Photosystem Disorder Could be the Key Cause for the Formation of Albino Leaf Phenotype in Pecan


Abstract
:1. Introduction
2. Results
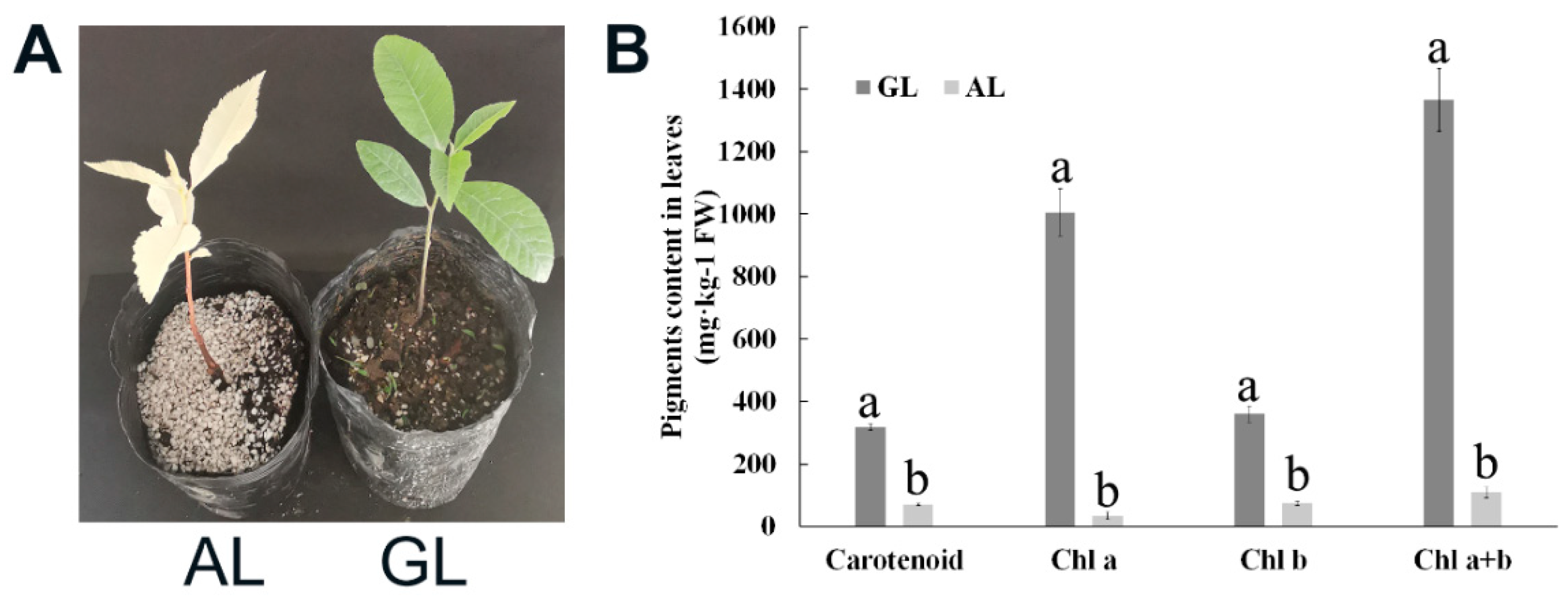
2.1. Content of Chlorophyll and Carotenoid in Green and Albino Leaves in Pecan
2.2. RNA Sequencing of Leaf Transcriptomes of the GL and AL Seedlings and Mapping of RNA Sequences to the Reference Genome
2.3. Differentially Expressed Gene Analysis
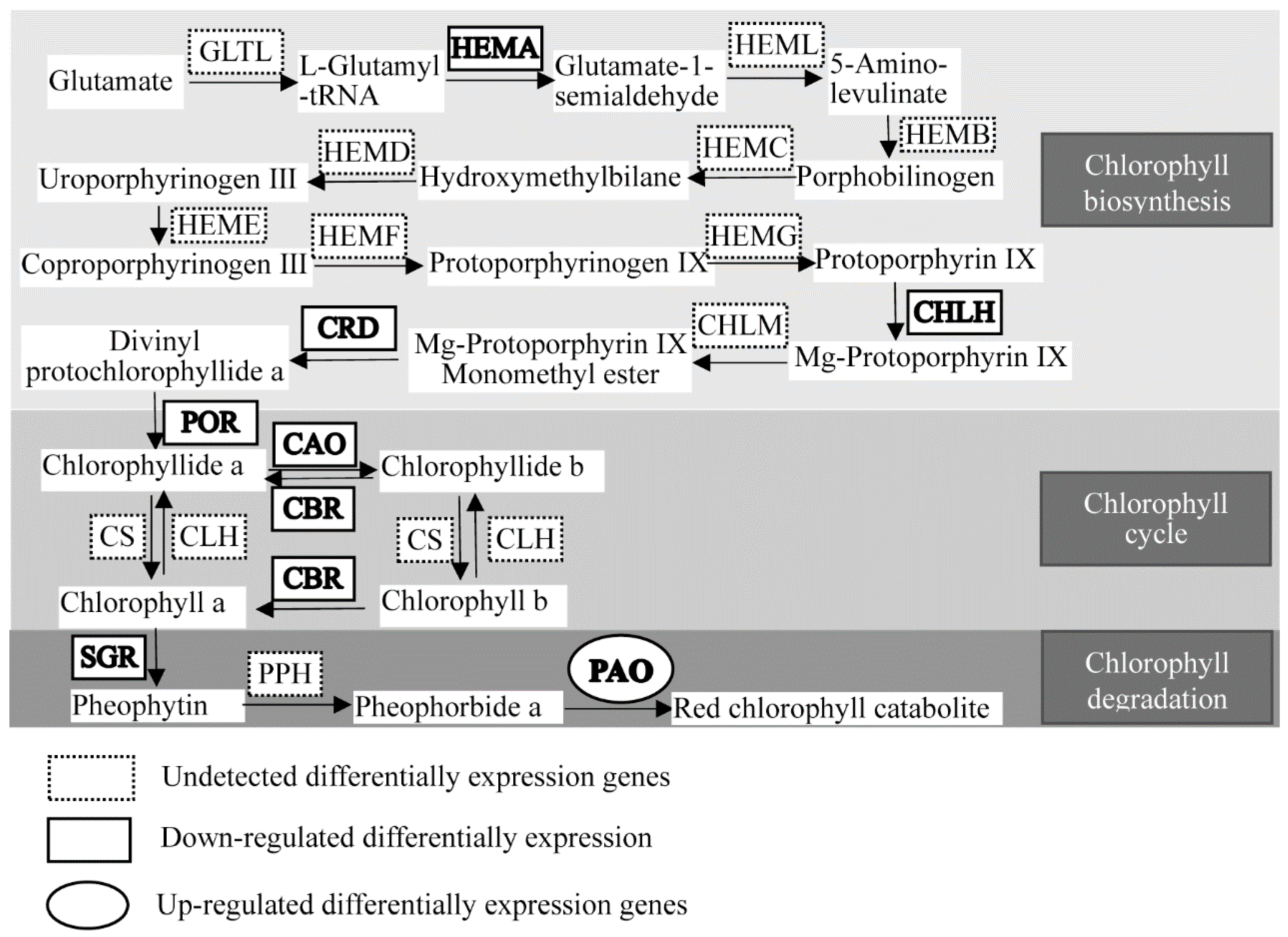
2.4. Chlorophyll Metabolism-Related Genes Expression Analysis
2.5. Identified Differentially Expressed Genes Involved in Photosynthesis
2.6. Response of Transcription Factors in the Comparison of AL vs. GL
3. Discussions
4. Materials and Methods
4.1. Plant Materials and Sample Preparation
4.2. Chlorophyll and Carotenoid Content Analysis
4.3. RNA Isolation, cDNA Library Preparation and Sequencing
4.4. Analysis of Differentially Expressed Genes
4.5. Illumina RNA-seq Result Validation by qRT-PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ma, C.; Cao, J.; Li, J.; Zhou, B.; Tang, J.; Miao, A. Phenotypic, histological and proteomic analyses reveal multiple differences associated with chloroplast development in yellow and variegated variants from Camellia sinensis. Sci. Rep. 2016, 6, 33369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Yue, C.; Cao, H.; Zhou, Y.; Zeng, J.; Yang, Y.; Wang, X. Biochemical and transcriptome analyses of a novel chlorophyll-deficient chlorina tea plant cultivar. BMC Plant Boil. 2014, 14, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Huang, J.; Liu, S.; Li, J.; Yang, X.; Liu, Y.; Liu, Z. Proteomic analysis of young leaves at three developmental stages in an albino tea cultivar. Proteome Sci. 2011, 9, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Chen, X.; Xu, B.; Li, Y.; Ma, Y.; Wang, G. Phenotype and transcriptome analysis reveals chloroplast development and pigment biosynthesis together influenced the leaf color formation in mutants of Anthurium andraeanum ‘Sonate’. Front. Plant Sci. 2015, 6, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Lu, X.; Xuan, Y.; Tang, F.; Wang, J.; Shi, D.; Fu, S.; Ren, J. Transcriptome analysis based on a combination of sequencing platforms provides insights into leaf pigmentation in Acer rubrum. BMC Plant Boil. 2019, 19, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Wang, M.; Zhang, Y.Z.; Du, L.F.; Pan, T. A chlorophyll-reduced seedling mutant in oilseed rape, Brassica napus, for utilization in F1 hybrid production. Plant Breed. 2000, 119, 131–135. [Google Scholar] [CrossRef]
- Jung, K.-H.; Hur, J.; Ryu, C.-H.; Choi, Y.; Chung, Y.-Y.; Miyao, A.; Hirochika, H.; An, G. Characterization of a rice chlorophyll-deficient mutant using the T-DNA gene-trap system. Plant Cell Physiol. 2003, 44, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.-H.; Hsu, C.-Y.; Cheng, H.-Y.; Chang, H.; Chen, H.-H.; Ger, M.-J. Downregulation of putative UDP-glucose: Flavonoid 3-O-glucosyltransferase gene alters flower coloring in Phalaenopsis. Plant Cell Rep. 2011, 30, 1007–1017. [Google Scholar] [CrossRef]
- Yasumura, Y.; Moylan, E.C.; Langdale, J. A conserved transcription factor mediates nuclear control of organelle biogenesis in anciently diverged land plants. Plant Cell 2005, 17, 1894–1907. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Xiao, L.; Zhang, Z.; Zhang, R.; Wang, Z.; Huang, C.; Huang, R.; Luan, Y.; Fan, T.; Wang, J.; et al. The genomes of pecan and Chinese hickory provide insights into Carya evolution and nut nutrition. GigaScience 2019, 8, 5. [Google Scholar] [CrossRef]
- Guo, W.; Chen, J.; Li, J.; Huang, J.; Wang, Z.; Lim, K.-J. Portal of Juglandaceae: A comprehensive platform for Juglandaceae study. Hortic. Res. 2020, 7, 35–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormac, A.C.; Terry, M.J. Light-signalling pathways leading to the co-ordinated expression of HEMA1 and Lhcb during chloroplast development in Arabidopsis thaliana. Plant J. 2002, 32, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Hong, X.; Hu, K.; Wang, Y.; Wang, X.; Du, S.; Li, Y.; Hu, D.; Cheng, K.; An, B.; et al. Impaired magnesium protoporphyrin IX methyltransferase (ChlM) impedes chlorophyll synthesis and plant growth in rice. Front. Plant Sci. 2017, 8, 1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inagaki, N.; Kinoshita, K.; Kagawa, T.; Tanaka, A.; Ueno, O.; Shimada, H.; Takano, M. Phytochrome B mediates the regulation of chlorophyll biosynthesis through transcriptional regulation of ChlH and GUN4 in rice seedlings. PLoS ONE 2015, 10, e0135408. [Google Scholar] [CrossRef]
- Shi, C.; Shi, X.-M. Characterization of three genes encoding the subunits of light-independent protochlorophyllide reductase in chlorella protothecoides CS-41. Biotechnol. Prog. 2006, 22, 1050–1055. [Google Scholar] [CrossRef]
- Stephenson, P.G.; Terry, M.J. Light signalling pathways regulating the Mg-chelatase branchpoint of chlorophyll synthesis during de-etiolation in Arabidopsis thaliana. Photochem. Photobiol. Sci. 2008, 7, 1243. [Google Scholar] [CrossRef]
- Ji-Yu, Z.; Pan, D.-L.; Jia, Z.-H.; Wang, T.; Wang, G.; Guo, Z. Chlorophyll, carotenoid and vitamin C metabolism regulation in Actinidia chinensis ’Hongyang’ outer pericarp during fruit development. PLoS ONE 2018, 13, e0194835. [Google Scholar] [CrossRef]
- Tanaka, R.; Tanaka, A. Chlorophyll cycle regulates the construction and destruction of the light-harvesting complexes. Biochim. Biophys. Acta (BBA) Bioenerg. 2011, 1807, 968–976. [Google Scholar] [CrossRef] [Green Version]
- Rüdiger, W. Biosynthesis of chlorophyll b and the chlorophyll cycle. Photosynth. Res. 2002, 74, 187–193. [Google Scholar] [CrossRef]
- Kanematsu, S.; Sakuraba, Y.; Tanaka, A.; Tanaka, R. Characterization of Arabidopsis mutants defective in the regulation of chlorophyllide a oxygenase. Photochem. Photobiol. Sci. 2008, 7, 1196. [Google Scholar] [CrossRef]
- Tanaka, R.; Koshino, Y.; Sawa, S.; Ishiguro, S.; Okada, K.; Tanaka, A. Overexpression of chlorophyllide a oxygenase (CAO) enlarges the antenna size of photosystem II in Arabidopsis thaliana. Plant J. 2001, 26, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, S.; Ito, H.; Tanaka, R.; Tanaka, A. Chlorophyll b reductase plays an essential role in maturation and storability of Arabidopsis seeds. Plant Physiol. 2012, 160, 261–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Liu, T.; Xu, Y.; Chen, W.; Liu, B.; Zhang, L.; Liu, D.-C.; Zhang, H.; Zhang, B. Comparative transcriptome analysis of two selenium-accumulating genotypes of Aegilops tauschii Coss. in response to selenium. BMC Genet. 2019, 20, 9. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Xu, X.; Hu, X.; Liu, Y.; Cao, H.; Chan, H.; Gong, Z.; Yuan, Y.; Luo, Y.; Feng, B.; et al. SlMYB72 regulates the metabolism of chlorophylls, carotenoids, and flavonoids in tomato fruit. Plant Physiol. 2020, 183, 854–868. [Google Scholar] [CrossRef] [PubMed]
- Ampomah-Dwamena, C.; Thrimawithana, A.H.; Dejnoprat, S.; Lewis, D.; Espley, R.V.; Allan, A.C. A kiwifruit (Actinidia deliciosa) R2R3-MYB transcription factor modulates chlorophyll and carotenoid accumulation. New Phytol. 2018, 221, 309–325. [Google Scholar] [CrossRef] [Green Version]
- Wen, C.-H.; Lin, S.-S.; Chu, F.-H. Transcriptome analysis of a subtropical deciduous tree: Autumn leaf senescence gene expression profile of Formosan Gum. Plant Cell Physiol. 2014, 56, 163–174. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Xu, Y.-H.; Jiang, S.-C.; Lu, K.; Lu, Y.-F.; Feng, X.-J.; Wu, Z.; Liang, S.; Yu, Y.-T.; Wang, X.-F.; et al. Light-harvesting chlorophyll a/b-binding proteins, positively involved in abscisic acid signalling, require a transcription repressor, WRKY40, to balance their function. J. Exp. Bot. 2013, 64, 5443–5456. [Google Scholar] [CrossRef] [Green Version]
- Ma, N.-N.; Feng, H.; Meng, X.; Li, D.; Yang, D.; Wu, C.; Meng, Q. Overexpression of tomato SlNAC1 transcription factor alters fruit pigmentation and softening. BMC Plant Boil. 2014, 14, 351. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Chen, G.; Zhou, S.; Tu, Y.; Wang, Y.; Dong, T.; Hu, Z. A New Tomato NAC (NAM/ATAF1/2/CUC2) transcription factor, SlNAC4, functions as a positive regulator of fruit ripening and carotenoid accumulation. Plant Cell Physiol. 2013, 55, 119–135. [Google Scholar] [CrossRef] [Green Version]
- Montefiori, M.; McGhie, T.; Hallett, I.C.; Costa, G. Changes in pigments and plastid ultrastructure during ripening of green-fleshed and yellow-fleshed kiwifruit. Sci. Hortic. 2009, 119, 377–387. [Google Scholar] [CrossRef]
- Tong, Z.; Qu, S.; Zhang, J.; Wang, F.; Tao, J.; Gao, Z.; Zhang, Z. A Modified protocol for RNA extraction from different peach tissues suitable for gene isolation and real-time PCR analysis. Mol. Biotechnol. 2011, 50, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, D.-L.; Wang, G.; Wang, T.; Jia, Z.-H.; Guo, Z.; Ji-Yu, Z. AdRAP2.3, a novel ethylene response factor VII from Actinidia deliciosa, enhances waterlogging resistance in transgenic tobacco through improving expression levels of PDC and ADH genes. Int. J. Mol. Sci. 2019, 20, 1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, Z.; Feng, G.; Su, W.; Liu, Z.; Peng, F. Transcriptomic analysis provides insights into grafting union development in pecan (Carya illinoinensis). Genes 2018, 9, 71. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)). Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Group Name | GL | AL |
|---|---|---|
| No. of total reads (×106) | 38.51 ± 5.36 | 38.68 ± 4.23 |
| No. of mapped reads (×106) | 35.87 ± 4.90 | 35.78 ± 4.08 |
| Mapped percentage | 93.15% ± 0.27% | 92.46% ± 0.47% |
| Unique Mapped reads (×106) | 34.93 ± 3.87 | 34.90 ± 3.21 |
| Database | 37, 254 All Unigenes | 5171 Differentially Expressed Genes | ||
|---|---|---|---|---|
| Number of Annotated Sequences | Percentage of Annotated Sequences | Number of Annotated Sequences | Percentage of Annotated Sequences | |
| NR (E-value < 10−5) | 33,859 | 90.89 | 4389 | 84.88 |
| SwissProt | 26,558 | 71.29 | 3596 | 69.54 |
| COG | 11,789 | 31.64 | 1546 | 29.90 |
| KEGG | 7701 | 20.67 | 939 | 18.16 |
| GO | 24,670 | 66.22 | 3337 | 64.53 |
| Pathway | Pathway ID | Q-Value | DEGs |
|---|---|---|---|
| Photosynthesis | ko00195 | 1.86 × 10−24 | 51 |
| Photosynthesis-antenna proteins | ko00196 | 2.86 × 10−14 | 22 |
| Metabolic pathways | ko01100 | 7.84 × 10−11 | 360 |
| Nitrogen metabolism | ko00910 | 1.66 × 10−7 | 32 |
| Biosynthesis of secondary metabolites | ko01110 | 1.70 × 10−6 | 182 |
| Carbon fixation in photosynthetic organisms | ko00710 | 2.41 × 10−6 | 35 |
| Starch and sucrose metabolism | ko00500 | 1.01 × 10−5 | 57 |
| Glyoxylate and dicarboxylate metabolism | ko00630 | 0.0008132 | 24 |
| Alanine, aspartate and glutamate metabolism | ko00250 | 0.0008106 | 22 |
| Glycine, serine and threonine metabolism | ko00260 | 0.0008446 | 25 |
| Genes ID | Protein | Gene | log2(AL/GL) | p-Value | Regulated |
|---|---|---|---|---|---|
| CIL0203S0025 | PREDICTED: glutamyl-tRNA reductase 1, chloroplastic-like | HEMA | −1.16362526 | 1.22 × 10−7 | down |
| CIL1034S0072 | PREDICTED: magnesium-chelatase subunit ChlH, chloroplastic | CHLH | −2.02856009 | 5.50 × 10−24 | down |
| CIL1166S0028 | PREDICTED: magnesium-protoporphyrin (oxidative) cyclase, chloroplastic | CRD | −2.02856009 | 5.50 × 10−24 | down |
| CIL1444S0041 | PREDICTED: chloroplastic | POR | −1.85284122 | 1.14 × 10−19 | down |
| CIL1192S0055 | PREDICTED: chlorophyllide a oxygenase, chloroplastic isoform ×1 | CAO | −1.97608744 | 4.99 × 10−43 | down |
| CIL1335S0038 | PREDICTED: chlorophyllide a oxygenase, chloroplastic-like | CAO | −2.03958666 | 1.67 × 10−16 | down |
| CIL0897S0166 | PREDICTED: probable chlorophyll(ide) b reductase NYC1, chloroplastic | CBR | −1.31052847 | 1.92 × 10−21 | down |
| CIL1224S0038 | PREDICTED: probable chlorophyll(ide) b reductase NYC1, chloroplastic | CBR | −1.21082238 | 1.61 × 10−15 | down |
| CIL0897S0167 | PREDICTED: probable chlorophyll(ide) b reductase NYC1, chloroplastic | CBR | −1.31052847 | 1.92 × 10−21 | down |
| CIL1230S0045 | PREDICTED: protein STAY-GREEN LIKE, chloroplastic-like | SGR | −1.67002437 | 1.22 × 10−13 | down |
| CIL0946S0047 | PREDICTED: pheophorbide a oxygenase, chloroplastic-like | PAO | 1.48825352 | 2.60 × 10−58 | up |
| CIL1523S0003 | PREDICTED: pheophorbide a oxygenase, chloroplastic-like | PAO | 1.232943538 | 1.36 × 10−8 | up |
| Genes ID | Protein | Gene | log2 (AL/GL) | p-Value | Regulated |
|---|---|---|---|---|---|
| Photosystem I | |||||
| MSTRG.12608 | Photosystem I P700 apoprotein A1 (chloroplast) | PsaA | −1.70701 | 1.18 × 10−9 | down |
| MSTRG.1756 | Photosystem I iron-sulfur center | PsaC | 1.571126 | 5.12 × 10−13 | up |
| CIL1061S0088 | PREDICTED: photosystem I reaction center subunit II, chloroplastic-like | PsaD | −2.06416 | 6.02 × 10−46 | down |
| CIL1130S0012 | PREDICTED: photosystem I reaction center subunit IV, chloroplastic-like | PsaE | −2.64087 | 1.29 × 10−66 | down |
| CIL1293S0081 | PREDICTED: photosystem I reaction center subunit IV B, chloroplastic-like | PsaE2 | −2.24447 | 1.54 × 10−28 | down |
| CIL0957S0008 | PREDICTED: photosystem I reaction center subunit III, chloroplastic-like | PsaF | −2.27227 | 4.74 × 10−43 | down |
| CIL1454S0024 | PREDICTED: photosystem I reaction center subunit V, chloroplastic | PsaG | −2.15252 | 1.79 × 10−45 | down |
| CIL0121S0006 | PREDICTED: photosystem I reaction center subunit VI, chloroplastic-like | PsaH | −3.31839 | 1.62 × 10−68 | down |
| CIL1184S0026 | PREDICTED: photosystem I reaction center subunit VI-2, chloroplastic | PsaH2 | −1.80355 | 9.55 × 10−31 | down |
| CIL0225S0008 | PREDICTED: photosystem I reaction center subunit psaK, chloroplastic-like | PsaK | −2.88506 | 7.68 × 10−70 | down |
| CIL1120S0015 | PREDICTED: photosystem I reaction center subunit psaK, chloroplastic-like | PsaK | −2.10015 | 0.000806 | down |
| CIL0070S0007 | PREDICTED: photosystem I reaction center subunit XI, chloroplastic | PsaL | −2.34772 | 8.07 × 10−41 | down |
| CIL0479S0005 | PREDICTED: photosystem I reaction center subunit N, chloroplastic-like | PsaN | −2.98776 | 1.44 × 10−13 | down |
| CIL1492S0029 | PREDICTED: photosystem I reaction center subunit N, chloroplastic | PsaN | −2.11471 | 2.07 × 10−42 | down |
| CIL0899S0004 | PREDICTED: photosystem I subunit O | PsaO | −2.38235 | 8.89 × 10−52 | down |
| Photosystem II | |||||
| CIL0840S0001 | Putative photosystem II protein D1(Helianthus annuus) | PsbA | −3.25158 | 9.61 × 10−43 | down |
| MSTRG.6696 | Photosystem II CP47 chlorophyll apoprotein, partial (plastid) | PsbB | −2.09893 | 0.001337 | down |
| MSTRG.28118 | PsbB, partial (chloroplast) | PsbB | −1.90447 | 0.000545 | down |
| MSTRG.12382 | Photosystem II protein K (chloroplast) | psbK | −2.14062 | 5.63 × 10−16 | down |
| MSTRG.11850 | Photosystem II protein K (chloroplast) | psbK | −2.0512 | 4.08 × 10−29 | down |
| CIL1409S0029 | PREDICTED: oxygen-evolving enhancer protein 1, chloroplastic-like | PsbO | −3.25073 | 1.79 × 10−72 | down |
| CIL0990S0108 | PREDICTED: psbP-like protein 1, chloroplastic | PsbP | −1.03085 | 9.74 × 10−12 | down |
| CIL0990S0110 | PREDICTED: photosynthetic NDH subunit of lumenal location 1, chloroplastic | PsbP | −1.82176 | 6.22 × 10−33 | down |
| CIL1351S0018 | PREDICTED: oxygen-evolving enhancer protein 2, chloroplastic | PsbP | −1.81357 | 1.90 × 10−31 | down |
| CIL1099S0039 | PREDICTED: oxygen-evolving enhancer protein 3-2, chloroplastic-like | PsbQ | −2.36913 | 7.49 × 10−56 | down |
| CIL1258S0023 | PREDICTED: photosynthetic NDH subunit of lumenal location 2, chloroplastic | PsbQ | −2.24784 | 9.86 × 10−43 | down |
| CIL1577S0034 | PREDICTED: photosynthetic NDH subunit of lumenal location 3, chloroplastic | PsbQ | −2.0615 | 1.01 × 10−36 | down |
| CIL1048S0062 | PREDICTED: photosystem II 10 kDa polypeptide, chloroplastic | PsbR | −1.39211 | 3.63 × 10−21 | down |
| CIL1112S0006 | PREDICTED: photosystem II 22 kDa protein, chloroplastic | PsbS | −1.62697 | 6.31 × 10−13 | down |
| CIL1192S0058 | PREDICTED: photosystem II 22 kDa protein, chloroplastic | PsbS | −1.7504 | 6.05 × 10−43 | down |
| CIL0970S0081 | PREDICTED: photosystem II reaction center W protein, chloroplastic-like | PsbW | −2.1531 | 1.81 × 10−36 | down |
| CIL1034S0038 | PREDICTED: photosystem II reaction center W protein, chloroplastic-like isoform X1 | PsbW | −1.00199 | 4.32 × 10−5 | down |
| CIL0929S0055 | PREDICTED: photosystem II core complex proteins psbY, chloroplastic | PsbY | −2.07095 | 1.48 × 10−28 | down |
| CIL1040S0004 | PREDICTED: photosystem II repair protein PSB27-H1, chloroplastic | PSB27-H1 | −2.45399 | 7.91 × 10−65 | down |
| Cytochrome b6-f complex | |||||
| MSTRG.12087 | Cytochrome b6 (chloroplast) | petB | −1.12581 | 7.22 × 10−5 | down |
| CIL1135S0039 | PREDICTED: cytochrome b6-f complex iron-sulfur subunit 1, chloroplastic-like | petC | −1.62437 | 1.20 × 10−24 | down |
| CIL1405S0081 | PREDICTED: cytochrome b6-f complex iron-sulfur subunit 1, chloroplastic-like | petC | −1.64744 | 2.11 × 10−47 | down |
| Photosynthetic electron transport | |||||
| CIL0131S0022 | Plastocyanin | petE | −3.7498 | 9.31 × 10−59 | down |
| CIL1192S0070 | PREDICTED: plastocyanin B'/B'&apos | petE | −2.19915 | 7.47 × 10−48 | down |
| CIL1082S0115 | PREDICTED: ferredoxin-like | petF | −1.92418 | 1.88 × 10−68 | down |
| CIL1146S0002 | ferredoxin--NADP reductase, leaf-type isozyme, chloroplastic | petH | −1.84692 | 6.72 × 10−27 | down |
| CIL1219S0022 | PREDICTED: ferredoxin--NADP reductase, root isozyme, chloroplastic | petH | 1.558778 | 2.18 × 10−11 | up |
| CIL1099S0062 | PREDICTED: cytochrome c6, chloroplastic-like | petJ | −1.07258 | 1.22 × 10−7 | down |
| F-Type ATPase | |||||
| CIL0009S0016 | ATP synthase beta subunit, partial (chloroplast) | atpB | −2.28295 | 1.64 × 10−10 | down |
| CIL0922S0023 | PREDICTED: ATP synthase gamma chain, chloroplastic | atpC | −1.31948 | 1.26 × 10−32 | down |
| CIL1157S0078 | PREDICTED: ATP synthase delta chain, chloroplastic-like | atpD | −2.8914 | 8.90 × 10−61 | down |
| CIL1064S0097 | PREDICTED: ATP synthase subunit delta, chloroplastic-like | atpH | −1.74555 | 4.63 × 10−24 | down |
| CIL0936S0006 | ATP synthase F0, A subunit | A | 1.238679 | 3.76 × 10−5 | up |
| CIL1413S0005 | REDICTED: ATP synthase subunit b' chloroplastic | B | −1.73514 | 7.32 × 10−46 | down |
| MSTRG.3669 | ATPase subunit 8 (mitochondrion) | B | 1.377949 | 0.007137 | up |
| Photosynthesis—antenna proteins | |||||
| CIL1196S0069 | PREDICTED: chlorophyll a-b binding protein 6A, chloroplastic-like | LHCA1 | −3.0003 | 1.28 × 10−57 | down |
| CIL1348S0032 | PREDICTED: chlorophyll a-b binding protein 6, chloroplastic-like | LHCA1 | −3.25903 | 1.92 × 10−53 | down |
| CIL1458S0020 | Chlorophyll a-b binding protein, chloroplastic | LHCA2 | −2.35787 | 3.49 × 10−50 | down |
| CIL1486S0034 | PREDICTED: chlorophyll a-b binding protein, chloroplastic | LHCA2 | −2.93568 | 1.46 × 10−48 | down |
| CIL1118S0087 | PREDICTED: chlorophyll a-b binding protein 8, chloroplastic | LHCA3 | −2.10753 | 3.33 × 10−45 | down |
| CIL1578S0015 | Chlorophyll a-b binding protein 4, chloroplastic | LHCA4 | −3.58717 | 1.06 × 10−73 | down |
| CIL1507S0004 | PREDICTED: photosystem I chlorophyll a/b-binding protein 5, chloroplastic | LHCA5 | −1.20413 | 1.03 × 10−14 | down |
| CIL1582S0038 | PREDICTED: photosystem I chlorophyll a/b-binding protein 6, chloroplastic | LHCA6 | −2.17269 | 8.46 × 10−33 | down |
| CIL0258S0015 | Chlorophyll a-b binding protein of LHCII type 1 | LHCB1 | −2.83684 | 3.34 × 10−44 | down |
| CIL1047S0026 | Chlorophyll a-b binding protein of LHCII type 1 | LHCB1 | −3.63831 | 9.87 × 10−74 | down |
| CIL1187S0057 | Chlorophyll a-b binding protein of LHCII type 1 | LHCB1 | −2.82388 | 1.67 × 10−29 | down |
| CIL1393S0038 | Chlorophyll a-b binding protein of LHCII type 1 | LHCB1 | −4.16137 | 5.33 × 10−123 | down |
| CIL1384S0010 | PREDICTED: chlorophyll a-b binding protein 151, chloroplastic | LHCB2 | −3.65164 | 3.05 × 10−102 | down |
| CIL1078S0110 | PREDICTED: chlorophyll a-b binding protein 13, chloroplastic | LHCB3 | −4.01404 | 5.76 × 10−57 | down |
| CIL1135S0055 | PREDICTED: chlorophyll a-b binding protein 13, chloroplastic | LHCB3 | −6.02437 | 4.65 × 10−51 | down |
| CIL1122S0046 | PREDICTED: chlorophyll a-b binding protein CP29.1, chloroplastic-like | LHCB4 | −2.90723 | 5.76 × 10−59 | down |
| CIL1204S0057 | PREDICTED: chlorophyll a-b binding protein CP29.3, chloroplastic | LHCB4 | −3.62946 | 1.16 × 10−51 | down |
| CIL0010S0054 | PREDICTED: chlorophyll a-b binding protein CP26, chloroplastic-like | LHCB5 | −2.54986 | 7.61 × 10−42 | down |
| CIL0948S0124 | PREDICTED: chlorophyll a-b binding protein CP26, chloroplastic | LHCB5 | −1.60257 | 0.000693 | down |
| CIL0948S0127 | PREDICTED: chlorophyll a-b binding protein CP26, chloroplastic | LHCB5 | −2.17918 | 5.06 × 10−35 | down |
| CIL0424S0004 | PREDICTED: chlorophyll a-b binding protein CP24 10A, chloroplastic | LHCB6 | −2.83352 | 1.64 × 10−41 | down |
| CIL1082S0041 | PREDICTED: chlorophyll a-b binding protein CP24 10A, chloroplastic-like | LHCB6 | −3.78467 | 7.49 × 10−69 | down |
| Category | Total | Upregulation | Downregulation |
|---|---|---|---|
| MYB | 277 | 16 | 24 |
| AP2/ERF | 197 | 23 | 12 |
| NAC | 122 | 25 | 8 |
| C2C2 | 110 | 6 | 16 |
| C2H2 | 149 | 14 | 6 |
| bHLH | 159 | 8 | 11 |
| WRKY | 93 | 7 | 12 |
| HB | 112 | 9 | 4 |
| bZIP | 81 | 7 | 5 |
| GARP | 63 | 3 | 5 |
| LOB | 54 | 4 | 4 |
| B3 | 69 | 3 | 4 |
| MADS | 63 | 2 | 5 |
| AUX/IAA | 43 | 1 | 5 |
| GRF | 14 | 6 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.-Y.; Wang, T.; Jia, Z.-H.; Guo, Z.-R.; Liu, Y.-Z.; Wang, G. Photosystem Disorder Could be the Key Cause for the Formation of Albino Leaf Phenotype in Pecan. Int. J. Mol. Sci. 2020, 21, 6137. https://doi.org/10.3390/ijms21176137
Zhang J-Y, Wang T, Jia Z-H, Guo Z-R, Liu Y-Z, Wang G. Photosystem Disorder Could be the Key Cause for the Formation of Albino Leaf Phenotype in Pecan. International Journal of Molecular Sciences. 2020; 21(17):6137. https://doi.org/10.3390/ijms21176137
Chicago/Turabian StyleZhang, Ji-Yu, Tao Wang, Zhan-Hui Jia, Zhong-Ren Guo, Yong-Zhi Liu, and Gang Wang. 2020. "Photosystem Disorder Could be the Key Cause for the Formation of Albino Leaf Phenotype in Pecan" International Journal of Molecular Sciences 21, no. 17: 6137. https://doi.org/10.3390/ijms21176137