; Manning, G.; Dillin, A. RPN-6 determines C. elegans longevity under proteotoxic stress conditions. Nature 2012, 489, 263–268. [Google Scholar] [CrossRef]
- Schwartz, T.S.; Pearson, P.; Dawson, J.; Allison, D.B.; Gohlke, J.M. Effects of fluctuating temperature and food availability on reproduction and lifespan. Exp. Gerontol. 2016, 86, 62–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanford, J.I.; Blanford, S.; Crane, R.G.; Mann, M.E.; Paaijmans, K.P.; Schreiber, K.V.; Thomas, M.B. Implications of temperature variation for malaria parasite development across Africa. Sci. Rep. 2013, 3, 1300. [Google Scholar] [CrossRef] [PubMed]
- Kassahun, H.; SenGupta, T.; Schiavi, A.; Maglioni, S.; Skjeldam, H.K.; Arczewska, K.; Brockway, N.L.; Estes, S.; Eide, L.; Ventura, N.; et al. Constitutive MAP-kinase activation suppresses germline apoptosis in NTH-1 DNA glycosylase deficient C. elegans. DNA Repair 2018, 61, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [Green Version]
- Barz, T.; Ackermann, K.; Dubois, G.; Eils, R.; Pyerin, W. Genome-wide expression screens indicate a global role for protein kinase CK2 in chromatin remodeling. J. Cell Sci. 2003, 116, 1563–1577. [Google Scholar] [CrossRef] [Green Version]
- Bonni, A.; Brunet, A.; West, A.E.; Datta, S.R.; Takasu, M.A.; Greenberg, M.E. Cell survival promoted by the Ras-MAPK signaling pathway by transcription-dependent and -independent mechanisms. Science 1999, 286, 1358–1362. [Google Scholar] [CrossRef] [Green Version]
- Latrasse, D.; Jegu, T.; Li, H.; de Zelicourt, A.; Raynaud, C.; Legras, S.; Gust, A.; Samajova, O.; Veluchamy, A.; Rayapuram, N.; et al. MAPK-triggered chromatin reprogramming by histone deacetylase in plant innate immunity. Genome Biol. 2017, 18, 131. [Google Scholar] [CrossRef]
- House, S.J.; Ginnan, R.G.; Armstrong, S.E.; Singer, H.A. Calcium/calmodulin-dependent protein kinase II-delta isoform regulation of vascular smooth muscle cell proliferation. Am. J. Physiol. Cell Physiol. 2007, 292, C2276–C2287. [Google Scholar] [CrossRef] [Green Version]
- Rao, M.S.; Mattson, M.P. Stem cells and aging: Expanding the possibilities. Mech. Ageing Dev. 2001, 122, 713–734. [Google Scholar] [CrossRef]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar]
- Fabian, T.J.; Johnson, T.E. Production of age-synchronous mass cultures of Caenorhabditis elegans. J. Gerontol. 1994, 49, B145–B156. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods. 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Thingholm, T.E.; Jorgensen, T.J.; Jensen, O.N.; Larsen, M.R. Highly selective enrichment of phosphorylated peptides using titanium dioxide. Nat. Protoc. 2006, 1, 1929–1935. [Google Scholar] [CrossRef] [PubMed]
- Dennis, G.J.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol. 2003, 4, P3. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, D.; Gygi, S.P. An iterative statistical approach to the identification of protein phosphorylation motifs from large-scale data sets. Nat. Biotechnol. 2005, 23, 1391–1398. [Google Scholar] [CrossRef]
Figure 1.
The quantitative phosphoproteome of Caenorhabditis elegans exposed to a mild temperature over a short-term period. (A) Scheme of the phosphoproteomic strategy. (B) Principal component analysis (PCA) of all quantified phosphoproteins. (C) The overlap of phosphoproteins in different triplicates of experiments. (D) The number distribution of phosphopeptide with different phosphosites. (E) The percentage of phosphorylated serine (pSer), threonine (pThr), and tyrosine (pTyr).
Figure 1.
The quantitative phosphoproteome of Caenorhabditis elegans exposed to a mild temperature over a short-term period. (A) Scheme of the phosphoproteomic strategy. (B) Principal component analysis (PCA) of all quantified phosphoproteins. (C) The overlap of phosphoproteins in different triplicates of experiments. (D) The number distribution of phosphopeptide with different phosphosites. (E) The percentage of phosphorylated serine (pSer), threonine (pThr), and tyrosine (pTyr).

Figure 2.
The determination of differentially regulated phosphoproteins (DRPs). Volcano plot analysis of DRPs in C. elegans exposed to 15 °C (A) or 25 °C (B). Heatmap of DRPs in C. elegans at 15 °C (C) or 25 °C (D). Subcellular localization of the phosphoproteins at 15 °C (E) or 25 °C (F).
Figure 2.
The determination of differentially regulated phosphoproteins (DRPs). Volcano plot analysis of DRPs in C. elegans exposed to 15 °C (A) or 25 °C (B). Heatmap of DRPs in C. elegans at 15 °C (C) or 25 °C (D). Subcellular localization of the phosphoproteins at 15 °C (E) or 25 °C (F).

Figure 3.
Functional classifications of DRPs. Biological process (BP) enrichment analysis of DRPs at 15 °C (A) and 25 °C (B). KEGG pathway enrichment analysis of DRPs at 15 °C (C) and 25 °C (D).
Figure 3.
Functional classifications of DRPs. Biological process (BP) enrichment analysis of DRPs at 15 °C (A) and 25 °C (B). KEGG pathway enrichment analysis of DRPs at 15 °C (C) and 25 °C (D).

Figure 4.
Protein–protein interaction (PPI) networks of DRPs. (A) PPI analysis of DRPs in C. elegans exposed to 15 °C. (B) PPI analysis of DRPs in C. elegans exposed to 25 °C.
Figure 4.
Protein–protein interaction (PPI) networks of DRPs. (A) PPI analysis of DRPs in C. elegans exposed to 15 °C. (B) PPI analysis of DRPs in C. elegans exposed to 25 °C.

Figure 5.
The features of enriched phosphorylation motifs. (A) Enriched phosphorylation motif of peptides upregulated at 15 °C. (B) Enriched phosphorylation motifs of peptide downregulated at 15 °C. (C) Enriched phosphorylation motif of peptides upregulated at 25 °C. (D) Enriched phosphorylation motifs of peptide downregulated at 25 °C.
Figure 5.
The features of enriched phosphorylation motifs. (A) Enriched phosphorylation motif of peptides upregulated at 15 °C. (B) Enriched phosphorylation motifs of peptide downregulated at 15 °C. (C) Enriched phosphorylation motif of peptides upregulated at 25 °C. (D) Enriched phosphorylation motifs of peptide downregulated at 25 °C.

Figure 6.
Interaction analysis of kinase substrate relationships of DRPs. (A) Kinase substrate interaction analysis of these proteins exposed to 15 °C. (B) Kinase substrate interaction analysis of these proteins exposed to 25 °C.
Figure 6.
Interaction analysis of kinase substrate relationships of DRPs. (A) Kinase substrate interaction analysis of these proteins exposed to 15 °C. (B) Kinase substrate interaction analysis of these proteins exposed to 25 °C.

Figure 7.
Temperature-responsive phosphoproteins are essential modulators of the worm lifespan. (A) The lifespans of egl-27 and xnp-1 mutants were significantly reduced at 15 °C (p < 0.01, log-rank test). The lifespans of sax2 and par-1 mutants were not reduced at 15 °C (p > 0.05, log-rank test). (B) The lifespans of egl-27 and xnp-1 mutants were significantly reduced at 20 °C (p < 0.01, log-rank test). The lifespans of sax2 and par-1 mutants were not reduced at 20 °C (p > 0.05, log-rank test). (C) The lifespans of egl-27 and xnp-1 mutants were significantly reduced at 25 °C (p < 0.01, log-rank test). The sax2 and par-1 mutants were not significantly reduced at 25 °C (p > 0.05, log-rank test).
Figure 7.
Temperature-responsive phosphoproteins are essential modulators of the worm lifespan. (A) The lifespans of egl-27 and xnp-1 mutants were significantly reduced at 15 °C (p < 0.01, log-rank test). The lifespans of sax2 and par-1 mutants were not reduced at 15 °C (p > 0.05, log-rank test). (B) The lifespans of egl-27 and xnp-1 mutants were significantly reduced at 20 °C (p < 0.01, log-rank test). The lifespans of sax2 and par-1 mutants were not reduced at 20 °C (p > 0.05, log-rank test). (C) The lifespans of egl-27 and xnp-1 mutants were significantly reduced at 25 °C (p < 0.01, log-rank test). The sax2 and par-1 mutants were not significantly reduced at 25 °C (p > 0.05, log-rank test).

Figure 8.
Temperature-responsive phosphoproteins are important regulators of the lifespan of C. elegans. (A) One-day-old egl-27, xnp-1, and par-1 mutants were less resistant to heat stress (p < 0.01, log-rank test). One-day-old sax-2 mutants showed thermal resistance (p < 0.01, log-rank test). (B) One-day-old egl-27, xnp-1, and par-1 mutants were shorter than wild-type strains under hydrogen peroxide treatment (p < 0.01, log-rank test). One-day-old sax-2 mutants were shorter than wild-type strains under hydrogen peroxide treatment (p < 0.05, log-rank test).
Figure 8.
Temperature-responsive phosphoproteins are important regulators of the lifespan of C. elegans. (A) One-day-old egl-27, xnp-1, and par-1 mutants were less resistant to heat stress (p < 0.01, log-rank test). One-day-old sax-2 mutants showed thermal resistance (p < 0.01, log-rank test). (B) One-day-old egl-27, xnp-1, and par-1 mutants were shorter than wild-type strains under hydrogen peroxide treatment (p < 0.01, log-rank test). One-day-old sax-2 mutants were shorter than wild-type strains under hydrogen peroxide treatment (p < 0.05, log-rank test).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The aging-related phosphoproteins in C. elegans at low temperatures.
| Accession | Gene Names | Sequence | Log2FC | Protein Names |
|---|---|---|---|---|
| O17003 | natc-1 | EYEGAEENSDEKSpEESpTSDPTPSSEAK | −0.614 | N-alpha-AcetylTransferase C complex subunit |
| O02639 | rpl-19 | VWLDPNEVSpEISGANSR | −0.456 | 60S ribosomal protein L19 |
| Q17446 | pmk-1 | QTDSEMTpGYpVATR | −0.416 | Stress-activated protein kinase pmk-1 |
| O61955 | ife-3 | GPAATTSPSNPGTEATGTSPATpPTpP | −0.369 | Eukaryotic translation initiation factor 4E-3 |
| P91457 | npp-7 | TASTASTTTINSpSNSR | −0.315 | Nuclear Pore complex Protein |
| Q18212 | hel-1 | LANDCPHIVVGTpPGR | −0.28 | Spliceosome RNA helicase DDX39B homolog |
| Q8I0P7 | pde-3 | KESpGTHVDTVVTTIDGQR | −0.267 | Probable 3’,5’-cyclic phosphodiesterase |
| O45865 | ant-1.1 | LLLQVQDASpK | 0.273 | Adenine Nucleotide Translocator |
| H2L003 | lmd-3 | SpIQGTVTSGAEK | 0.296 | LysM Domain (Peptidoglycan binding) protein |
| E5QCE7 | dnj-14 | EESpPAADHSHDPK | 0.31 | DNaJ domain (Prokaryotic heat shock protein) |
| Q9NEN6 | rps-6 | SSpASpHHSpESEVK | 0.327 | 40S ribosomal protein S6 |
| A3QM97 | cnnm-1 | SMSpIVGTSELSAR | 0.396 | Metal transporter cnnm-1 |
| Q21219 | pept-1 | GYSESRSpESpVSpSK | 0.463 | Peptide transporter family 1 (Di-/tri-peptide transporter CPTB) |
| G5EFL5 | alp-1 | AAYHPQVNTpARPVSpVSpPAPSpAGSK | 0.478 | ALP/Enigma encoding |
| H2KZZ2 | hlh-30 | QVVSSSpAPTSSIDIEK | 0.654 | Helix Loop Helix |
| Q86DA5 | tir-1 | FLIQQDSpVVNPSTK | 0.671 | Sterile alpha and TIR motif-containing protein |
Table 2.
The aging-related phosphoproteins in C. elegans at high temperatures.
| Accession | Gene Names | Sequence | Log2FC | Protein Names |
|---|---|---|---|---|
| G5EFF5 | daf-12 | LEESSGACGSpPDDGLLDSSEESR | −0.606 | Nuclear hormone receptor family member |
| Q18688 | daf-21 | IEGAEEDASpRMEEVD | −0.477 | Heat shock protein 90 |
| O45962 | rle-1 | DNSHNSpPQTpPPK | −0.394 | Regulation of longevity by E3 ubiquitin-protein ligase |
| Q9XVS4 | dao-5 | ADLSSDFSpDDEAPAK | −0.393 | Dauer or Aging adult Overexpression |
| G5EE01 | daf-18 | VTpPPPDVPSTSTR | −0.343 | Phosphatidylinositol 3,4,5-trisphosphate 3-phosphatas |
| Q8I0P7 | pde-3 | RDSpPLDSDLSQ | −0.333 | Probable 3’,5’-cyclic phosphodiesterase |
| Q9XTG7 | akt-2 | EFTSMPVQLTpPPR | −0.329 | Serine/threonine-protein kinase |
| P91457 | npp-7 | TASTASTTTINSpSNSR | −0.329 | Nuclear Pore complex Protein |
| Q8MQ70 | hpk-1 | NSpGQSTDLNSR | −0.322 | Homeodomain-interacting protein kinase 1 |
| O76360 | egl-4 | KPSpDQPNGNQVQVGTR | −0.271 | cGMP-dependent protein kinase |
| A3QM97 | cnnm-1 | SMSpIVGTSELSAR | 0.34 | Metal transporter |
| O18696 | pde-1 | SpYDNAPALESLEK | 0.403 | Probable 3’,5’-cyclic phosphodiesterase |
| Q10663 | icl-1 | AGSpVVNRIPEAADLLEK | 0.414 | Bifunctional glyoxylate cycle protein |
| Q17446 | pmk-1 | QTDSEMTpGYpVATR | 0.537 | Stress-activated protein kinase |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Huang, J.; Wu, Z.; Zhang, X. Short-Term Mild Temperature-Stress-Induced Alterations in the C. elegans Phosphoproteome. Int. J. Mol. Sci. 2020, 21, 6409. https://doi.org/10.3390/ijms21176409
AMA Style
Huang J, Wu Z, Zhang X. Short-Term Mild Temperature-Stress-Induced Alterations in the C. elegans Phosphoproteome. International Journal of Molecular Sciences. 2020; 21(17):6409. https://doi.org/10.3390/ijms21176409
Chicago/Turabian StyleHuang, Jichang, Zhen Wu, and Xumin Zhang. 2020. "Short-Term Mild Temperature-Stress-Induced Alterations in the C. elegans Phosphoproteome" International Journal of Molecular Sciences 21, no. 17: 6409. https://doi.org/10.3390/ijms21176409
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.