Non-Starch Polysaccharides in Durum Wheat: A Review

 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Dietary Fibre and Nutritional Aspects
3. (1,3;1,4)-β-Glucan in Durum Wheat: Structure and Biosynthesis
4. Physicochemical and Functional Properties of (1,3;1,4)-β-Glucans
5. (1,3;1,4)-β-Glucan Content and QTL Regions in Durum Wheat
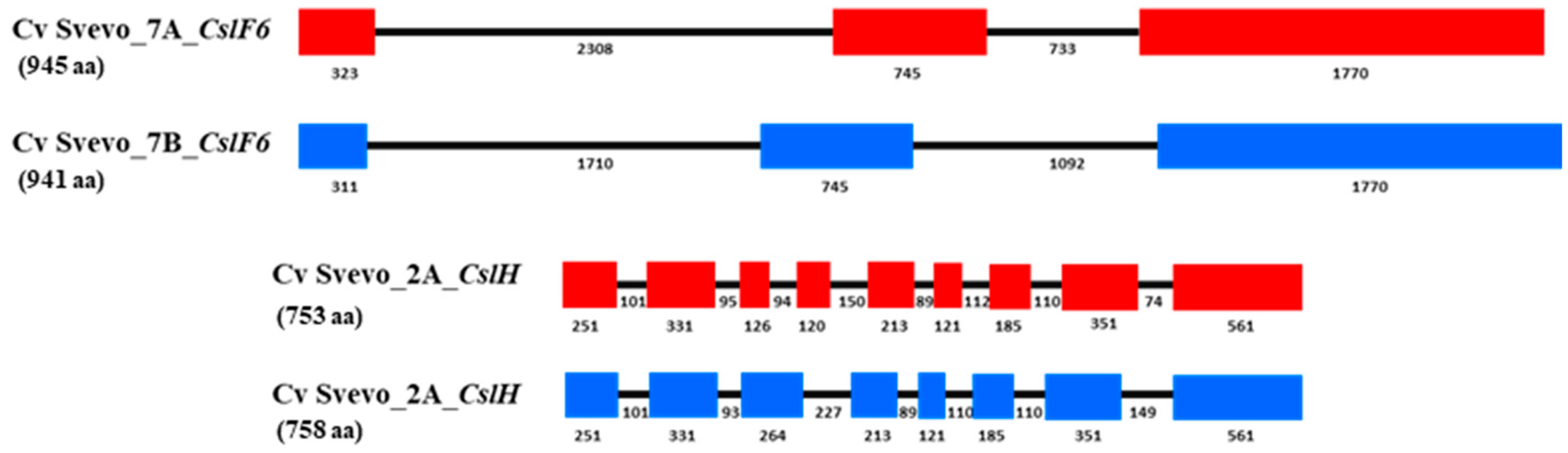
6. Candidate Genes for (1,3;1,4)-β-Glucan in Durum
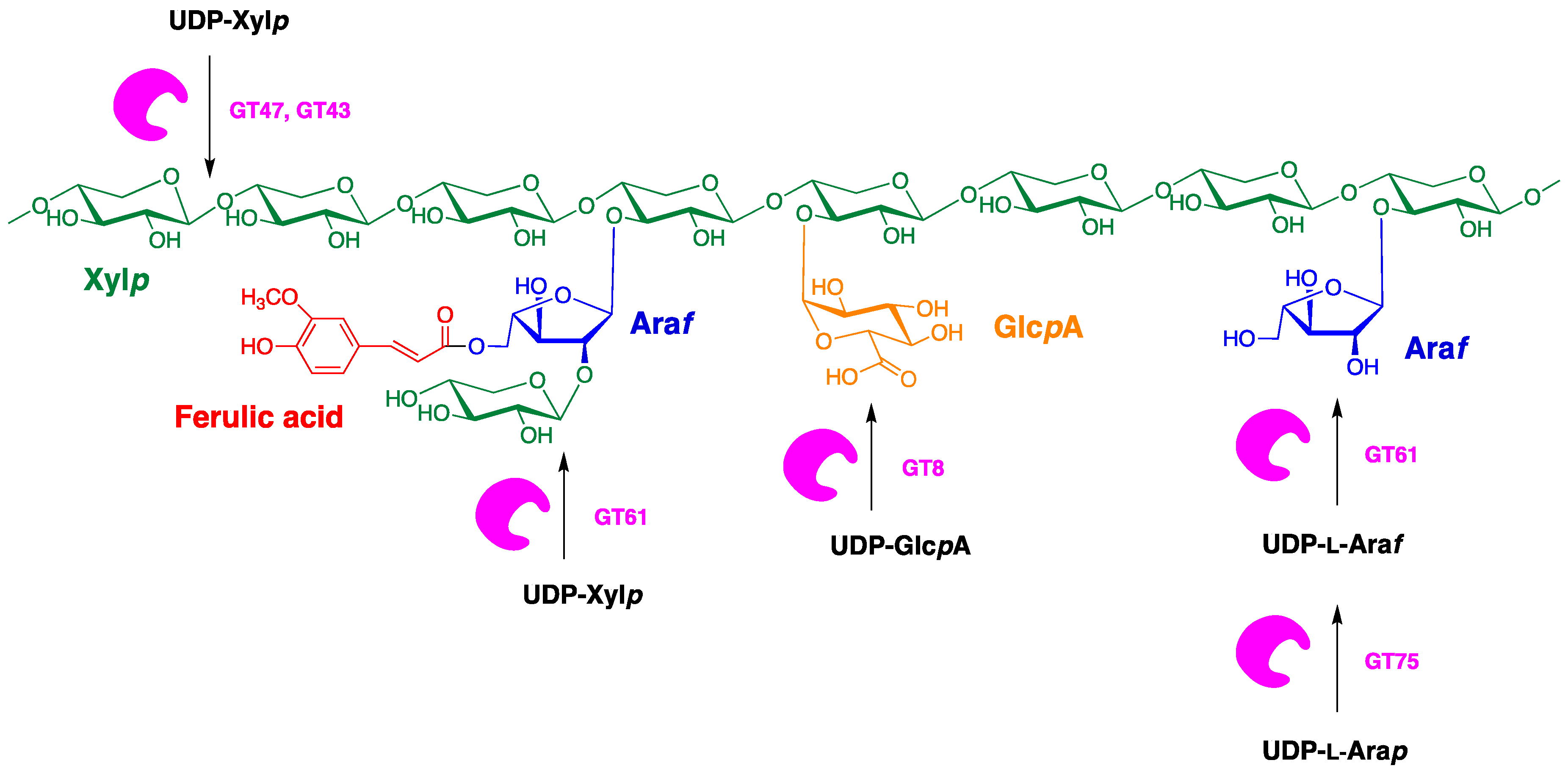
7. Arabinoxylan in Durum Kernels: Structure and Biosynthesis
8. Physio-Chemical and Functional Property of Arabinoxylan
9. Cell Walls in Plant–Pathogen Interactions
10. Arabinoxylan Content and QTL in Durum
11. Summary and Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- International Grains Council. Available online: https://www.igc.int/en/default.aspx (accessed on 13 January 2020).
- Wiese, M.V. Compendium of wheat diseases; American Phytopathological Society: St Paul, MN, USA, 1987. [Google Scholar]
- Amadou Tidiane, S.; Chiari, T.; Legesse, W.; Kemal, S.; Ortiz, R.; van Ginkel, M.; Bassi, F. Durum Wheat (Triticum durum Desf.): Origin, Cultivation and Potential Expansion in Sub-Saharan Africa. Agronomy 2019, 9, 263. [Google Scholar] [CrossRef] [Green Version]
- Colasuonno, P.; Marcotuli, I.; Blanco, A.; Maccaferri, M.; Condorelli, G.E.; Tuberosa, R.; Parada, R.; de Camargo, A.C.; Schwember, A.R.; Gadaleta, A. Carotenoid Pigment Content in Durum Wheat (Triticum turgidum L. var durum): An Overview of Quantitative Trait Loci and Candidate Genes. Front. Plant. Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Juneja, L.R.; Wilczynska, A.; Singh, R.B.; Takahashi, T.; Pella, D.; Chibisov, S.; Abramova, M.; Hristova, K.; Fedacko, J.; Pella, D.; et al. Chapter 5—Evolutionary Diet and Evolution of Man. In The Role of Functional Food Security in Global Health; Singh, R.B., Watson, R.R., Takahashi, T., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 71–85. [Google Scholar]
- Fincher, G.B. Exploring the evolution of (1,3;1,4)-beta-D-glucans in plant cell walls: Comparative genomics can help! Curr. Opin. Plant Biol. 2009, 12, 140–147. [Google Scholar] [CrossRef]
- Fedacko, J.; Takahashi, T.; Singh, R.B.; Pella, D.; Chibisov, S.; Hristova, K.; Pella, D.; Elkilany, G.N.; Tomar, R.S.; Juneja, L.R. Chapter 6—Globalization of Diets and Risk of Noncommunicable Diseases. In The Role of Functional Food Security in Global Health; Singh, R.B., Watson, R.R., Takahashi, T., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 87–107. [Google Scholar]
- Dhingra, D.; Michael, M.; Rajput, H.; Patil, R.T. Dietary fibre in foods: A review. J. Food Sci. Technol. 2012, 49, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Lambo, A.M.; Öste, R.; Nyman, M.E. Dietary fibre in fermented oat and barley β- glucan rich concentrates. Food Chem. 2005, 89, 283–293. [Google Scholar] [CrossRef]
- Bingham, S.A.; Day, N.E.; Luben, R. Dietary fibre in food and protection against colorectal cancer in the European Prospective Investigation into Cancer and Nutrition (EPIC): An observational study. Lancet 2003, 362, 1496–1501. [Google Scholar] [CrossRef]
- Ricciardi, R.; Clark, S.; Roberts, P.L. Chapter 157-Diverticular Disease Management. In Shackelford’s Surgery of the Alimentary Tract, 2 Volume Set (Eighth Edition); Yeo, C.J., Ed.; Elsevier: Philadelphia, PA, USA, 2019; pp. 1826–1847. [Google Scholar]
- Crowe, F.L.; Appleby, P.N.; Allen, N.E.; Key, T.J. Diet and risk of diverticular disease in Oxford cohort of European Prospective Investigation into Cancer and Nutrition (EPIC): Prospective study of British vegetarians and non-vegetarians. BMJ 2011, 343. [Google Scholar] [CrossRef] [Green Version]
- Bader Ul Ain, H.; Saeed, F.; Ahmad, N.; Imran, A.; Niaz, B.; Afzaal, M.; Imran, M.; Tufail, T.; Javed, A. Functional and health-endorsing properties of wheat and barley cell wall’s non-starch polysaccharides. Int. J. Food Prop. 2018, 21, 1463–1480. [Google Scholar] [CrossRef] [Green Version]
- Collins, H.M.; Burton, R.A.; Topping, D.L.; Liao, M.-L.; Bacic, A.; Fincher, G.B. Variability in Fine Structures of Noncellulosic Cell Wall Polysaccharides from Cereal Grains: Potential Importance in Human Health and Nutrition. Cereal Chem. 2010, 87, 272–282. [Google Scholar] [CrossRef]
- Zhang, D.; Williams, B.A.; Mikkelsen, D.; Li, X.; Keates, H.L.; Lisle, A.T.; Collins, H.M.; Fincher, G.B.; Bird, A.R.; Topping, D.L.; et al. Soluble arabinoxylan alters digesta flow and protein digestion of red meat-containing diets in pigs. Nutrition 2015, 31, 1141–1147. [Google Scholar] [CrossRef]
- Allison, M.A.; Criqui, M.H.; Wright, C.M. Patterns and Risk Factors for Systemic Calcified Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 331–336. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, L.R.; Harris, P.J. The dietary fibre debate: More food for thought. Lancet 2003, 361, 1487–1488. [Google Scholar] [CrossRef]
- Grabber, J.H.; Hatfield, R.D.; Ralph, J.; Zoń, J.; Amrhein, N. Ferulate cross-linking in cell walls isolated from maize cell suspensions. Phytochemistry 1995, 40, 1077–1082. [Google Scholar] [CrossRef]
- Hsieh, Y.S.; Harris, P.J. Xyloglucans of monocotyledons have diverse structures. Mol. Plant. 2009, 2, 943–965. [Google Scholar] [CrossRef] [PubMed]
- Dinelli, G.; Segura-Carretero, A.; Di Silvestro, R.; Marotti, I.; Arraez-Roman, D.; Benedettelli, S.; Ghiselli, L.; Fernadez-Gutierrez, A. Profiles of phenolic compounds in modern and old common wheat varieties determined by liquid chromatography coupled with time-of-flight mass spectrometry. J. Chromatogr. A 2011, 1218, 7670–7681. [Google Scholar] [CrossRef]
- Tan, H.T.; Corbin, K.R.; Fincher, G.B. Emerging Technologies for the Production of Renewable Liquid Transport Fuels from Biomass Sources Enriched in Plant Cell Walls. Front. Plant. Sci. 2016, 7, 1854. [Google Scholar] [CrossRef] [Green Version]
- Corbin, K.R.; Hsieh, Y.S.; Betts, N.S.; Byrt, C.S.; Henderson, M.; Stork, J.; DeBolt, S.; Fincher, G.B.; Burton, R.A. Grape marc as a source of carbohydrates for bioethanol: Chemical composition, pre-treatment and saccharification. Bioresour. Technol. 2015, 193, 76–83. [Google Scholar] [CrossRef]
- Corbin, K.R.; Byrt, C.S.; Bauer, S.; DeBolt, S.; Chambers, D.; Holtum, J.A.M.; Karem, G.; Henderson, M.; Lahnstein, J.; Beahan, C.T.; et al. Prospecting for Energy-Rich Renewable Raw Materials: Agave Leaf Case Study. PLoS ONE 2015, 10, e0135382. [Google Scholar] [CrossRef] [Green Version]
- Burton, R.A.; Gidley, M.J.; Fincher, G.B. Heterogeneity in the chemistry, structure and function of plant cell walls. Nat. Chem. Biol. 2010, 6, 724–732. [Google Scholar] [CrossRef]
- Fincher, G.B. Revolutionary times in our understanding of cell wall biosynthesis and remodeling in the grasses. Plant. Physiol. 2009, 149, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Bulone, V.; Schwerdt, J.G.; Fincher, G.B. Co-evolution of Enzymes Involved in Plant Cell Wall Metabolism in the Grasses. Front. Plant. Sci. 2019, 10, 1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcotuli, I.; Houston, K.; Waugh, R.; Fincher, G.B.; Burton, R.A.; Blanco, A.; Gadaleta, A. Genome Wide Association Mapping for Arabinoxylan Content in a Collection of Tetraploid Wheats. PLoS ONE 2015, 10, e0132787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fincher, G.B.; Stone, B.A. Chemistry of nonstarch polysaccharides. In Encyclopedia of Grain Science; Wrigley, C., Corke, H., Walker, C.E., Eds.; Elsevier: Oxford, UK, 2004; pp. 206–223. [Google Scholar]
- Woodward, J.R.; Fincher, G.B.; Stone, B. Water-soluble (1->3,1->4)-D-glucans from barley (Hordeum vulgare) endosperm. II. Fine structure. Carbohydr. Polym. 1983, 3, 207–225. [Google Scholar] [CrossRef]
- Lazaridou, A.; Biliaderis, C.G. Cryogelation of cereal β-glucans: Structure and molecular size effects. Food Hydrocolloids 2004, 18, 933–947. [Google Scholar] [CrossRef]
- Staudte, R.; Woodward, J.; Fincher, G.; Stone, B. Water-soluble (13,14)-D-glucans in barley (Hordeum vulgare) endosperm. III. Distribution of celltriosyl and cellotetraosyl residues. Carbohydr. Polym. 1983, 3, 299–312. [Google Scholar] [CrossRef]
- Woodward, J.; Fincher, G. Water soluble barley-glucans. Fine structure, solution behaviour and organization in the cell wall. Brew. Dig. 1983, 58, 28–32. [Google Scholar]
- Brennan, C.S.; Cleary, L.J. The potential use of cereal (1,3;1,4)-beta-D-glucans as functional food ingredients. J. Cereal Sci. 2005, 42, 1–13. [Google Scholar] [CrossRef]
- Doblin, M.S.; Pettolino, F.A.; Wilson, S.M.; Campbell, R.; Burton, R.A.; Fincher, G.B.; Newbigin, E.; Bacic, A. A barley cellulose synthase-like CSLH gene mediates (1,3;1,4)-beta-D-glucan synthesis in transgenic Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 5996–6001. [Google Scholar] [CrossRef] [Green Version]
- Wilson, S.M.; Ho, Y.Y.; Lampugnani, E.R.; Van de Meene, A.M.; Bain, M.P.; Bacic, A.; Doblin, M.S. Determining the subcellular location of synthesis and assembly of the cell wall polysaccharide (1,3; 1,4)-beta-D-glucan in grasses. Plant Cell 2015, 27, 754–771. [Google Scholar] [CrossRef] [Green Version]
- Trafford, K.; Haleux, P.; Henderson, M.; Parker, M.; Shirley, N.J.; Tucker, M.R.; Fincher, G.B.; Burton, R.A. Grain development in Brachypodium and other grasses: Possible interactions between cell expansion, starch deposition, and cell-wall synthesis. J. Exp. Bot. 2013, 64, 5033–5047. [Google Scholar] [CrossRef] [Green Version]
- Buckeridge, M.S.; Rayon, C.; Urbanowicz, B.; Tine, M.A.S.; Carpita, N.C. Mixed linkage (1,3;1,4)-beta-D-glucans of grasses. Cereal Chem. 2004, 81, 115–127. [Google Scholar] [CrossRef]
- Carpita, N.C.; Defernez, M.; Findlay, K.; Wells, B.; Shoue, D.A.; Catchpole, G.; Wilson, R.H.; McCann, M.C. Cell Wall Architecture of the Elongating Maize Coleoptile. Plant Physiol. 2001, 127, 551–565. [Google Scholar] [CrossRef]
- Gibeaut, D.M.; Pauly, M.; Bacic, A.; Fincher, G.B. Changes in cell wall polysaccharides in developing barley (Hordeum vulgare) coleoptiles. Planta 2005, 221, 729. [Google Scholar] [CrossRef]
- Gibeaut, D.M.; Carpita, N.C. Synthesis of (1→3), (1→4)-b-D-glucan in the Golgi apparatus of maize coleoptiles. Proc. Natl. Acad. Sci. USA 1993, 90, 3850–3854. [Google Scholar] [CrossRef] [Green Version]
- Trethewey, J.A.K.; Campbell, L.M.; Harris, P.J. (1,3;1,4)-beta-D-Glucans in the cell walls of the poales (sensu lato): An immunogold labeling study using a monoclonal antibody. Am. J. Bot. 2005, 92, 1660–1674. [Google Scholar] [CrossRef]
- Burton, R.A.; Fincher, G.B. Evolution and Development of Cell Walls in Cereal Grains. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Morrall, P.; Briggs, D. Changes in cell wall polysaccharides of germinating barley grains. Phytochemistry 1978, 17, 1495–1502. [Google Scholar] [CrossRef]
- Guillon, F.; Larre, C.; Petipas, F.; Berger, A.; Moussawi, J.; Rogniaux, H.; Santoni, A.; Saulnier, L.; Jamme, F.; Miquel, M.; et al. A comprehensive overview of grain development in Brachypodium distachyon variety Bd21. J. Exp. Bot. 2012, 63, 739–755. [Google Scholar] [CrossRef] [Green Version]
- McCleary, B.V.; Codd, R. Measurement of (1-3),(1-4)-β-D-glucan in barley and oats: A streamlined enzymatic procedure. J. Sci. Food Agric. 1991, 55, 303–312. [Google Scholar] [CrossRef]
- Pritchard, J.R.; Lawrence, G.J.; Larroque, O.; Li, Z.; Laidlaw, H.K.C.; Morell, M.K.; Rahman, S. A survey of β-glucan and arabinoxylan content in wheat. J. Sci. Food Agric. 2011, 91, 1298–1303. [Google Scholar] [CrossRef]
- Marcotuli, I.; Houston, K.; Schwerdt, J.G.; Waugh, R.; Fincher, G.B.; Burton, R.A.; Blanco, A.; Gadaleta, A. Genetic Diversity and Genome Wide Association Study of beta-Glucan Content in Tetraploid Wheat Grains. PLoS ONE 2016, 11, e0152590. [Google Scholar] [CrossRef] [Green Version]
- Marcotuli, I.; Gadaleta, A.; Mangini, G.; Signorile, A.M.; Zacheo, S.A.; Blanco, A.; Simeone, R.; Colasuonno, P. Development of a High-Density SNP-Based Linkage Map and Detection of QTL for beta-Glucans, Protein Content, Grain Yield per Spike and Heading Time in Durum Wheat. Int. J. Mol. Sci. 2017, 18, 1329. [Google Scholar] [CrossRef] [PubMed]
- Marcotuli, I.; Colasuonno, P.; Cutillo, S.; Simeone, R.; Blanco, A.; Gadaleta, A. β-glucan content in a panel of Triticum and Aegilops genotypes. Genet. Resour. Crop Ev. 2019, 66, 897–907. [Google Scholar] [CrossRef]
- Colasuonno, P.; Marcotuli, I.; Cutillo, S.; Simeone, R.; Blanco, A.; Gadaleta, A. Effect of barley chromosomes on the β-glucan content of wheat. Genet. Resour. Crop Ev. 2020, 67, 561–567. [Google Scholar] [CrossRef]
- Danilova, T.V.; Akhunova, A.R.; Akhunov, E.D.; Friebe, B.; Gill, B.S. Major structural genomic alterations can be associated with hybrid speciation in Aegilops markgrafii (Triticeae). Plant. J. 2017, 92, 317–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakszegi, M.; Molnar, I.; Lovegrove, A.; Darko, E.; Farkas, A.; Lang, L.; Bedo, Z.; Dolezel, J.; Molnar-Lang, M.; Shewry, P. Addition of Aegilops U and M Chromosomes Affects Protein and Dietary Fiber Content of Wholemeal Wheat Flour. Front. Plant. Sci. 2017, 8, 1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Türkösi, E.; Cseh, A.; Darkó, É.; Molnár-Láng, M. Addition of Manas barley chromosome arms to the hexaploid wheat genome. BMC Genet. 2016, 17, 87. [Google Scholar] [CrossRef] [Green Version]
- Cseh, A.; Soós, V.; Rakszegi, M.; Türkösi, E.; Balázs, E.; Molnár-Láng, M. Expression ofHvCslF9andHvCslF6barley genes in the genetic background of wheat and their influence on the wheat β-glucan content. Ann. Appl. Biol. 2013, 163, 142–150. [Google Scholar] [CrossRef]
- Molnár-Láng, M.; Kruppa, K.; Cseh, A.; Bucsi, J.; Linc, G. Identification and phenotypic description of new wheat-six-rowed winter barley disomic additions. Genome 2012, 55, 302–311. [Google Scholar] [CrossRef]
- Manickavelu, A.; Kawaura, K.; Imamura, H.; Mori, M.; Ogihara, Y. Molecular mapping of quantitative trait loci for domestication traits and β-glucan content in a wheat recombinant inbred line population. Euphytica 2010, 177, 179–190. [Google Scholar] [CrossRef]
- Maccaferri, M.; Ricci, A.; Salvi, S.; Milner, S.G.; Noli, E.; Martelli, P.L.; Casadio, R.; Akhunov, E.; Scalabrin, S.; Vendramin, V.; et al. A high-density, SNP-based consensus map of tetraploid wheat as a bridge to integrate durum and bread wheat genomics and breeding. Plant. Biotechnol. J. 2015, 13, 648–663. [Google Scholar] [CrossRef]
- Little, A.; Schwerdt, J.G.; Shirley, N.J.; Khor, S.F.; Neumann, K.; O’Donovan, L.A.; Lahnstein, J.; Collins, H.M.; Henderson, M.; Fincher, G.B.; et al. Revised Phylogeny of the Cellulose Synthase Gene Superfamily: Insights into Cell Wall Evolution. Plant. Physiol. 2018, 177, 1124–1141. [Google Scholar] [CrossRef] [Green Version]
- Pear, J.R.; Kawagoe, Y.; Schreckengost, W.E.; Delmer, D.P.; Stalker, D.M. Higher plants contain homologs of the bacterial Cela genes encoding the catalytic subunit of cellulose synthase. Proc. Natl. Acad. Sci. USA 1996, 93, 12637–12642. [Google Scholar] [CrossRef] [Green Version]
- Arioli, T.; Liangcai, P.; Andreas, S.B.; Joanne, B.; Werner, W.; Werner, H.; Christine, C.; Herman, H.; Jacek, P.; Rosemary, B.; et al. Molecular Analysis of Cellulose Biosynthesis in Arabidopsis. Science 1998, 279, 717. [Google Scholar] [CrossRef] [Green Version]
- Burton, R.A.; Gibeaut, D.M.; Bacic, A.; Findlay, K.; Roberts, K.; Hamilton, A.; Baulcombe, D.C.; Fincher, G.B. Virus-induced silencing of a plant cellulose synthase gene. Plant Cell 2000, 12, 691–705. [Google Scholar] [CrossRef] [Green Version]
- Scheible, W.R.; Eshed, R.; Richmond, T.; Delmer, D.; Somerville, C. Modifications of cellulose synthase confer resistance to isoxaben and thiazolidinone herbicides in Arabidopsis Ixr1 mutants. Proc. Natl. Acad. Sci. USA 2001, 98, 10079–10084. [Google Scholar] [CrossRef] [Green Version]
- Houston, K.; Burton, R.A.; Sznajder, B.; Rafalski, A.J.; Dhugga, K.S.; Mather, D.E.; Taylor, J.; Steffenson, B.J.; Waugh, R.; Fincher, G.B. A Genome-Wide Association Study for Culm Cellulose Content in Barley Reveals Candidate Genes Co-Expressed with Members of the CELLULOSE SYNTHASE A Gene Family. PLoS ONE 2015, 10, e0130890. [Google Scholar] [CrossRef] [Green Version]
- Burton, R.A.; Wilson, S.M.; Hrmova, M.; Harvey, A.J.; Shirley, N.J.; Medhurst, A.; Stone, B.A.; Newbigin, E.J.; Bacic, A.; Fincher, G.B. Cellulose synthase-like CslF genes mediate the synthesis of cell wall (1,3;1,4)-ß-D-glucans. Science 2006, 311, 1940–1942. [Google Scholar] [CrossRef]
- Schwerdt, J.G.; MacKenzie, K.; Wright, F.; Oehme, D.; Wagner, J.M.; Harvey, A.J.; Shirley, N.J.; Burton, R.A.M.; Halpin, C.; Zimmer, J. Evolutionary Dynamics of the Cellulose Synthase Gene Superfamily in Grasses. Plant. Physiol. 2015, 168, 968–983. [Google Scholar] [CrossRef] [Green Version]
- Richmond, T.; Somerville, C. The cellulose synthase superfamily. Plant. Physiol. 2000, 124, 495–498. [Google Scholar] [CrossRef] [Green Version]
- Farrokhi, N.; Burton, R.; Brownfield, L.; Hrmova, M.; Wilson, S.; Bacic, A.; Fincher, G. Plant cell wall biosynthesis: Genetic, biochemical and functional genomics approaches to the identification of key genes. Plant. Biotechnol. J. 2006, 4, 145–167. [Google Scholar] [CrossRef]
- Burton, R.A.; Collins, H.M.; Kibble, N.A.J.; Smith, J.A.; Shirley, N.J.; Jobling, S.A.; Henderson, M.; Singh, R.R.; Pettolino, F.; Wilson, S.M.; et al. Over-expression of specific HvCslF cellulose synthase-like genes in transgenic barley increases the levels of cell wall (1,3;1,4)-ß-D-glucans and alters their fine structure. Plant. Biotechnol. J. 2011, 9, 117–135. [Google Scholar] [CrossRef]
- Houston, K.; Russell, J.; Schreiber, M.; Halpin, C.; Oakey, H.; Washington, J.; Booth, A.; Shirley, N.; Burton, R.; Fincher, G.; et al. A genome wide association scan for (1,3;1,4)-beta-glucan content in the grain of contemporary 2-row Spring and Winter barleys. BMC Genom. 2014, 15, 907. [Google Scholar] [CrossRef] [Green Version]
- Cu, S.T.; March, T.J.; Stewart, S.; Degner, S.; Coventry, S.; Box, A.; Stewart, D.; Skadhauge, B.; Burton, R.A.; Fincher, G.B.; et al. Genetic analysis of grain and malt quality in an elite barley population. Mol. Breed. 2016, 36. [Google Scholar] [CrossRef]
- Cu, S.; Collins, H.M.; Betts, N.S.; March, T.J.; Janusz, A.; Stewart, D.C.; Skadhauge, B.; Eglinton, J.; Kyriacou, B.; Little, A.; et al. Water uptake in barley grain: Physiology; genetics and industrial applications. Plant. Sci. 2016, 242, 260–269. [Google Scholar] [CrossRef]
- Burton, R.; Jobling, S.; Harvey, A.; Shirley, N.; Mather, D.; Bacic, A.; Fincher, G. The Genetics and Transcriptional Profiles of the Cellulose Synthase-Like HvCslF Gene Family in Barley. Plant. Physiol. 2008, 146, 1821–1833. [Google Scholar] [CrossRef] [Green Version]
- Marcotuli, I.; Colasuonno, P.; Blanco, A.; Gadaleta, A. Expression analysis of cellulose synthase-like genes in durum wheat. Sci. Rep. 2018, 8, 15675. [Google Scholar] [CrossRef]
- Dimitroff, G.; Little, A.; Lahnstein, J.; Schwerdt, J.G.; Srivastava, V.; Bulone, V.; Burton, R.A.; Fincher, G.B. (1,3;1,4)-beta-Glucan Biosynthesis by the CSLF6 Enzyme: Position and Flexibility of Catalytic Residues Influence Product Fine Structure. Biochemistry 2016, 55, 2054–2061. [Google Scholar] [CrossRef]
- Andersson, R.; Westerlund, E.; Aman, P. Natural variations in the contents of structural elements of water-extractable non-starch polysaccharides in white flour. J. Cereal Sci. 1994, 19, 77–82. [Google Scholar] [CrossRef]
- Nieduszynski, I.; Marchessault, R.H. Structure of beta-D-(1,4′)-xylan hydrate. Nature 1971, 232, 46–47. [Google Scholar] [CrossRef]
- Courtin, C.M.; Gys, W.; Gebruers, K.; Delcour, J.A. Evidence for the Involvement of Arabinoxylan and Xylanases in Refrigerated Dough Syruping. J. Agric. Food Chem. 2005, 53, 7623–7629. [Google Scholar] [CrossRef]
- Smith, M.M.; Hartley, R.D. Occurrence and nature of ferulic acid substitution of cell-wall polysaccharides in graminaceous plants. Carbohydr. Res. 1983, 118, 65–80. [Google Scholar] [CrossRef]
- Stone, B.A.; Morell, M. Carbohydrates. In Wheat: Chemistry and Technology; Khan, K., Shewry, P., Eds.; AACC International Inc.: St Paul, MN, USA, 2007; Volume 4. [Google Scholar]
- Busse-Wicher, M.; Li, A.; Silveira, R.L.; Pereira, C.S.; Tryfona, T.; Gomes, T.C.F.; Skaf, M.S.; Dupree, P. Evolution of Xylan Substitution Patterns in Gymnosperms and Angiosperms: Implications for Xylan Interaction with Cellulose. Plant. Physiol. 2016, 171, 2418–2431. [Google Scholar] [CrossRef]
- Viëtor, R.; Angelino, S.; Voragen, A. Structural features of arabinoxylans from barley and malt cell wall material. J. Cereal Sci 1992, 15, 213–222. [Google Scholar] [CrossRef]
- Andersson, R.; Westerlund, E.; Tilly, A.C.; Aman, P. Natural variations in the chemical composition of white flour. J. Cereal Sci. 1993, 17, 183–189. [Google Scholar] [CrossRef]
- Lee, J.N.; Lee, D.Y.; Ji, I.H.; Kim, G.E.; Kim, H.N.; Sohn, J.; Kim, S.; Kim, C.W. Purification of soluble beta-glucan with immune-enhancing activity from the cell wall of yeast. Biosci. Biotechnol. Biochem. 2001, 65, 837–841. [Google Scholar] [CrossRef] [Green Version]
- Carpita, N.C. Cell wall development in maize coleoptiles. Plant. Physiol. 1984, 76, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Iiyama, K.; Lam, T.B.T.; Stone, B.A. Covalent cross-links in the cell-wall. Plant. Physiol. 1994, 104, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Waldron, K.W.; Brett, C.T. A glucuronyltransferase involved in glucuronoxylan synthesis in pea (Pisum sativum) epicotyls. Biochem. J. 1983, 213, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.M.; Goubet, F.; Vicky, W.W.A.; Goodacre, R.; Stephens, E.; Dupree, P.; Turner, S.R. Comparison of five xylan synthesis mutants reveals new insight into the mechanisms of xylan synthesis. Plant. J. 2007, 52, 1154–1168. [Google Scholar] [CrossRef]
- Mitchell, R.A.C.; Dupree, P.; Shewry, P.R. A novel bioinformatics approach identifies candidate genes for the synthesis and feruloylation of arabinoxylan. Plant. Physiol. 2007, 144, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Pena, M.J.; Zhong, R.; Zhou, G.-K.; Richardson, E.A.; O’Neill, M.A.; Darvill, A.G.; York, W.S.; Ye, Z.-H. Arabidopsis irregular xylem8 and irregular xylem9: Implications for the Complexity of Glucuronoxylan Biosynthesis. Plant. Cell 2007, 19, 549–563. [Google Scholar] [CrossRef] [Green Version]
- Persson, S.; Paredez, A.; Carroll, A.; Palsdottir, H.; Doblin, M.; Poindexter, P.; Khitrov, N.; Auer, M.; Somerville, C.R. Genetic evidence for three unique components in primary cell-wall cellulose synthase complexes in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 15566–15571. [Google Scholar] [CrossRef] [Green Version]
- Pellny, T.K.; Lovegrove, A.; Freeman, J.; Tosi, P.; Love, C.G.; Knox, J.P.; Shewry, P.R.; Mitchell, R.A. Cell walls of developing wheat starchy endosperm: Comparison of composition and RNA-Seq transcriptome. Plant. Physiol. 2012, 158, 612–627. [Google Scholar] [CrossRef] [Green Version]
- Anders, N.; Wilkinson, M.D.; Lovegrove, A.; Freeman, J.; Tryfona, T.; Pellny, T.K.; Weimar, T.; Mortimer, J.C.; Stott, K.; Baker, J.M.; et al. Glycosyl transferases in family 61 mediate arabinofuranosyl transfer onto xylan in grasses. Proc. Natl. Acad. Sci. USA 2012, 109, 989–993. [Google Scholar] [CrossRef] [Green Version]
- Lovegrove, A.; Wilkinson, M.D.; Freeman, J.; Pellny, T.K.; Tosi, P.; Saulnier, L.; Shewry, P.R.; Mitchell, R.A.C. RNA Interference Suppression of Genes in Glycosyl Transferase Families 43 and 47 in Wheat Starchy Endosperm Causes Large Decreases in Arabinoxylan Content. Plant. Physiol. 2013, 163, 95–107. [Google Scholar] [CrossRef] [Green Version]
- Jensen, J.K.; Johnson, N.R.; Wilkerson, C.G. Arabidopsis thaliana IRX10 and two related proteins from psyllium and Physcomitrella patens are xylan xylosyltransferases. Plant. J. 2014, 80, 207–215. [Google Scholar] [CrossRef]
- Urbanowicz, B.R.; Peña, M.J.; Moniz, H.A.; Moremen, K.W.; York, W.S. Two Arabidopsis proteins synthesize acetylated xylan in vitro. Plant. J. 2014, 80, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Gruppen, H.; Kormelink, F.J.M.; Voragen, A.G.J. Enzymatic Degradation of Water-Unextractable Cell-Wall Material and Arabinoxylans from Wheat-Flour. J. Cereal Sci. 1993, 18, 129–143. [Google Scholar] [CrossRef]
- Izydorczyk, M.S.; Biliaderis, C.G. Studies on the structure of wheat endosperm arabinoxylans. Carbohydr. Polym. 1994, 24, 61–71. [Google Scholar] [CrossRef]
- Guillon, F.; Bouchet, B.; Jamme, F.; Robert, P.; Quemener, B.; Barron, C.; Larre, C.; Dumas, P.; Saulnier, L. Brachypodium distachyon grain: Characterization of endosperm cell walls. J. Exp. Bot. 2011, 62, 1001–1015. [Google Scholar] [CrossRef] [Green Version]
- Marcotuli, I.; Hsieh, Y.S.; Lahnstein, J.; Yap, K.; Burton, R.A.; Blanco, A.; Fincher, G.B.; Gadaleta, A. Structural Variation and Content of Arabinoxylans in Endosperm and Bran of Durum Wheat (Triticum turgidum L.). J. Agric. Food Chem. 2016, 64, 2883–2892. [Google Scholar] [CrossRef]
- Andrewartha, K.A.; Phillips, D.R.; Stone, B.A. Solution properties of wheat-flour arabinoxylans and enzymatically modified arabinoxylans. Carbohydr. Res. 1979, 77, 191. [Google Scholar] [CrossRef]
- Mendis, M.; Simsek, S. Arabinoxylans and human health. Food Hydrocoll. 2014, 42, 239–243. [Google Scholar] [CrossRef]
- do Prado, S.B.R.; Castro-Alves, V.C.; Ferreira, G.F.; Fabi, J.P. Ingestion of Non-digestible Carbohydrates From Plant-Source Foods and Decreased Risk of Colorectal Cancer: A Review on the Biological Effects and the Mechanisms of Action. Front. Nutr. 2019, 6, 72. [Google Scholar] [CrossRef]
- Garcia-Angulo, P.; Willats, W.G.T.; Encina, A.E.; Alonso-Simon, A.; Alvarez, J.M.; Acebes, J.L. Immunocytochemical characterization of the cell walls of bean cell suspensions during habituation and dehabituation to dichlobenil. Physiol. Plant. 2006, 127, 87–99. [Google Scholar] [CrossRef]
- Mendis, M.; Leclerc, E.; Simsek, S. Arabinoxylans, gut microbiota and immunity. Carbohydr. Polym. 2016, 139, 159–166. [Google Scholar] [CrossRef]
- Schneeman, B.O. Gastrointestinal physiology and functions. British J. Nutr. 2002, 88, S159–S163. [Google Scholar] [CrossRef] [Green Version]
- Courtin, C.M.; Delcour, J.A. Arabinoxylans and endoxylanases in wheat flour bread-making. J. Cereal Sci. 2002, 35, 225–243. [Google Scholar] [CrossRef]
- Frederix, S.A.; Van hoeymissen, K.E.; Courtin, C.M.; Delcour, J.A. Water-Extractable and Water-Unextractable Arabinoxylans Affect Gluten Agglomeration Behavior during Wheat Flour Gluten Starch Separation. J. Agric. Food Chem. 2004, 52, 7950–7956. [Google Scholar] [CrossRef]
- Simsek, S.; Ohm, J.B. Structural changes of arabinoxylans in refrigerated dough. Carbohydr. Polym. 2009, 77, 87–94. [Google Scholar] [CrossRef]
- Bedford, M.R.; Schulze, H. Exogenous enzymes for pigs and poultry. Nutr. Res. Rev. 1998, 11, 91–114. [Google Scholar] [CrossRef]
- Ryals, J.A.; Neuenschwander, U.H.; Willits, M.G.; Molina, A.; Steiner, H.Y.; Hunt, M.D. Systemic acquired resistance. Plant. Cell 1996, 8, 1809–1819. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, J.; Henderson, M.; Schweizer, P.; Burton, R.A.; Fincher, G.B.; Little, A. Differential accumulation of callose, arabinoxylan and cellulose in nonpenetrated versus penetrated papillae on leaves of barley infected with Blumeria graminis f. sp. hordei. New Phytol. 2014, 204, 650–660. [Google Scholar] [CrossRef]
- Chowdhury, J.; Lück, S.; Rajaraman, J.; Douchkov, D.; Shirley, N.J.; Schwerdt, J.G.; Schweizer, P.; Fincher, G.B.; Burton, R.A.; Little, A. Altered Expression of Genes Implicated in Xylan Biosynthesis Affects Penetration Resistance against Powdery Mildew. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Giancaspro, A.; Lionetti, V.; Giove, S.L.; Zito, D.; Fabri, E.; Reem, N.; Zabotina, O.A.; De Angelis, E.; Monaci, L.; Bellincampi, D.; et al. Cell wall features transferred from common into durum wheat to improve Fusarium Head Blight resistance. Plant. Sci. 2018, 274, 121–128. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
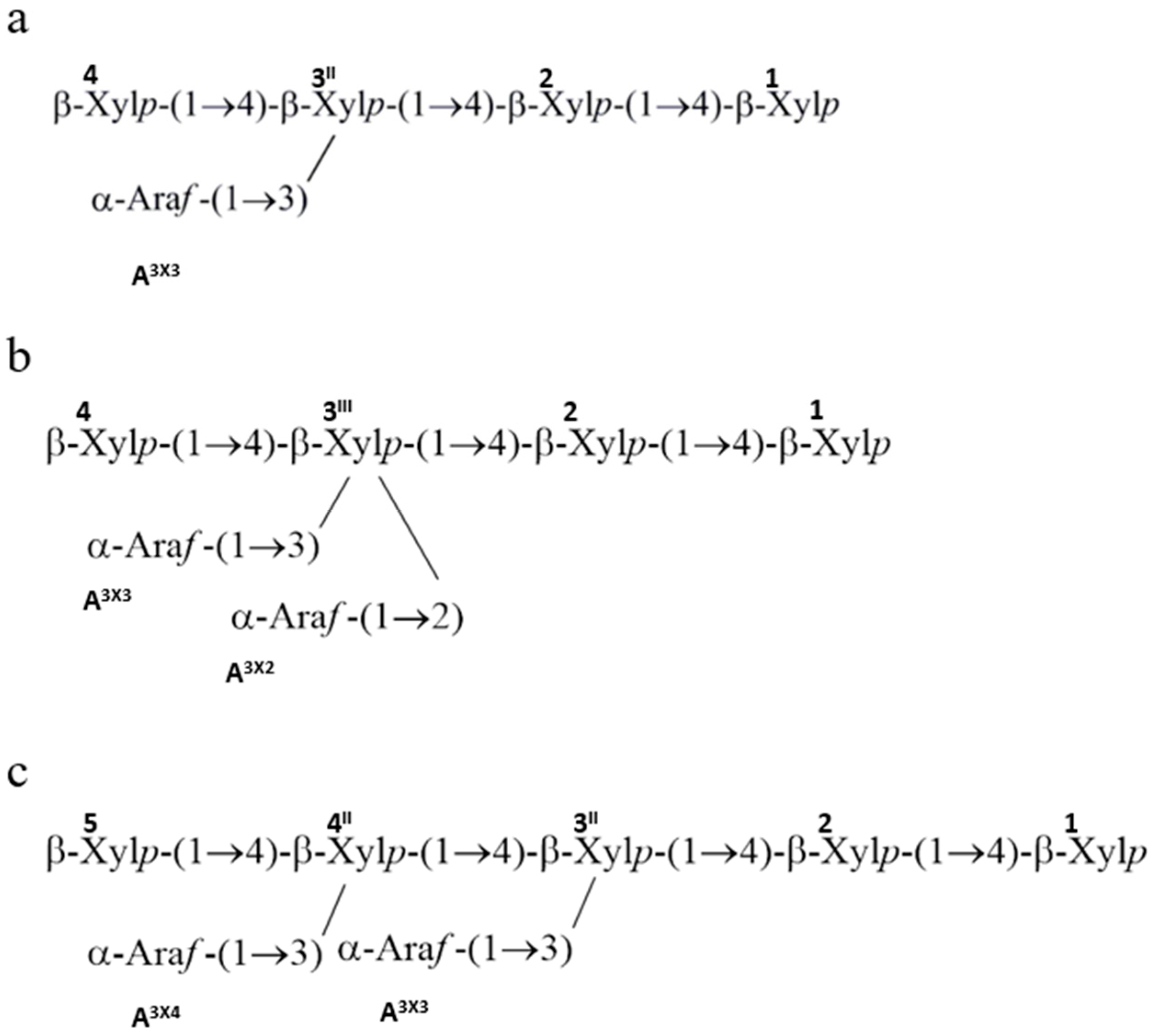{kind=link}
| Nutrients | Triticum Turgidum Durum Wheat | Triticum Aestivum Bread Wheat | Hordeum Vulgare Barley | Avena Sativa Oats | Oryza Sativa Rice |
|---|---|---|---|---|---|
| Starch (%DM) | 71 | 48 | 55 | 44 | 64 |
| Protein (g) | 14 | 12 | 13 | 10 | 4.4 |
| Fat (g) | 2.5 | 1.7 | 1.6 | 1.6 | 0.4 |
| Dietary fibre (g) | 1.6 | 2.4 | 10 | 15 | 0.6 |
| Thiamin B1 (mg) | 0.4 | 0.1 | 0.2 | 0.7 | 0.1 |
| Riboflavin B2 (mg) | 0.1 | 0.0 | 0.1 | 0.1 | 0.0 |
| Sodium (mg) | 3.8 | 2.0 | 4.0 | 2.0 | 2.0 |
| Potassium (mg) | 431 | 141 | 309 | 510 | 86 |
| Calcium (mg) | 34 | 15 | 32 | 24 | 5.6 |
| Magnesium (mg) | 42 | 13 | 12 | 16 | 34 |
| Iron (mg) | 6.8 | 3.6 | 2.7 | 2.0 | 2.8 |
| Zinc (mg) | 4.2 | 0.7 | 2 | 2.1 | 1.1 |
| Selenium (mg) | 70.7 | 33.9 | 37.7 | - | - |
| Chr | Marker | Map Position | QTL Type | LOD | Encoding Gene | References |
|---|---|---|---|---|---|---|
| 1A | IWB42976 | 10.6 | GWAS | 3.2 | - | [47] |
| 1B | Xhbg406 | 75.0 | RIL | 3.3 | - | [56] |
| 2A | IWB66738 | 11.2 | GWAS | 3.3 | Starch synthase II | [47] |
| 2A | IWB1280 | 47.8 | RIL | 4.5 | - | [48] |
| 2A | IWB26593 | 197.0 | GWAS | 3.1 | β-amylase | [47] |
| 2B | IWB30115 | 2.1 | RIL | 4.7 | - | [48] |
| 2B | IWB1898 | 14.5 | GWAS | 3.5 | (1,4)-beta-xylanase | [47] |
| 2B | IWB23783 | 42.3 | RIL | 3.8 | β-glucosidase 1a | [48] |
| 3A | Xbarc45 | 28.5 | RIL | 2.8 | - | [56] |
| 5B | Xgwm540 | 49.0 | RIL | 5.3 | - | [56] |
| 5B | IWB70546 | 166.0 | GWAS | 3.2 | - | [47] |
| 6D | Xcfd80 | 29.9 | RIL | 3.1 | - | [56] |
| 7A | IWB74166 | 49.7 | GWAS | 3.4 | Isoamylase | [47] |
| 7A | IWB68797 | 90.9 | GWAS | 3.2 | Fructan 1-exohydrolase | [47] |
| Chrom | Marker | Map Position | QTL Type | LOD | Encoding Gene | References |
|---|---|---|---|---|---|---|
| 1A | Y34Ukr-RH13_TOTAX * | - | RIL | 3.2 | - | [93] |
| 1A | IWA3952 | 70.1 | GWAS | 3.1 | Glycosyl hydrolase | [27] |
| 1A | IWB64556 | 113.8 | GWAS | 3.0 | Glycosyltransferase | [27] |
| 1B | Y34Ukr-RH13_TOTAX * | - | RIL | 3.2 | - | [93] |
| 1B | IWB20653 | 62.6 | GWAS | 3.0 | - | [27] |
| 1B | IWB8152 | 110.2 | GWAS | 3.1 | - | [27] |
| 1B | wPt-4129 | 137.0 | RIL | 3.9 | - | [1] |
| 1B | BA00789946 | 277.0 | RIL | 5.1 | - | [93] |
| 1B | Xbcd508 | 116.0 | RIL | 2.9 | - | [2] |
| 1B | bcd508b | 116.0 | RIL | >2 | - | [3] |
| 2A | IWB10662 | 65.7 | GWAS | 3.1 | - | [27] |
| 2A | wpt-3114 | 85.7 | RIL | 22.4 | - | [4] |
| 2A | IWB32084 | 167.9 | GWAS | 3.0 | - | [27] |
| 2B | IWB71648 | 107.4 | GWAS | 3.0 | Cis-zeatin O-glucosyltransferase 1 | [27] |
| 3A | wPt-9422 | 79.6 | RIL | 4.1 | - | [1] |
| 3A | IWB41859 | 122.7 | GWAS | 3.1 | Glycosyl hydrolase | [27] |
| 3B | IWB34238 | 57.2 | GWAS | 3.0 | - | [27] |
| 3D | wpt-0485 | 0.0 | RIL | 9.9 | - | [4] |
| 4B | IWB71022 | 55.6 | GWAS | 3.0 | - | [27] |
| 4D | gpw-95001 | 46.0 | RIL | 37.7 | - | [4] |
| 5A | IWB21456 | 26.5 | GWAS | 3.0 | Glycosyltransferase | [27] |
| 5A | IWB10271 | 43.3 | GWAS | 4.0 | Cellulose synthase | [27] |
| 5A | IWB75097 | 63.7 | GWAS | 3.0 | Glucuronosyltransferase | [27] |
| 6A | IWB2064 | 126.2 | GWAS | 3.0 | - | [27] |
| 6B | IWB9021 | 82.6 | GWAS | 4.0 | - | [27] |
| 6B | gwm680 | 112.4 | RIL | 9.2 | - | [4] |
| 7A | IWB73134 | 42.1 | GWAS | 6.1 | 1,3-β-D-glucan synthase | [27] |
| 7A | IWA2658 | 130.3 | GWAS | 3.0 | β-1,4-endo glucanase | [27] |
| 7A | IWB34095 | 179.2 | GWAS | 3.0 | - | [27] |
| 7B | wPt-0963 | 42.2 | RIL | 4.0 | - | [1] |
| 7B | IWB45276 | 83.1 | GWAS | 3.0 | - | [27] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marcotuli, I.; Colasuonno, P.; Hsieh, Y.S.Y.; Fincher, G.B.; Gadaleta, A. Non-Starch Polysaccharides in Durum Wheat: A Review. Int. J. Mol. Sci. 2020, 21, 2933. https://doi.org/10.3390/ijms21082933
Marcotuli I, Colasuonno P, Hsieh YSY, Fincher GB, Gadaleta A. Non-Starch Polysaccharides in Durum Wheat: A Review. International Journal of Molecular Sciences. 2020; 21(8):2933. https://doi.org/10.3390/ijms21082933
Chicago/Turabian StyleMarcotuli, Ilaria, Pasqualina Colasuonno, Yves S. Y. Hsieh, Geoffrey B. Fincher, and Agata Gadaleta. 2020. "Non-Starch Polysaccharides in Durum Wheat: A Review" International Journal of Molecular Sciences 21, no. 8: 2933. https://doi.org/10.3390/ijms21082933
APA StyleMarcotuli, I., Colasuonno, P., Hsieh, Y. S. Y., Fincher, G. B., & Gadaleta, A. (2020). Non-Starch Polysaccharides in Durum Wheat: A Review. International Journal of Molecular Sciences, 21(8), 2933. https://doi.org/10.3390/ijms21082933