Identification and Expression Analysis of Hormone Biosynthetic and Metabolism Genes in the 2OGD Family for Identifying Genes That May Be Involved in Tomato Fruit Ripening


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
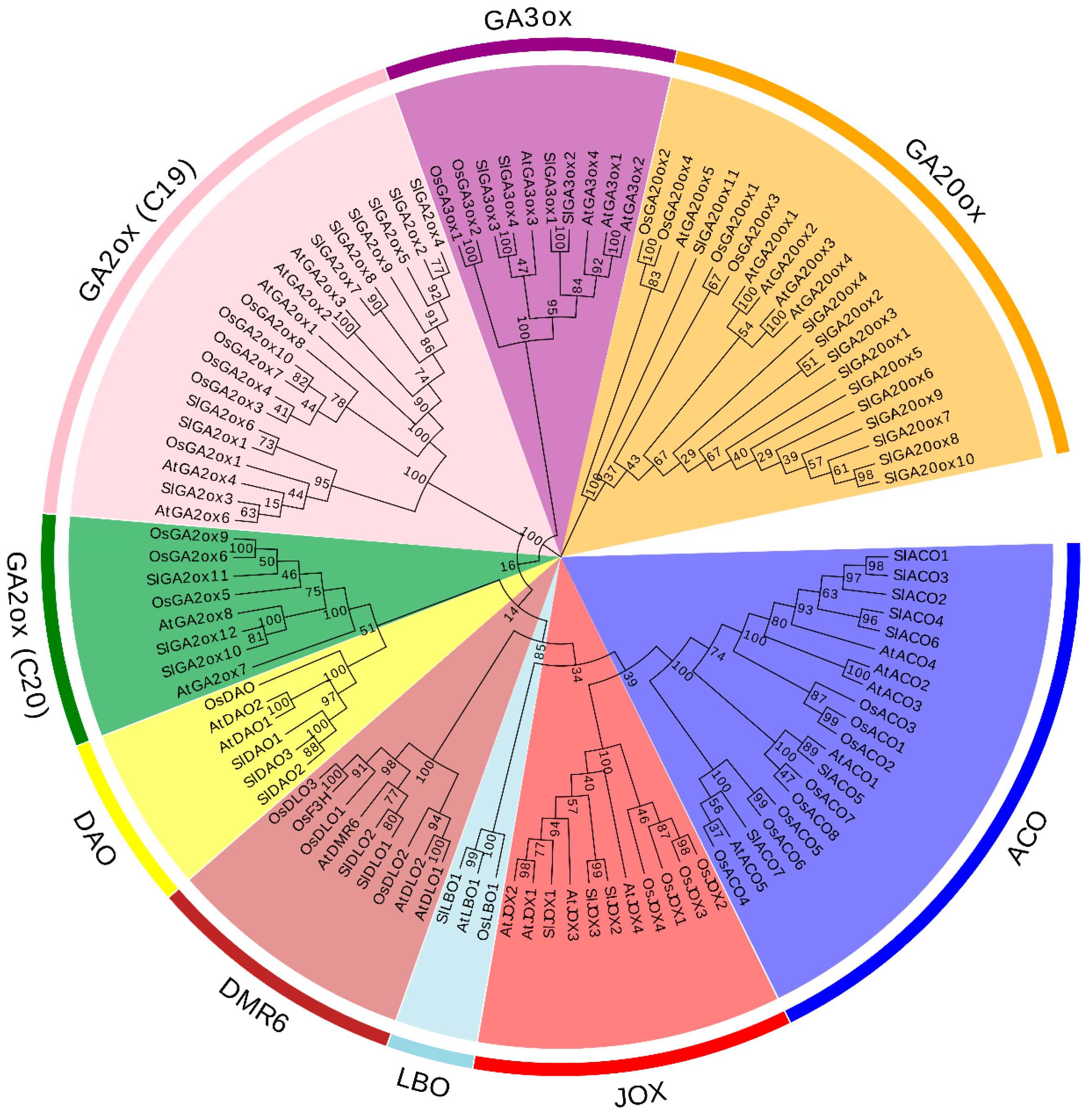
2.1. Identification and Phylogenetic Analysis of Hormone Biosynthetic and Metabolism Proteins in 2OGD Superfamily
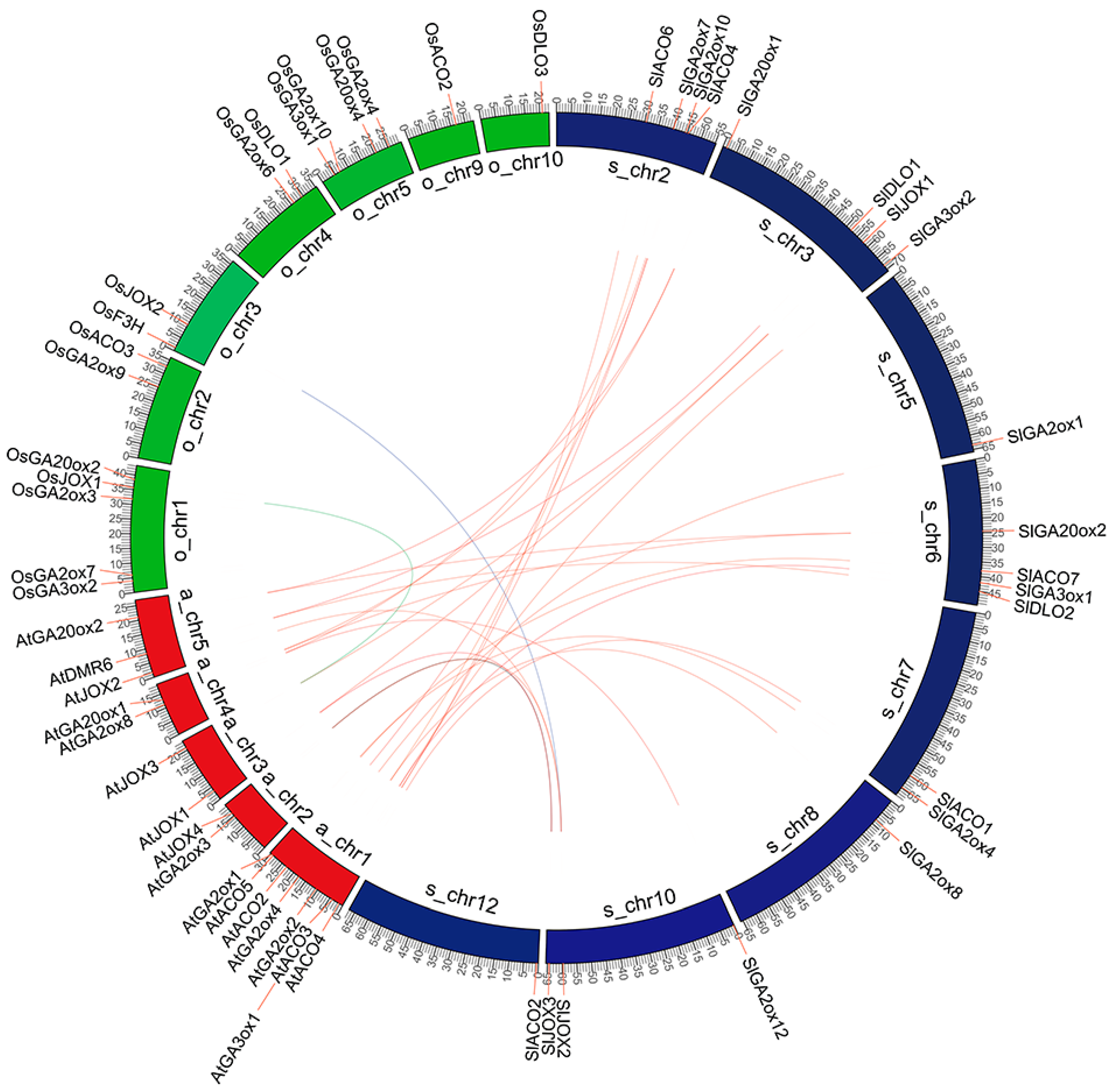
2.2. Synteny and Duplication Analysis of Hormone Biosynthetic and Metabolism Proteins in 2OGD Superfamily
2.3. Multiple Sequence Alignment and Motif Composition Analysis of Hormone Biosynthetic and Metabolism 2OGD Proteins
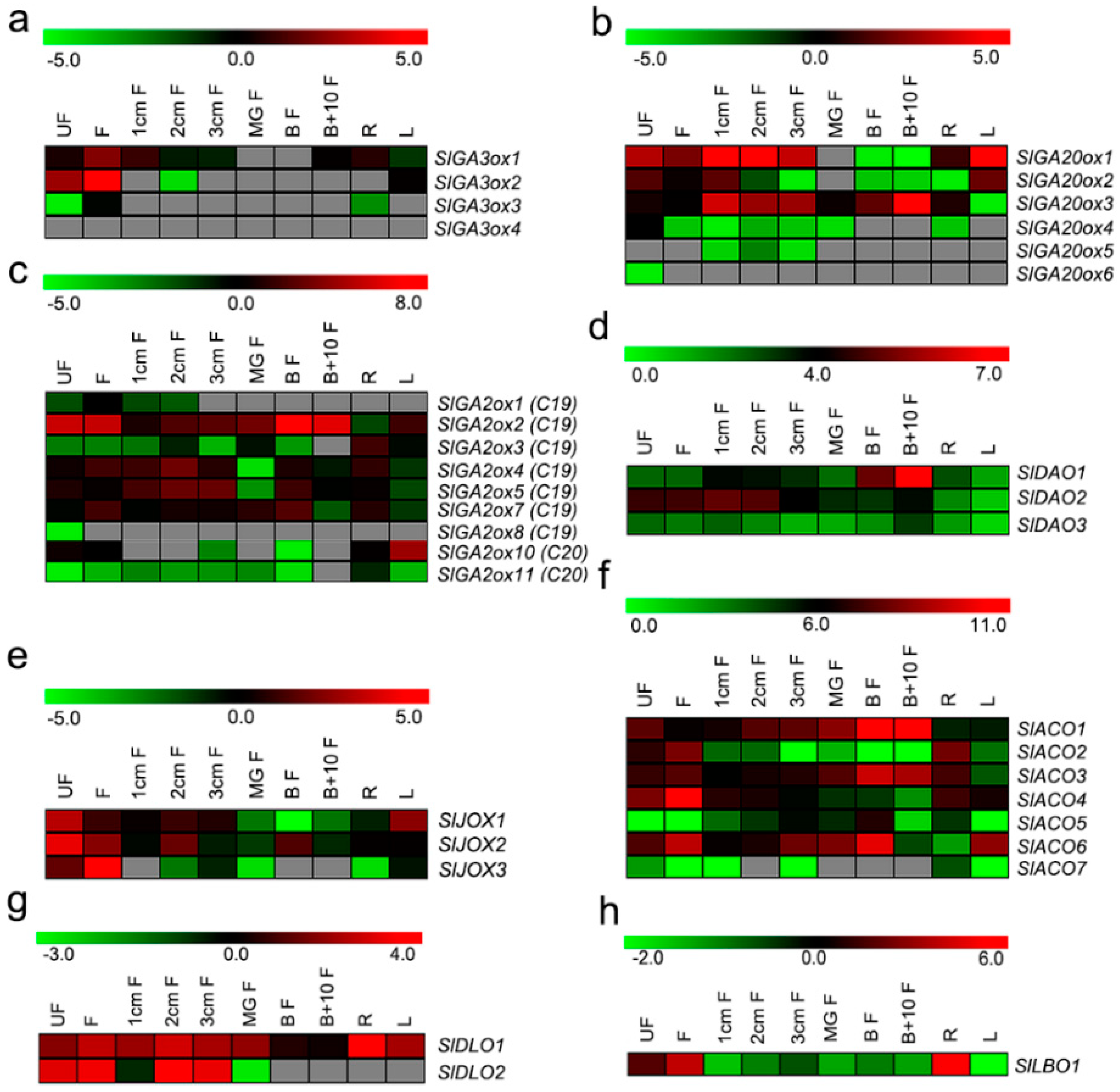
2.4. Expression of Hormone Biosynthetic and Metabolism 2OGD Genes in Tomato
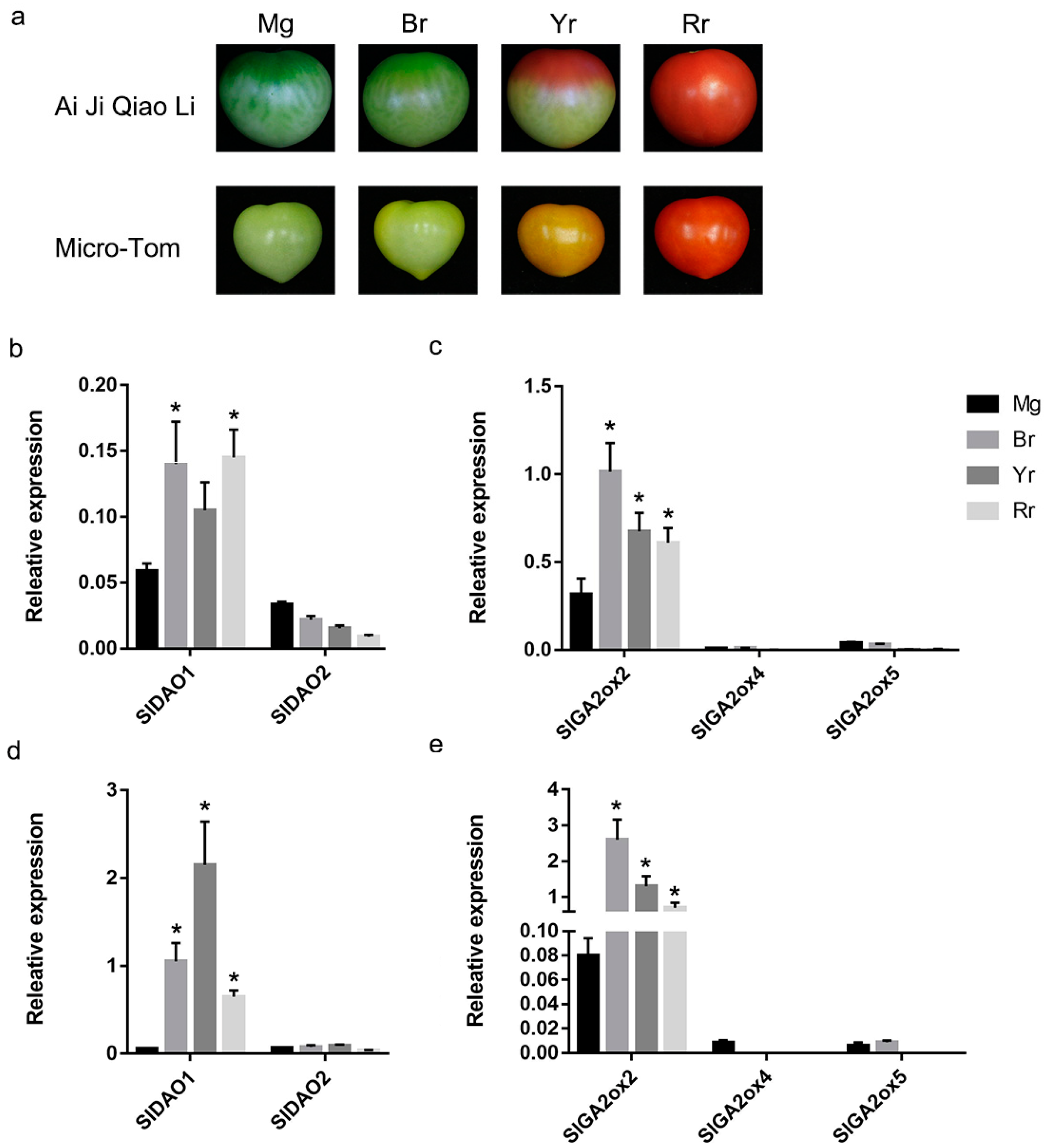
2.5. Expression of SlGA2ox and SlDAO Genes during Tomato Fruit Ripening
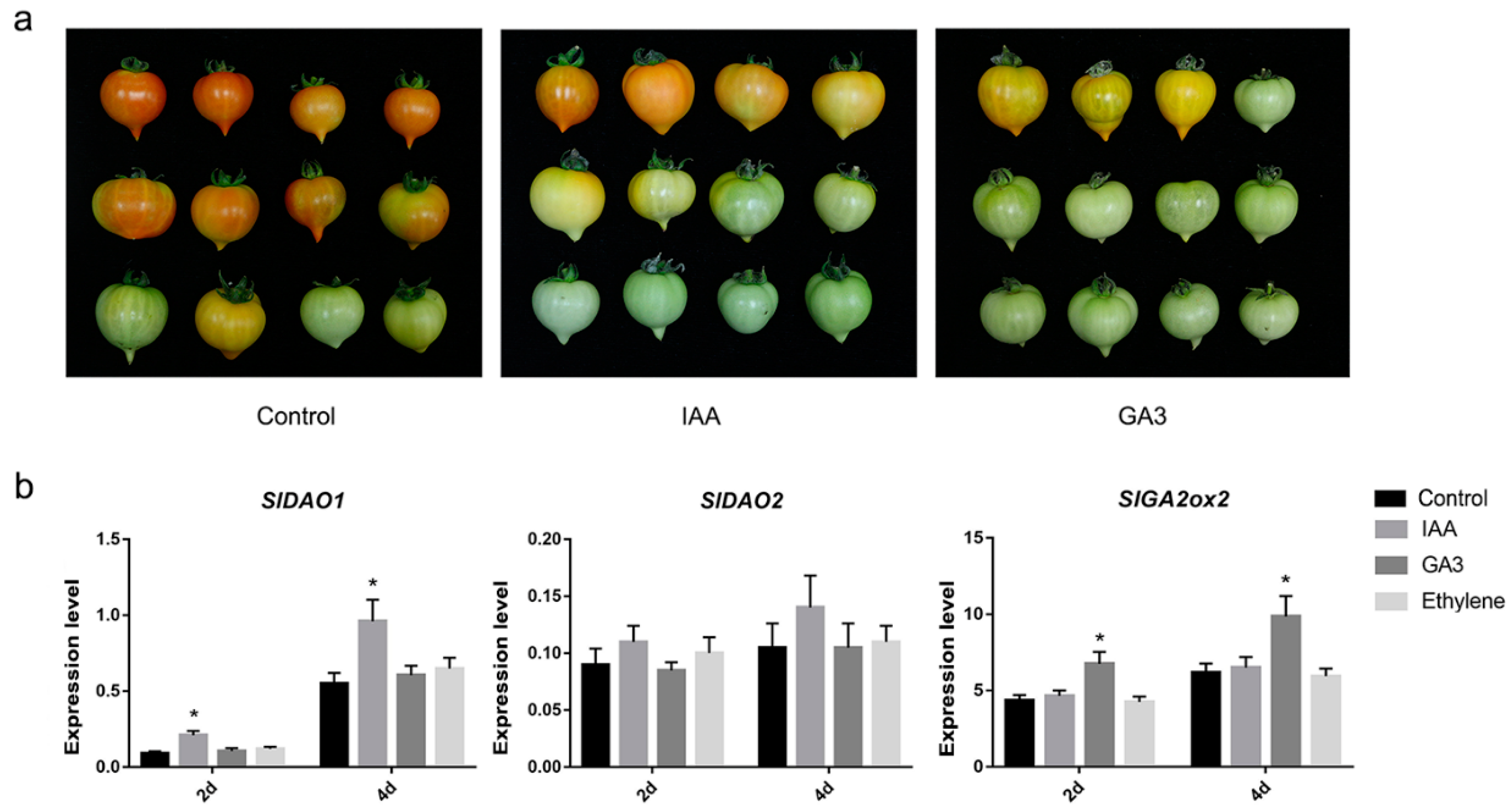
2.6. Effects of Auxin, GA3, and Ethylene on the Expression of SlDAO1, SlDAO2, and SlGA2ox2
3. Discussion
3.1. Identification of Hormone Biosynthetic and Metabolism Genes from 2OGD Family
3.2. Functional Analysis of Hormone Biosynthetic and Metabolism Genes in 2OGD Family
3.3. SlGA2ox2 and SlDAO1 May Play a Role in GA and Auxin Metabolism for Normal Ripening of Tomato Fruits
4. Materials and Methods
4.1. Identification and Phylogenetic Analysis of Hormone Biosynthesis and Metabolism Related DOXC Proteins
4.2. Chromosomal Location and Synteny Analysis
4.3. Multiple Sequence Alignment and Motif Composition Analysis
4.4. Expression Analysis
4.5. Plant Materials and Hormone Treatments
4.6. RNA Extraction and qPCR Analysis of Selected Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| GAs | Gibberellins |
| JA | Jasmonate |
| SA | Salicylic acid |
| SL | Strigolactone |
| GA20ox | GA20-oxidase |
| GA3ox | GA3-oxidase |
| GA2ox | GA2-oxidase |
| DAO | Dioxygenase for Auxin Oxidation |
| ACO | 1-aminocyclopropane-1-carboxylic acid oxidase |
| JOX | JASMONATE-INDUCED OXYGENASE |
| DMR6 | Downy Mildew Resistant6 |
| DLO | DMR6-LIKE OXYGENASE |
| LBO | LATERAL BRANCHING OXIDOREDUCTASE |
| FPKM | Fragments Per Kilobase Per Million |
References
- Su, L.; Diretto, G.; Purgatto, E.; Danoun, S.; Zouine, M.; Li, Z.; Roustan, J.P.; Bouzayen, M.; Giuliano, G.; Chervin, C. Carotenoid accumulation during tomato fruit ripening is modulated by the auxin-ethylene balance. BMC Plant Biol. 2015, 15, 114–125. [Google Scholar] [CrossRef]
- Li, J.Y.; Tao, X.Y.; Li, L.; Mao, L.; Luo, Z.S.; Khan, Z.U.; Ying, T. Comprehensive RNA-Seq analysis on the regulation of tomato ripening by exogenous auxin. PLoS ONE 2016, 11, e0156453. [Google Scholar] [CrossRef]
- Sravankumar, T.; Akash; Naik, N.; Kumar, R. A ripening-induced SlGH3-2 gene regulates fruit ripening via adjusting auxin-ethylene levels in tomato (Solanum lycopersicum L.). Plant Mol. Biol. 2018, 98, 455–469. [Google Scholar] [CrossRef]
- Li, H.; Wu, H.; Qi, Q.; Li, H.; Li, Z.; Chen, S.; Ding, Q.; Wang, Q.; Yan, Z.; Gai, Y.; et al. Gibberellins play a role in regulating tomato fruit ripening. Plant Cell Physiol. 2019, 60, 1619–1629. [Google Scholar] [CrossRef]
- Farrow, S.C.; Facchini, P.J. Functional diversity of 2-oxoglutarate/Fe(II)-dependent dioxygenases in plant metabolism. Front Plant Sci. 2014, 5, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedden, P.; Thomas, S.G. Gibberellin biosynthesis and its regulation. Biochem. J. 2012, 444, 11–25. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Zhang, Y.; Liu, X.; Zhang, X.; Liu, S.; Yu, X.; Ren, Y.; Zheng, X.; Zhou, K.; Jiang, L.; et al. A role for a dioxygenase in auxin metabolism and reproductive development in rice. Dev. Cell. 2013, 27, 113–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argueso, C.T.; Hansen, M.; Kieber, J.J. Regulation of ethylene biosynthesis. J. Plant Growth Regul. 2007, 26, 92–105. [Google Scholar] [CrossRef]
- Caarls, L.; Elberse, J.; Awwanah, M.; Ludwig, N.R.; Vries, D.; Zeilmaker, T.; Wees, S.C.M.V.; Schuurink, R.C.; Ackerveken, G.V. Arabidopsis JASMONATE-INDUCED OXYGENASES down-regulate plant immunity by hydroxylation and inactivation of the hormone jasmonic acid. Proc. Natl. Acad. Sci. USA 2017, 114, 6388–6393. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhao, L.; Zhao, J.; Li, Y.; Wang, J.; Guo, R.; Gan, S.; Liu, C.; Zhang, W. S5H/DMR6 encodes a salicylic acid 5-hydroxylase that fine-tunes salicylic acid homeostasis. Plant Physiol. 2017, 175, 1082–1093. [Google Scholar] [CrossRef] [Green Version]
- Brewer, P.B.; Kaori, Y.; Fiona, F.; Emma, M.; Adrian, S.; Tancred, F.; Kohki, A.; Yoshiya, S.; Elizabeth, A.D.; Julia, E.C.; et al. Later BRANCHING OXIDOREDUCTASE acts in the final stages of strigolactone biosynthesis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2016, 113, 6301–6306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aik, W.; McDonough, M.A.; Thalhammer, A.; Chowdhury, A.; Schofield, C.J. Role of the jelly-roll fold in substrate binding by 2-oxoglutarate oxygenases. Curr. Opin. Struc. Biol. 2012, 22, 691–700. [Google Scholar] [CrossRef]
- Clifton, I.J.; McDonough, M.A.; Ehrismann, D.; Kershaw, N.J.; Granatino, N.; Schofield, C.J. Structural studies on 2-oxoglutarate oxygenases and related double-stranded beta-helix fold proteins. J. Inorg. Biochem. 2006, 100, 644–669. [Google Scholar] [CrossRef]
- Kawal, Y.; Ono, E.; Mizutani, M. Evolution and diversity of the 2–oxoglutarate-dependent dioxygenase superfamily in plants. Plant J. 2014, 78, 328–343. [Google Scholar]
- Kataoka, H.; Yamamoto, Y.; Sekiguchi, M. A new gene (alkB) of Escherichia coli that controls sensitivity to methyl methane sulfonate. J. Bacteriol. 1983, 153, 1301–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keskiaho, K.; Hieta, R.; Sormunen, R.; Myllyharju, J. Chlamydomonasreinhardtii has multiple prolyl 4–hydroxylases, one of which is essential for proper cell wall assembly. Plant Cell 2007, 19, 256–269. [Google Scholar] [CrossRef] [Green Version]
- Takehara, S.; Sakuraba, S.; Mikami, B.; Yoshida, H.; Yoshimura, H.; Itoh, A.; Endo, M.; Watanabe, N.; Nagae, T.; Matsuoka, M.; et al. A common allosteric mechanism regulates homeostatic inactivation of auxin and gibberellin. Nat. Commun. 2020, 11, 2143. [Google Scholar] [CrossRef]
- Yoo, A.; Seo, Y.S.; Jung, J.W.; Sung, S.K.; Kim, W.T.; Lee, W.; Yang, D.R. Lys296 and Arg299 residues in the C-terminus of MD-ACO1 are essential for a 1-aminocyclopropane-1-carboxylate oxidase enzyme activity. J. Struct. Biol. 2006, 156, 407–420. [Google Scholar] [CrossRef]
- Xiao, J.; Li, H.; Zhang, J.; Chen, R.; Zhang, Y.; Ouyang, B.; Wang, T.; Ye, Z. Dissection of GA 20-oxisdase members affecting tomato morphology by RNAi-mediated silencing. Plant Growth Regul. 2006, 50, 179–189. [Google Scholar] [CrossRef]
- Chen, S.; Wang, X.; Zhang, L.; Lin, S.; Liu, D.; Wang, Q.; Cai, S.; El-Tanbouly, R.; Gan, L.; Wu, H.; et al. Identification and characterization of tomato gibberellin 2-oxidases (GA2oxs) and effects of fruit-specific SlGA2ox1 overexpression on fruit and seed growth and development. Hortic. Res. 2016, 3. [Google Scholar] [CrossRef] [Green Version]
- Serrani, J.C.; Fos, M.; Atares, A.; Garcia-Martinez, J.L. Effect of gibberellin and auxin on parthenocarpic fruit growth induction in the cv Micro-Tom of tomato. J. Plant Growth Regul. 2007, 26, 211–221. [Google Scholar] [CrossRef]
- Martinez-Bello, L.; Moritz, T.; Lopez-Diaz, L. Silencing C19-GA 2-oxidases induces parthenocarpic development and inhibits lateral branching in tomato plants. Plant Physiol. 2015, 66, 5897–5910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jong, M.; Wolters-Arts, M.; Garcia-Martinez, J.L.; Mariani, C.; Vriezen, W.H. The Solanum lycopersicum AUXIN RESPONSE FACTOR 7 (SlARF7) mediates cross-talk between auxin and gibberellin signaling during tomato fruit set and development. J. Exp. Bot. 2011, 62, 617–626. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Lin, J.E.; Harris, C.; Pereira, F.C.M.; Wu, F.; Blakeslee, J.J.; Peer, A.J. DAO1 catalyzes temporal and tissue-specific oxidative inactivation of auxin in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2016, 113, 11010–11015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinozaki, Y.; Hao, S.; Kojima, M.; Sakakobara, H.; Ozeki-Lida, Y.; Zheng, Y.; Fei, Z.; Zhong, S.; Giovannoni, J.J.; Rose, J.K.; et al. Ethylene suppresses tomato (Solanum lycopersicum) fruit set through modification of gibberellin metabolism. Plant J. 2015, 83, 237–251. [Google Scholar] [CrossRef]
- Zhai, Q.; Zhang, X.; Wu, F.; Feng, H.; Deng, L.; Xu, L.; Zhang, M.; Wang, Q.; Li, C. Transcriptional mechanism of jasmonate receptor COI1-mediated delay of flowering time in Arabidopsis. Plant Cell 2015, 27, 2814–2828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomazella, D.P.; De, P.; Brail, Q.; Dahlbeck, D.; Staskawicz, B. CRISPR-Cas9 mediated mutagenesis of a DMR6 ortholog in tomato confers broad-spectrum disease resistance. bioRxiv 2016. [Google Scholar] [CrossRef] [Green Version]
- Waters, M.; Gutjahr, C.; Bennett, T.; Nelson, D. Strigolactone signaling and evolution. Annu. Rev. Plant Biol. 2017, 68, 291–322. [Google Scholar] [CrossRef]
- Nasir, F.; Tian, L.; Shi, S.; Chang, C.; Ma, L.; Gao, Y.; Tian, C. Strigolactones positively regulate defense against Magnaporthe oryzae in rice (Oryza sativa). Plant Physiol. Biochem. 2017, 142, 106–116. [Google Scholar] [CrossRef]
- Wu, H.; Li, H.; Chen, H.; Qi, Q.; Ding, Q.; Xue, J.; Ding, J.; Jiang, X.; Hou, X.; Li, Y. Identification and expression of strigolactone biosynthetic and signaling genes reveal strigolactones are involved in fruit development of the woodland strawberry (Fragaria vesca). BMC Plant Biol. 2019, 19, 73. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y. Essential roles of local auxin biosynthesis in plant development and in adaptation to environmental changes. Annu. Rev. Plant Biol. 2018, 69, 417–435. [Google Scholar] [CrossRef]
- Exposito, R.M.; Borges, A.A.; Borges-Perez, A.; Perez, J.A. Gene structure and spatiotemporal expression profile of tomato genes encoding YUCCA-like flavin monooxygenases: The ToFZY gene family. Plant Physiol. Biochem. 2011, 49, 782–791. [Google Scholar] [CrossRef]
- Epstein, E.; Cohen, J.D.; Slovin, J.P. The biosynthetic pathway for indole-3-acetic acid changes during tomato fruit development tomato fruit development. Plant Growth Regul. 2002, 38, 15–20. [Google Scholar] [CrossRef]
- Kowalczyk, M.; Sandberg, M. Quantitative analysis of indole-3-acetic acid metabolites in Arabidopsis. Plant Physiol. 2001, 127, 1845–1853. [Google Scholar] [CrossRef] [PubMed]
- Pencik, A.; Simonovik, B.; Petersson, S.V.; Henykova, E.; Simon, S.; Greenham, K.; Zhang, Y.; Kowalczyk, M.; Estelle, M.; Zazimalova, E.; et al. Regulation of auxin homeostasis and gradients in Arabidopsis roots through the formation of the indole-3-acetic acid catabolite 2-oxindole-3-acetic acid. Plant Cell 2013, 25, 3858–3870. [Google Scholar] [CrossRef] [Green Version]
- Ostin, A.; Kowalyczk, M.; Bhalerao, R.P.; Sandberg, G. Metabolism of indole-3-acetic acid in Arabidopsis. Plant Physiol. 1998, 118, 285–296. [Google Scholar] [CrossRef] [Green Version]
- Stepanova, A.N.; Alonso, J.M. Auxin catabolism unplugged: Role of IAA oxidation in auxin homeostasis. Proc. Natl. Acad. Sci. USA 2016, 113, 10742–10744. [Google Scholar] [CrossRef] [Green Version]
- JGI. Available online: http://phytozome.jgi.doe.gov/ (accessed on 30 January 2020).
- PFAM. Available online: http://pfam.xfam.org/search (accessed on 1 February 2020).
- SMART. Available online: http://smart.embl-heidelberg.de/ (accessed on 1 February 2020).
- Lee, T.H.; Tang, H.; Wang, X.; Paterson, A.H. PGDD: A database of gene and genome duplication in plants. Nucleic Acids Res. 2013, 41, 1152–1158. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40. [Google Scholar] [CrossRef] [Green Version]
- MEME. Available online: http://memesuite.org/tools/meme (accessed on 5 February 2020).
- Tomato Functional Genomics Database. Available online: http://ted.bti.cornell.edu/cgi-bin/TFGD/digital/home.cgi (accessed on 10 February 2020).






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, Q.; Wang, F.; Xue, J.; Yang, X.; Fan, J.; Chen, H.; Li, Y.; Wu, H. Identification and Expression Analysis of Hormone Biosynthetic and Metabolism Genes in the 2OGD Family for Identifying Genes That May Be Involved in Tomato Fruit Ripening. Int. J. Mol. Sci. 2020, 21, 5344. https://doi.org/10.3390/ijms21155344
Ding Q, Wang F, Xue J, Yang X, Fan J, Chen H, Li Y, Wu H. Identification and Expression Analysis of Hormone Biosynthetic and Metabolism Genes in the 2OGD Family for Identifying Genes That May Be Involved in Tomato Fruit Ripening. International Journal of Molecular Sciences. 2020; 21(15):5344. https://doi.org/10.3390/ijms21155344
Chicago/Turabian StyleDing, Qiangqiang, Feng Wang, Juan Xue, Xinxin Yang, Junmiao Fan, Hong Chen, Yi Li, and Han Wu. 2020. "Identification and Expression Analysis of Hormone Biosynthetic and Metabolism Genes in the 2OGD Family for Identifying Genes That May Be Involved in Tomato Fruit Ripening" International Journal of Molecular Sciences 21, no. 15: 5344. https://doi.org/10.3390/ijms21155344