Can Ceylon Leadwort (Plumbago zeylanica L.) Acclimate to Lead Toxicity?—Studies of Photosynthetic Apparatus Efficiency

 , , , , , , and
, , , , , , and
Abstract
:
1. Introduction
2. Results
2.1. Growth and Development of P. zeylanica
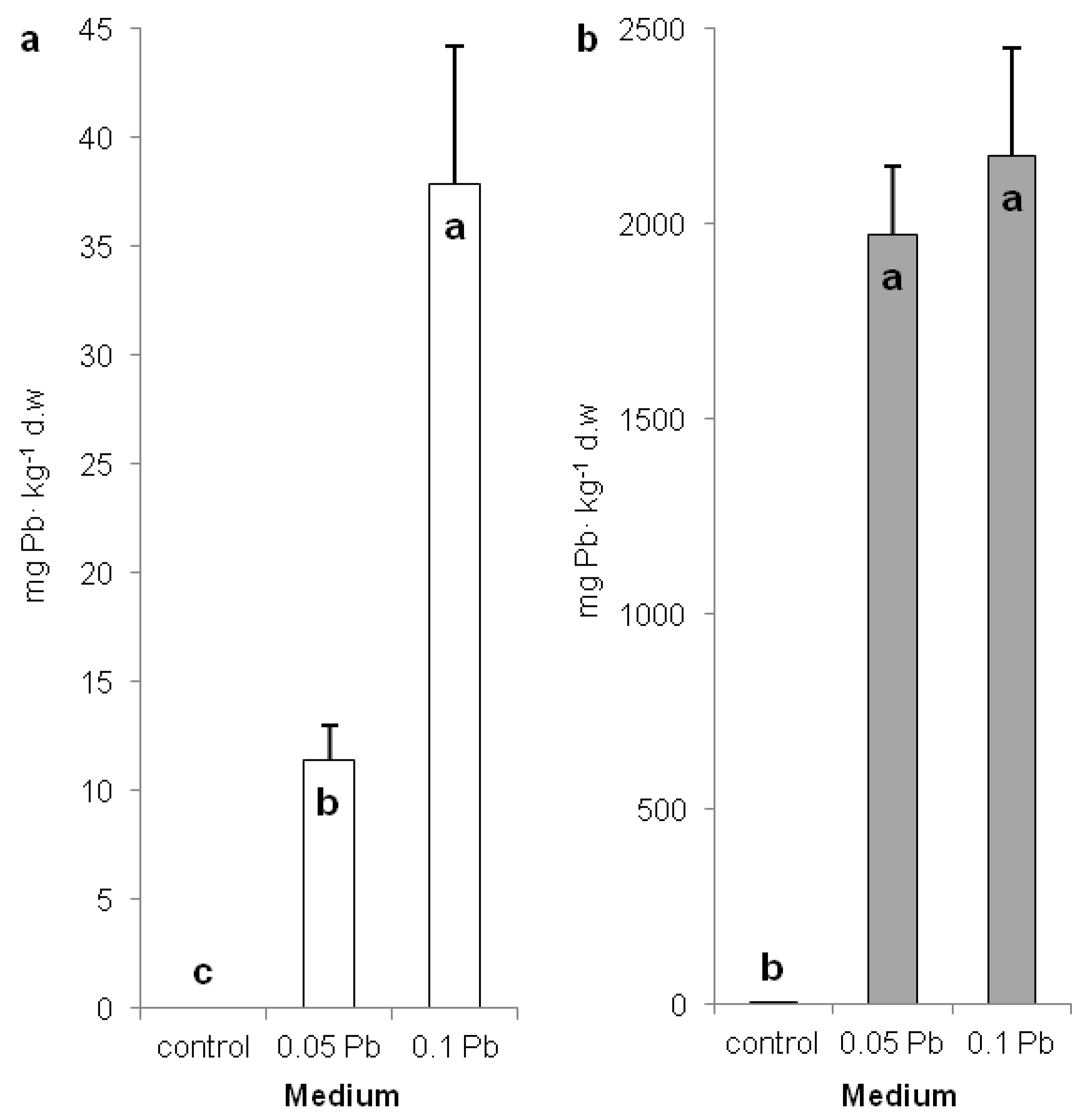
2.2. Lead Concentration in Shoots and Roots
2.3. The Level of Cell Membranes Damage
2.4. Accumulation of Protecting Secondary Metabolites
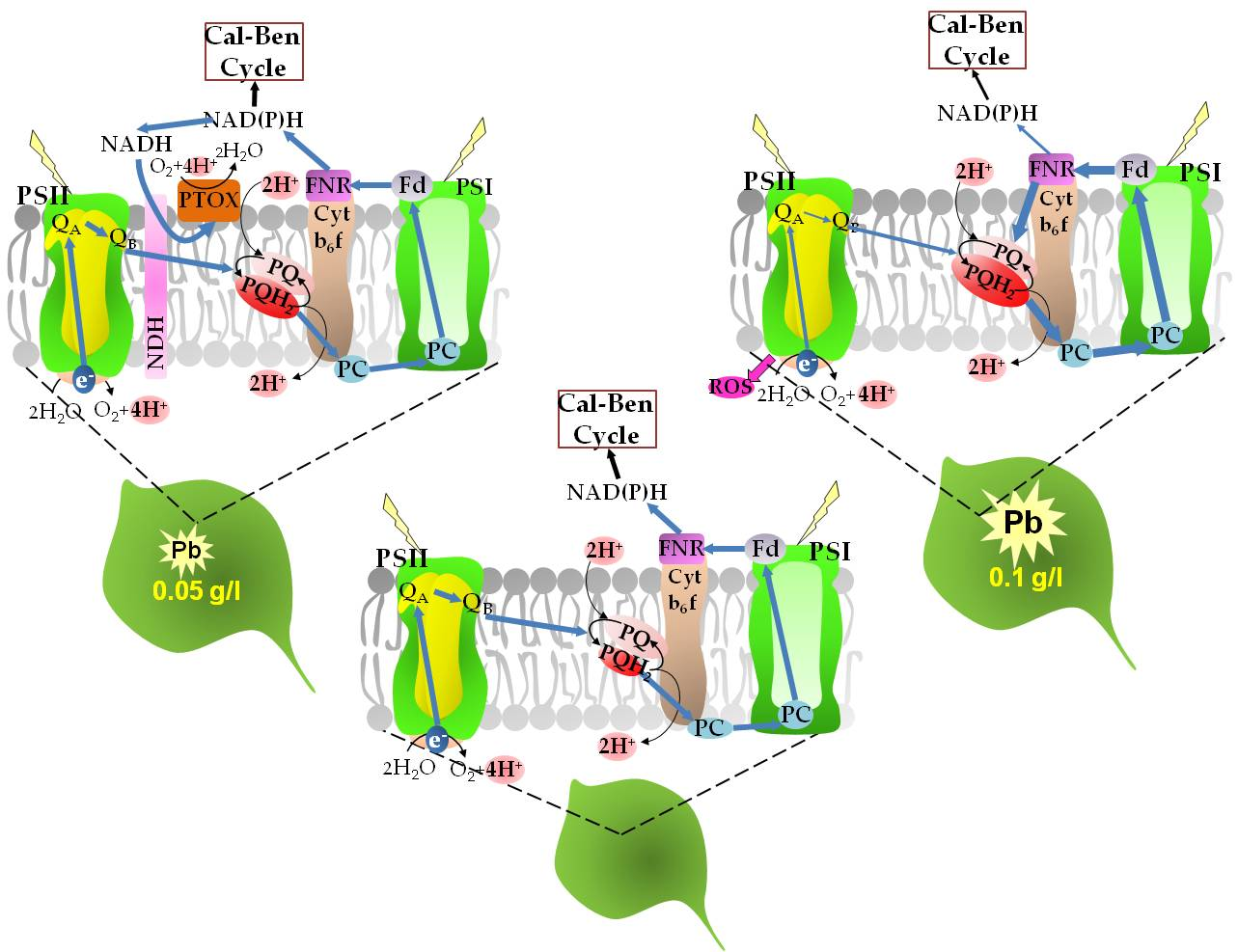
2.5. Effect of Lead on Photosynthetic Apparatus Efficiency
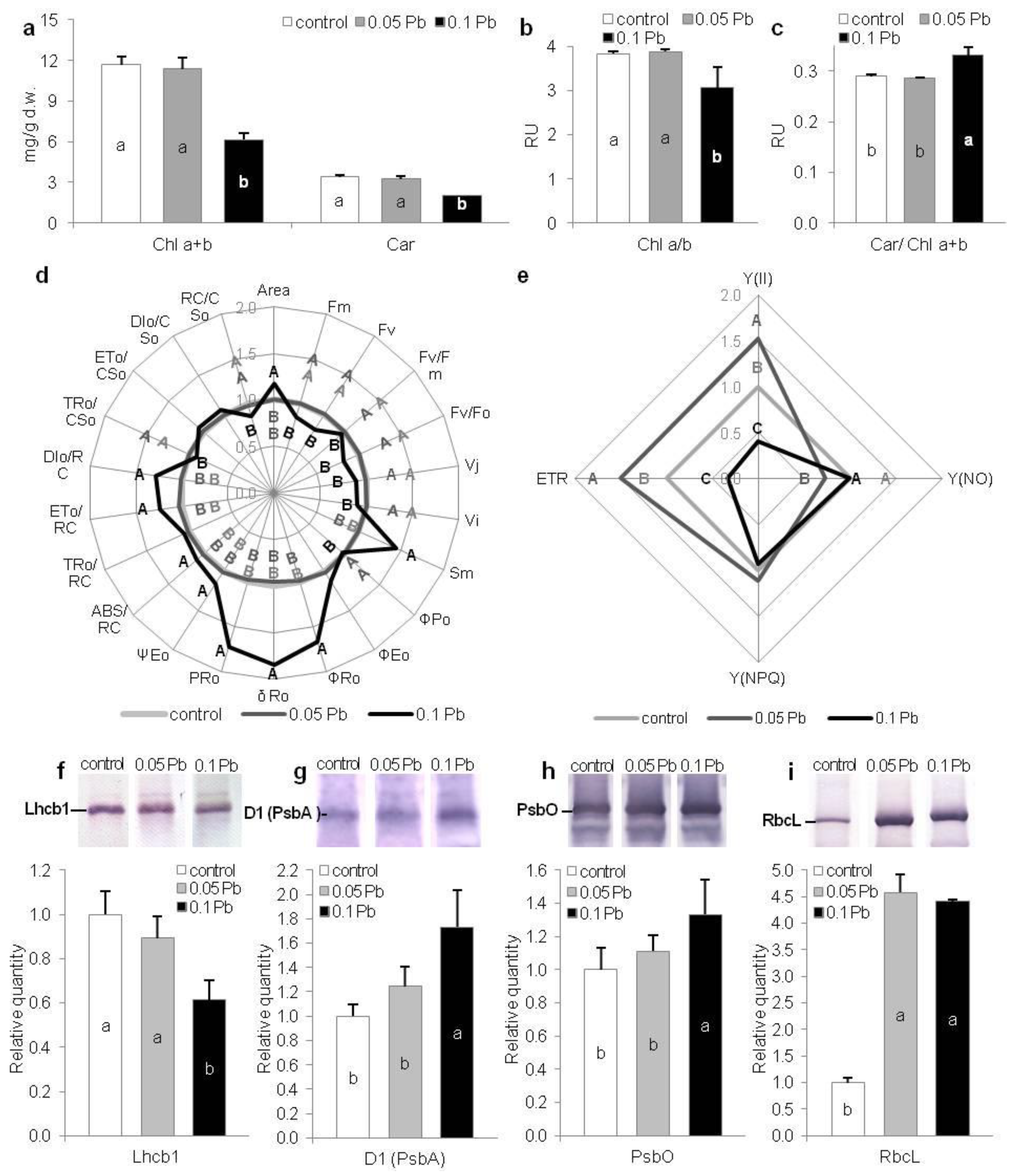
2.5.1. Photosynthetic Pigments’ Content
2.5.2. Chlorophyll a Fluorescence
2.5.3. Key Photosynthetic Proteins
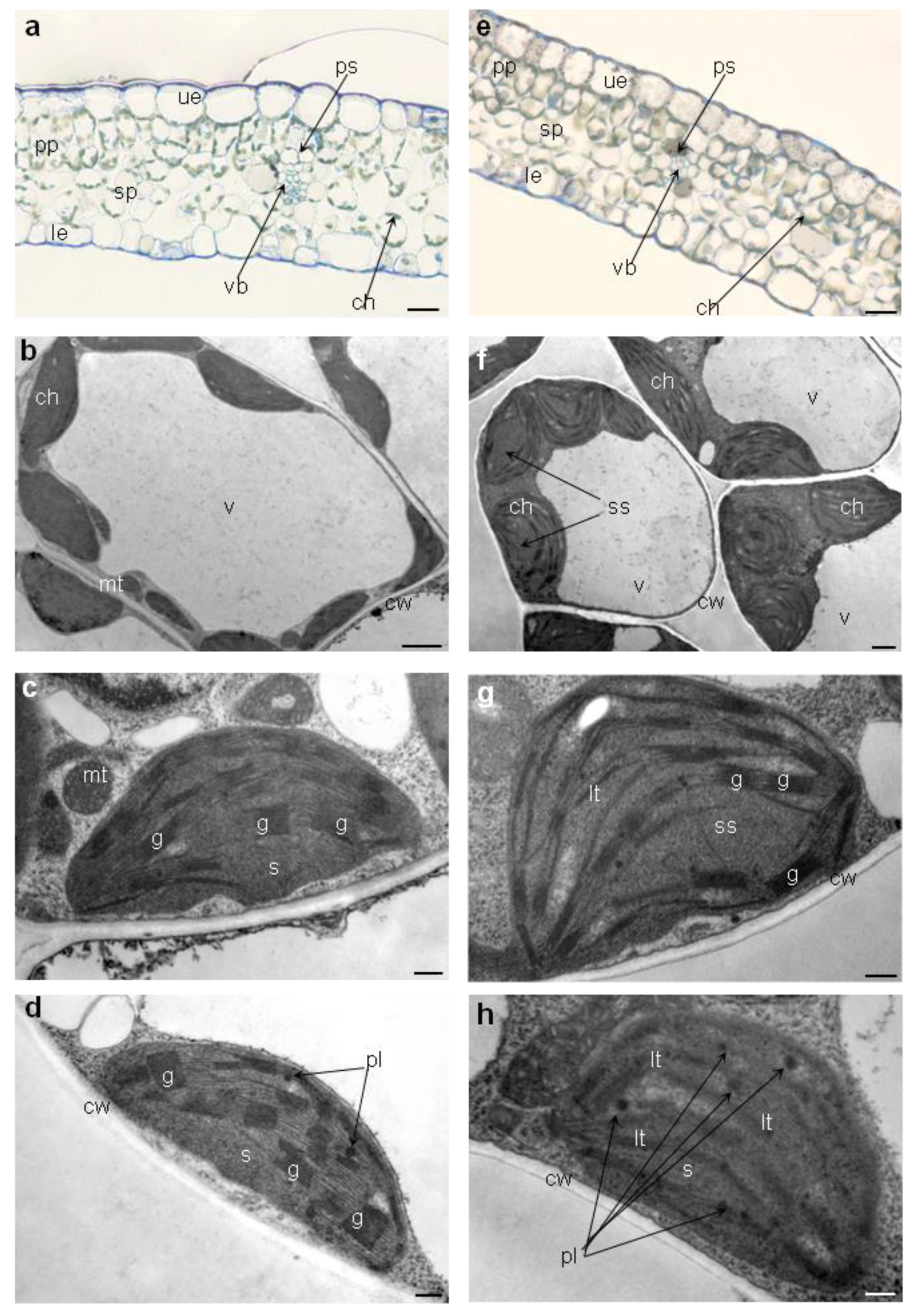
2.5.4. Leaves’ Anatomy and Chloroplasts’ Ultrastructure
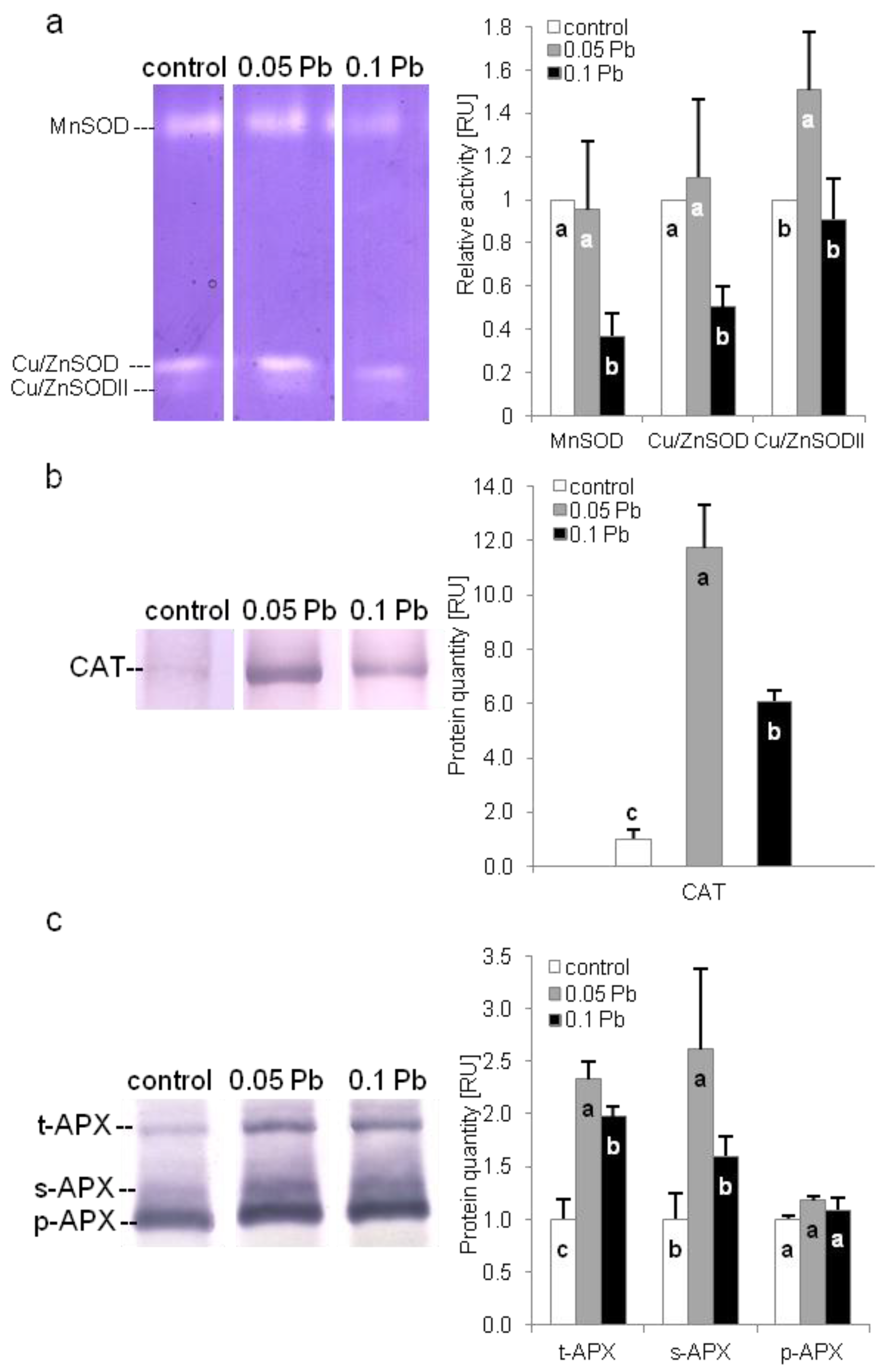
2.5.5. Antioxidant Enzymes Protection
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Experimental Conditions and Stress Treatment
4.3. Biometric Assay
4.4. Estimation of Lead Concentration in Plant Tissue
4.5. Estimation of Lipid Membranes Peroxidation Level
4.6. Phenolic Compounds’ Content Determination
4.6.1. Total Phenols
4.6.2. Specific Phenol Groups
4.6.3. Plumbagin
4.7. Evaluation of Photosynthetic Apparatus Efficiency
4.7.1. Photosynthetic Pigment Content Estimation
4.7.2. Chlorophyll a Fluorescence Measurement
4.7.3. Protein Content Determination
4.7.4. Histological and TEM Observation
4.7.5. Evaluation of Antioxidant Enzymes
4.8. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALAD | alpha amino levulinic dehydrogenase |
| APX | ascorbate peroxidase |
| BCIP | 5-bromo-4-chloro-3-indolyl phosphate |
| BSA | bovine serum albumin |
| CAD | cinnamic acid derivatives |
| CAT | catalase |
| CEF | cyclic electron flow |
| CS | cross section |
| Cu/ZnSOD | copper- and zinc-containing superoxide dismutase |
| Cu/ZnSODII | chloroplastic copper- and zinc-containing superoxide dismutase |
| DTT | dithiothreitol |
| DW | dry weight |
| EDTA | ethylenediaminetetraacetic acid |
| ETR | electron transport rate |
| F-C | Folin–Ciocalteu |
| Fd | ferredoxin |
| FFW | ferredoxin |
| FQR | ferredoxin-plastoquinone oxidoreductase |
| GI | growth index |
| HPLC | high performance liquid chromatography |
| IFW | initial fresh weight |
| Lhcb1 | light-harvesting chlorophyll a/b binding protein |
| LHCII | light-harvesting complex of PSII |
| MDA | malondialdehyde |
| MEP | methyl eritrythol phosphate pathway |
| MnSOD | manganese superoxide dismutase |
| MQ | Milli-Q |
| MS | Murashige and Skoog medium |
| NADP+ | nicotinamide adenine dinucleotide phosphate |
| NADPH | reduced nicotinamide adenine dinucleotide phosphate |
| NBT | nitro blue tetrazolium |
| NDH | NAD(P)H dehydrogenase complex |
| NPQ | non-photochemical quenching |
| OEC | oxygen evolving complex |
| PC | plastocyanin |
| PQ | plastoquinone |
| PsbA | Photosystem II reaction center protein A |
| PsbO | oxygen-evolving enhancer protein 1 |
| PSI | Photosystem I |
| PSII | Photosystem II |
| PTOX | terminal plastoquinone oxidase |
| PVPD | polyvinylidene fluoride membrane |
| PVPP | polyvinylpolypyrrolidone |
| RC | reaction center |
| RbcL | rubisco large subunit |
| ROS | reactive oxygen species |
| Rubisco | ribulose-1,5-bisphosphate carboxylase |
| SDS-PAGE | sodium dodecyl sulfate–polyacrylamide gel electrophoresis |
| SOD | superoxide dismutase |
| TBA | thiobarbituric acid |
| TCA | trichloroacetic acid |
| TEM | transmission electron microscopy |
| THF | tetrahydrofuran |
References
- Alloway, B.J. Sources of heavy metals and metalloids in soils. In Heavy Metals in Soils. Environmental Pollution, 3rd ed.; Alloway, B., Ed.; Springer: Dordrecht, The Netherlands, 2013; Volume 22, pp. 11–50. [Google Scholar]
- Rozpądek, P.; Domka, A.; Ważny, R.; Nosek, M.; Jędrzejczyk, R.; Tokarz, K.; Turnau, K. How does the endophytic fungus Mucor sp. improve Arabidopsis arenosa vegetation in the degraded environment of a mine dump? Environ. Exp. Bot. 2018, 147, 31–42. [Google Scholar] [CrossRef]
- De Lurdes Dinis, M.; Fiúza, A. Exposure assessment to heavy metals in the environment: Measures to eliminate or reduce the exposure to critical receptors. In Environmental Heavy Metal Pollution and Effects on Child Mental Development. NATO Science for Peace and Security Series C: Environmental Securit; Simeonov, L., Kochubovski, M., Simeonova, B., Eds.; Springer: Dordrecht, The Netherlands, 2011; Volume 1, pp. 27–50. [Google Scholar]
- Tong, S.; Schirnding, Y.E.V.; Prapamontol, T. Environmental lead exposure: A public health problem of global dimensions. B World Health. Organ. 2000, 78, 1068–1077. [Google Scholar]
- Gupta, D.; Huang, H.; Yang, X.; Razafindrabe, B.; Inouhe, M. The detoxification of lead in Sedum alfredii H. is not related to phytochelatins but the glutathione. J. Hazard. Mater. 2010, 177, 437–444. [Google Scholar] [CrossRef]
- Krzesłowska, M.; Lenartowska, M.; Mellerowicz, E.J.; Samardakiewicz, S.; Woźny, A. Pectinous cell wall thickenings formation—a response of moss protonemata cells to lead. Environ. Exp. Bot. 2009, 656565, 119–131. [Google Scholar] [CrossRef]
- Maestri, E.; Marmiroli, M.; Visioli, G.; Marmiroli, N. Metal tolerance and hyperaccumulation:costs and trade-offs between traits and environment. Environ. Exp. Bot. 2010, 688, 1–13. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Lead toxicity in plants. Braz. J. Plant Physiol. 2005, 1717, 35–52. [Google Scholar] [CrossRef] [Green Version]
- Küpper, H. Lead toxicity in plants. In Lead—Its Effects on Environment and Health; Astrid, S., Sigel, H., Sigel, R.K.O., Eds.; Walter de Gruyter: Berlin, Germany, 2017; Volume 17, pp. 491–500. [Google Scholar]
- Cegiełkowska, W.; Michaska-Kacymirow, M.; Wierzbicka, M. Heavy metals in the environment. In Ecotoxicology of Plants, Soils, Metals; Wierzbicka, M., Ed.; University of Warsaw Publisher: Warszawa, Poland, 2015; Volume 1, pp. 22–36. (In Polish) [Google Scholar]
- Shahid, M.; Pourrut, B.; Dumat, C.; Nadeem, M.; Aslam, M.; Pinelli, E. Heavy-metal-induced reactive oxygen species: Phytotoxicity and physicochemical changes in plants. In Reviews of Environmental Contamination and Toxicology; Whitacre, D.M., Ed.; Springer International Publishing: Cham, Switzerland, 2014; Volume 232, pp. 114–131. [Google Scholar]
- Yilmaz, K.; Akinci, İ.E.; Akinci, S. Effect of lead accumulation on growth and mineral composition of eggplant seedlings (Solarium melongena). N. Z. J. Crop Hort. 2009, 37, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Pourrut, B.; Shahid, M.; Dumat, C.; Winterton, P.; Pinelli, E. Lead uptake, toxicity and detoxification in plants. Rev. Environ. Contam. Toxicol. 2011, 213, 113–136. [Google Scholar]
- Sengar, R.S.; Gautam, M.; Sengar, R.S.; Garg, S.K.; Senger, K.; Chaudhary, R. Lead stress effects on physiobiochemical activities of higher plants. Rev. Environ. Contam. Toxicol. 2008, 196, 73–93. [Google Scholar]
- Rodriguez, E.; da Conceição Santos, M.; Azevedo, R.; Correia, C.; Moutinho-Pereira, J.; de Oliveira, J.M.P.F.; Dias, M.C. Photosynthesis light-independent reactions are sensitive biomarkers to monitor lead phytotoxicity in a Pb-tolerant Pisum sativum cultivar. Environ. Sci. Pollut. R. 2015, 22, 574–585. [Google Scholar] [CrossRef]
- Kaur, G.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Pb-inhibited mitotic activity in onion roots involves DNA damage and disruption of oxidative metabolism. Ecotoxicology 2014, 23, 1292–1304. [Google Scholar] [CrossRef]
- Patra, M.; Bhowmik, N.; Bandopadhyay, B.; Sharma, A. Comparison of mercury, lead and arsenic with respect to genotoxic effects on plant systems and the development of genetic tolerance. Environ. Exp. Bot. 2004, 52, 199–223. [Google Scholar] [CrossRef]
- Samardakiewicz, S.; Krzeszowiec-Jeleń, W.; Bednarski, W.; Jankowski, A.; Suski, S.; Gabryś, H.; Woźny, A. Pb-induced avoidance-like chloroplast movements in fronds of Lemna trisulca L. PLoS ONE 2015, 10, 0116757. [Google Scholar] [CrossRef]
- Kopittke, P.M.; Asher, C.J.; Kopittke, R.A.; Menzies, N.W. Toxic effects of Pb2+ on growth of cowpea (Vigna unguiculata). Environ. Pollut. 2007, 150, 280–287. [Google Scholar] [CrossRef] [Green Version]
- Piwowarczyk, B.; Tokarz, K.; Muszyńska, E.; Makowski, W.; Jędrzejczyk, R.; Gajewski, Z.; Hanus-Fajerska, E. The acclimatization strategies of kidney vetch (Anthyllis. vulneraria L.) to Pb toxicity. Environ. Sci. Pollut. R. 2018, 25, 19739–19752. [Google Scholar] [CrossRef] [Green Version]
- Nalina, L.; Shanthanu, K.; Rajamani, K.; John Joe, A.; Dhanani, T.; Singh, R.; Kumar, S. Morphological, yield and quality variability among the accessions of Plumbago zeylanica. Electron. J. Plant Breed 2017, 8, 680–687. [Google Scholar] [CrossRef]
- Gawronski, S.W.; Greger, M.; Gawronska, H. Plant taxonomy and metal phytoremediation. In Soil Biology. Detoxification of Heavy Metals; Sherameti, I., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; Volume 30, pp. 91–109. [Google Scholar]
- Saji, A.; Antony, V.T. Plumbago auriculata Lam. Int. J. Pharm. Sci. Rev. Res. 2015, 33, 281–284. [Google Scholar]
- Tyagi, R.I.; Menghani, E. A review on Plumbago zeylanica a compelling herb. Int. J. Pharm. Sci. Rev. Res. 2014, 5, 119–126. [Google Scholar]
- Vogel-Mikuš, K.; Pongrac, P.; Kump, P.; Nečemer, M.; Regvar, M. Colonisation of a Zn, Cd and Pb hyperaccumulator Thlaspi praecox Wulfen with indigenous arbuscular mycorrhizal fungal mixture induces changes in heavy metal and nutrient uptake. Environ. Pollut. 2006, 139, 362–371. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Golemiec, E.; Tokarz, K.; Wielanek, M.; Niewiadomska, E. A dissection of the effects of ethylene, H2O2 and high irradiance on antioxidants and several genes associated with stress and senescence in tobacco leaves. J. Plant Physiol. 2014, 171, 269–275. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta. Physiol. Plant 2016, 38, 102. [Google Scholar] [CrossRef] [Green Version]
- Rozpądek, P.; Nosek, M.; Domka, A.; Ważny, R.; Jędrzejczyk, R.; Tokarz, K.; Pilarska, M.; Niewiadomska, E.; Turnau, K. Acclimation of the photosynthetic apparatus and alterations in sugar metabolism in response to inoculation with endophytic fungi. Plant Cell Environ. 2019, 42, 1408–1423. [Google Scholar] [CrossRef]
- Stirbet, A.; Riznichenko, G.Y.; Rubin, A.B. Modeling chlorophyll a fluorescence transient: Relation to photosynthesis. Biochem. Mosc. 2014, 79, 291–323. [Google Scholar] [CrossRef]
- Marmiroli, M.; Antonioli, G.; Maestri, E.; Marmiroli, N. Evidence of the involvement of plant ligno-cellulosic structure in the sequestration of Pb: An X-ray spectroscopy-based analysis. Environ. Pollut. 2005, 134, 217–227. [Google Scholar] [CrossRef]
- Wierzbicka, M.H.; Przedpełska, E.; Ruzik, R.; Ouerdane, L.; Połec-Pawlak, K.; Jarosz, M.; Szpunar, J.; Szakiel, A. Comparison of the toxicity and distribution of cadmium and lead in plant cells. Protoplasma 2007, 231, 99–111. [Google Scholar] [CrossRef]
- Krzesłowska, M.; Lenartowska, M.; Samardakiewicz, S.; Bilski, H.; Woźny, A. Lead deposited in the cell wall of Funaria hygrometrica protonemata is not stable–a remobilization can occur. Environ. Pollut. 2010, 158, 325–338. [Google Scholar] [CrossRef]
- Seregin, I.V.; Shpigun, L.K.; Ivanov, V.B. Distribution and toxic effects of cadmium and lead on maize roots. Russ. J. Plant Physiol. 2004, 51, 525–533. [Google Scholar] [CrossRef]
- Rascio, N.; Navari-Izzo, F. Heavy metal hyperaccumulating plants: How and why do they do it? And what makes them so interesting? Plant Sci. 2011, 180, 169–181. [Google Scholar] [CrossRef]
- Pourrut, B.; Shahid, M.; Douay, F.; Dumat, C.; Pinelli, E. Molecular mechanisms involved in lead uptake, toxicity and detoxification in higher plants. In Heavy Metal Stress in Plants; Corpas, F.J., Palma, J.M., Gupta, D.K., Eds.; Springer: Berlin, Germany, 2013; pp. 121–147. [Google Scholar]
- Tokarz, K.; Piwowarczyk, B.; Makowski, W. Mechanisms involved in photosynthetic apparatus protection against lead toxicity. In Lead in Plants and the Environment. Radionuclides and Heavy Metals in the Environment; Gupta, D., Chatterjee, S., Walther, C., Eds.; Springer: Cham, Switzerland, 2020; pp. 117–128. [Google Scholar]
- Bhattacharjee, S. The language of reactive oxygen species signaling in plants. J. Bot. 2012, 2012, 985298. [Google Scholar] [CrossRef] [Green Version]
- Cenkci, S.; Cigerci, I.H.; Yildiz, M.; Özay, C.; Bozdag, A.; Terzi, H. Lead contamination reduces chlorophyll biosynthesis and genomic template stability in Brassica rapa L. Environ. Exp. Bot. 2010, 67, 467–473. [Google Scholar] [CrossRef]
- Giuliano, G. Plant carotenoids: Genomics meets multi-gene engineering. Curr. Opin. Plant. Biol. 2014, 19, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Z.; Zhao, F.; Li, M. Lead toxicity in Brassica pekinensis Rupr.: Effect on nitrate assimilation and growth. Environ. Toxicol. 2006, 21, 147–153. [Google Scholar] [CrossRef]
- Rottet, S.; Besagni, C.; Kessler, F. The role of plastoglobules in thylakoid lipid remodeling during plant development. BBA-Bioenergetics 2015, 1847, 889–899. [Google Scholar] [CrossRef] [Green Version]
- Arena, C.; Figlioli, F.; Sorrentino, M.C.; Izzo, L.G.; Capozzi, F.; Giordano, S.; Spagnuolo, V. Ultrastructural, protein and photosynthetic alterations induced by Pb and Cd in Cynara cardunculus L., and its potential for phytoremediation. Ecotox. Environ. Saf. 2017, 145, 83–89. [Google Scholar] [CrossRef]
- Figlioli, F.; Sorrentino, M.C.; Memoli, V.; Arena, C.; Maisto, G.; Giordano, S.; Capozzi, F.; Spagnuolo, V. Overall plant responses to Cd and Pb metal stress in maize: Growth pattern, ultrastructure, and photosynthetic activity. Environ. Sci. Pollut. R 2019, 26, 1781–1790. [Google Scholar] [CrossRef]
- Khan, M.M.; Islam, E.; Irem, S.; Akhtar, K.; Ashraf, M.Y.; Iqbal, J.; Liu, D. Pb-induced phytotoxicity in para grass (Brachiaria mutica) and Castorbean (Ricinus communis L.): Antioxidant and ultrastructural studies. Chemosphere 2018, 200, 257–265. [Google Scholar] [CrossRef]
- Reis, G.S.M.; de Almeida, A.A.F.; de Almeida, N.M.; de Castro, A.V.; Mangabeira, P.A.O.; Pirovani, C.P. Molecular, biochemical and ultrastructural changes induced by Pb toxicity in seedlings of Theobroma cacao L. PLoS ONE 2015, 10, e0129696. [Google Scholar] [CrossRef]
- Shen, J.; Song, L.; Müller, K.; Hu, Y.; Song, Y.; Yu, W.; Wang, H.; Wu, J. Magnesium alleviates adverse effects of lead on growth, photosynthesis, and ultrastructural alterations of Torreya grandis seedlings. Front. Plant Sci. 2016, 7, 1819. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Jiang, Z.; Ma, J.; Yang, L.; Wei, Y. The effects of lead stress on photosynthetic function and chloroplast ultrastructure of Robinia pseudoacacia seedlings. Environ. Sci. Pollut. R. 2017, 24, 10718–10726. [Google Scholar] [CrossRef]
- Piotrowska, A.; Bajguz, A.; Godlewska-Żyłkiewicz, B.; Czerpak, R.; Kamińska, M. Jasmonic acid as modulator of lead toxicity in aquatic plant Wolffia arrhiza (Lemnaceae). Environ. Exp. Bot. 2009, 66, 507–513. [Google Scholar] [CrossRef]
- Dekker, J.P.; Boekema, E.J. Supramolecular organization of thylakoid membrane proteins in green plants. BBA-Bioenergetics 2005, 1706, 12–39. [Google Scholar] [CrossRef] [Green Version]
- Garstka, M. Structural background of photosynthetic light reaction. Adv. Cell Biol. 2007, 34, 445–476. (In Polish) [Google Scholar]
- Gupta, D.K.; Nicoloso, F.T.; Schetinger, M.R.C.; Rossato, L.V.; Pereira, L.B.; Castro, G.Y.; Srivastava, S.; Tripathi, R.D. Antioxidant defense mechanism in hydroponically grown Zea mays seedlings under moderate lead stress. J. Hazard Mater. 2009, 172, 479–484. [Google Scholar] [CrossRef]
- De Las Rivas, J.; Barber, J. Analysis of the structure of the PsbO protein and its implications. Photosynth. Res. 2004, 81, 329–343. [Google Scholar] [CrossRef]
- Sersen, F.; Kralova, K.; Pesko, M.; Cigan, M. Effect of Pb2+ ions on photosynthetic apparatus. Gen. Physiol. Biophys. 2014, 33, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Dao, L.H.; Beardall, J. Effects of lead on two green microalgae Chlorella and cenedesmus: Photosystem II activity and heterogeneity. Algal. Res. 2016, 16, 150–159. [Google Scholar] [CrossRef]
- Janik, E.; Szczepaniuk, J.; Maksymiec, W. Organization and functionality of chlorophyll-protein complexes in thylakoid membranes isolated from Pb-treated Secale cereale. J. Photoch. Photobio. B. 2013, 125, 98–104. [Google Scholar] [CrossRef]
- Harpaz-Saad, S.; Azoulay, T.; Arazi, T.; Ben-Yaakov, E.; Mett, A.; Shiboleth, Y.M.; Hörtensteiner, S.; Gidoni, D.; Gal-On, A.; Goldschmidt, E.E.; et al. Chlorophyllase is a rate-limiting enzyme in chlorophyll catabolism and is posttranslationally regulated. Plant Cell 2007, 19, 1007–1022. [Google Scholar] [CrossRef] [Green Version]
- Romanowska, E.; Wróblewska, B.; Drożak, A.; Zienkiewicz, M.; Siedlecka, M. Effect of Pb ions on superoxide dismutase and catalase activities in leaves of pea plants grown in high and low irradiance. Biol. Plantarum. 2008, 52, 80. [Google Scholar] [CrossRef]
- Qufei, L.; Fashui, H. Effects of Pb2+ on the structure and function of photosystem II of Spirodela polyrrhiza. Biol. Trace. Elem. Res. 2009, 129, 251. [Google Scholar] [CrossRef]
- Force, L.; Critchley, C.; van Rensen, J.J. New fluorescence parameters for monitoring photosynthesis in plants. Photosynth. Res. 2003, 78, 17. [Google Scholar] [CrossRef]
- Belatik, A.; Hotchandani, S.; Carpentier, R. Inhibition of the water oxidizing complex of photosystem II and the reoxidation of the quinone acceptor QA− by Pb2+. PLoS ONE 2013, 8, e68142. [Google Scholar] [CrossRef] [Green Version]
- Doncheva, S.; Ananieva, K.; Stefanov, D.; Vassilev, A.; Gesheva, E.; Dinev, N. Photosynthetic electron transport and antioxidant defense capacity of sunflower plants under combined heavy metal stress. Genet Plant Physiol. 2018, 8, 3–23. [Google Scholar]
- Jiang, H.X.; Chen, L.S.; Zheng, J.G.; Han, S.; Tang, N.; Smith, B.R. Aluminum-induced effects on photosystem II photochemistry in Citrus leaves assessed by the chlorophyll a fluorescence transient. Tree Physiol. 2008, 28, 1863–1871. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Tripathi, R.D.; Dwivedi, S.; Kumar, A.; Trivedi, P.K.; Chakrabarty, D. Lead bioaccumulation potential of an aquatic macrophyte Najas indica are related to antioxidant system. Bioresour. Technol. 2010, 101, 3025–3032. [Google Scholar] [CrossRef] [PubMed]
- McDonald, A.E.; Ivanov, A.G.; Bode, R.; Maxwell, D.P.; Rodermel, S.R.; Hüner, N.P. Flexibility in photosynthetic electron transport: The physiological role of plastoquinol terminal oxidase (PTOX). BBA-Bioenergetics 2011, 1807, 954–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokarz, K.; Makowski, W.; Banasiuk, R.; Krolicka, A.; Piwowarczyk, B. Response of Dionaea muscipula J. Ellis to light stress in in vitro: Physiological study. Plant Cell Tiss. Organ. Cult. 2018, 134, 65–77. [Google Scholar] [CrossRef] [Green Version]
- Tokarz, K.; Piwowarczyk, B.; Wysocka, A.; Wójtowicz, T.; Makowski, W.; Golemiec, E. Response of grass pea (Lathyrus sativus L.) photosynthetic apparatus to short-term intensive UV-A: Red radiation. Acta. Physiol. Plant 2019, 41, 168. [Google Scholar] [CrossRef] [Green Version]
- Krause, G.H.; Jahns, P. Non-photochemical energy dissipation determined by chlorophyll fluorescence quenching: Characterization and function. In Chlorophyll a Fluorescence. Advances in Photosynthesis and Respiration; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; Volume 19, pp. 463–495. [Google Scholar]
- Makowski, W.; Tokarz, B.; Banasiuk, R.; Królicka, A.; Dziurka, M.; Wojciechowska, R.; Tokarz, K.M. Is a blue–red light a good elicitor of phenolic compounds in the family Droseraceae? A comparative study. J. Photochem. Photobiol. B 2019, 201, 111679. [Google Scholar] [CrossRef] [PubMed]
- Klughammer, C.; Schreiber, U. Complementary PS II quantum yields calculated from simple fluorescence parameters measured by PAM fluorometry and the saturation pulse method. PAM Appl. Notes 2008, 1, 201–247. [Google Scholar]
- Huang, W.; Zhang, S.B.; Cao, K.F. Evidence for leaf fold to remedy the deficiency of physiological photoprotection for photosystem II. Photosynth. Res. 2012, 110, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Milward, S.E.; Fan, D.Y.; Chow, W.S.; Badger, M.R. How does cyclic electron flow alleviate photoinhibition in Arabidopsis? Plant Physiol. 2009, 149, 1560–1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laisk, A.; Eichelmann, H.; Oja, V.; Rasulov, B.; Rämma, H. Photosystem II cycle and alternative electron flow in leaves. Plant Cell Physiol. 2006, 47, 972–983. [Google Scholar] [CrossRef] [PubMed]
- Rumeau, D.; Peltier, G.; Cournac, L. Chlororespiration and cyclic electron flow around PSI during photosynthesis and plant stress response. Plant Cell Environ. 2007, 30, 1041–1051. [Google Scholar] [CrossRef] [PubMed]
- Stirbet, A.; Lazár, D.; Guo, Y.; Govindjee, G. Photosynthesis: Basics, history and modelling. Ann. Bot. in press. [CrossRef]
- Tikhonov, A.N. Induction events and short-term regulation of electron transport in chloroplasts: An overview. Photosynth. Res. 2015, 125, 65–94. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Swain, T.; Hillis, W.E. Phenolic constituents of Prunus domestica. I Quantitative analysis of phenolic constituents. J. Sci. Food Agr. 1959, 10, 63–68. [Google Scholar] [CrossRef]
- Fukumoto, L.; Mazza, G. Assessing antioxidant and prooxidant activities of phenolic compounds. J. Agric. Food Chem. 2000, 48, 3597–3604. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Bosa, K.; Kościelniak, J.; Żuk-Gołaszewska, K. Effects of salt stress on photosystem II efficiency and CO2 assimilation of two Syrian barley landraces. Environ. Exp. Bot. 2011, 73, 64–72. [Google Scholar] [CrossRef]
- Laureau, C.; De Paepe, R.; Latouche, G.; Moreno-Chacon, M.; Finazzi, G.; Kuntz, M.; Cornic, G.; Streb, P. Plastid terminal oxidase (PTOX) has the potential to act as a safety valve for excess excitation energy in the alpine plant species Ranunculus glacialis L. Plant Cell Environ. 2013, 36, 1296–1310. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein -dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Miszalski, Z.; Ślesak, I.; Niewiadomska, E.; Baczek-Kwinta, R.; Lüttge, U.; Ratajczak, R. Subcellular localization and stress responses of superoxide dismutase isoforms from leaves in the C3-CAM intermediate halophyte Mesembryanthemum crystallinum L. Plant Cell Environ. 1998, 21, 169–179. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Sharma, U.; Agrawal, V. In vitro shoot regeneration and enhanced synthesis of plumbagin in root callus of Plumbago zeylanica L.—an important medicinal herb. In Vitro Cell. Dev. Plant 2018, 54, 423–435. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pb Concentration [g/l] | Parameters [±SD] | ||||
|---|---|---|---|---|---|
| Growth Index [%] | Shoot Dry Weight Content [%] | Rooting [%] | Root Length [cm] | Root Dry Weight Content [%] | |
| 0.0 (control) | 69.0a ±12.2 | 11.1b ±1.1 | 92.0a ±9.8 | 6.3a ±1.6 | 12.7a ±1.7 |
| 0.05 | 73.5a ±10.3 | 11.2b ±0.9 | 96.0a ±8.9 | 5.0b ±1.0 | 13.3a ±1.2 |
| 0.1 | 71.8a ±13.9 | 12.4a ±0.9 | 74.3b ±19.0 | 4.3c ±1.3 | 12.2a ±1.3 |
| Pb Concentration [g/l] | Organ | Parameters [±SD] | |||||
|---|---|---|---|---|---|---|---|
| MDA* [nM∙g−1DW] | TP [mg∙g−1DW] | CAD [mg∙g−1DW] | FLAV [mg∙g−1DW] | ANT [mg∙g−1DW] | PLUMB [mg∙g−1DW] | ||
| 0.0 (control) | shoot | 180.9b ± 2.4 | 44.6a ± 0.7 | 12.0a ± 0.7 | 11.0a ± 0.7 | 4.4b ± 0.4 | 33.3a ± 3.5 |
| 0.05 | 179.8b ± 5.1 | 41.6b ± 0.7 | 12.4a ± 0.5 | 11.5a ± 0.5 | 3.0a ± 0.2 | 30.4a± 0.5 | |
| 0.1 | 185.7a ± 2.8 | 39.5c ± 2.0 | 11.2a ± 1.3 | 9.6b ± 1.2 | 1.7c ± 0.4 | 28.5a ± 1.0 | |
| 0.0 (control) | root | 261.3a± 16.3 | 27.8b ± 1.5 | 5.1a ± 0.2 | 2.7a ± 0.1 | 1.0a ± 0.1 | 14.7a ± 1.0 |
| 0.05 | 236.7a± 11.5 | 37.5a ± 0.8 | 4.3b ± 0.2 | 2.3b ± 0.2 | 0.7b ± 0.1 | 13.2b ± 0.8 | |
| 0.1 | 251.3a± 5.9 | 43.9a ± 8.1 | 3.7c ± 0.3 | 2.2b ± 0.2 | 0.7b ± 0.1 | 12.1b ± 0.0 | |
| Extracted parameters | Calculated parameters | ||
| FO | Minimum fluorescence, when all PSII reaction centers (RCs) are open | Fv | variable fluorescence; Fm − F0 |
| FM | Maximum fluorescence, when all PSII reaction centers are closed | Fv/Fm | maximum quantum yield of PSII; (Fm − F0)/Fm |
| F50μs, F100μs, F300μs, F2ms, F30 ms | Fluorescence intensities at 50, 100, 300 μs, 2, 30 ms, respectively | Fv/F0 | activity of the water-splitting complex on the donor side of the PSII; (Fm − F0)/F0 |
| Area | Total complementary area between fluorescence induction curve and F = Fm | ||
| OJIP parameters (calculated) | Yields or flux ratios (calculated) | ||
| VJ | Relative variable fluorescence at 2 ms (J-step); VJ = (F2ms − Fo)/(Fm − Fo) | φPo | Maximum quantum yield of primary photochemistry at t = 0; φPo = 1 − Fo/Fm = Fv/Fm |
| VI | Relative variable fluorescence at 30 ms (I-step); VI = (F30ms − Fo)/(Fm − Fo) | φEo | Quantum yield for electron transport at t = 0; φEo = (Fv/Fm)(1 − VJ) |
| Sm | Normalized total complementary area above the OJIP transient (reflecting multiple-turnover QA reduction events) or total electron carriers r RC; Sm = Area/(Fm − Fo) | ψEo | Probability (at time 0) that trapped exciton moves an electron into the electron transport chain beyond; ψEo = 1 − VJ |
| ρRo | Efficiency with which a trapped exciton can move an electron into the electron transport chain from QA‾ to the PSI and electron acceptors; ρRo = ψEoδRo = (1 − VJ)(1 − VI)/(1 − VJ) | ||
| δRo | Efficiency with which an electron can move from the reduced intersystem electron acceptors to the PSI end electron acceptors; δRo = REo/ETo = (1 − VI)/(1 − VJ) | ||
| φRo | Quantum yield for the reduction of end acceptors of PSI per photon absorbed; φRo = REo/ABS = φPoψEoδRo | ||
| Specific fluxes or activities per reaction center (RC) (calculated) | Phenomenological fluxes or activities per excited cross section (CS) (calculated) | ||
| ABS/RC | Absorption flux per RC; ABS/RC = Mo/VJ = 4(F300μs − Fo)/(Fm − Fo)/VJ | TRo/CSo | Trapped energy flux per CS at t = 0; TRo/CSo = (ABS/CSo)φPo |
| TRo/RC | Trapped energy flux per RC at t = 0; TRo/RC = Mo/VJ | ETo/CSo | Electron transport flux per CS at t = 0; ETo/CSo = (ABS/CSo)φEo |
| ETo/RC | Electron transport flux per RC at t = 0; ETo/RC = (Mo/VJ)ψEo | DIo/CSo | Dissipated energy flux per CS at t = 0; DIo/CSo = ABS CSo − TRo/CSo |
| DIo/RC | Dissipated energy flux per RC at t = 0; DIo/RC = ABS/RC − TRo/RC | ||
| Density of reaction centers (calculated) | |||
| RC/CSo | Amount of active PSII RCs per CS at t = 0; RC/CSo = φPo(ABS/CSo)(VJ/Mo) | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tokarz, K.M.; Makowski, W.; Tokarz, B.; Hanula, M.; Sitek, E.; Muszyńska, E.; Jędrzejczyk, R.; Banasiuk, R.; Chajec, Ł.; Mazur, S. Can Ceylon Leadwort (Plumbago zeylanica L.) Acclimate to Lead Toxicity?—Studies of Photosynthetic Apparatus Efficiency. Int. J. Mol. Sci. 2020, 21, 1866. https://doi.org/10.3390/ijms21051866
Tokarz KM, Makowski W, Tokarz B, Hanula M, Sitek E, Muszyńska E, Jędrzejczyk R, Banasiuk R, Chajec Ł, Mazur S. Can Ceylon Leadwort (Plumbago zeylanica L.) Acclimate to Lead Toxicity?—Studies of Photosynthetic Apparatus Efficiency. International Journal of Molecular Sciences. 2020; 21(5):1866. https://doi.org/10.3390/ijms21051866
Chicago/Turabian StyleTokarz, Krzysztof M., Wojciech Makowski, Barbara Tokarz, Monika Hanula, Ewa Sitek, Ewa Muszyńska, Roman Jędrzejczyk, Rafał Banasiuk, Łukasz Chajec, and Stanisław Mazur. 2020. "Can Ceylon Leadwort (Plumbago zeylanica L.) Acclimate to Lead Toxicity?—Studies of Photosynthetic Apparatus Efficiency" International Journal of Molecular Sciences 21, no. 5: 1866. https://doi.org/10.3390/ijms21051866