Full Rescue of F508del-CFTR Processing and Function by CFTR Modulators Can Be Achieved by Removal of Two Regulatory Regions

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
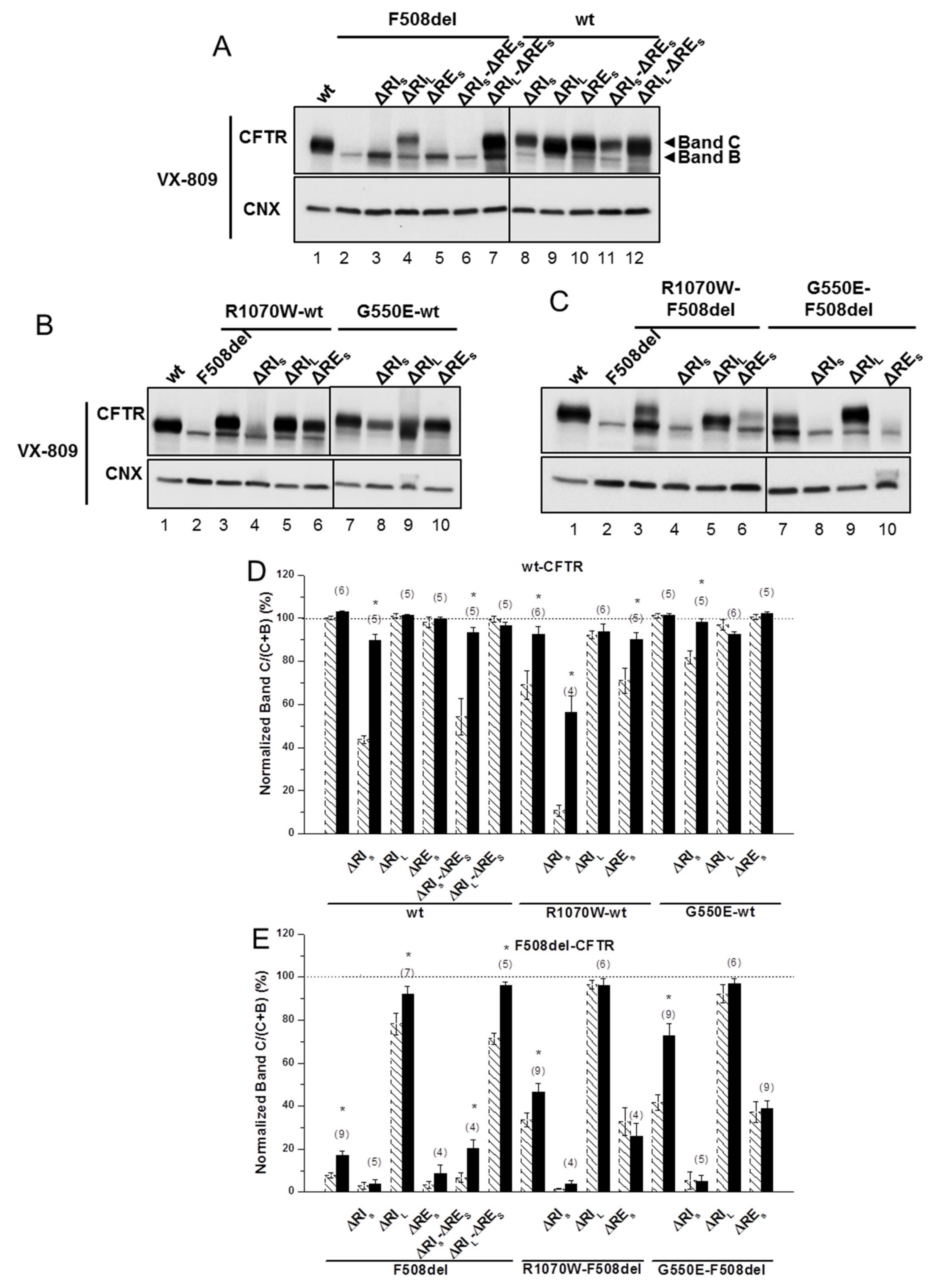
2.1. Removal of Short Regulatory Extension (RES) Alone or with Regulatory Insertion (RI) has No Impact on 508del-CFTR Processing
2.2. Simultaneous Removal of Long Regulatory Extension (REL) and Helix H9 Significantly Reduces wt-CFTR Processing but Increases Levels of F508del-CFTR Immature Form
2.3. ΔRIS, but Not ΔRES, Abolishes the Plasma Membrane Rescue of F508del-CFTR by Revertants
2.4. ΔRIL Synergises with VX-809, but Not with Revertants to Rescue ΔRES-F508del-CFTR Processing
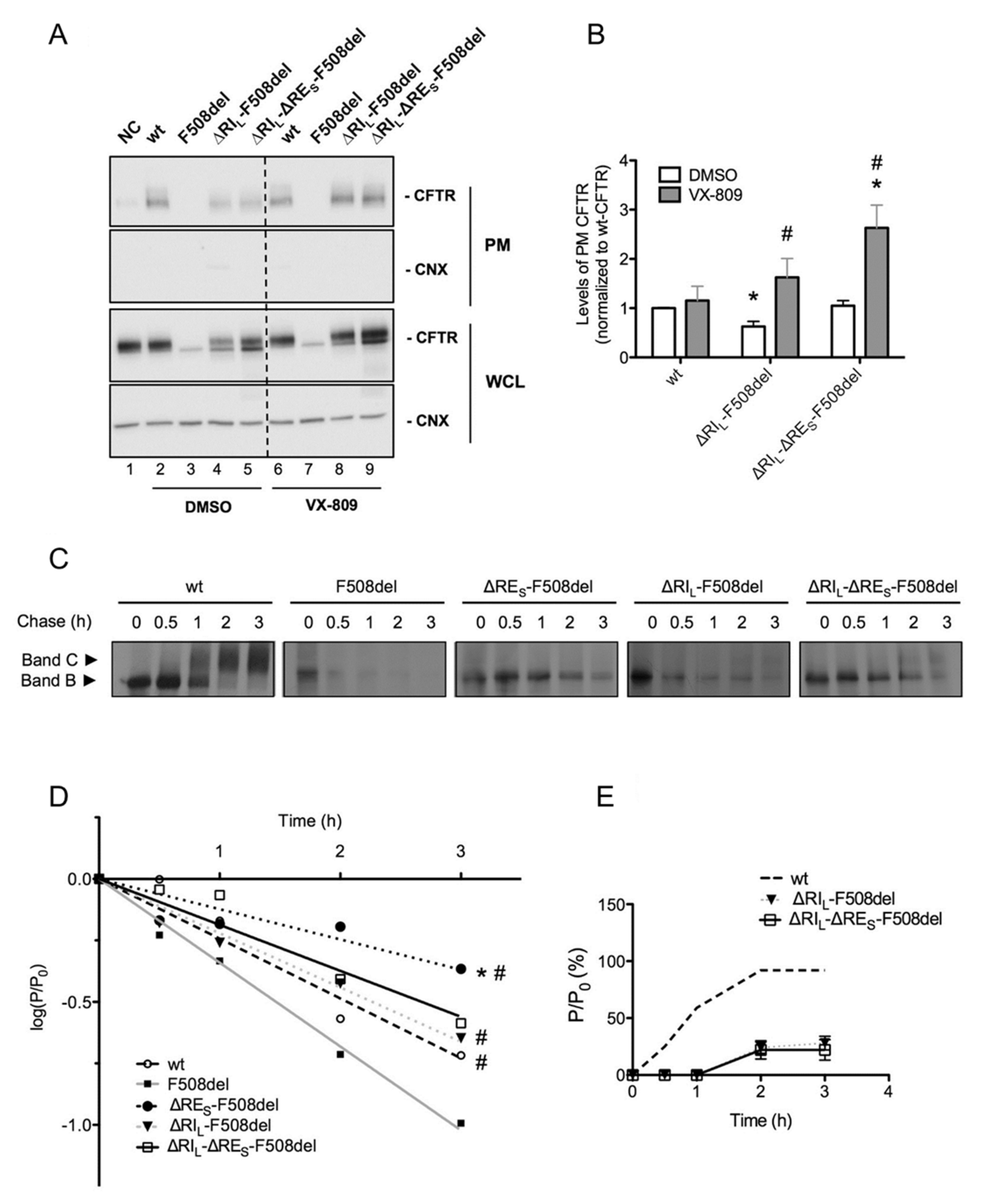
2.5. ΔRIL-ΔRES-F508del-CFTR Levels at the Plasma Membrane are Equivalent to Those of wt-CFTR
2.6. VX-809 Jointly with RE and RI Removal Completely Restored F508del-CFTR Function as Chloride Channel
2.7. VX-770-Stimulated Currents of CFTR Variants are Dramatically Decreased by ΔRIL but not by ΔRIL-ΔRES
3. Discussion
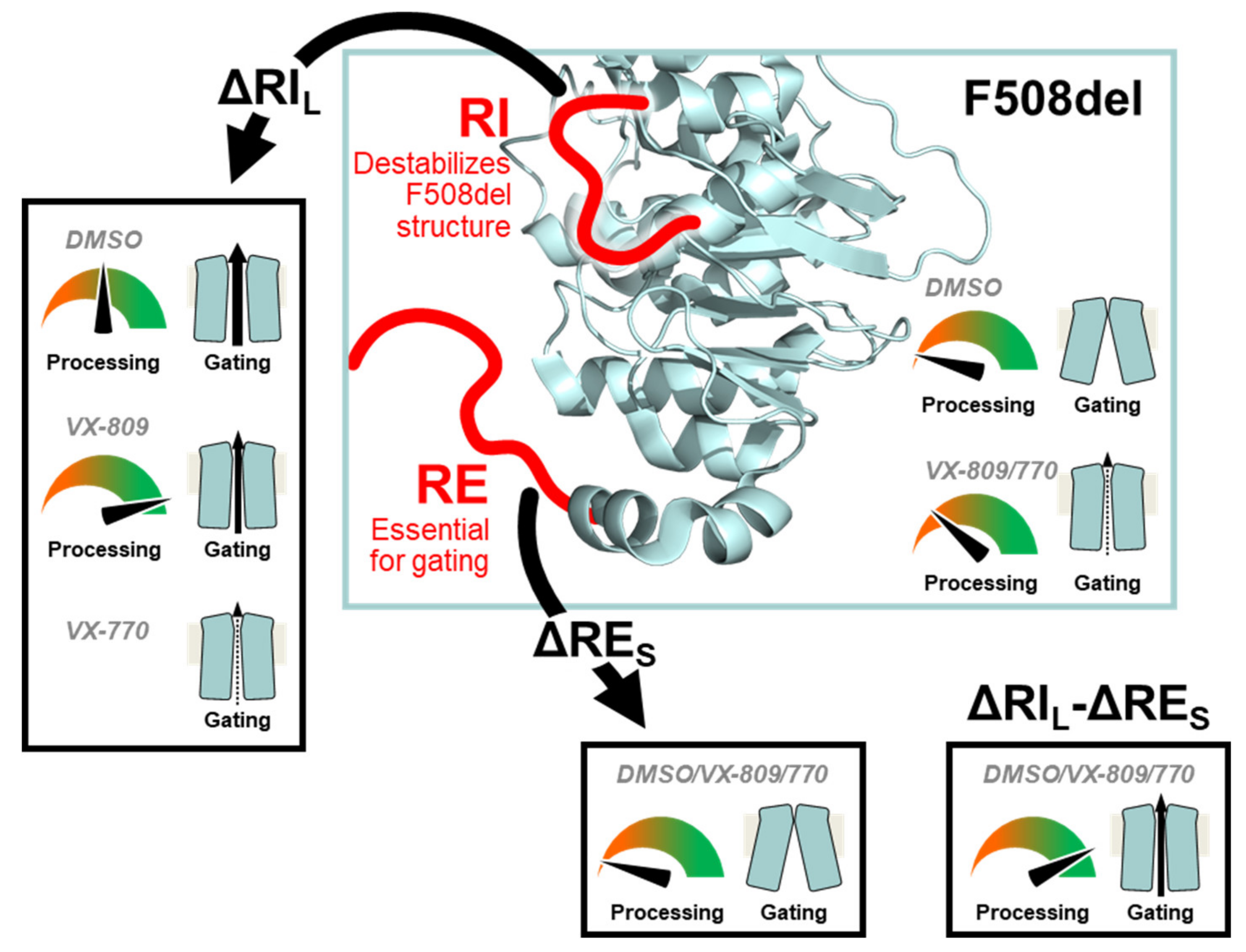
3.1. Impact of RE and RI on CFTR Processing and Function
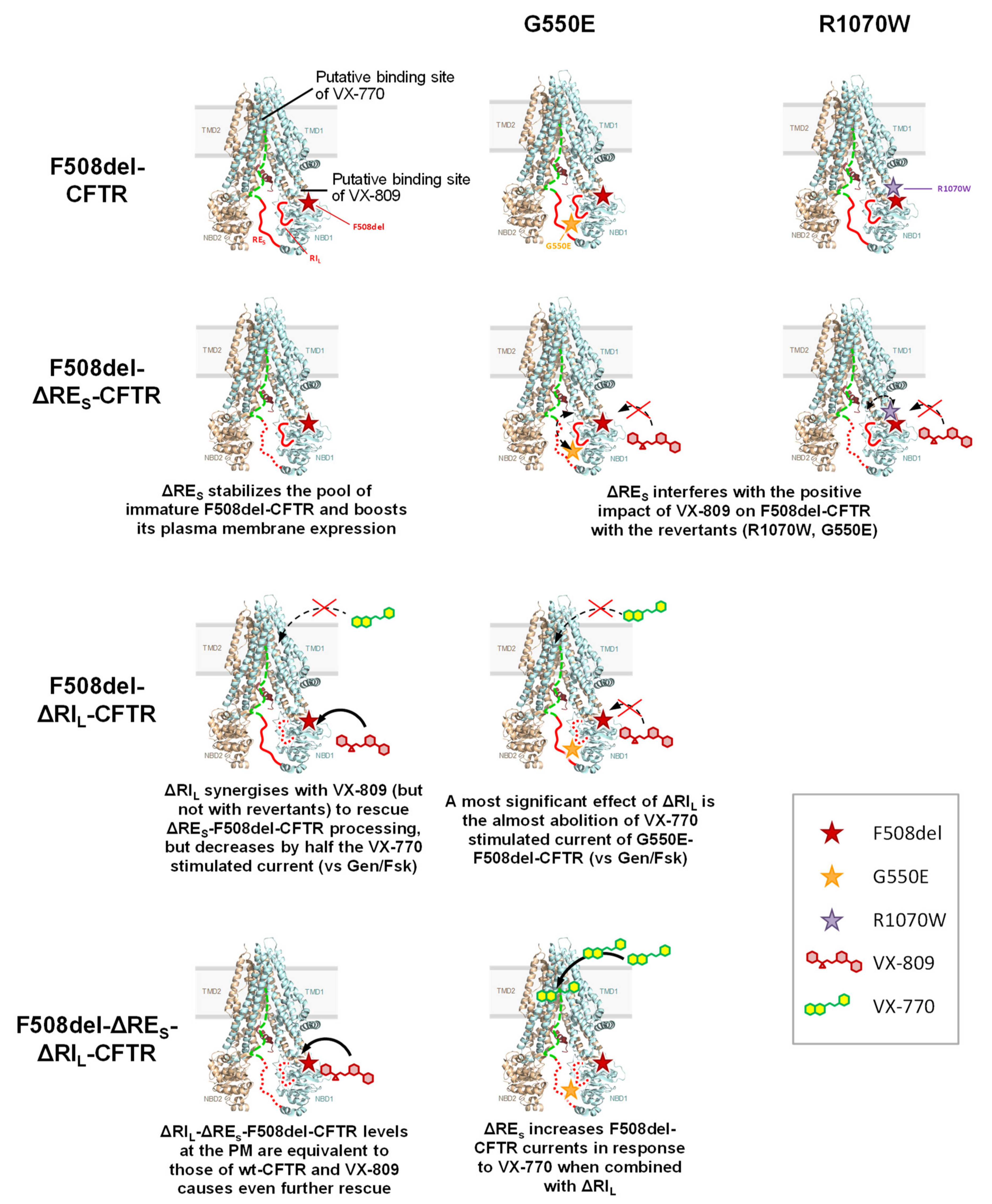
3.2. Effect of VX-809 on F508del-CFTR Variants Lacking RE and RI
3.3. Impact of F508del-Revertants on CFTR Variants Lacking RE and RI
4. Materials and Methods
4.1. CFTR Variants, Cells, and Culture Conditions
4.2. Western Blot
4.3. Pulse-Chase and Immunoprecipitation
4.4. Iodide Efflux
4.5. Biochemical Determination of the Plasma Membrane Levels of CFTR
4.6. Multiple Sequence Alignment
4.7. Data and Statistical Analyses
4.8. Reagents
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Riordan, J.R. CFTR Function and Prospects for Therapy. Annu. Rev. Biochem. 2008, 77, 701–726. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, D.N.; Welsh, M.J. Structure and Function of the CFTR Chloride Channel. Physiol. Rev. 1999, 79, S23–S45. [Google Scholar] [CrossRef] [PubMed]
- Kanelis, V.; Hudson, R.P.; Thibodeau, P.H.; Thomas, P.J.; Forman-Kay, J.D. NMR Evidence for Differential Phosphorylation-Dependent Interactions in WT and DeltaF508 CFTR. EMBO J. 2010, 29, 263–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serohijos, A.W.R.; Hegedus, T.; Aleksandrov, A.A.; He, L.; Cui, L.; Dokholyan, N.V.; Riordan, J.R. Phenylalanine-508 Mediates a Cytoplasmic-Membrane Domain Contact in the CFTR 3D Structure Crucial to Assembly and Channel Function. Proc. Natl. Acad. Sci. USA 2008, 105, 3256–3261. [Google Scholar] [CrossRef] [Green Version]
- Farinha, C.M.; King-Underwood, J.; Sousa, M.; Correia, A.R.; Henriques, B.J.; Roxo-Rosa, M.; Da Paula, A.C.; Williams, J.; Hirst, S.; Gomes, C.M.; et al. Revertants, Low Temperature, and Correctors Reveal the Mechanism of F508del-CFTR Rescue by VX-809 and Suggest Multiple Agents for Full Correction. Chem. Biol. 2013, 20, 943–955. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Aleksandrov, A.A.; Serohijos, A.W.R.; Hegedus, T.; Aleksandrov, L.A.; Cui, L.; Dokholyan, N.V.; Riordan, J.R. Multiple Membrane-Cytoplasmic Domain Contacts in the Cystic Fibrosis Transmembrane Conductance Regulator (CFTR) Mediate Regulation of Channel Gating. J. Biol. Chem. 2008, 283, 26383–26390. [Google Scholar] [CrossRef] [Green Version]
- Denning, G.M.; Anderson, M.P.; Amara, J.F.; Marshall, J.; Smith, A.E.; Welsh, M.J. Processing of Mutant Cystic Fibrosis Transmembrane Conductance Regulator Is Temperature-Sensitive. Nature 1992, 358, 761–764. [Google Scholar] [CrossRef]
- Teem, J.L.; Berger, H.A.; Ostedgaard, L.S.; Rich, D.P.; Tsui, L.C.; Welsh, M.J. Identification of Revertants for the Cystic Fibrosis Delta F508 Mutation Using STE6-CFTR Chimeras in Yeast. Cell 1993, 73, 335–346. [Google Scholar] [CrossRef]
- Teem, J.L.; Carson, M.R.; Welsh, M.J. Mutation of R555 in CFTR-Delta F508 Enhances Function and Partially Corrects Defective Processing. Recept. Channels 1996, 4, 63–72. [Google Scholar]
- DeCarvalho, A.C.V.; Gansheroff, L.J.; Teem, J.L. Mutations in the Nucleotide Binding Domain 1 Signature Motif Region Rescue Processing and Functional Defects of Cystic Fibrosis Transmembrane Conductance Regulator Delta F508. J. Biol. Chem. 2002, 277, 35896–35905. [Google Scholar] [CrossRef] [Green Version]
- Roxo-Rosa, M.; Xu, Z.; Schmidt, A.; Neto, M.; Cai, Z.; Soares, C.M.; Sheppard, D.N.; Amaral, M.D. Revertant Mutants G550E and 4RK Rescue Cystic Fibrosis Mutants in the First Nucleotide-Binding Domain of CFTR by Different Mechanisms. Proc. Natl. Acad. Sci. USA 2006, 103, 17891–17896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loo, T.W.; Bartlett, M.C.; Clarke, D.M. The V510D Suppressor Mutation Stabilizes DeltaF508-CFTR at the Cell Surface. Biochemistry 2010, 49, 6352–6357. [Google Scholar] [CrossRef]
- Aleksandrov, A.A.; Kota, P.; Aleksandrov, L.A.; He, L.; Jensen, T.; Cui, L.; Gentzsch, M.; Dokholyan, N.V.; Riordan, J.R. Regulatory Insertion Removal Restores Maturation, Stability and Function of DeltaF508 CFTR. J. Mol. Biol. 2010, 401, 194–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Goor, F.; Hadida, S.; Grootenhuis, P.D.J.; Burton, B.; Stack, J.H.; Straley, K.S.; Decker, C.J.; Miller, M.; McCartney, J.; Olson, E.R.; et al. Correction of the F508del-CFTR Protein Processing Defect in Vitro by the Investigational Drug VX-809. Proc. Natl. Acad. Sci. USA 2011, 108, 18843–18848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, H.A.; Buchanan, S.G.; Burley, S.K.; Conners, K.; Dickey, M.; Dorwart, M.; Fowler, R.; Gao, X.; Guggino, W.B.; Hendrickson, W.A.; et al. Structure of Nucleotide-Binding Domain 1 of the Cystic Fibrosis Transmembrane Conductance Regulator. EMBO J. 2004, 23, 282–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahan, D.; Evagelidis, A.; Hanrahan, J.W.; Hinkson, D.A.; Jia, Y.; Luo, J.; Zhu, T. Regulation of the CFTR Channel by Phosphorylation. Pflüg. Arch. Eur. J. Physiol. 2001, 443, S92–S96. [Google Scholar] [CrossRef]
- Pasyk, S.; Molinski, S.; Ahmadi, S.; Ramjeesingh, M.; Huan, L.-J.; Chin, S.; Du, K.; Yeger, H.; Taylor, P.; Moran, M.F.; et al. The Major Cystic Fibrosis Causing Mutation Exhibits Defective Propensity for Phosphorylation. Proteomics 2015, 15, 447–461. [Google Scholar] [CrossRef] [PubMed]
- Dawson, J.E.; Farber, P.J.; Forman-Kay, J.D. Allosteric Coupling between the Intracellular Coupling Helix 4 and Regulatory Sites of the First Nucleotide-Binding Domain of CFTR. PLoS ONE 2013, 8, e74347. [Google Scholar] [CrossRef]
- Lewis, H.A.; Zhao, X.; Wang, C.; Sauder, J.M.; Rooney, I.; Noland, B.W.; Lorimer, D.; Kearins, M.C.; Conners, K.; Condon, B.; et al. Impact of the DeltaF508 Mutation in First Nucleotide-Binding Domain of Human Cystic Fibrosis Transmembrane Conductance Regulator on Domain Folding and Structure. J. Biol. Chem. 2005, 280, 1346–1353. [Google Scholar] [CrossRef] [Green Version]
- Csanády, L.; Chan, K.W.; Nairn, A.C.; Gadsby, D.C. Functional Roles of Nonconserved Structural Segments in CFTR’s NH2-Terminal Nucleotide Binding Domain. J. Gen. Physiol. 2005, 125, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Bozoky, Z.; Krzeminski, M.; Muhandiram, R.; Birtley, J.R.; Al-Zahrani, A.; Thomas, P.J.; Frizzell, R.A.; Ford, R.C.; Forman-Kay, J.D. Regulatory R Region of the CFTR Chloride Channel Is a Dynamic Integrator of Phospho-Dependent Intra- and Intermolecular Interactions. Proc. Natl. Acad. Sci. USA 2013, 110, E4427–E4436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Zhang, Z.; Csanády, L.; Gadsby, D.C.; Chen, J. Molecular Structure of the Human CFTR Ion Channel. Cell 2017, 169, 85–95.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jih, K.-Y.; Hwang, T.-C. Vx-770 Potentiates CFTR Function by Promoting Decoupling between the Gating Cycle and ATP Hydrolysis Cycle. Proc. Natl. Acad. Sci. USA 2013, 110, 4404–4409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chappe, V.; Irvine, T.; Liao, J.; Evagelidis, A.; Hanrahan, J.W. Phosphorylation of CFTR by PKA Promotes Binding of the Regulatory Domain. EMBO J. 2005, 24, 2730–2740. [Google Scholar] [CrossRef] [Green Version]
- Mense, M.; Vergani, P.; White, D.M.; Altberg, G.; Nairn, A.C.; Gadsby, D.C. In Vivo Phosphorylation of CFTR Promotes Formation of a Nucleotide-Binding Domain Heterodimer. EMBO J. 2006, 25, 4728–4739. [Google Scholar] [CrossRef]
- Hegedus, T.; Serohijos, A.W.R.; Dokholyan, N.V.; He, L.; Riordan, J.R. Computational Studies Reveal Phosphorylation-Dependent Changes in the Unstructured R Domain of CFTR. J. Mol. Biol. 2008, 378, 1052–1063. [Google Scholar] [CrossRef] [Green Version]
- Hudson, R.P.; Chong, P.A.; Protasevich, I.I.; Vernon, R.; Noy, E.; Bihler, H.; An, J.L.; Kalid, O.; Sela-Culang, I.; Mense, M.; et al. Conformational Changes Relevant to Channel Activity and Folding within the First Nucleotide Binding Domain of the Cystic Fibrosis Transmembrane Conductance Regulator. J. Biol. Chem. 2012, 287, 28480–28494. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Liu, F.; Chen, J. Molecular Structure of the ATP-Bound, Phosphorylated Human CFTR. Proc. Natl. Acad. Sci. USA 2018, 115, 12757–12762. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Zhang, Z.; Levit, A.; Levring, J.; Touhara, K.K.; Shoichet, B.K.; Chen, J. Structural Identification of a Hotspot on CFTR for Potentiation. Science 2019, 364, 1184–1188. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, J. Atomic Structure of the Cystic Fibrosis Transmembrane Conductance Regulator. Cell 2016, 167, 1586–1597.e9. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Liu, F.; Chen, J. Conformational Changes of CFTR upon Phosphorylation and ATP Binding. Cell 2017, 170, 483–491.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aleksandrov, A.A.; Kota, P.; Cui, L.; Jensen, T.; Alekseev, A.E.; Reyes, S.; He, L.; Gentzsch, M.; Aleksandrov, L.A.; Dokholyan, N.V.; et al. Allosteric Modulation Balances Thermodynamic Stability and Restores Function of ΔF508 CFTR. J. Mol. Biol. 2012, 419, 41–60. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, B.; Callebaut, I.; Mornon, J.-P.; (Sorbonne University, Paris, France). Personal communication, 2016.
- Hudson, R.P.; Dawson, J.E.; Chong, P.A.; Yang, Z.; Millen, L.; Thomas, P.J.; Brouillette, C.G.; Forman-Kay, J.D. Direct Binding of the Corrector VX-809 to Human CFTR NBD1: Evidence of an Allosteric Coupling between the Binding Site and the NBD1:CL4 Interface. Mol. Pharmacol. 2017, 92, 124–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, H.Y.; Grove, D.E.; De La Rosa, O.; Houck, S.A.; Sopha, P.; Van Goor, F.; Hoffman, B.J.; Cyr, D.M. VX-809 Corrects Folding Defects in Cystic Fibrosis Transmembrane Conductance Regulator Protein through Action on Membrane-Spanning Domain 1. Mol. Biol. Cell 2013, 24, 3016–3024. [Google Scholar] [CrossRef] [PubMed]
- Thibodeau, P.H.; Richardson, J.M.; Wang, W.; Millen, L.; Watson, J.; Mendoza, J.L.; Du, K.; Fischman, S.; Senderowitz, H.; Lukacs, G.L.; et al. The Cystic Fibrosis-Causing Mutation DeltaF508 Affects Multiple Steps in Cystic Fibrosis Transmembrane Conductance Regulator Biogenesis. J. Biol. Chem. 2010, 285, 35825–35835. [Google Scholar] [CrossRef] [Green Version]
- Eckford, P.D.W.; Li, C.; Ramjeesingh, M.; Bear, C.E. Cystic Fibrosis Transmembrane Conductance Regulator (CFTR) Potentiator VX-770 (Ivacaftor) Opens the Defective Channel Gate of Mutant CFTR in a Phosphorylation-Dependent but ATP-Independent Manner. J. Biol. Chem. 2012, 287, 36639–36649. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Zeltwanger, S.; Yang, I.C.-H.; Nairn, A.C.; Hwang, T.-C. Actions of Genistein on Cystic Fibrosis Transmembrane Conductance Regulator Channel Gating. J. Gen. Physiol. 1998, 111, 477–490. [Google Scholar] [CrossRef] [Green Version]
- Moyer, B.D.; Loffing, J.; Schwiebert, E.M.; Loffing-Cueni, D.; Halpin, P.A.; Karlson, K.H.; Ismailov, I.I.; Guggino, W.B.; Langford, G.M.; Stanton, B.A. Membrane Trafficking of the Cystic Fibrosis Gene Product, Cystic Fibrosis Transmembrane Conductance Regulator, Tagged with Green Fluorescent Protein in Madin-Darby Canine Kidney Cells. J. Biol. Chem. 1998, 273, 21759–21768. [Google Scholar] [CrossRef] [Green Version]
- Swiatecka-Urban, A.; Duhaime, M.; Coutermarsh, B.; Karlson, K.H.; Collawn, J.; Milewski, M.; Cutting, G.R.; Guggino, W.B.; Langford, G.; Stanton, B.A. PDZ Domain Interaction Controls the Endocytic Recycling of the Cystic Fibrosis Transmembrane Conductance Regulator. J. Biol. Chem. 2002, 277, 40099–40105. [Google Scholar] [CrossRef] [Green Version]
- UniProt Consortium. UniProt: A Hub for Protein Information. Nucleic Acids Res. 2015, 43, D204–D212. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2--a Multiple Sequence Alignment Editor and Analysis Workbench. Bioinforma. Oxf. Engl. 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Notredame, C.; Higgins, D.G.; Heringa, J. T-Coffee: A Novel Method for Fast and Accurate Multiple Sequence Alignment. J. Mol. Biol. 2000, 302, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Curtis, M.J.; Bond, R.A.; Spina, D.; Ahluwalia, A.; Alexander, S.P.A.; Giembycz, M.A.; Gilchrist, A.; Hoyer, D.; Insel, P.A.; Izzo, A.A.; et al. Experimental Design and Analysis and Their Reporting: New Guidance for Publication in BJP. Br. J. Pharmacol. 2015, 172, 3461–3471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, L.; Aleksandrov, L.; Chang, X.-B.; Hou, Y.-X.; He, L.; Hegedus, T.; Gentzsch, M.; Aleksandrov, A.; Balch, W.E.; Riordan, J.R. Domain Interdependence in the Biosynthetic Assembly of CFTR. J. Mol. Biol. 2007, 365, 981–994. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| wt-Background | F508del-Background | ||||
|---|---|---|---|---|---|
| Control | VX-809 | Control | VX-809 | ||
| CFTR Variant | % to wt | % to wt | % to wt | % to wt | |
| - | 100 ± 1 | 103 ± 1 | 8 ± 1 | 17 ± 2 | |
| ΔRES | - | 98 ± 2 | 100 ± 1 | 3 ± 2 | 9 ± 4 |
| ΔRIS | 54 ± 9 | 93 ± 3 | 7 ± 2 | 20 ± 4 | |
| ΔRIL | 100 ± 1 | 97 ± 2 | 71 ± 3 | 96 ± 2 | |
| R1070W | 71 ± 6 | 90 ± 3 | 33 ± 7 | 26 ± 6 | |
| G550E | 101 ± 1 | 102 ± 1 | 37 ± 5 | 39 ± 4 | |
| ΔREL | - | 88 ± 5 | 87 ± 3 | 9 ± 2 | 8 ± 2 |
| ΔH9 | 55 ± 3 | 69 ± 3 | 0 ± 0 | 0 ± 0 | |
| ΔH9 | 72 ± 4 | 69 ± 3 | 1 ± 0 | 0 ± 0 | |
| ΔRIS | - | 44 ± 2 | 90 ± 3 | 3 ± 2 | 4 ± 3 |
| R1070W | 11 ± 3 | 56 ± 8 | 2 ± 0 | 4 ± 2 | |
| G550E | 82 ± 3 | 98 ± 1 | 6 ± 4 | 5 ± 3 | |
| ΔRIL | - | 101 ± 1 | 101 ± 0 | 78 ± 5 | 92 ± 4 |
| R1070W | 92 ± 2 | 94 ± 3 | 96 ± 2 | 96 ± 3 | |
| G550E | 97 ± 2 | 92 ± 2 | 92 ± 4 | 97 ± 2 | |
| R1070W | 69 ± 7 | 93 ± 4 | 34 ± 3 | 46 ± 4 | |
| G550E | 101 ± 1 | 101 ± 1 | 42 ± 4 | 73 ± 6 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uliyakina, I.; Botelho, H.M.; da Paula, A.C.; Afonso, S.; Lobo, M.J.; Felício, V.; Farinha, C.M.; Amaral, M.D. Full Rescue of F508del-CFTR Processing and Function by CFTR Modulators Can Be Achieved by Removal of Two Regulatory Regions. Int. J. Mol. Sci. 2020, 21, 4524. https://doi.org/10.3390/ijms21124524
Uliyakina I, Botelho HM, da Paula AC, Afonso S, Lobo MJ, Felício V, Farinha CM, Amaral MD. Full Rescue of F508del-CFTR Processing and Function by CFTR Modulators Can Be Achieved by Removal of Two Regulatory Regions. International Journal of Molecular Sciences. 2020; 21(12):4524. https://doi.org/10.3390/ijms21124524
Chicago/Turabian StyleUliyakina, Inna, Hugo M. Botelho, Ana C. da Paula, Sara Afonso, Miguel J. Lobo, Verónica Felício, Carlos M. Farinha, and Margarida D. Amaral. 2020. "Full Rescue of F508del-CFTR Processing and Function by CFTR Modulators Can Be Achieved by Removal of Two Regulatory Regions" International Journal of Molecular Sciences 21, no. 12: 4524. https://doi.org/10.3390/ijms21124524