The Cbs Locus Affects the Expression of Senescence Markers and mtDNA Copy Number, but not Telomere Dynamics in Mice


Abstract
:1. Introduction
2. Results
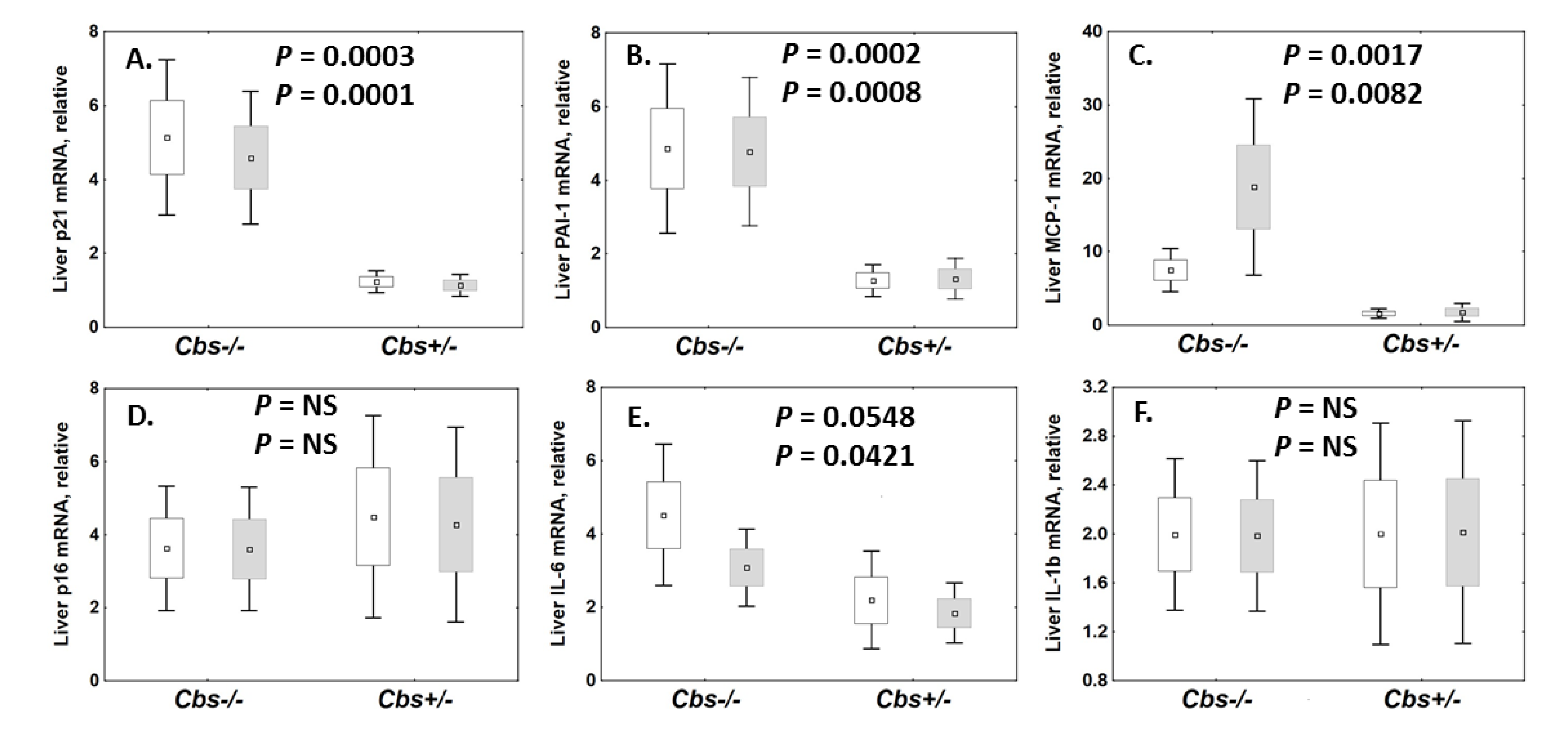
2.1. Cbs Genotype Affects the Expression of Senescence-Related mRNAs in Mice
2.1.1. Liver
2.1.2. Brain
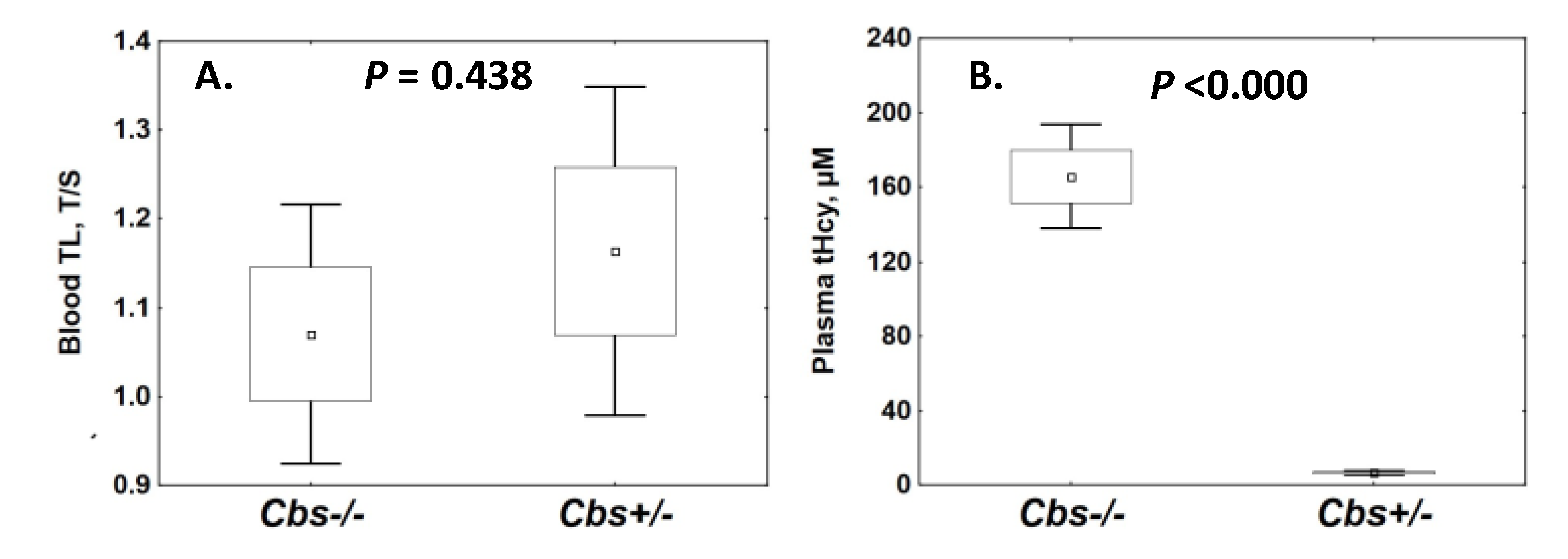
2.2. Cbs Genotype and TL
2.2.1. Blood TL
Effects of Age on Blood TL in Cbs+/− and Cbs−/− Mice
Effects of Sex on Blood TL in Cbs+/− and Cbs−/− Mice
2.2.2. Brain TL and Tert mRNA
2.2.3. Liver TL and Tert mRNA
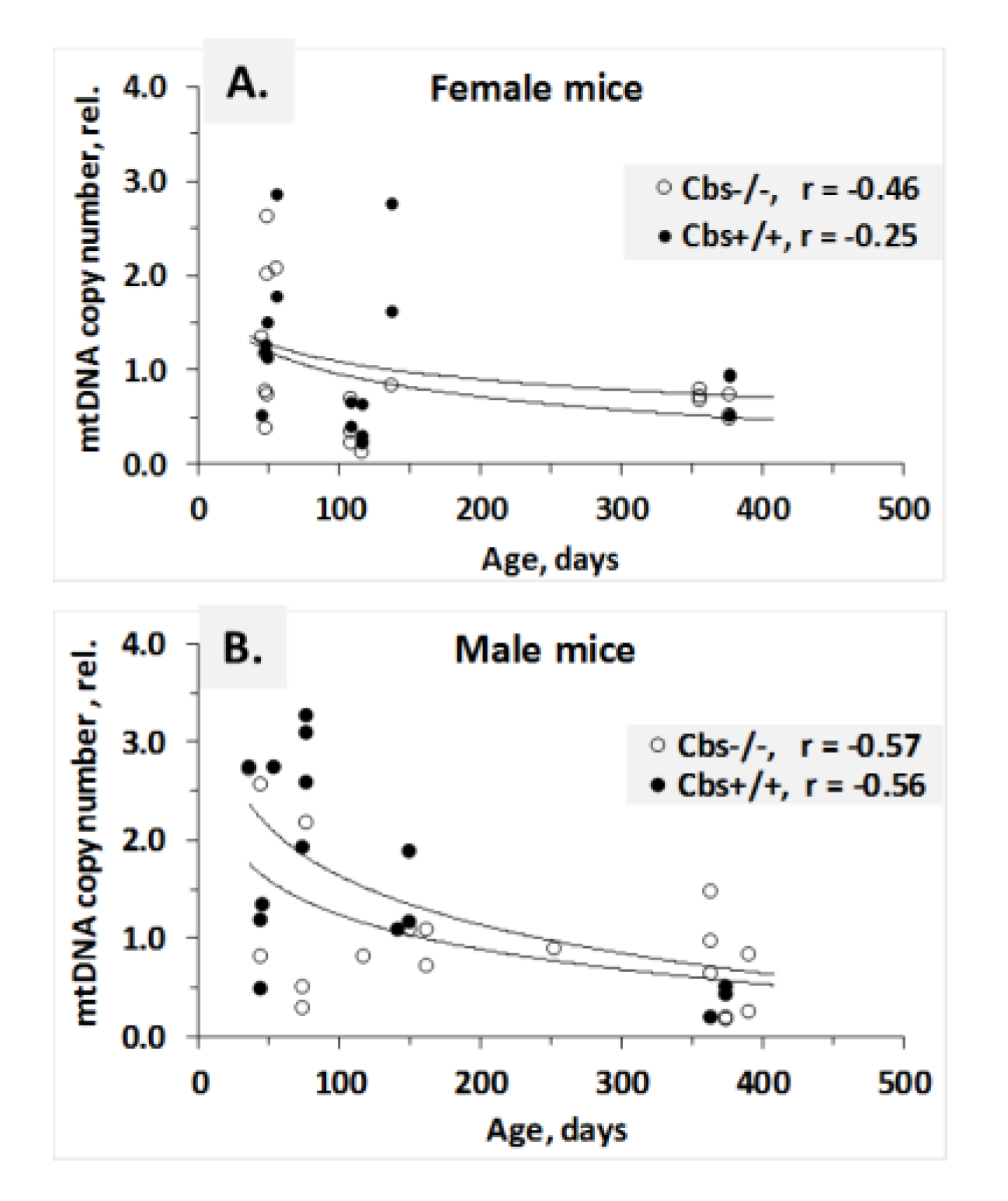
2.3. Cbs Genotype and mtDNA Copy Number in Blood, Liver, and Brain
2.3.1. Blood mtDNA
2.3.2. Brain and Liver mtDNA
2.4. Determinants of Blood TL in Mice
2.5. Determinants of TL in Mouse Brain and Liver
2.6. Determinants of Tert mRNA Expression in Mouse Brain and Liver
2.7. Determinants of Blood, Brain, and Liver mtDNA
2.7.1. Pearson Correlations
2.7.2. Multiple Regression
2.8. Determinants of Senescence-Related mRNAs in Mice
2.8.1. Liver
2.8.2. Brain
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Blood and Tissue Collection
4.3. Total Hcy Assays
4.4. DNA Extraction
4.5. Telomere Length Analysis
4.6. mtDNA Quantification
4.7. RNA Extraction
4.8. Quantification of Tert and Senescence-Related mRNAs by Real-Time qPCR
4.9. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Childs, B.G.; Durik, M.; Baker, D.J.; van Deursen, J.M. Cellular senescence in aging and age-related disease: From mechanisms to therapy. Nat. Med. 2015, 21, 1424–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughan, D.E.; Rai, R.; Khan, S.S.; Eren, M.; Ghosh, A.K. Plasminogen Activator Inhibitor-1 Is a Marker and a Mediator of Senescence. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1446–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.S.; Shah, S.J.; Klyachko, E.; Baldridge, A.S.; Eren, M.; Place, A.T.; Aviv, A.; Puterman, E.; Lloyd-Jones, D.M.; Heiman, M.; et al. A null mutation in SERPINE1 protects against biological aging in humans. Sci. Adv. 2017, 3, eaao1617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorgoulis, V.; Adams, P.D.; Alimonti, A.; Bennett, D.C.; Bischof, O.; Bishop, C.; Campisi, J.; Collado, M.; Evangelou, K.; Ferbeyre, G.; et al. Cellular Senescence: Defining a Path Forward. Cell 2019, 179, 813–827. [Google Scholar] [CrossRef]
- Shay, J.W.; Wright, W.E. Telomeres and telomerase: Three decades of progress. Nat. Rev. Genet. 2019, 20, 299–309. [Google Scholar] [CrossRef]
- Calado, R.T.; Dumitriu, B. Telomere dynamics in mice and humans. Semin. Hematol. 2013, 50, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Kim, N.W.; Piatyszek, M.A.; Prowse, K.R.; Harley, C.B.; West, M.D.; Ho, P.L.; Coviello, G.M.; Wright, W.E.; Weinrich, S.L.; Shay, J.W. Specific association of human telomerase activity with immortal cells and cancer. Science 1994, 266, 2011–2015. [Google Scholar] [CrossRef]
- Shay, J.W.; Bacchetti, S. A survey of telomerase activity in human cancer. Eur. J. Cancer 1997, 33, 787–791. [Google Scholar] [CrossRef]
- Cawthon, R.M.; Smith, K.R.; O’Brien, E.; Sivatchenko, A.; Kerber, R.A. Association between telomere length in blood and mortality in people aged 60 years or older. Lancet 2003, 361, 393–395. [Google Scholar] [CrossRef]
- Sanders, J.L.; Boudreau, R.M.; Penninx, B.W.; Simonsick, E.M.; Kritchevsky, S.B.; Satterfield, S.; Harris, T.B.; Bauer, D.C.; Newman, A.B.; Health, A.B.C.S. Association of a Modified Physiologic Index with mortality and incident disability: The Health, Aging, and Body Composition study. J. Gerontol. A. Biol. Sci. Med. Sci. 2012, 67, 1439–1446. [Google Scholar] [CrossRef] [Green Version]
- Whittemore, K.; Vera, E.; Martinez-Nevado, E.; Sanpera, C.; Blasco, M.A. Telomere shortening rate predicts species life span. Proc. Natl. Acad. Sci. USA 2019, 116, 15122–15127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Codd, V.; Nelson, C.P.; Albrecht, E.; Mangino, M.; Deelen, J.; Buxton, J.L.; Hottenga, J.J.; Fischer, K.; Esko, T.; Surakka, I.; et al. Identification of seven loci affecting mean telomere length and their association with disease. Nat. Genet. 2013, 45, 422–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, J.L.; Newman, A.B. Telomere length in epidemiology: A biomarker of aging, age-related disease, both, or neither? Epidemiol. Rev. 2013, 35, 112–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yakes, F.M.; Van Houten, B. Mitochondrial DNA damage is more extensive and persists longer than nuclear DNA damage in human cells following oxidative stress. Proc. Natl. Acad. Sci. USA 1997, 94, 514–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baek, J.H.; Son, H.; Jeong, Y.H.; Park, S.W.; Kim, H.J. Chronological Aging Standard Curves of Telomere Length and Mitochondrial DNA Copy Number in Twelve Tissues of C57BL/6 Male Mouse. Cells 2019, 8, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagouge, M.; Larsson, N.G. The role of mitochondrial DNA mutations and free radicals in disease and ageing. J. Intern. Med. 2013, 273, 529–543. [Google Scholar] [CrossRef] [Green Version]
- Moore, A.Z.; Ding, J.; Tuke, M.A.; Wood, A.R.; Bandinelli, S.; Frayling, T.M.; Ferrucci, L. Influence of cell distribution and diabetes status on the association between mitochondrial DNA copy number and aging phenotypes in the InCHIANTI study. Aging Cell 2018, 17, e12683. [Google Scholar] [CrossRef] [Green Version]
- Kujoth, G.C.; Hiona, A.; Pugh, T.D.; Someya, S.; Panzer, K.; Wohlgemuth, S.E.; Hofer, T.; Seo, A.Y.; Sullivan, R.; Jobling, W.A.; et al. Mitochondrial DNA mutations, oxidative stress, and apoptosis in mammalian aging. Science 2005, 309, 481–484. [Google Scholar] [CrossRef]
- Ylikallio, E.; Tyynismaa, H.; Tsutsui, H.; Ide, T.; Suomalainen, A. High mitochondrial DNA copy number has detrimental effects in mice. Hum. Mol. Genet. 2010, 19, 2695–2705. [Google Scholar] [CrossRef] [Green Version]
- Jakubowski, H.; Boers, G.H.; Strauss, K.A. Mutations in cystathionine beta-synthase or methylenetetrahydrofolate reductase gene increase N-homocysteinylated protein levels in humans. FASEB J. 2008, 22, 4071–4076. [Google Scholar] [CrossRef]
- Chwatko, G.; Boers, G.H.; Strauss, K.A.; Shih, D.M.; Jakubowski, H. Mutations in methylenetetrahydrofolate reductase or cystathionine beta-synthase gene, or a high-methionine diet, increase homocysteine thiolactone levels in humans and mice. FASEB J. 2007, 21, 1707–1713. [Google Scholar] [CrossRef] [PubMed]
- Jakubowski, H. Homocysteine in Protein Structure/Function and Human Disease—Chemical Biology of Homocysteine-Containing Proteins; Springer: Wien, Austria, 2013. [Google Scholar]
- Jakubowski, H. Homocysteine Modification in Protein Structure/Function and Human Disease. Physiol. Rev. 2019, 99, 555–604. [Google Scholar] [CrossRef] [PubMed]
- Mudd, S.H.; Skovby, F.; Levy, H.L.; Pettigrew, K.D.; Wilcken, B.; Pyeritz, R.E.; Andria, G.; Boers, G.H.; Bromberg, I.L.; Cerone, R.; et al. The natural history of homocystinuria due to cystathionine beta-synthase deficiency. Am. J. Hum. Genet. 1985, 37, 1–31. [Google Scholar] [PubMed]
- Mudd, S.H.; Levy, H.L.; Kraus, J.P. Disorders of transsulfuration. In The Metabolic and Molecular Bases of Inherited Disease, 8th ed.; Scriver, C.R., Beaudet, A.L., Sly, W.S., Valle, D., Childs, B., Kinzler, K.W., Vogelstein, B., Eds.; Mc Graw-Hill: New York, NY, USA, 2001; Volume 2, pp. 2007–2056. [Google Scholar]
- Watanabe, M.; Osada, J.; Aratani, Y.; Kluckman, K.; Reddick, R.; Malinow, M.R.; Maeda, N. Mice deficient in cystathionine beta-synthase: Animal models for mild and severe homocyst(e)inemia. Proc. Natl. Acad. Sci. USA 1995, 92, 1585–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Kuhnisch, J.; Mustafa, A.; Lhotak, S.; Schlachterman, A.; Slifker, M.J.; Klein-Szanto, A.; High, K.A.; Austin, R.C.; Kruger, W.D. Mouse models of cystathionine beta-synthase deficiency reveal significant threshold effects of hyperhomocysteinemia. FASEB J. 2009, 23, 883–893. [Google Scholar] [CrossRef] [Green Version]
- Jakubowski, H.; Perla-Kajan, J.; Finnell, R.H.; Cabrera, R.M.; Wang, H.; Gupta, S.; Kruger, W.D.; Kraus, J.P.; Shih, D.M. Genetic or nutritional disorders in homocysteine or folate metabolism increase protein N-homocysteinylation in mice. FASEB J. 2009, 23, 1721–1727. [Google Scholar] [CrossRef] [Green Version]
- Perla-Kajan, J.; Utyro, O.; Rusek, M.; Malinowska, A.; Sitkiewicz, E.; Jakubowski, H. N-Homocysteinylation impairs collagen cross-linking in cystathionine beta-synthase-deficient mice: A novel mechanism of connective tissue abnormalities. FASEB J. 2016, 30, 3810–3821. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Segura, A.; de Jong, T.V.; Melov, S.; Guryev, V.; Campisi, J.; Demaria, M. Unmasking Transcriptional Heterogeneity in Senescent Cells. Curr. Biol. 2017, 27, 2652–2660.e4. [Google Scholar] [CrossRef] [Green Version]
- Sharpless, N.E.; Sherr, C.J. Forging a signature of in vivo senescence. Nat. Rev. Cancer 2015, 15, 397–408. [Google Scholar] [CrossRef]
- Guillen, N.; Navarro, M.A.; Arnal, C.; Noone, E.; Arbones-Mainar, J.M.; Acin, S.; Surra, J.C.; Muniesa, P.; Roche, H.M.; Osada, J. Microarray analysis of hepatic gene expression identifies new genes involved in steatotic liver. Physiol. Genomics 2009, 37, 187–198. [Google Scholar] [CrossRef] [Green Version]
- Masuyama, M.; Iida, R.; Takatsuka, H.; Yasuda, T.; Matsuki, T. Quantitative change in mitochondrial DNA content in various mouse tissues during aging. Biochim. Biophys. Acta 2005, 1723, 302–308. [Google Scholar] [CrossRef]
- Gadaleta, M.N.; Rainaldi, G.; Lezza, A.M.; Milella, F.; Fracasso, F.; Cantatore, P. Mitochondrial DNA copy number and mitochondrial DNA deletion in adult and senescent rats. Mutat. Res. 1992, 275, 181–193. [Google Scholar] [CrossRef]
- Fries, G.R.; Bauer, I.E.; Scaini, G.; Wu, M.J.; Kazimi, I.F.; Valvassori, S.S.; Zunta-Soares, G.; Walss-Bass, C.; Soares, J.C.; Quevedo, J. Accelerated epigenetic aging and mitochondrial DNA copy number in bipolar disorder. Transl. Psychiatry 2017, 7, 1283. [Google Scholar] [CrossRef]
- Hernandez-Segura, A.; Nehme, J.; Demaria, M. Hallmarks of Cellular Senescence. Trends Cell Biol. 2018, 28, 436–453. [Google Scholar] [CrossRef]
- Huang, T.; Rivera-Perez, J.A. Senescence-associated beta-galactosidase activity marks the visceral endoderm of mouse embryos but is not indicative of senescence. Genesis 2014, 52, 300–308. [Google Scholar] [CrossRef] [Green Version]
- Khayati, K.; Antikainen, H.; Bonder, E.M.; Weber, G.F.; Kruger, W.D.; Jakubowski, H.; Dobrowolski, R. The amino acid metabolite homocysteine activates mTORC1 to inhibit autophagy and form abnormal proteins in human neurons and mice. FASEB J. 2017, 31, 598–609. [Google Scholar] [CrossRef] [Green Version]
- Jakubowski, H. New method for the determination of protein N-linked homocysteine. Anal. Biochem. 2008, 380, 257–261. [Google Scholar] [CrossRef]
- Cawthon, R.M. Telomere length measurement by a novel monochrome multiplex quantitative PCR method. Nucleic Acids Res. 2009, 37, e21. [Google Scholar] [CrossRef] [Green Version]
- Cawthon, R.M. Telomere measurement by quantitative PCR. Nucleic Acids Res. 2002, 30, e47. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Malik, A.N.; Czajka, A.; Cunningham, P. Accurate quantification of mouse mitochondrial DNA without co-amplification of nuclear mitochondrial insertion sequences. Mitochondrion 2016, 29, 59–64. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Variable | Pearson Correlation | Multiple Regression | ||||
|---|---|---|---|---|---|---|
| Model 1, Hcy | Model 2, Cbs Genotype | |||||
| β | p | β | p | β | p | |
| Female (n = 39) | ||||||
| Age * | 0.14 | 0.473 | 0.15 | 0.478 | 0.10 | 0.559 |
| Hcy # | 0.25 | 0.897 | −0.03 | 0.878 | ||
| Cbs genotype | −0.28 | 0.884 | −0.08 | 0.611 | ||
| F = 0.27, p = 0.768, R2 = 0.02 | F = 0.36, p = 0.698, R2 = 0.02 | |||||
| Male (n = 41) | ||||||
| Age * | −0.56 | 0.002 | −0.50 | 0.008 | −0.38 | 0.021 |
| Hcy # | −0.34 | 0.077 | −0.16 | 0.361 | ||
| Cbs genotype | 0.40 | 0.033 | 0.11 | 0.493 | ||
| F = 6.40, p = 0.006, R2 = 0.34 | F = 4.01, p = 0.027, R2 = 0.18 | |||||
| All (n = 80) | ||||||
| Sex | 0.07 | 0.627 | 0.03 | 0.816 | 0.11 | 0.774 |
| Age * | −0.28 | 0.036 | −0.24 | 0.094 | −0.190 | 0.102 |
| Hcy # | −0.17 | 0.203 | −0.08 | 0.553 | ||
| Cbs genotype | 0.22 | 0.108 | 0.04 | 0.701 | ||
| F = 1.6, p = 0.195, R2 = 0.08 | F = 1.6, p = 0.193, R2 = 0.06 | |||||
| Variable | TL in | |||||
|---|---|---|---|---|---|---|
| Blood | Brain | Liver | ||||
| β | p | β | p | β | p | |
| Female (n = 40) | Female (n = 20) | |||||
| Age * | 0.10 | 0.559 | 0.53 | 0.023 | 0.20 | 0.392 |
| Cbs genotype | −0.08 | 0.611 | −0.10 | 0.641 | 0.29 | 0.228 |
| F = 0.36, p = 0.698, R2 = 0.02 | F = 3.25, p = 0.066, R2 = 0.20 | F = 1.11, p = 0.351, R2 = 0.012 | ||||
| Male (n = 41) | Male (n = 23) | |||||
| Age * | −0.38 | 0.021 | 0.39 | 0.078 | −0.29 | 0.181 |
| Cbs genotype | 0.11 | 0.493 | −0.17 | 0.423 | 0.18 | 0.417 |
| F = 4.01, p = 0.027, R2 = 0.18 | F = 1.65, p = 0.217, R2 = 0.14 | |||||
| All (n = 80) | All (n = 43) | |||||
| Sex | 0.11 | 0.774 | −0.55 | 0.000 | 0.48 | 0.001 |
| Age * | −0.190 | 0.102 | 0.38 | 0.003 | −0.006 | 0.964 |
| Cbs genotype | 0.04 | 0.701 | −0.12 | 0.340 | 0.23 | 0.110 |
| F = 1.61, p = 0.193, R2 = 0.06 | F = 9.70, p = 0.000, R2 = 0.39 | F = 4.60, p = 0.008, R2 = 0.26 | ||||
| Variable | Blood mtDNA Cbs−/− (n = 40), Cbs+/− (n = 40) | Brain mtDNA Cbs−/− (n = 19), Cbs+/− (n = 24) | Liver mtDNA Cbs−/− (n = 19), Cbs+/− (n = 24) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pearson Correlation | Multiple Regression | Pearson Correlation | Multiple Regression | Pearson Correlation | Multiple Regression | |||||||||
| Model 1, Hcy | Model 2, Cbs | |||||||||||||
| β | p | β | p | β | p | β | p | β | p | β | p | β | p | |
| Females (n = 39) | Females (n = 20) | Females (n = 20) | ||||||||||||
| Age * | −0.01 | 0.960 | −0.05 | 0.832 | −0.29 | 0.090 | 0.49 | 0.03 | 0.49 | 0.032 | 0.71 | 0.000 | 0.72 | 0.000 |
| tHcy # | 0.08 | 0.703 | 0.10 | 0.670 | ||||||||||
| Cbs genotype | 0.02 | 0.918 | 0.09 | 0.600 | 0.08 | 0.74 | 0.10 | 0.627 | 0.31 | 0.177 | 0.35 | 0.030 | ||
| F = 0.1, p = 0.91, R2= 0.01 | F = 1.8, p = 0.18, R2 = 0.10 | F = 2.2, p = 0.09, R2 = 0.25 | F = 14.1, p = 0.000, R2 = 0.63 | |||||||||||
| Males (n = 41) | Males (n = 23) | Males (n = 23) | ||||||||||||
| Age * | −0.57 | 0.005 | -0.50 | 0.023 | −0.58 | 0.000 | 0.49 | 0.02 | 0.49 | 0.023 | 0.60 | 0.003 | 0.62 | 0.003 |
| tHcy # | −0.38 | 0.070 | -0.17 | 0.419 | ||||||||||
| Cbs genotype | 0.50 | 0.014 | 0.27 | 0.058 | −0.11 | 0.62 | 0.000 | 0.988 | −0.04 | 0.858 | 0.10 | 0.580 | ||
| F = 5.3, p = 0.014, R2 = 0.35 | F = 13.2, p = 0.000, R2 = 0.48 | F =3.28, p = 0.063, R2 = 0.24 | F = 5.9, p = 0.009, R2 = 0.37 | |||||||||||
| All (n = 80) | All (n = 43) | All (n = 43) | ||||||||||||
| Sex | −0.33 | 0.021 | −0.36 | 0.008 | −0.25 | 0.018 | −0.40 | 0.007 | −0.41 | 0.003 | −0.11 | 0.476 | −0.10 | 0.357 |
| Age * | −0.32 | 0.025 | −0.33 | 0.028 | −0.45 | 0.000 | 0.42 | 0.005 | 0.44 | 0.002 | 0.67 | 0.000 | 0.71 | 0.000 |
| tHcy # | −0.21 | 0.144 | −0.08 | 0.584 | ||||||||||
| Cbs genotype | 0.29 | 0.044 | 0.18 | 0.088 | 0.01 | 0.97 | 0.02 | 0.860 | 0.14 | 0.354 | 0.24 | 0.041 | ||
| F = 4.8, p = 0.006, R2 = 0.24 | F = 9.0, p = 0.000, R2 = 0.29 | F = 7.0, p = 0.001, R2 = 0.35 | F = 14.3, p = 0.000, R2 = 0.44 | |||||||||||
| Variable | Pai-1 mRNA | p21 mRNA | Mcp-1 mRNA | Il-6 mRNA | Kl mRNA | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Brain | Liver | Brain | Liver | Brain | Liver | Brain | Liver | Brain | ||||||||||
| β | p | β | p | β | p | β | p | β | p | β | p | β | p | β | p | β | p | |
| Females (n = 18–20) | ||||||||||||||||||
| Age * | −0.15 | 0.524 | −0.21 | 0.362 | 0.14 | 0.621 | 0.23 | 0.476 | 0.69 | 0.001 | −0.15 | 0.497 | −0.48 | 0.043 | ||||
| Cbs gene | −0.16 | 0.520 | −0.34 | 0.021 | −0.66 | 0.007 | −0.62 | 0.006 | −0.34 | 0.016 | ||||||||
| TL | −0.11 | 0.436 | 0.16 | 0.536 | 0.14 | 0.609 | ||||||||||||
| Tert | 0.42 | 0.116 | 0.71 | 0.006 | −0.10 | 0.696 | −0.05 | 0.851 | 0.15 | 0.006 | 0.39 | 0.037 | 0.62 | 0.004 | 0.17 | 0.443 | ||
| mtDNA | −0.21 | 0.419 | 0.64 | 0.002 | 0.04 | 0.856 | 0.61 | 0.002 | ||||||||||
| NS | F = 15.7, p = 0.000, R2 = 0.87 | NS | F = 5.4, p = 0.02, R2 = 0.03 | NS | F = 4.84, p = 0.015, R2=0.49 | F = 9.76, p = 0.002, R2=0.57 | F = 20.0, p = 0.000, R2 = 0.86 | F = 3.70, p = 0.047, R2 = 0.32 | ||||||||||
| Males (n = 20–21) | ||||||||||||||||||
| Age * | −0.31 | 0.207 | −0.30 | 0.327 | 1.22 | 0.002 | 0.52 | 0.005 | 0.68 | 0.039 | 0.62 | 0.015 | −0.48 | 0.060 | ||||
| Cbs gene | 0.31 | 0.207 | −0.30 | 0.139 | −0.47 | 0.034 | −0.44 | 0.013 | −0.14 | 0.321 | ||||||||
| TL | −0.72 | 0.002 | −0.03 | 0.880 | −1.13 | 0.003 | −1.10 | 0.003 | ||||||||||
| Tert | 0.76 | 0.020 | 0.11 | 0.554 | −0.50 | 0.082 | 0.01 | 0.960 | 0.10 | 0.645 | −0.65 | 0.016 | 0.18 | 0.468 | ||||
| mtDNA | 0.36 | 0.189 | 0.73 | 0.001 | ||||||||||||||
| F = 4.7, p = 0.031, R2=0.61 | F = 4.5, p = 0.020, R2 = 0.35 | F = 8.78, p = 0.004, R2 = 0.57 | NS | F = 6.17, p = 0.010, R2 = 0.63 | F = 13.0, p = 0.000, R2 = 0.78 | F = 5.19, p = 0.016, R2 = 0.56 | F = 8.9, p = 0.000, R2 = 0.66 | NS | ||||||||||
| All (n = 39–43) | ||||||||||||||||||
| Sex | −0.35 | 0.044 | −0.15 | 0.270 | 0.46 | 0.010 | ||||||||||||
| Age * | -0.22 | 0.140 | 0.32 | 0.061 | 0.43 | 0.002 | 0.45 | 0.008 | 0.25 | 0.093 | −0.40 | 0.011 | ||||||
| Cbs gene | 0.19 | 0.270 | -0.52 | 0.001 | −0.52 | 0.001 | −0.43 | 0.001 | −0.19 | 0.112 | ||||||||
| TL | −0.36 | 0.028 | −0.20 | 0.259 | −0.28 | 0.095 | ||||||||||||
| Tert | 0.15 | 0.378 | 0.001 | 0.993 | 0.38 | 0.019 | 0.31 | 0.044 | ||||||||||
| mtDNA | −0.43 | 0.018 | 0.56 | 0.000 | ||||||||||||||
| F = 3.3, p = 0.037, R2 = 0.27 | F = 7.1, p = 0.003, R2 = 0.29 | F = 6.0, p = 0.002, R2 = 0.30 | F = 8.5, p = 0.001, R2 = 0.31 | F = 3.2, p = 0.036, R2 = 0.23 | F = 13.5, p = 0.000, R2 = 0.43 | F = 4.9, p = 0.007, R2 = 0.26 | F = 16.9, p = 0.000, R2 = 0.57 | F = 8.4, p = 0.001, R2 = 0.34 | ||||||||||
| Variable | p16 mRNA | Il-1β mRNA | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Brain | Liver | Brain | Liver | |||||||||
| β | p | β | p | β | p | β | p | |||||
| Females (n = 18–20) | ||||||||||||
| Age * | 0.84 | 0.000 | 0.78 | 0.085 | 0.30 | 0.214 | 0.34 | 0.099 | ||||
| Cbs gene | 0.25 | 0.065 | 0.33 | 0.086 | 0.14 | 0.561 | 0.08 | 0.616 | ||||
| TL | ||||||||||||
| Tert | −0.07 | 0.806 | −0.18 | 0.466 | −0.38 | 0.015 | ||||||
| mtDNA | 0.63 | 0.008 | ||||||||||
| F = 23.8, p = 0.000, R2 = 0.74 | F = 7.2, p = 0.003, R2 = 0.59 | NS | F = 11.8, p = 0.000, R2=0.69 | |||||||||
| Males (n = 20–21) | ||||||||||||
| Age * | 0.82 | 0.000 | 0.41 | 0.078 | 0.46 | 0.142 | 0.13 | 0.603 | ||||
| Cbs gene | −0.05 | 0.714 | 0.04 | 0.790 | 0.02 | 0.939 | −0.02 | 0.876 | ||||
| TL | ||||||||||||
| Tert | 0.45 | 0.052 | 0.06 | 0.834 | 0.08 | 0.772 | ||||||
| mtDNA | 0.65 | 0.004 | ||||||||||
| F = 20.7, p = 0.000, R2 = 0.70 | F = 12.4, p = 0.000, R2 = 0.66 | NS | F = 7.5, p = 0.001, R2 = 0.54 | |||||||||
| All (n = 39–43) | ||||||||||||
| Sex | 0.09 | 0.014 | −0.18 | 0.180 | −0.18 | 0.140 | ||||||
| Age * | 0.09 | 0.000 | 0.41 | 0.058 | 0.17 | 0.392 | ||||||
| Cbs gene | 0.09 | 0.114 | 0.15 | 0.278 | 0.03 | 0.854 | −0.02 | 0.863 | ||||
| TL | −0.01 | 0.923 | 0.13 | 0.419 | −0.19 | 0.184 | ||||||
| Tert | 0.30 | 0.137 | −0.17 | 0.307 | −0.02 | 0.924 | ||||||
| mtDNA | 0.04 | 0.785 | 0.46 | 0.005 | 0.70 | 0.000 | ||||||
| F = 27.8, p = 0.000, R2 = 0.69 | F = 11.7, p = 0.000, R2 = 0.56 | F = 3.4, p = 0.021, R2 = 0.31 | F = 10.4, p = 0.000, R2 = 0.64 | |||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Utyro, O.; Perła-Kaján, J.; Jakubowski, H. The Cbs Locus Affects the Expression of Senescence Markers and mtDNA Copy Number, but not Telomere Dynamics in Mice. Int. J. Mol. Sci. 2020, 21, 2520. https://doi.org/10.3390/ijms21072520
Utyro O, Perła-Kaján J, Jakubowski H. The Cbs Locus Affects the Expression of Senescence Markers and mtDNA Copy Number, but not Telomere Dynamics in Mice. International Journal of Molecular Sciences. 2020; 21(7):2520. https://doi.org/10.3390/ijms21072520
Chicago/Turabian StyleUtyro, Olga, Joanna Perła-Kaján, and Hieronim Jakubowski. 2020. "The Cbs Locus Affects the Expression of Senescence Markers and mtDNA Copy Number, but not Telomere Dynamics in Mice" International Journal of Molecular Sciences 21, no. 7: 2520. https://doi.org/10.3390/ijms21072520