SLC38A10 (SNAT10) is Located in ER and Golgi Compartments and Has a Role in Regulating Nascent Protein Synthesis


Abstract
:1. Introduction
2. Results
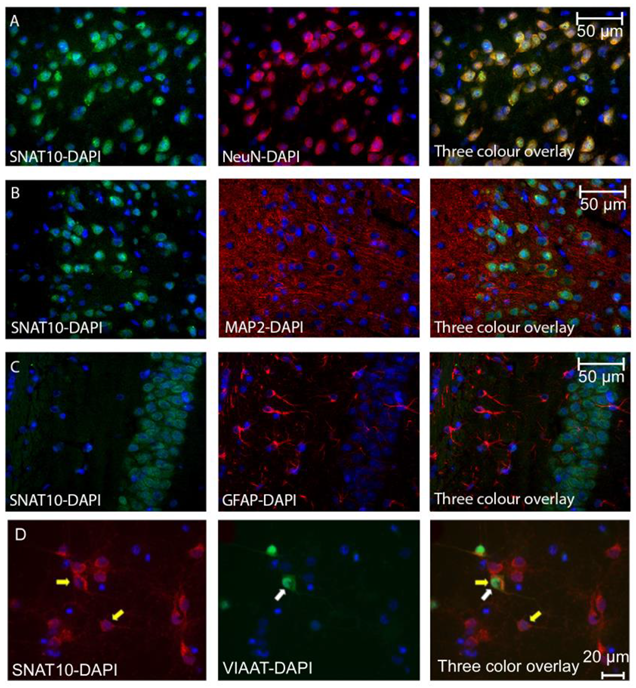
2.1. SNAT10 Is Expressed in Both Excitatory and Inhibitory Neurons
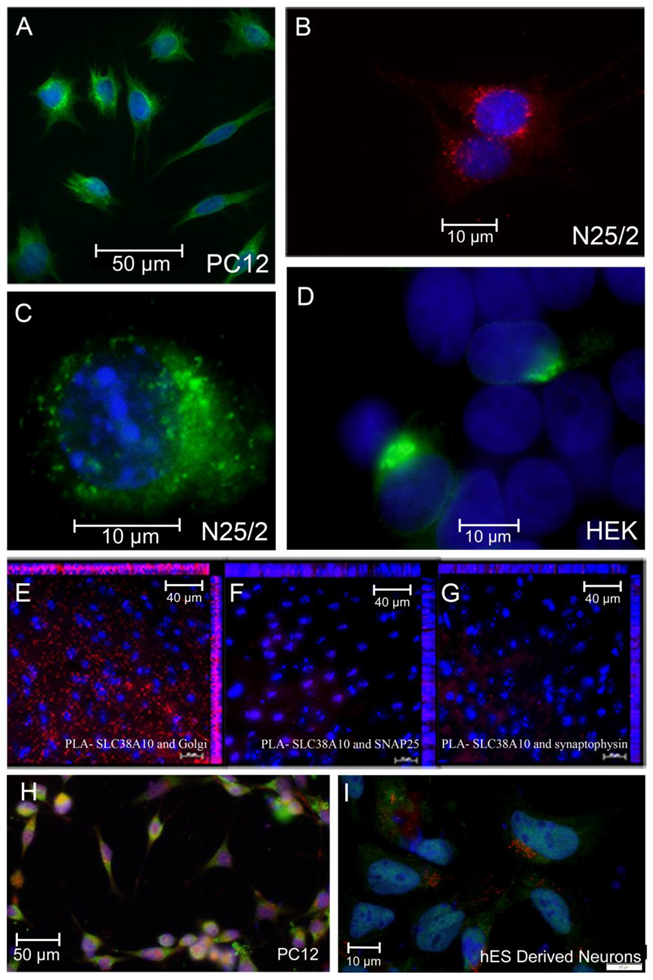
2.2. The Subcellular Localization of SNAT10
2.3. Proximity Ligation Assay Exhibits a High Level of Interaction between SNAT10 and Golgi Proteins
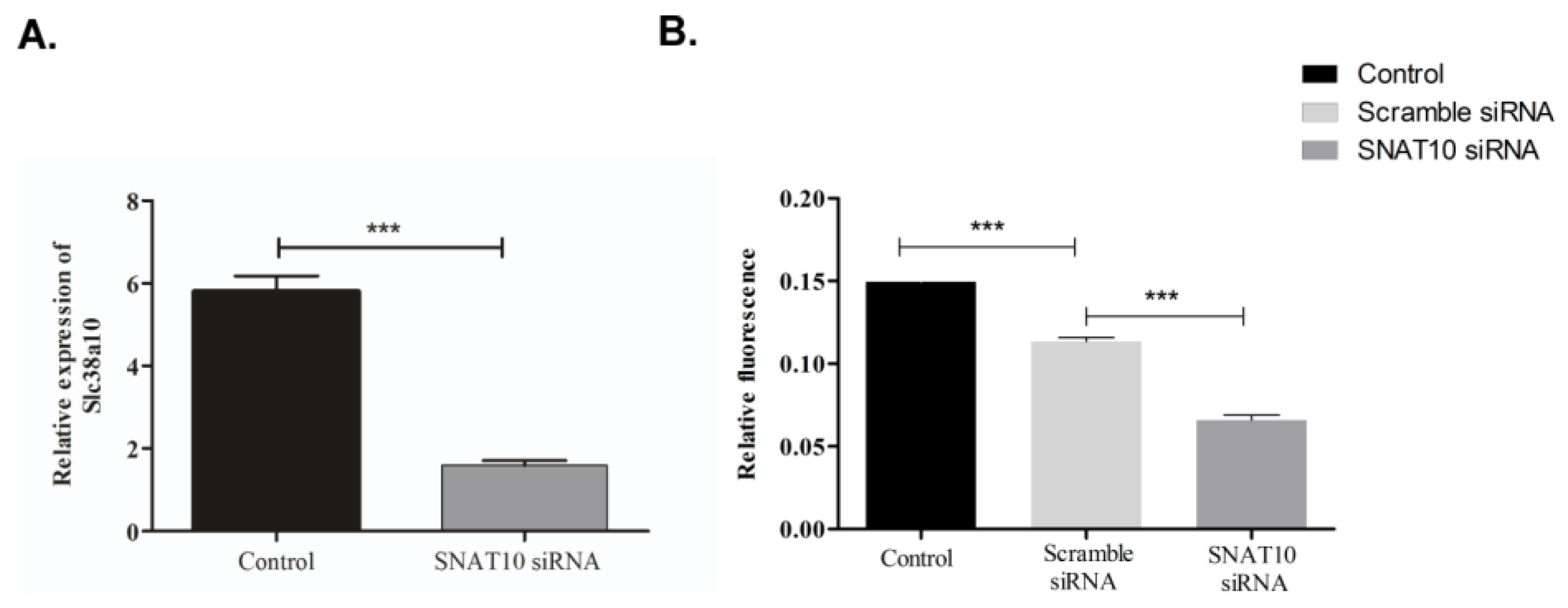
2.4. Knockdown of SNAT10 Affects Protein Synthesis
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. Cell Cultures
- The immortalized rat adrenal gland cell line PC12 Adh (ATCC, CRL-1721.1) was cultured in ATCC-formulated complete growth media F-12K (catalog no. 30-2004) supplemented with 12.5 mL fetal bovine serum (FBS), 75 mL horse serum, 5 mL penicillin-streptomycin (Pen-Strep) to a 500 mL F-12K media flask. All cells were incubated at 37 °C with 5% CO2. The cells were seeded on glass slides (coated with 10 μg/mL poly-L-lycine) for 40 h prior to immuno-staining. All chemicals/media were from Gibco, Stockholm, Sweden, except another company name is stated. Cell passage nine was used for experiments.
- The immortalized embryonic mouse hypothalamus cell line N25/2ATCC (mHypoE-N25/2, Cellutions Biosystems Inc., Burlington, Ontario, Canada) was cultured in Dulbecco’s modified eagle medium (DMEM (+) 4.5g/L D-Glucose, (+) L-glutamine, (+) pyruvate) supplemented with 50 mL fetal bovine serum (FBS), 5 mL penicillin-streptomycin (Pen-Strep). Cell passage eight was used for experiments.
- The immortalized human embryonic kidney cell line (HEK293)ATCC, was cultured in Dulbecco’s modified eagle medium (DMEM [+] 4.5g/L D-Glucose, (+) L-glutamine, (+) pyruvate), supplemented with 50 mL fetal bovine serum (FBS) and 5 mL penicillin-streptomycin (Pen-Strep). Cell passage six was used for experiments.
- Neuronal stem cells were differentiated from human embryonic stem cell H9 line (Wi cell, Madison, Wisconsin, USA) for a duration of seven days using 1:49 ratio of Gibco Neurobasal® medium and Gibco® Neural Induction Supplement. After the cells were differentiated they were harvested and expanded in neural expansion medium. We prepared 50 mL of the medium each time consisting of 24.5 mL of Neurobasal® Medium and Advanced™ DMEM⁄F12 (Thermofisher, Uppsala, Sweden). The cells were seeded on geltrex and splitted 1:5 ratio every 4–5 days. After the second passage post-thaw, they were used for the experiments.
4.3. Construct and Transfection Protocols
4.4. Mouse Primary Cortical Culture
4.5. Fluorescent Immunocytochemistry
4.6. Co-Localization Immunostaining
4.7. Image Analysis for Immunocytochemistry
4.8. Proximity Ligation Assay (PLA)
4.9. Image Analysis for PLA
4.10. Flouorescent Immunohistology
4.11. Knock down of Slc38a10 in PC12 Cells
4.12. Measurement of Protein Synthesis Using OPP Kit
4.13. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Perland, E.; Fredriksson, R. Classification Systems of Secondary Active Transporters. Trends Pharmacol. Sci. 2017, 38, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Cesar-Razquin, A.; Snijder, B.; Frappier-Brinton, T.; Isserlin, R.; Gyimesi, G.; Bai, X.; Reithmeier, R.A.; Hepworth, D.; Hediger, M.A.; Edwards, A.M.; et al. A Call for Systematic Research on Solute Carriers. Cell 2015, 162, 478–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Closs, E.I.; Simon, A.; Vekony, N.; Rotmann, A. Plasma membrane transporters for arginine. J. Nutr. 2004, 134, 2752S–2759S. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, S.; Baomar, H.A.; Al-Walai, S.; Al-Sadi, S.; Fredriksson, R. Histological analysis of SLC38A6 (SNAT6) expression in mouse brain shows selective expression in excitatory neurons with high expression in the synapses. PLoS ONE 2014, 9, e95438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takamori, S. VGLUTs: ‘exciting’ times for glutamatergic research? Neurosci. Res. 2006, 55, 343–351. [Google Scholar] [CrossRef]
- Di Noia, M.A.; Todisco, S.; Cirigliano, A.; Rinaldi, T.; Agrimi, G.; Iacobazzi, V.; Palmieri, F. The human SLC25A33 and SLC25A36 genes of solute carrier family 25 encode two mitochondrial pyrimidine nucleotide transporters. J. Biol. Chem. 2014, 289, 33137–33148. [Google Scholar] [CrossRef] [Green Version]
- Visser, W.F.; van Roermund, C.W.; Waterham, H.R.; Wanders, R.J. Identification of human PMP34 as a peroxisomal ATP transporter. Biochem. Biophys. Res. Commun. 2002, 299, 494–497. [Google Scholar] [CrossRef]
- Rebsamen, M.; Pochini, L.; Stasyk, T.; de Araujo, M.E.; Galluccio, M.; Kandasamy, R.K.; Snijder, B.; Fauster, A.; Rudashevskaya, E.L.; Bruckner, M.; et al. SLC38A9 is a component of the lysosomal amino acid sensing machinery that controls mTORC1. Nature 2015, 519, 477–481. [Google Scholar] [CrossRef]
- Fredriksson, R.; Nordstrom, K.J.; Stephansson, O.; Hagglund, M.G.; Schioth, H.B. The solute carrier (SLC) complement of the human genome: Phylogenetic classification reveals four major families. FEBS Lett. 2008, 582, 3811–3816. [Google Scholar] [CrossRef] [Green Version]
- Mackenzie, B.; Erickson, J.D. Sodium-coupled neutral amino acid (System N/A) transporters of the SLC38 gene family. Pflugers Arch. 2004, 447, 784–795. [Google Scholar] [CrossRef]
- Sundberg, B.E.; Waag, E.; Jacobsson, J.A.; Stephansson, O.; Rumaks, J.; Svirskis, S.; Alsio, J.; Roman, E.; Ebendal, T.; Klusa, V.; et al. The evolutionary history and tissue mapping of amino acid transporters belonging to solute carrier families SLC32, SLC36, and SLC38. J. Mol. Neurosci. 2008, 35, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Schioth, H.B.; Roshanbin, S.; Hagglund, M.G.; Fredriksson, R. Evolutionary origin of amino acid transporter families SLC32, SLC36 and SLC38 and physiological, pathological and therapeutic aspects. Mol. Aspects Med. 2013, 34, 571–585. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Roderick, H.L.; Camacho, P.; Jiang, J.X. Identification and characterization of an amino acid transporter expressed differentially in liver. Proc. Natl. Acad. Sci. USA 2000, 97, 3230–3235. [Google Scholar] [CrossRef] [PubMed]
- Hagglund, M.G.; Hellsten, S.V.; Bagchi, S.; Philippot, G.; Lofqvist, E.; Nilsson, V.C.; Almkvist, I.; Karlsson, E.; Sreedharan, S.; Tafreshiha, A.; et al. Transport of l-Glutamine, l-Alanine, l-Arginine and l-Histidine by the Neuron-Specific Slc38a8 (SNAT8) in CNS. J. Mol. Biol. 2015, 427, 1495–1512. [Google Scholar] [CrossRef]
- Hagglund, M.G.; Sreedharan, S.; Nilsson, V.C.; Shaik, J.H.; Almkvist, I.M.; Backlin, S.; Wrange, O.; Fredriksson, R. Identification of SLC38A7 (SNAT7) protein as a glutamine transporter expressed in neurons. J. Biol. Chem. 2011, 286, 20500–20511. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, T.; Kekuda, R.; Fei, Y.J.; Hatanaka, T.; Sugawara, M.; Martindale, R.G.; Leibach, F.H.; Prasad, P.D.; Ganapathy, V. Cloning and functional characterization of a new subtype of the amino acid transport system N. Am. J. Physiol. Cell Physiol. 2001, 281, C1757–C1768. [Google Scholar] [CrossRef]
- Sugawara, M.; Nakanishi, T.; Fei, Y.J.; Martindale, R.G.; Ganapathy, M.E.; Leibach, F.H.; Ganapathy, V. Structure and function of ATA3, a new subtype of amino acid transport system A, primarily expressed in the liver and skeletal muscle. Biochim. Biophys. Acta 2000, 1509, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Varoqui, H.; Zhu, H.; Yao, D.; Ming, H.; Erickson, J.D. Cloning and functional identification of a neuronal glutamine transporter. J. Biol. Chem. 2000, 275, 4049–4054. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Tsun, Z.Y.; Wolfson, R.L.; Shen, K.; Wyant, G.A.; Plovanich, M.E.; Yuan, E.D.; Jones, T.D.; Chantranupong, L.; Comb, W.; et al. Metabolism. Lysosomal amino acid transporter SLC38A9 signals arginine sufficiency to mTORC1. Science 2015, 347, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Yao, D.; Mackenzie, B.; Ming, H.; Varoqui, H.; Zhu, H.; Hediger, M.A.; Erickson, J.D. A novel system A isoform mediating Na+/neutral amino acid cotransport. J. Biol. Chem. 2000, 275, 22790–22797. [Google Scholar] [CrossRef] [Green Version]
- Perland, E.; Lekholm, E.; Eriksson, M.M.; Bagchi, S.; Arapi, V.; Fredriksson, R. The Putative SLC Transporters Mfsd5 and Mfsd11 Are Abundantly Expressed in the Mouse Brain and Have a Potential Role in Energy Homeostasis. PLoS ONE 2016, 11, e0156912. [Google Scholar] [CrossRef] [PubMed]
- Hatanaka, T.; Hatanaka, Y.; Tsuchida, J.; Ganapathy, V.; Setou, M. Amino acid transporter ATA2 is stored at the trans-Golgi network and released by insulin stimulus in adipocytes. J. Biol. Chem. 2006, 281, 39273–39284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varoqui, H.; Erickson, J.D. Selective up-regulation of system a transporter mRNA in diabetic liver. Biochem. Biophys. Res. Commun. 2002, 290, 903–908. [Google Scholar] [CrossRef] [PubMed]
- Cubelos, B.; Gonzalez-Gonzalez, I.M.; Gimenez, C.; Zafra, F. Amino acid transporter SNAT5 localizes to glial cells in the rat brain. Glia 2005, 49, 230–244. [Google Scholar] [CrossRef] [PubMed]
- Desforges, M.; Lacey, H.A.; Glazier, J.D.; Greenwood, S.L.; Mynett, K.J.; Speake, P.F.; Sibley, C.P. SNAT4 isoform of system A amino acid transporter is expressed in human placenta. Am. J. Physiol. Cell Physiol. 2006, 290, C305–C312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, S.; Villegas, C.J.; Jiang, J.X. Differential regulation of amino acid transporter SNAT3 by insulin in hepatocytes. J. Biol. Chem. 2005, 280, 26055–26062. [Google Scholar] [CrossRef] [Green Version]
- Rubio-Aliaga, I.; Wagner, C.A. Regulation and function of the SLC38A3/SNAT3 glutamine transporter. Channels 2016, 10, 440–452. [Google Scholar] [CrossRef]
- Boulland, J.L.; Osen, K.K.; Levy, L.M.; Danbolt, N.C.; Edwards, R.H.; Storm-Mathisen, J.; Chaudhry, F.A. Cell-specific expression of the glutamine transporter SN1 suggests differences in dependence on the glutamine cycle. Eur. J. Neurosci. 2002, 15, 1615–1631. [Google Scholar] [CrossRef]
- Boulland, J.L.; Rafiki, A.; Levy, L.M.; Storm-Mathisen, J.; Chaudhry, F.A. Highly differential expression of SN1, a bidirectional glutamine transporter, in astroglia and endothelium in the developing rat brain. Glia 2003, 41, 260–275. [Google Scholar] [CrossRef]
- Conti, F.; Melone, M. The glutamine commute: Lost in the tube? Neurochem. Int. 2006, 48, 459–464. [Google Scholar] [CrossRef]
- Mackenzie, B.; Schafer, M.K.; Erickson, J.D.; Hediger, M.A.; Weihe, E.; Varoqui, H. Functional properties and cellular distribution of the system A glutamine transporter SNAT1 support specialized roles in central neurons. J. Biol. Chem. 2003, 278, 23720–23730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amara, S.G.; Fontana, A.C. Excitatory amino acid transporters: Keeping up with glutamate. Neurochem. Int. 2002, 41, 313–318. [Google Scholar] [CrossRef]
- Kilberg, M.S.; Pan, Y.X.; Chen, H.; Leung-Pineda, V. Nutritional control of gene expression: How mammalian cells respond to amino acid limitation. Annu. Rev. Nutr. 2005, 25, 59–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hundal, H.S.; Taylor, P.M. Amino acid transceptors: Gate keepers of nutrient exchange and regulators of nutrient signaling. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E603–E613. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.; Genau, H.M.; Behrends, C. Amino Acid-Dependent mTORC1 Regulation by the Lysosomal Membrane Protein SLC38A9. Mol. Cell. Biol. 2015, 35, 2479–2494. [Google Scholar] [CrossRef] [Green Version]
- Hellsten, S.V.; Hagglund, M.G.; Eriksson, M.M.; Fredriksson, R. The neuronal and astrocytic protein SLC38A10 transports glutamine, glutamate, and aspartate, suggesting a role in neurotransmission. FEBS Open Bio. 2017, 7, 730–746. [Google Scholar] [CrossRef] [Green Version]
- McMahon, H.T.; Bolshakov, V.Y.; Janz, R.; Hammer, R.E.; Siegelbaum, S.A.; Sudhof, T.C. Synaptophysin, a major synaptic vesicle protein, is not essential for neurotransmitter release. Proc. Natl. Acad. Sci. USA 1996, 93, 4760–4764. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Xu, Y.; Stoleru, D.; Salic, A. Imaging protein synthesis in cells and tissues with an alkyne analog of puromycin. Proc. Natl. Acad. Sci. USA 2012, 109, 413. [Google Scholar] [CrossRef] [Green Version]
- Mikros, E.; Diallinas, G. Tales of tails in transporters. Open Biol. 2019, 9, 190083. [Google Scholar] [CrossRef] [Green Version]
- Korfali, N.; Wilkie, G.S.; Swanson, S.K.; Srsen, V.; de Las Heras, J.; Batrakou, D.G.; Malik, P.; Zuleger, N.; Kerr, A.R.; Florens, L.; et al. The nuclear envelope proteome differs notably between tissues. Nucleus 2012, 3, 552–564. [Google Scholar] [CrossRef] [Green Version]
- Lekholm, E.; Perland, E.; Eriksson, M.M.; Hellsten, S.V.; Lindberg, F.A.; Rostami, J.; Fredriksson, R. Putative Membrane-Bound Transporters MFSD14A and MFSD14B Are Neuronal and Affected by Nutrient Availability. Front. Mol. Neurosci. 2017, 10, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McQuin, C.; Goodman, A.; Chernyshev, V.; Kamentsky, L.; Cimini, B.A.; Karhohs, K.W.; Doan, M.; Ding, L.; Rafelski, S.M.; Thirstrup, D.; et al. CellProfiler 3.0: Next-generation image processing for biology. PLoS Biol. 2018, 16, e2005970. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Primary Antibodies | Species | Dilution | Company |
|---|---|---|---|
| Anti-SNAT10 | Rabbit | 1:100 | Innovagen |
| Anti-Synaptophysin | Mouse | 1:250 | BD Transduction lab |
| Anti-SNAP25 | Mouse | 1:500 | Millipore |
| Anti-Golgi 58k | Mouse | 1:50 | AbCam |
| Anti-KDEL | Mouse | 1:200 | Abcam |
| Anti-GFP | Chicken | 1:200 | Millipore |
| Anti-FLAG | Mouse/Rabbit | 1:200 | Invitrogen |
| Secondary antibodies | Species | Dilution | Company |
| Anti-rabbit-594 | Donkey | 1:800 | Invitrogen |
| Anti-mouse-488 | Goat | 1:800 | Invitrogen |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tripathi, R.; Hosseini, K.; Arapi, V.; Fredriksson, R.; Bagchi, S. SLC38A10 (SNAT10) is Located in ER and Golgi Compartments and Has a Role in Regulating Nascent Protein Synthesis. Int. J. Mol. Sci. 2019, 20, 6265. https://doi.org/10.3390/ijms20246265
Tripathi R, Hosseini K, Arapi V, Fredriksson R, Bagchi S. SLC38A10 (SNAT10) is Located in ER and Golgi Compartments and Has a Role in Regulating Nascent Protein Synthesis. International Journal of Molecular Sciences. 2019; 20(24):6265. https://doi.org/10.3390/ijms20246265
Chicago/Turabian StyleTripathi, Rekha, Kimia Hosseini, Vasiliki Arapi, Robert Fredriksson, and Sonchita Bagchi. 2019. "SLC38A10 (SNAT10) is Located in ER and Golgi Compartments and Has a Role in Regulating Nascent Protein Synthesis" International Journal of Molecular Sciences 20, no. 24: 6265. https://doi.org/10.3390/ijms20246265