Cytonemes, Their Formation, Regulation, and Roles in Signaling and Communication in Tumorigenesis


1
Instituto Cajal-CSIC. Av. del Doctor Arce, 37. 28002 Madrid, Spain
2
Department of Biochemistry and Genetics, La Trobe Institute for Molecular Science, La Trobe University, Melbourne, Victoria 3086, Australia
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(22), 5641; https://doi.org/10.3390/ijms20225641
Submission received: 23 September 2019
/
Revised: 8 November 2019
/
Accepted: 9 November 2019
/
Published: 11 November 2019
(This article belongs to the Special Issue Basic and Translational Models of Cooperative Oncogenesis)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Increasing evidence during the past two decades shows that cells interconnect and communicate through cytonemes. These cytoskeleton-driven extensions of specialized membrane territories are involved in cell–cell signaling in development, patterning, and differentiation, but also in the maintenance of adult tissue homeostasis, tissue regeneration, and cancer. Brain tumor cells in glioblastoma extend ultralong membrane protrusions (named tumor microtubes, TMs), which contribute to invasion, proliferation, radioresistance, and tumor progression. Here we review the mechanisms underlying cytoneme formation, regulation, and their roles in cell signaling and communication in epithelial cells and other cell types. Furthermore, we discuss the recent discovery of glial cytonemes in the Drosophila glial cells that alter Wingless (Wg)/Frizzled (Fz) signaling between glia and neurons. Research on cytoneme formation, maintenance, and cell signaling mechanisms will help to better understand not only physiological developmental processes and tissue homeostasis but also cancer progression.
Keywords:
Cytonemes; Drosophila; epithelial cells; Dpp; Hh; EGF; FGF; Wg; glioblastoma; tumourgenesis1. Introduction
Filopodia are long, thin, finger-like, actin-rich plasma-membrane protrusions that function as tentacles for cells to explore their local environment. Cells develop filopodia in response to chemo attractive signals in the microenvironment. These structures are 0.1–0.3 µm in diameter and contain parallel bundles of 10–30 actin filaments held together by actin-binding proteins, including tropomyosin and fascin, and their elongation is mediated by formins [1]. Filopodia dynamics are mainly regulated by the small Rho GTPase, Cdc42 [2]. Filopodia sense the extracellular environment at their tips using cell surface receptors, and they have been given different names usually according to their size or functions: microspikes [3], thin filopodia [4], thick filopodia [5], gliopodia [6], myopodia [7], growth cone filopodia, and dendritic spines involved in synapse formation [8] and in neuronal targeting and pathfinding [9], invadopodia (invasion) [10], podosomes (cell adhesion) [11,12], antigen presentation by dendritic cells of the immune system [13], telopodes [14], pseudopods, tunneling nanotubes [15], and cytonemes [16]. Other related functions include: force generation by macrophages [17], virus transmission [18], vasculogenesis [19], wound closure [20], dorsal closure during Drosophila embryonic development [21], Delta-Notch signaling [22], and growth factor signaling [16].
A cytoneme is defined as specialized types of signaling filopodia that exchange signaling proteins between cells. They were first noted as long cellular extensions that protrude from Drosophila wing imaginal disc cells, and are predominantly linear, with diameters estimated at 100–200 nm and lengths between 2–150 μm [16,23,24,25]. However, filopodia can extend more than 800 μm and have been measured with diameters of 100–500 nm [16,26,27]. Therefore, the macrostructural features contribute to the classification of the various types of filopodia-like structures.
In the vinegar fly, Drosophila melanogaster, cytonemes were initially found in wing and eye imaginal discs [16] and later in ovaries [28], trachea [29,30], and lymph glands [31]. They have also been described in other organisms, such as earthworms [32], earwig ovaries [33], spider embryos [34], in Rhodnius and Calpodes [35], and in several mammalian cell types including retroviral-infected cells [36], mast cells [37], B-lymphocytes [38], and neutrophils [39]. Recent observations suggest that cytonemes also have an important role during development of the zebrafish neural plate [40], where they transport Wnt8a between distant epithelial cells during development in chicken embryos where they mediate Wnt signaling [41,42], and of the chick limb where they transport Sonic Hedgehog (Shh) [25].
Studies in Drosophila have shown differences among cytoneme subtypes. There are cytonemes that send and others that receive signaling proteins. Moreover, cytonemes involved in Decapentaplegic (Dpp), Hedgehog (Hh), Epidermal growth factor (EGF), and Fibroblast growth factor (FGF) signaling can be distinguished by composition, location, and behavior (reviewed in [43]).
Here we will review the physiological role of cytonemes during development in different tissues and the role of cytonemes in tumorigenesis, focusing mainly on the Drosophila model organism.
2. Cytonemes in the Physiology of Epithelial Cells in Drosophila
In this section, we will review and summarize the available information about the structure and composition of cytonemes, available markers, and components required for cytoneme formation. Most of the available literature is based in Drosophila melanogaster. Later, we will review cytoneme physiological roles in cell–cell signaling in both epithelial and in non-epithelial cells.
2.1. Cytonemes: Structure, Composition, and Markers
The cytoneme core is composed of actin filaments that can be marked with actin fluorescent protein chimeras [23,30,44] and with actin-binding fluorescent protein chimeras, such as moesin (GMA:GFP) [23,45,46] and Diaphanous (Dia:GFP [47]). Dia is a member of the formin family [48], which are involved in actin polymerization and that associate with the growing end of actin filaments. Cytonemes can also be observed by using fluorescent tags to mark either the cytoneme membrane (e.g., CD8:GFP, CD8:Cherry, or Cherry-CAAX), or cytoneme components (e.g., signaling protein receptors such as Tkv [24,44], Breathless (Btl) [24,30], Patched (Ptc) [49], and Fibroblast Growth Factor Receptor (FGFR) [50]. Other components include signaling proteins such as Dpp [24] and Hh [23], and the following components of signaling pathways: Ihog, Brother of Ihog (Boi), Shifted (Shf/DmWif), Dallylike (Dlp), Dispatched (Disp) [23,51], Delta (Dl) [22], and cell adhesion proteins, including Neuroglian (Nrg:GFP), and Capricious (Caps:GFP)) [24,44,52]. Fluorescently labeled Flotillin-2 (Flo2/Reggie-1), CD4-Tomato, and glycosylphosphatidyl-inositol (GPI-YFP) also mark cytonemes [23].
Cytonemes can have different components within the same tissue. For instance, the wing disc’s apical cytonemes contain components of the Dpp pathway (Tkv) [44], the basal cytonemes contain components of the Hh pathway (Hh, Ihog, Dally, Dlp, Shf/DmWif, a secreted protein that positively modulates Hedgehog signaling, Disp, and Ptc) [23,49,51,53], and cytonemes in the air sac primordium (ASP) of the wing disc contain either Btl or Tkv, but not both [24]. It is possible that every signaling pathway has a dedicated set of cytonemes that mediate trafficking between specific signaling cells.
The appearance and physical characteristics of cytoneme tips suggest that they are specialized regions. The tips of ASP cytonemes concentrate over-expressed Nrg:GFP, activated Dia:GFP, indicating that cytoneme tips may be sites of actin nucleation, and Caps:GFP, and the only cytonemes that take up Dpp are those whose tips contact wing disc cells [43]. Many wing disc cytonemes had bright bulbous tips at apparent points of contact with ASP cells [52]. The shafts of cytonemes are marked with either membrane-tethered fluorescent proteins or constituent protein fluorescent constructs. They have a uniform diameter, visualized with fluorescence optics, and cytoneme tips are brighter and wider.
2.2. Regulatory Mechanisms in the Formation of Cytonemes
In epithelial tissues, cytonemes emanate from specific membrane territories that have an intrinsic basolateral polarization. Furthermore, in Drosophila, the signaling ligands Hh, Wg, Delta, and Spz, as well as their reception processes show basolateral positioning [22,49,54,55,56]. This subcellular localization implicates mechanisms that drive both signaling components and machinery for the initiation of protrusion to the basolateral side. In the Drosophila wing disc, an apico-basal activity gradient of the RhoGTPase, Rac, regulates filopodial polarization [57,58]. This Rac gradient is regulated by adherens-junction (AJ) proteins and drive both the position and shape of epithelial filopodia. Likewise, a vesicle-sorting mechanism has been described to transport signaling ligands to the basolateral side [59,60]. However, the regulatory mechanisms for cytoneme cargo upload have yet to be determined, and it is also unknown whether or not a vesicle-recycling mechanism could also contribute to cytoneme formation. In zebrafish, Wnt8a at the plasma membrane recruits transducer of CDC42-dependent assembly protein 1 (Toca-1), which locally activates cytoneme nucleation [40,61]. Hence, intracellular trafficking of the Wnt ligand could be key for the spatial localization of membrane protrusion and signaling.
In Drosophila, ectopic expression of a constitutively active form of Dia [62] concentrates at the tips of the ASP cytonemes, and cytonemes do not extend normally in the absence of Dia function [52]. The Rho family member, Vav, localizes to wing disc basal cytonemes [44]. Additionally, the Drosophila capping proteins SCAR and pico, which are actin-binding proteins, have been implicated in cytoneme function by genetic loss-of-function studies [23].
The ASP cells that extend the cytonemes containing receptor-bound Dpp are able to activate Dpp signal transduction. ASP cells that are genetically compromised for Dia and shibire (which encodes a dynamin) and Nrg or Caps (which encodes cell adhesion proteins), fail to make normal cytonemes and are signaling deficient [63]. Moreover, genetic conditions that deplete anterior cells of either Dia or SCAR, reduce the length and number of cytonemes and reduce both the Hh gradient and signaling in the anterior compartment of the wing disc. Furthermore, over-expression of Flotillin-2, a major component of membrane microdomains, increases cytoneme length and the extent of the Hh signaling domain, and it is able to induce numerous filopodia-like protrusions in various cell lines [64].
2.3. Signaling and Communication through Cytonemes
Cytonemes have been described to be associated with components of a specific signaling pathway, even when emerging from the same cells [24]. They can transport either the pathway ligand or the receptor, depending on whether they emanate from receiving or signal-producing cells, or from both. Cytonemes have been shown to be involved in the paracrine transport of the following signaling molecules, including Notch, Spi/EGF, Branchless (Bnl)/FGF, Dpp/BMP, Wingless (Wg)/WNT, and Hh/Shh. Cytoneme-mediated delivery of signaling ligands has been shown for Dpp in the Drosophila wing disc, Wnt in zebrafish [40,65], Hh in Drosophila, and Shh in chick limb bud [23,25,49,55,59]. Conversely, cytonemes emanating from signal-receiving cells have been shown to participate in the distribution of the Drosophila FGF receptor homolog Breathless (Btl) in the developing ASP [66].
Establishing the Dpp gradient in the Drosophila wing disc depends on cytonemes containing the receptor Tkv. These cytonemes extend from cells situated on both sides of the source territory containing Dpp from producer cells [16]. Cells that activate Dpp signal transduction extend cytonemes to the closest cells that produce Dpp. This specificity is evident in the wing disc, where cells in the wing blade primordium direct Tkv-containing cytonemes toward the Dpp-producing cells at the disc midline to which they are closest. Specificity is also evident in the ASP cells that direct Tkv-containing cytonemes toward nearby Dpp-producing cells of the wing disc, while also directing FGFR-containing cytonemes toward wing disc cells that express FGF. Furthermore, cells in the eye disc direct the epidermal growth factor receptor (EGFR)-containing cytonemes toward Spi/EGF-producing cells of the morphogenetic furrow (MF). However, experimental conditions that change the location of the signaling cells also change cytoneme distributions. Changes to the location of cells that express signaling proteins are reflected in altered distributions of cytonemes and blocking signal transduction, either after ectopic expression of a dominant-negative receptor or if expression of the signaling protein is reduced or eliminated, results in the absence of cytonemes [63].
Cytonemes orient uniformly towards the anterior–posterior (A/P) compartment boundary of the Drosophila wing pouch primordium [16]. Cells at the anterior–posterior (A/P) compartment boundary express Dpp (a member of the transforming growth factor-β (TGF-β) superfamily), and long filopodia that extend from wing disc receiving cells towards Dpp-expressing cells at the compartment border are involved in Dpp signaling [16,44]. Different mechanisms were proposed to mediate extracellular molecule distribution among epithelial cells, including extracellular diffusion [67], cell to cell transfer [68], vesicle transport [69], or cytoneme mediated movement [16]. The contribution of cytonemes to Dpp distribution seems to be the most accepted mechanism in the case of the tracheal system. This organization suggested that physical contacts arise at which Dpp transfers to its targets, as an alternative to the diffusion–secretion established model. This mechanism of direct delivery is similar to neurotransmitter release and uptake.
The Hh morphogen is required during development, and Hh signaling has been related to axon guidance, cell migration, stem cell maintenance, and oncogenesis [70]. The role of Hh as a cytoneme based signaling molecule, was originally described in Drosophila wing epithelial cells and in abdominal histoblasts [23]. Actin-based cytonemes are produced and transported by Hh-producing cells to deliver Hh to several cell diameter distances. This study showed that Hh gradient correlates with cytonemes formation, and mutations affecting cytoneme formation also disrupt the Hh gradient [23].
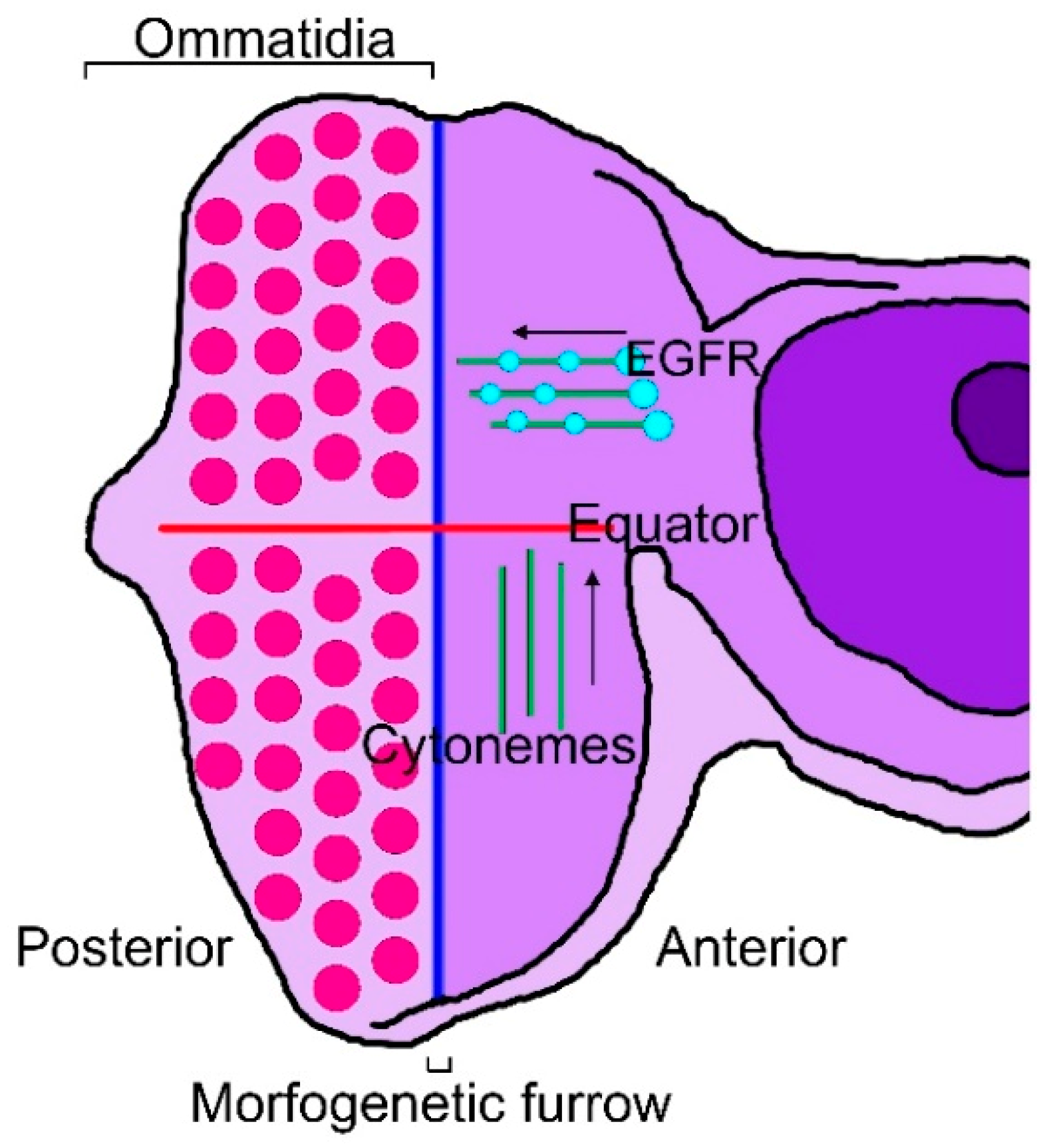
There are cytonemes that are specific to the Drosophila eye and wing discs or to tracheal cells. Eye differentiation in Drosophila is marked by a wave of cell division and differentiation, termed the morphogenetic furrow (MF), which initiates from the posterior margin of the eye imaginal disc just prior to metamorphosis. The MF progresses through the unpatterned dividing cells of the eye disc from posterior to anterior, leaving behind ordered cell clusters called ommatidia [71,72,73]. In the eye disc, cytonemes on the apical surface of columnar epithelial cells orient to either the MF or the equator, which is orthogonal to the MF and defines a line of mirror-image symmetry where dorsal and ventral ommatidia are juxtaposed [74]. EGFR is present in motile puncta in the cytonemes that orient to the MF, where the EGFR ligand, Spi/EGF, is produced [24]. In the wing disc, Thickveins (Tkv), a receptor for Dpp, is present in motile puncta in cytonemes that orient to the disc midline, where Dpp is expressed, and the cytoneme tips contact and appear to be directed only to Dpp-expressing cells, suggesting that they transport Dpp across the disc [44]. In the eye disc, cells extend two types of cytonemes, either orienting toward the MF or toward the equator (Figure 1).
The EGFR signaling pathway sustains multiple functions during eye development, including proliferation and differentiation [75,76,77,78]. In normal conditions, cells extend cytonemes to the usual signaling centers [63]. The EGF receptor concentrates in motile puncta in the MF-directed cytonemes but is not present in the cytonemes directed to the equator. When a dominant-negative EGFR is ectopically expressed throughout the eye disc, the long MF-directed cytonemes are absent, suggesting that they depend on Spi/EGF signaling [24]. Conversely, when the EGF ligand is uniformly over-expressed, the only eye disc cytonemes detected are short and lack a directional bias, suggesting that these cytonemes may have orientations and lengths that are dependent specifically on the source of spi/EGF [24,63]. These characteristics suggest that cytonemes in the eye disc selectively localize EGFR; the MF directed cytonemes mediate EGF signaling; cytonemes appear to link signal producing and receiving cells; and stable contacts via cytonemes require contributions from both signal-producing and signal-receiving cells.
In Drosophila, cytonemes containing the Fz receptor from the ASP cells contact the Wg-producing cell bodies in the wing disc epithelia for signal reception [54]. In the zebrafish embryo, the tips of cytonemes from Wnt8a-producing cells transfer the ligand by contacting the cell body of responding cells [40]. Moreover, cytoneme distributions and plasticity reveal their specificity for a specific signaling protein. Eye disc cytonemes change distribution after uniform overexpression of Spi/EGF and orient towards these ectopic sources, but they do not change after uniform overexpression of Dpp or Hh. Similarly, ASP cytonemes change after ectopic expression of FGF, and wing disc cytonemes change after ectopic expression of Dpp, but these cytonemes do not change after uniform overexpression of Spi/EGF or Hh. These behaviors suggest that stable contacts require contributions from both the signal-producing and signal-receiving cells.
The presence of the Tkv, Btl, and EGF receptor in different, specific cytonemes suggests that each of these cytoneme subtypes mediates the movement of Dpp, FGF, and Spi/EGF, respectively. Cells that activate the signal transduction pathways for these signaling proteins also extend cytonemes that contain the cognate receptor. Similarly, wing disc epithelial cells that activate Dpp signal transduction extend Tkv-containing cytonemes toward Dpp-expressing cells. Likewise, cells of the eye disc activate EGF signal transduction and extend EGFR-containing cytonemes towards the furrow. Moreover, the cells at the tip of the ASP that activate the FGF signal transduction, extend FGFR-containing cytonemes, and cells that activate Dpp signal transduction, extend Tkv-containing cytonemes [63].
Studies in several other systems have reported that signal transduction is associated with cell–cell contacts both for cells that are far apart and for cells separated by short distances. For example, Hh is involved in juxtacrine signaling in the Drosophila germline stem cell niche, and it is localized in cytonemes that extend from Hh-expressing cells [28]. In the Drosophila leg mechanosensory organ, Spi/EGF is produced in a socket cell and induces a specific neighbor to adopt a bract cell fate. Polarized protrusions that originate from the socket cell appear to target EGF signaling to the particular precursor cell [79]. Filopodia-mediated contacts between cells that are not immediate neighbors have also been implicated in Notch and Scabrous-dependent signaling that pattern the bristles of the adult thorax [22,80]. Cytonemes have been shown to link wing disc and ASP cells and are required for signaling. Dpp in transit between the wing disc and the ASP colocalizes with the Tkv receptor in puncta at cytoneme contacts, also known as cytoneme synapses due to their similarities to neuronal synapses [63], and moves along them [52]. Moreover, contacts at cytonemes from myoblasts carrying the Notch ligand, Delta (Dl), also contact the ASP, and vice versa, suggesting a possible cytoneme–cytoneme interaction to activate Notch signaling [54]. In the case of Hh distribution within Drosophila epithelia, a study revealed direct cytoneme–cytoneme contact sites between distant producer and receptor cells all along their length [55]. Similar contacts have been suggested for Shh signaling in the developing chick limb bud [25].
3. Cytonemes in Nonepithelial Tissues: Trachea, Myoblast, Ovary, Brain
Various laboratories described the existence and function of cytonemes in the trachea, myoblast, ovary, and brain cells of Drosophila. The functions of filopodia in neurons are related to path finding [9] and synapse formation [8], chemotaxis in Dictyostelium discoideum [81], cell migration and adhesion [82,83], cell signaling [22,40,52,55,84], and cancer progression and metastasis [83,85,86,87].
3.1. Trachea
The tracheal air sac primordium is a branch that interacts with the epithelial cells of the wing disc. Cytoneme extend from the basal surface of the tracheal epithelium and mediate Dpp signaling through its receptor Tkv [44] or FGF signaling through the FGF-receptor [30]. However, these two receptors are not present in the same cytoneme [52], which suggests a higher level of specificity in cytoneme-mediated signaling.
FGF mediates the budding of air sac precursors and tracheal branching during third instar larvae. These air sac precursors extend cytonemes towards FGF expressing cells and establish filopodial contacts. As a result, FGF signaling induces mitosis on differentiated tracheal cells [30].
A recent publication investigating the similarities between neuronal and cytoneme synapses, studied the roles of neuronal synapses components in the development of the Drosophila ASP. Signaling in the ASP was disrupted if genes associated to glutamate signaling were silenced in wing disc cells, thus cytoneme-mediated signaling in epithelial cells is glutamatergic. In more detail, partial loss-of-function conditions in the wing disc, which targeted essential components of presynaptic neuronal compartments, decreased the presence of cytonemes and signaling in the ASP, but targeting these genes in the ASP had no effect. Partial loss-of-function conditions in the ASP that targeted essential components of postsynaptic neuronal compartments, decreased signaling in the ASP, but targeting these genes in the wing disc had no effect on the ASP. This indicates glutamatergic functions of neuronal presynaptic compartments only in the signal transmitting cells of the wing disc, and glutamatergic functions of neuronal postsynaptic compartments only in the signal-receiving cells of the ASP [88]. Cytoneme contacts are characterized by GRASP fluorescence, a technique that marks sites of approximately 20–40 nm cell–cell apposition originally developed to identify neuronal synapses [52,55]. Cytoneme contacts and cytoneme mediated signaling depend on the adhesion proteins Caps and Nrg, which also have essential trans-synaptic roles in neuronal synapses [89,90]. This study provides evidence for additional components that are common to both cytoneme contacts and neuronal synapses, including voltage-activated glutamate transmission. It was also found that Dpp signaling in the ASP was compromised if disc cells lacked Synaptobrevin and Synaptotagmin-1 (which function in vesicle transport at neuronal synapses), the glutamate transporter, and a voltage-gated calcium channel, or if ASP cells lacked Synaptotagmin-4 or the glutamate receptor GluRII [88].
3.2. Myoblasts
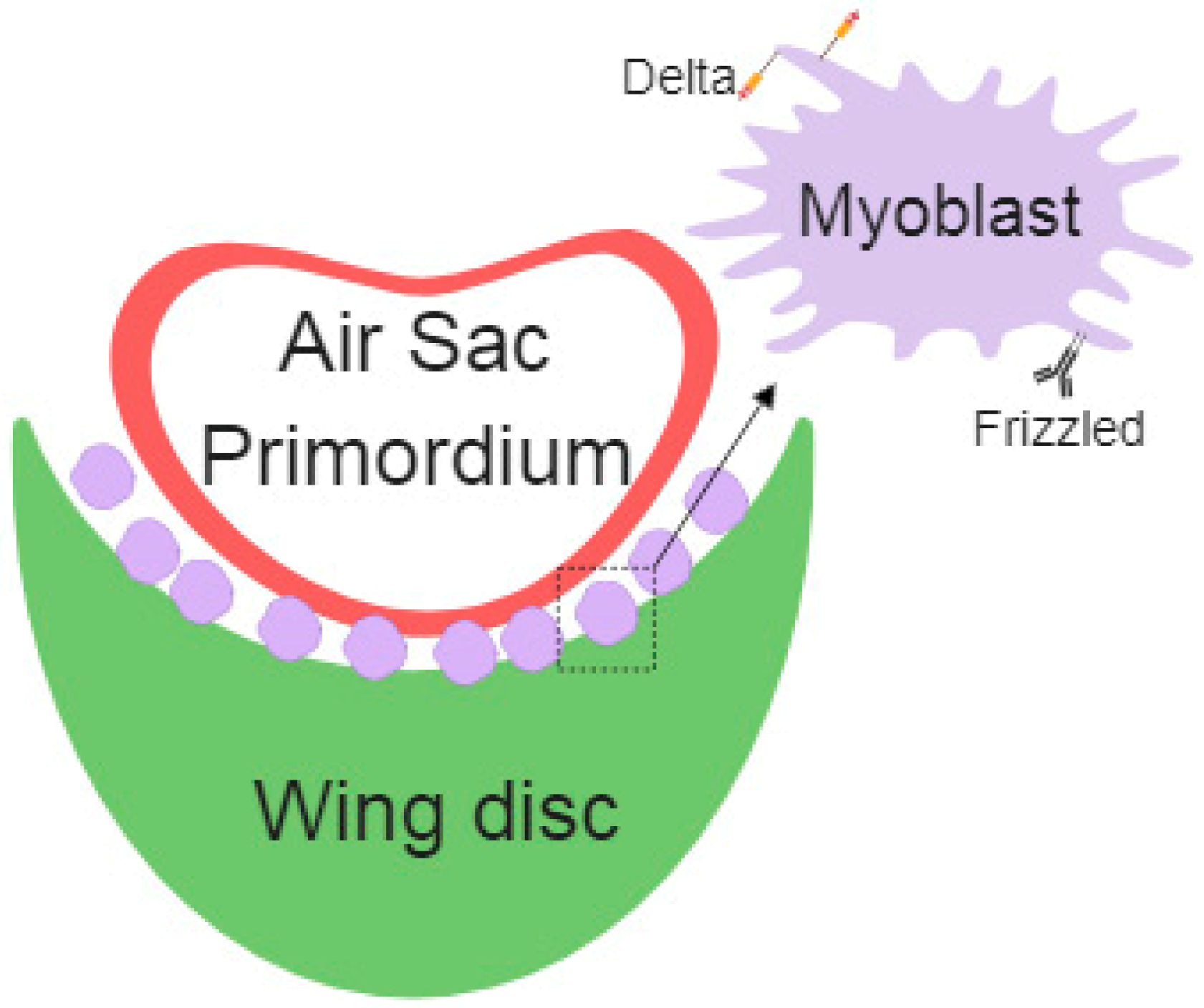
Myoblasts are the precursor cells of the muscle fibers during development; these cells develop together in the wing imaginal disc and later differentiate into the flight muscles. Long-range signals among cells assure the proper regulation of embryonic development. Cytonemes connect myoblasts with epithelial cells and the air sac primordium. Specific cytonemes from myoblast accumulate the Fz Receptor, which takes Wg from epithelial cells. Additionally, myoblast specific Delta (Dl) containing cytonemes contribute to Notch activation in the air sac primordium [54] (Figure 2).
3.3. Ovary/Testis
Stem cell niches maintain the proliferative condition of stem cells, but not of the neighboring daughter cells. Niche signals are limited to the stem cells through microtube-based nanotubes that mediate Dpp signaling. Nanotubes accumulate the Tkv receptor that, after interaction with the ligand Dpp, activates signaling within germ-line stem cells. This signaling (Tkv-Dpp) is sufficient to stimulate nanotube formation. Disruption of this signal leads to germline stem cell loss [91]. Microtube-based nanotubes can transport signaling molecules in a similar way compared to cytonemes, but are microtubule-based and F-actin independent [92].
The maintenance of the stem cell niche in the female germline depends on the Hh gradient and cytoneme formation [28]. In this case, Hh is produced in the cap cells of the ovary niche under the transcriptional regulation of Engrailed. Hh is secreted to the adjacent escort cells and stimulates the expression of Dpp and another Tkv ligand, Glass bottom boat (Gbb), by suppressing Janus kinase signal transducer (JAK/STAT) activity [93]. Whether escort cells are the functional and unique source of Dpp is a matter of controversy. It is also proposed that Dpp is produced by cap cells and functions over a short (one cell diameter) distance to promote GSC self-renewal, by suppressing the expression of the differentiation-promoting factor bags of marbles (Bam) [93]. Cytonemes are produced in the stem cell niche cap cells and mediate Hh transport and delivery to the escort cells. These cytonemes project directionally towards low Hh signaling in the niche even though the signals that modulate cytoneme length from the niche are still under debate. This system ensures the production of Dpp and Gbb to activate Dpp pathway in female germ-line stem cells to assure their proliferative condition [28].
3.4. Neurons
The nervous system cells, neurons, are morphologically distant from other cell types because they extend long protrusions (axons and dendrites) and are polarized and asymmetric. Neurons communicate with target cells or tissues by extending dendrites and axons that form specialized contacts (synapses). These synapses can release or take signals from other cells and ensure the specificity of the communication, and modulate the duration and amplitude of the message [94]. A main characteristic of neurons is the formation of a network that interconnects thousands of cells in the brain. This network facilitates the coordination among different brain regions, contributes to the exchange of molecules (neurotransmitters), and the localization of organelles such as mitochondria in specific zones [95]. These projections are named tunneling nanotubes (TNTs) and share some features with cytonemes [96]. TNTs are formed by F-actin TNTs. In addition, TNTs also contain microtubules, depending on the size and the corresponding delivering cargo. Among others, TNTs transport prion aggregates involved in neurodegenerative diseases and have been postulated as a source of neurodegeneration spreading through different areas of the brain [92].
3.5. Glia
Glial cells (microglia, oligodendrocyte, and astrocytes) are specialized nervous system cells that give metabolic support to the neurons and are responsible for the immune response in the brain, contribute to the spreading of the electric signals, recapture glutamate, and are involved in many other essential functions for the brain physiology [97]. Glial cells play a central role in neural diseases such as Alzheimer’s disease [98], amyotrophic lateral sclerosis (ALS) [99], or psychosis [100].
The coordination between glial cells and neurons is essential for axonal conduction, synaptic transmission, and information processing during development and during adult life; therefore, it is critical for brain function. Different methods of communication have been described in the glia–neuron two-way communication, including ion fluxes, neurotransmitters, cell adhesion molecules, extracellular vesicles, and signaling molecules [101,102]. Thus, glial cells have emerged as central players in the development and function of complex nervous systems from flies to humans [103].
In addition, a novel mechanism of cell to cell communication based on cellular protrusions (filopodia) has been described in glial cells [104]. Astrocytes in vivo extend thin processes around synapses that mediate the communication with neurons. These structures are known as peripheral astrocyte processes (PAPs) and are from 50 to 100 nm thick [104].
4. Cytonemes in Pathology: Tumorigenesis
The nature of these interconnecting structures and their similarities with epithelial cytonemes are currently under debate. Cytonemes have been proposed to mediate communication between neoplastic cells and cells in their microenvironment [87]. In a Drosophila wing disc tumor model utilizing ectopic expression of the EGFR and receptor protein-tyrosine kinase (Ret) oncogenes, cytoneme formation is required to receive signals from the neighboring cells. Genetic ablation of cytonemes prevents tumor progression, restores apico-basal polarity, and improves survival [87]. This recently established system serves as an optimal platform for novel pharmaceutical approaches against cancer progression in vivo. The authors identified pharmacological combinations against cytoneme-mediated oncogenic signals that prevent tumor progression and improve life span. The value of flies as a valid platform for human disease has accumulated evidences that favor this model for future preclinical studies. In particular, the high cost of testing single or combined pharmacological treatments in mice is several orders of magnitude higher than Drosophila based platforms, which has made preclinical trials risky and challenging. The molecular basis underlying cytonemes, the signals transduced by cytonemes, and the implications in tumorigenesis are hot topics for human disease that open novel avenues for potential future treatments.
In addition, the discovery of tunneling nanotubes (TNTs) brings a novel class of thin and long membrane protrusions that connect benign tumor cells [15]. These protrusions form complex networks that mediate the selective transfer of vesicles, organelles, and small molecules [105,106]. TNTs are a common phenomenon in different cell types and tissues that increase under pathological conditions, such as infections, cancer, or neurodegenerative diseases [105]. One considerable limitation to the study of TNTs is the fragility of these structures that makes TNTs difficult to preserve after fixation of tissues. This brought a controversy about their existence in vivo. However, intravital techniques enabled the study of TNTs in live animals [107], which revealed that TNTs are indeed relevant cellular structures in vivo.
In vivo microscopy methods have been used in recent years to study in detail cellular features of cancer cells. A recent study showed that TNTs are induced by stress in prostate cancer and they had a role in mediating intercellular communication that confer stress adaptive cell survival and treatment resistance on the tumoral cells [108]. Additionally, pancreatic cancer cells show TNTs and their formation is stimulated after chemotherapy exposure [109]. Furthermore, TNTs are involved in the communication between tumor cells and macrophages to promote macrophage-dependent tumor cell invasion both in vitro and in an in vivo zebrafish model [110]. Interestingly, colorectal cancer cells have the ability to form locomotory and invasive filopodia that promote invasion and metastasis, and this is suppressed by the phosphorylation of Vasodilator-Stimulated Phosphoprotein (VASP) [111]. Related to colorectal cancer, leucine-rich-repeat containing G-protein-coupled receptor 5 (Lgr5), which labels crypt stem cells, represents the cell of origin in gastrointestinal cancers [112], and Lgr5 promotes the formation of cytonemes in mammalian cells suggesting a possible role for cytonemes in gastrointestinal cancer cell survival, invasion, and metastasis [113]. Exo70, a key component of the Exocyst complex, induces extensive actin membrane protrusions resembling filopodia and blocking Exo70 function inhibits invadopodia formation [114]. Exo70 expression is upregulated in colon cancer samples and its expression is positively correlated with tumor size, invasion depth, and distant metastasis. Colon cancer patients with higher Exo70 expression have a poorer clinical outcome than those with lower Exo70 expression [115].
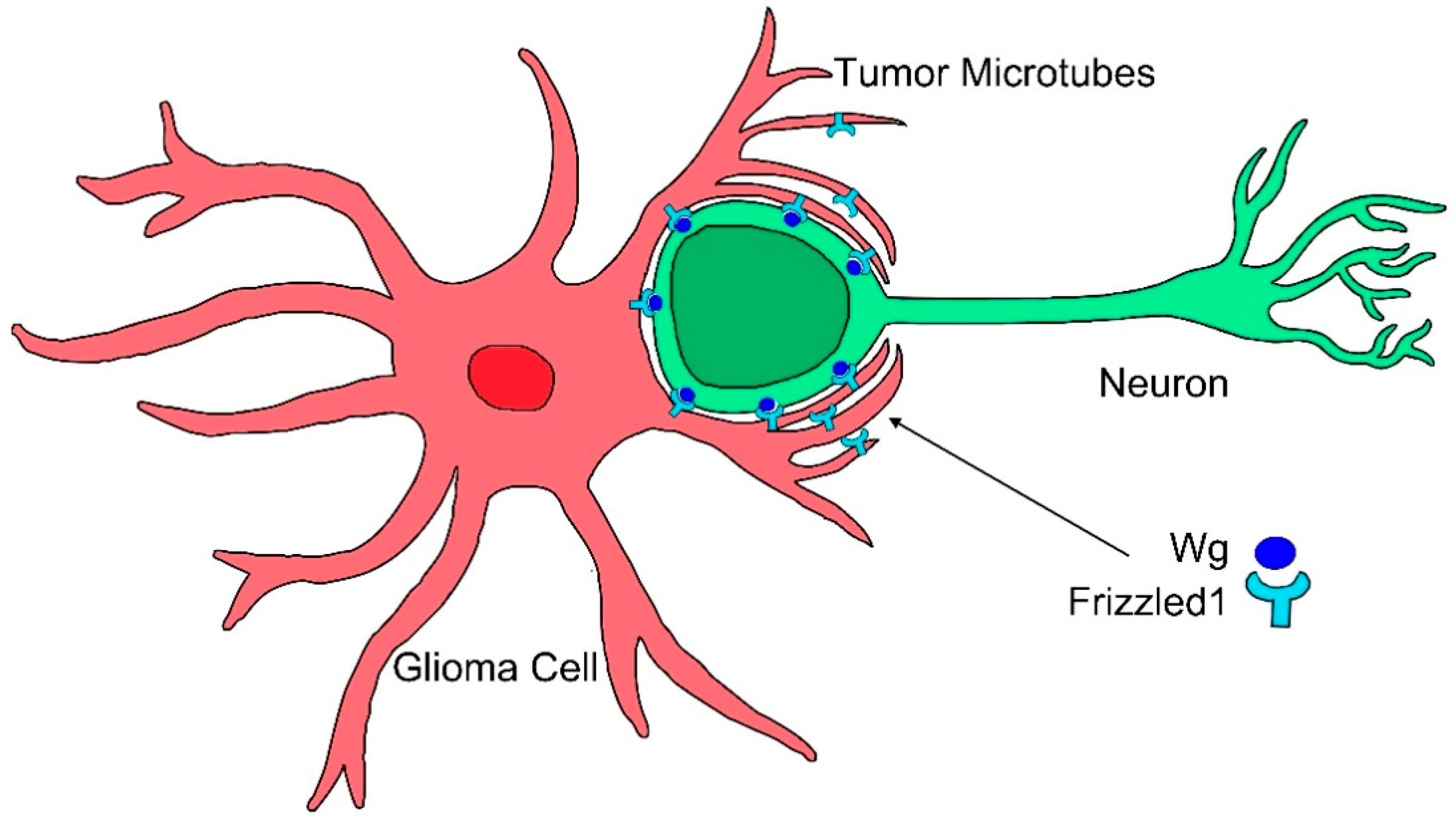
In particular, glioblastoma (GB) cells produce long cellular protrusions at the invasive edge of the tumor that scan the surrounding area and interconnect tumor cells. These protrusions are F-actin based and form a complex network that interconnects GB cells; therefore, they are named tumor microtubes (TMs) [116]. TMs contribute to invasion and proliferation, resulting in effective brain colonization by GB cells. Moreover, TMs constitute a multicellular network that connects GB cells over long distances (up to 500 µm length) [116]. This study found that Growth Associated Protein-43 (GAP43) is essential for the development of TMs and the tumor cell network associated with GB progression, and it drives TM-dependent tumor cell invasion, proliferation, interconnection, and radioresistance. TMs share many characteristics with cytonemes, they are actin-based projections and they are marked by several cytoneme markers, including Ihog, LifeActin, GMA, GPI, myosin light chain (MLC), and the nonmuscle type 2 myosin, spaghetti squash (sqh). Moreover, this study [86] showed in a Drosophila glioma model that the glioma network is dependent on the fly GAP43-like gene (igloo, igl), as has been described in human tumor cells. The glioma network does not develop upon igl silencing. TMs stability in GB is sensitive to GAP43 expression in tumoral cells. Moreover, downregulating Nrg (Nrg-RNAi), which is known to prevent epithelial cytoneme formation, resulted in a reduction of the TM-like processes in GB [86]. Moreover, TMs accumulate Frizzled1 receptor (Fz1) that mediates Wg signaling (Figure 3) [86]. Thus, there are molecular and functional similarities between cytonemes and TMs; however, the term cytoneme is used for physiological situations, and TMs is restricted to the tumoral condition.
TMs and TNTs share some structural features, but TMs are more stable, longer, and thicker (2 µm). In addition, TMs in human cells provide functional coordination to GB cells and facilitate cell repair, brain infiltration, and offer resistance to radiotherapy through dilution of Ca+2 intracellular peaks [116], which thereby increases the aggressiveness of GB.
5. Concluding Remarks
Filopodia are a cellular system of communication widely expanded among living organisms from bacteria to human cells. These protrusions mediate the interaction among cells and with their microenvironment and serve as sensors for the filopodia-emitting cells. Over decades, different forms of filopodia have been described according to their specific function, composition, dimensions, and stability.
Among them, cytonemes have emerged as a novel alternative for cell to cell communication that are involved in development, physiology, and disease. Cytonemes contribute to the directionality of the signals and the specificity of the interaction, as there are emitter and receiver cytonemes for specific signaling pathways. In particular, Wg/WNT, Dpp/BMP, and Hh signaling can be mediated by cytonemes during development and are essential for certain tumoral cell types progression. This feature brings a novel perspective for cancer biology and reveals potential targets for treatment. Therefore, there is a need to decipher the specific mechanisms underlying cytoneme formation, and in general, each type of filopodial protrusion.
Again, the discoveries from animal models, such as Drosophila, provide novel approaches to understand the role of cytonemes in central processes in biology and how they are involved in tumorigenesis. Since cytonemes are conserved structures in other animals, including human cells, the challenge for scientific research in the following years will be to understand the molecular basis of their function in normal physiology and cancer.
Author Contributions
Writing—original draft preparation, S.C.-T. and M.P.; writing—review and editing, S.C.-T. and M.P.; supervision, S.C.-T. and M.P.
Funding
Marta Portela is supported by a NHMRC grant APP116025 (to A/Prof. Helena Richardson).
Acknowledgments
The authors thank Helena Richardson and Paco Martin for comments on the manuscript. The authors apologize to the Drosophila community, if they have inadvertently missed any relevant literature in the writing of this review.
Conflicts of Interest
The authors declare that they have no conflict of interest regarding the publication of this article.
References
- Mellor, H. The role of formins in filopodia formation. Biochim. Biophys. Acta 2010, 1803, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Mattila, P.K.; Lappalainen, P. Filopodia: Molecular architecture and cellular functions. Nat. Rev. Mol. Cell Biol. 2008, 9, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Svitkina, T.M.; Bulanova, E.A.; Chaga, O.Y.; Vignjevic, D.M.; Kojima, S.; Vasiliev, J.M.; Borisy, G.G. Mechanism of filopodia initiation by reorganization of a dendritic network. J. Cell Biol. 2003, 160, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.; Fraser, S.E.; McClay, D. Dynamics of thin filopodia during sea urchin gastrulation. Development 1995, 121, 2501–2511. [Google Scholar] [PubMed]
- McClay, D.R. The role of thin filopodia in motility and morphogenesis. Exp. Cell Res. 1999, 253, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Vasenkova, I.; Luginbuhl, D.; Chiba, A. Gliopodia extend the range of direct glia-neuron communication during the CNS development in Drosophila. Mol. Cell Neurosci. 2006, 31, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Ritzenthaler, S.; Suzuki, E.; Chiba, A. Postsynaptic filopodia in muscle cells interact with innervating motoneuron axons. Nat. Neurosci. 2000, 3, 1012–1017. [Google Scholar] [CrossRef] [PubMed]
- Yuste, R.; Bonhoeffer, T. Genesis of dendritic spines: Insights from ultrastructural and imaging studies. Nat. Rev. Neurosci. 2004, 5, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Bentley, D.; Toroian-Raymond, A. Disoriented pathfinding by pioneer neurone growth cones deprived of filopodia by cytochalasin treatment. Nature 1986, 323, 712–715. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.T. Proteolytic activity of specialized surface protrusions formed at rosette contact sites of transformed cells. J. Exp. Zool. 1989, 251, 167–185. [Google Scholar] [CrossRef] [PubMed]
- Tarone, G.; Cirillo, D.; Giancotti, F.G.; Comoglio, P.M.; Marchisio, P.C. Rous sarcoma virus-transformed fibroblasts adhere primarily at discrete protrusions of the ventral membrane called podosomes. Exp. Cell Res. 1985, 159, 141–157. [Google Scholar] [CrossRef]
- Vasioukhin, V.; Bauer, C.; Yin, M.; Fuchs, E. Directed actin polymerization is the driving force for epithelial cell-cell adhesion. Cell 2000, 100, 209–219. [Google Scholar] [CrossRef]
- Raghunathan, A.; Sivakamasundari, R.; Wolenski, J.; Poddar, R.; Weissman, S.M. Functional analysis of B144/LST1: A gene in the tumor necrosis factor cluster that induces formation of long filopodia in eukaryotic cells. Exp. Cell Res. 2001, 268, 230–244. [Google Scholar] [CrossRef] [PubMed]
- Popescu, L.M.; Faussone-Pellegrini, M.S. TELOCYTES—A case of serendipity: The winding way from Interstitial Cells of Cajal (ICC), via Interstitial Cajal-Like Cells (ICLC) to TELOCYTES. J. Cell Mol. Med. 2010, 14, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Rustom, A.; Saffrich, R.; Markovic, I.; Walther, P.; Gerdes, H.H. Nanotubular highways for intercellular organelle transport. Science 2004, 303, 1007–1010. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Weber, F.A.; Kornberg, T.B. Cytonemes: Cellular processes that project to the principal signaling center in Drosophila imaginal discs. Cell 1999, 97, 599–607. [Google Scholar] [CrossRef]
- Kress, H.; Stelzer, E.H.; Holzer, D.; Buss, F.; Griffiths, G.; Rohrbach, A. Filopodia act as phagocytic tentacles and pull with discrete steps and a load-dependent velocity. Proc. Natl. Acad. Sci. USA 2007, 104, 11633–11638. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, M.J.; Sherer, N.M.; Marks, C.B.; Pypaert, M.; Mothes, W. Actin- and myosin-driven movement of viruses along filopodia precedes their entry into cells. J. Cell Biol. 2005, 170, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Lawson, N.D.; Weinstein, B.M. In vivo imaging of embryonic vascular development using transgenic zebrafish. Dev. Biol. 2002, 248, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Crosson, C.E.; Klyce, S.D.; Beuerman, R.W. Epithelial wound closure in the rabbit cornea. A biphasic process. Investig. Ophthalmol. Vis. Sci. 1986, 27, 464–473. [Google Scholar]
- Jacinto, A.; Wood, W.; Balayo, T.; Turmaine, M.; Martinez-Arias, A.; Martin, P. Dynamic actin-based epithelial adhesion and cell matching during Drosophila dorsal closure. Curr. Biol. 2000, 10, 1420–1426. [Google Scholar] [CrossRef]
- Cohen, M.; Georgiou, M.; Stevenson, N.L.; Miodownik, M.; Baum, B. Dynamic filopodia transmit intermittent Delta-Notch signaling to drive pattern refinement during lateral inhibition. Dev. Cell 2010, 19, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, M.; Gradilla, A.C.; Seijo, I.; Andres, G.; Rodriguez-Navas, C.; Gonzalez-Mendez, L.; Guerrero, I. Cytonemes are required for the establishment of a normal Hedgehog morphogen gradient in Drosophila epithelia. Nat. Cell Biol. 2013, 15, 1269–1281. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Hsiung, F.; Kornberg, T.B. Specificity of Drosophila cytonemes for distinct signaling pathways. Science 2011, 332, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Sanders, T.A.; Llagostera, E.; Barna, M. Specialized filopodia direct long-range transport of SHH during vertebrate tissue patterning. Nature 2013, 497, 628–632. [Google Scholar] [CrossRef] [PubMed]
- Mitchison, T.J.; Cramer, L.P. Actin-based cell motility and cell locomotion. Cell 1996, 84, 371–379. [Google Scholar] [CrossRef]
- Sheetz, M.P.; Wayne, D.B.; Pearlman, A.L. Extension of filopodia by motor-dependent actin assembly. Cell Motil. Cytoskelet. 1992, 22, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Rios, P.; Guerrero, I.; Gonzalez-Reyes, A. Cytoneme-mediated delivery of hedgehog regulates the expression of bone morphogenetic proteins to maintain germline stem cells in Drosophila. PLoS Biol. 2012, 10, e1001298. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, C.; Ebner, A.; Affolter, M. In vivo imaging reveals different cellular functions for FGF and Dpp signaling in tracheal branching morphogenesis. Dev. Cell 2002, 2, 677–683. [Google Scholar] [CrossRef]
- Sato, M.; Kornberg, T.B. FGF is an essential mitogen and chemoattractant for the air sacs of the Drosophila tracheal system. Dev. Cell 2002, 3, 195–207. [Google Scholar] [CrossRef]
- Mandal, L.; Martinez-Agosto, J.A.; Evans, C.J.; Hartenstein, V.; Banerjee, U. A Hedgehog- and Antennapedia-dependent niche maintains Drosophila haematopoietic precursors. Nature 2007, 446, 320–324. [Google Scholar] [CrossRef] [PubMed]
- Kasschau, M.R.; Ngo, T.D.; Sperber, L.M.; Tran, K.L. Formation of filopodia in earthworm (Lumbricus terrestris) coelomocytes in response to osmotic stress. Zoology 2007, 110, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Tworzydlo, W.; Kloc, M.; Bilinski, S.M. Female germline stem cell niches of earwigs are structurally simple and different from those of Drosophila melanogaster. J. Morphol. 2010, 271, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Akiyama-Oda, Y.; Oda, H. Early patterning of the spider embryo: A cluster of mesenchymal cells at the cumulus produces Dpp signals received by germ disc epithelial cells. Development 2003, 130, 1735–1747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locke, M. The very rapid induction of filopodia in insect cells. Tissue Cell 1987, 19, 301–318. [Google Scholar] [CrossRef]
- Sherer, N.M.; Lehmann, M.J.; Jimenez-Soto, L.F.; Horensavitz, C.; Pypaert, M.; Mothes, W. Retroviruses can establish filopodial bridges for efficient cell-to-cell transmission. Nat. Cell Biol. 2007, 9, 310–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fifadara, N.H.; Beer, F.; Ono, S.; Ono, S.J. Interaction between activated chemokine receptor 1 and FcepsilonRI at membrane rafts promotes communication and F-actin-rich cytoneme extensions between mast cells. Int. Immunol. 2010, 22, 113–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, N.; DeFranco, A.L. Visualizing lipid raft dynamics and early signaling events during antigen receptor-mediated B-lymphocyte activation. Mol. Biol. Cell 2003, 14, 432–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galkina, S.I.; Molotkovsky, J.G.; Ullrich, V.; Sud’ina, G.F. Scanning electron microscopy study of neutrophil membrane tubulovesicular extensions (cytonemes) and their role in anchoring, aggregation and phagocytosis. The effect of nitric oxide. Exp. Cell. Res. 2005, 304, 620–629. [Google Scholar] [CrossRef] [PubMed]
- Stanganello, E.; Hagemann, A.I.; Mattes, B.; Sinner, C.; Meyen, D.; Weber, S.; Schug, A.; Raz, E.; Scholpp, S. Filopodia-based Wnt transport during vertebrate tissue patterning. Nat. Commun. 2015, 6, 5846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prols, F.; Wiegreffe, C.; Scaal, M. Communication between distant epithelial cells by filopodia-like protrusions during embryonic development. Development 2015, 142, 665–671. [Google Scholar] [CrossRef] [Green Version]
- Serralbo, O.; Marcelle, C. Migrating cells mediate long-range WNT signaling. Development 2014, 141, 2057–2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornberg, T.B. Cytonemes and the dispersion of morphogens. Wiley Interdiscip. Rev. Dev. Biol. 2014, 3, 445–463. [Google Scholar] [CrossRef] [PubMed]
- Hsiung, F.; Ramirez-Weber, F.A.; Iwaki, D.D.; Kornberg, T.B. Dependence of Drosophila wing imaginal disc cytonemes on Decapentaplegic. Nature 2005, 437, 560–563. [Google Scholar] [CrossRef] [PubMed]
- Cabernard, C.; Affolter, M. Distinct roles for two receptor tyrosine kinases in epithelial branching morphogenesis in Drosophila. Dev. Cell 2005, 9, 831–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, K.A.; Demsky, M.; Montague, R.A.; Weymouth, N.; Kiehart, D.P. GFP-moesin illuminates actin cytoskeleton dynamics in living tissue and demonstrates cell shape changes during morphogenesis in Drosophila. Dev. Biol. 1997, 191, 103–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guha, A.; Kornberg, T.B. Tracheal branch repopulation precedes induction of the Drosophila dorsal air sac primordium. Dev. Biol. 2005, 287, 192–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castrillon, D.H.; Wasserman, S.A. Diaphanous is required for cytokinesis in Drosophila and shares domains of similarity with the products of the limb deformity gene. Development 1994, 120, 3367–3377. [Google Scholar] [PubMed]
- Chen, W.; Huang, H.; Hatori, R.; Kornberg, T.B. Essential basal cytonemes take up Hedgehog in the Drosophila wing imaginal disc. Development 2017, 144, 3134–3144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koizumi, K.; Takano, K.; Kaneyasu, A.; Watanabe-Takano, H.; Tokuda, E.; Abe, T.; Watanabe, N.; Takenawa, T.; Endo, T. RhoD activated by fibroblast growth factor induces cytoneme-like cellular protrusions through mDia3C. Mol. Biol. Cell 2012, 23, 4647–4661. [Google Scholar] [CrossRef] [PubMed]
- Bilioni, A.; Sanchez-Hernandez, D.; Callejo, A.; Gradilla, A.C.; Ibanez, C.; Mollica, E.; Carmen Rodriguez-Navas, M.; Simon, E.; Guerrero, I. Balancing Hedgehog, a retention and release equilibrium given by Dally, Ihog, Boi and shifted/DmWif. Dev. Biol. 2013, 376, 198–212. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Huang, H.; Liu, S.; Kornberg, T.B. Cytoneme-mediated contact-dependent transport of the Drosophila decapentaplegic signaling protein. Science 2014, 343, 1244624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callejo, A.; Bilioni, A.; Mollica, E.; Gorfinkiel, N.; Andres, G.; Ibanez, C.; Torroja, C.; Doglio, L.; Sierra, J.; Guerrero, I. Dispatched mediates Hedgehog basolateral release to form the long-range morphogenetic gradient in the Drosophila wing disk epithelium. Proc. Natl. Acad. Sci. USA 2011, 108, 12591–12598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Kornberg, T.B. Myoblast cytonemes mediate Wg signaling from the wing imaginal disc and Delta-Notch signaling to the air sac primordium. Elife 2015, 4, e06114. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Mendez, L.; Seijo-Barandiaran, I.; Guerrero, I. Cytoneme-mediated cell-cell contacts for Hedgehog reception. Elife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Steinhauer, J.; Liu, H.H.; Miller, E.; Treisman, J.E. Trafficking of the EGFR ligand Spitz regulates its signaling activity in polarized tissues. J. Cell Sci. 2013, 126, 4469–4478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgiou, M.; Baum, B. Polarity proteins and Rho GTPases cooperate to spatially organise epithelial actin-based protrusions. J. Cell Sci. 2010, 123, 1089–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couto, A.; Mack, N.A.; Favia, L.; Georgiou, M. An apicobasal gradient of Rac activity determines protrusion form and position. Nat. Commun. 2017, 8, 15385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gradilla, A.C.; Guerrero, I. Cytoneme-mediated cell-to-cell signaling during development. Cell Tissue Res. 2013, 352, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, I.; Kornberg, T.B. Hedgehog and its circuitous journey from producing to target cells. Semin Cell Dev. Biol. 2014, 33, 52–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, H.Y.; Rohatgi, R.; Lebensohn, A.M.; Le, M.; Li, J.; Gygi, S.P.; Kirschner, M.W. Toca-1 mediates Cdc42-dependent actin nucleation by activating the N-WASP-WIP complex. Cell 2004, 118, 203–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousso, T.; Shewan, A.M.; Mostov, K.E.; Schejter, E.D.; Shilo, B.Z. Apical targeting of the formin Diaphanous in Drosophila tubular epithelia. Elife 2013, 2, e00666. [Google Scholar] [CrossRef] [PubMed]
- Kornberg, T.B.; Roy, S. Cytonemes as specialized signaling filopodia. Development 2014, 141, 729–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumann-Giesen, C.; Falkenbach, B.; Beicht, P.; Claasen, S.; Luers, G.; Stuermer, C.A.; Herzog, V.; Tikkanen, R. Membrane and raft association of reggie-1/flotillin-2: Role of myristoylation, palmitoylation and oligomerization and induction of filopodia by overexpression. Biochem. J. 2004, 378, 509–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattes, B.; Dang, Y.; Greicius, G.; Kaufmann, L.T.; Prunsche, B.; Rosenbauer, J.; Stegmaier, J.; Mikut, R.; Ozbek, S.; Nienhaus, G.U.; et al. Wnt/PCP controls spreading of Wnt/beta-catenin signals by cytonemes in vertebrates. Elife 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Roy, S. Imaging Cytonemes in Drosophila Embryos. Methods Mol. Biol. 2018, 1863, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Strigini, M.; Cohen, S.M. Wingless gradient formation in the Drosophila wing. Curr. Biol. 2000, 10, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Entchev, E.V.; Schwabedissen, A.; Gonzalez-Gaitan, M. Gradient formation of the TGF-beta homolog Dpp. Cell 2000, 103, 981–991. [Google Scholar] [CrossRef] [Green Version]
- Panakova, D.; Sprong, H.; Marois, E.; Thiele, C.; Eaton, S. Lipoprotein particles are required for Hedgehog and Wingless signalling. Nature 2005, 435, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Briscoe, J.; Therond, P.P. The mechanisms of Hedgehog signalling and its roles in development and disease. Nat. Rev. Mol. Cell Biol. 2013, 14, 416–429. [Google Scholar] [CrossRef] [PubMed]
- Heberlein, U.; Moses, K. Mechanisms of Drosophila retinal morphogenesis: The virtues of being progressive. Cell 1995, 81, 987–990. [Google Scholar] [CrossRef] [Green Version]
- Treisman, J.E.; Heberlein, U. Eye development in Drosophila: Formation of the eye field and control of differentiation. Curr. Top. Dev. Biol. 1998, 39, 119–158. [Google Scholar] [PubMed]
- Bonini, N.M.; Choi, K.W. Early decisions in Drosophila eye morphogenesis. Curr. Opin. Genet. Dev. 1995, 5, 507–515. [Google Scholar] [CrossRef]
- Reifegerste, R.; Moses, K. Genetics of epithelial polarity and pattern in the Drosophila retina. Bioessays 1999, 21, 275–285. [Google Scholar] [CrossRef]
- Dominguez, M.; Wasserman, J.D.; Freeman, M. Multiple functions of the EGF receptor in Drosophila eye development. Curr. Biol. 1998, 8, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Rubin, G.M. Analysis of genetic mosaics in developing and adult Drosophila tissues. Development 1993, 117, 1223–1237. [Google Scholar] [PubMed]
- Freeman, M. Reiterative use of the EGF receptor triggers differentiation of all cell types in the Drosophila eye. Cell 1996, 87, 651–660. [Google Scholar] [CrossRef] [Green Version]
- Baker, N.E.; Rubin, G.M. Ellipse mutations in the Drosophila homologue of the EGF receptor affect pattern formation, cell division, and cell death in eye imaginal discs. Dev. Biol. 1992, 150, 381–396. [Google Scholar] [CrossRef]
- Peng, Y.; Han, C.; Axelrod, J.D. Planar polarized protrusions break the symmetry of EGFR signaling during Drosophila bract cell fate induction. Dev. Cell 2012, 23, 507–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renaud, O.; Simpson, P. scabrous modifies epithelial cell adhesion and extends the range of lateral signalling during development of the spaced bristle pattern in Drosophila. Dev. Biol. 2001, 240, 361–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.H.; Chung, C.Y.; Wessels, D.; Stephens, S.; Titus, M.A.; Soll, D.R.; Firtel, R.A. Requirement of a vasodilator-stimulated phosphoprotein family member for cell adhesion, the formation of filopodia, and chemotaxis in dictyostelium. J. Biol. Chem. 2002, 277, 49877–49887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, F.; Janzen, D.M.; Knecht, D.A. Contribution of Filopodia to Cell Migration: A Mechanical Link between Protrusion and Contraction. Int. J. Cell Biol. 2010, 2010, 507821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arjonen, A.; Kaukonen, R.; Ivaska, J. Filopodia and adhesion in cancer cell motility. Cell Adhes. Migr. 2011, 5, 421–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanganello, E.; Scholpp, S. Role of cytonemes in Wnt transport. J. Cell Sci. 2016, 129, 665–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machesky, L.M. Lamellipodia and filopodia in metastasis and invasion. FEBS Lett. 2008, 582, 2102–2111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteban, M.P.; Venkataramani, V.; Fahey-Lozano, N.; Seco, E.; Losada-Perez, M.; Winkler, F.; Casas-Tinto, S. WNT vampirization by glioblastoma leads to tumor growth and neurodegeneration. biorXiv 2019, biorXiv 428953. [Google Scholar] [CrossRef]
- Fereres, S.; Hatori, R.; Hatori, M.; Kornberg, T.B. Cytoneme-mediated signaling essential for tumorigenesis. PLoS Genet. 2019, 15, e1008415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Liu, S.; Kornberg, T.B. Glutamate signaling at cytoneme synapses. Science 2019, 363, 948–955. [Google Scholar] [CrossRef] [PubMed]
- Enneking, E.M.; Kudumala, S.R.; Moreno, E.; Stephan, R.; Boerner, J.; Godenschwege, T.A.; Pielage, J. Transsynaptic coordination of synaptic growth, function, and stability by the L1-type CAM Neuroglian. PLoS Biol. 2013, 11, e1001537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shishido, E.; Takeichi, M.; Nose, A. Drosophila synapse formation: Regulation by transmembrane protein with Leu-rich repeats, CAPRICIOUS. Science 1998, 280, 2118–2121. [Google Scholar] [CrossRef] [PubMed]
- Inaba, M.; Buszczak, M.; Yamashita, Y.M. Nanotubes mediate niche-stem-cell signalling in the Drosophila testis. Nature 2015, 523, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Mattes, B.; Scholpp, S. Emerging role of contact-mediated cell communication in tissue development and diseases. Histochem. Cell Biol. 2018, 150, 431–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Zhong, G.; Chai, P.C.; Luo, L.; Liu, S.; Yang, Y.; Baeg, G.H.; Cai, Y. Coordinated niche-associated signals promote germline homeostasis in the Drosophila ovary. J. Cell Biol. 2015, 211, 469–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornberg, T.B.; Roy, S. Communicating by touch--neurons are not alone. Trends Cell Biol. 2014, 24, 370–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Gerdes, H.H. Transfer of mitochondria via tunneling nanotubes rescues apoptotic PC12 cells. Cell Death Differ. 2015, 22, 1181–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delage, E.; Cervantes, D.C.; Penard, E.; Schmitt, C.; Syan, S.; Disanza, A.; Scita, G.; Zurzolo, C. Differential identity of Filopodia and Tunneling Nanotubes revealed by the opposite functions of actin regulatory complexes. Sci. Rep. 2016, 6, 39632. [Google Scholar] [CrossRef] [PubMed]
- Jakel, S.; Dimou, L. Glial Cells and Their Function in the Adult Brain: A Journey through the History of Their Ablation. Front. Cell Neurosci. 2017, 11, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Nievas, B.G.; Serrano-Pozo, A. Editorial: The Role of Glia in Alzheimer’s Disease. Front. Neurol. 2018, 9, 1161. [Google Scholar] [CrossRef] [PubMed]
- Strohm, L.; Behrends, C. Glia-specific autophagy dysfunction in ALS. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar] [CrossRef]
- Lewandowski, K.E.; Du, F.; Fan, X.; Chen, X.; Huynh, P.; Ongur, D. Role of glia in prefrontal white matter abnormalities in first episode psychosis or mania detected by diffusion tensor spectroscopy. Schizophr. Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Fruhbeis, C.; Frohlich, D.; Kuo, W.P.; Kramer-Albers, E.M. Extracellular vesicles as mediators of neuron-glia communication. Front. Cell Neurosci. 2013, 7, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fields, R.D.; Stevens-Graham, B. New insights into neuron-glia communication. Science 2002, 298, 556–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Losada-Perez, M. Glia: From ‘just glue’ to essential players in complex nervous systems: A comparative view from flies to mammals. J. Neurogenet. 2018, 32, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Aumann, G.; Friedlander, F.; Thummler, M.; Keil, F.; Brunkhorst, R.; Korf, H.W.; Derouiche, A. Quantifying Filopodia in Cultured Astrocytes by an Algorithm. Neurochem. Res. 2017, 42, 1795–1809. [Google Scholar] [CrossRef] [PubMed]
- Ariazi, J.; Benowitz, A.; De Biasi, V.; Den Boer, M.L.; Cherqui, S.; Cui, H.; Douillet, N.; Eugenin, E.A.; Favre, D.; Goodman, S.; et al. Tunneling Nanotubes and Gap Junctions-Their Role in Long-Range Intercellular Communication during Development, Health, and Disease Conditions. Front. Mol. Neurosci. 2017, 10, 333. [Google Scholar] [CrossRef] [PubMed]
- Okafo, G.; Prevedel, L.; Eugenin, E. Tunneling nanotubes (TNT) mediate long-range gap junctional communication: Implications for HIV cell to cell spread. Sci. Rep. 2017, 7, 16660. [Google Scholar] [CrossRef] [PubMed]
- Rehberg, M.; Nekolla, K.; Sellner, S.; Praetner, M.; Mildner, K.; Zeuschner, D.; Krombach, F. Intercellular Transport of Nanomaterials is Mediated by Membrane Nanotubes In Vivo. Small 2016, 12, 1882–1890. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, A.; Zhang, F.; Somasekharan, S.P.; Tse, C.; Leachman, L.; Gleave, A.; Li, B.; Asmaro, I.; Huang, T.; Kotula, L.; et al. Stress-induced tunneling nanotubes support treatment adaptation in prostate cancer. Sci. Rep. 2019, 9, 7826. [Google Scholar] [CrossRef] [PubMed]
- Desir, S.; O’Hare, P.; Vogel, R.I.; Sperduto, W.; Sarkari, A.; Dickson, E.L.; Wong, P.; Nelson, A.C.; Fong, Y.; Steer, C.J.; et al. Chemotherapy-Induced Tunneling Nanotubes Mediate Intercellular Drug Efflux in Pancreatic Cancer. Sci. Rep. 2018, 8, 9484. [Google Scholar] [CrossRef] [PubMed]
- Hanna, S.J.; McCoy-Simandle, K.; Leung, E.; Genna, A.; Condeelis, J.; Cox, D. Tunneling nanotubes, a novel mode of tumor cell-macrophage communication in tumor cell invasion. J. Cell Sci. 2019, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuzga, D.S.; Pelta-Heller, J.; Li, P.; Bombonati, A.; Waldman, S.A.; Pitari, G.M. Phosphorylation of vasodilator-stimulated phosphoprotein Ser239 suppresses filopodia and invadopodia in colon cancer. Int. J. Cancer 2012, 130, 2539–2548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, N.; van Es, J.H.; Kuipers, J.; Kujala, P.; van den Born, M.; Cozijnsen, M.; Haegebarth, A.; Korving, J.; Begthel, H.; Peters, P.J.; et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature 2007, 449, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Snyder, J.C.; Rochelle, L.K.; Marion, S.; Lyerly, H.K.; Barak, L.S.; Caron, M.G. Lgr4 and Lgr5 drive the formation of long actin-rich cytoneme-like membrane protrusions. J. Cell Sci. 2015, 128, 1230–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Zhao, Y.; Sun, Y.; He, B.; Yang, C.; Svitkina, T.; Goldman, Y.E.; Guo, W. Exo70 stimulates the Arp2/3 complex for lamellipodia formation and directional cell migration. Curr. Biol. 2012, 22, 1510–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Zheng, K.; Lv, X.; Hou, J.; Xu, L.; Zhao, Y.; Song, F.; Fan, Y.; Cao, H.; Zhang, W.; et al. Exo70 is an independent prognostic factor in colon cancer. Sci. Rep. 2017, 7, 5039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osswald, M.; Jung, E.; Sahm, F.; Solecki, G.; Venkataramani, V.; Blaes, J.; Weil, S.; Horstmann, H.; Wiestler, B.; Syed, M.; et al. Brain tumour cells interconnect to a functional and resistant network. Nature 2015, 528, 93–98. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Cytonemes of the eye imaginal disc. Diagram of an eye disc in which the MF (blue line) has progressed from posterior to anterior, showing differentiated ommatidia in the posterior region of the eye disc (red dots). The equator is perpendicular to the MF (red line). There are two types of cytoneme (green lines) extending from cells anterior to the MF: (1) cytonemes oriented toward the equator and (2) cytonemes oriented toward the MF and populated with EGFR-containing puncta (light blue dots).
Figure 1.
Cytonemes of the eye imaginal disc. Diagram of an eye disc in which the MF (blue line) has progressed from posterior to anterior, showing differentiated ommatidia in the posterior region of the eye disc (red dots). The equator is perpendicular to the MF (red line). There are two types of cytoneme (green lines) extending from cells anterior to the MF: (1) cytonemes oriented toward the equator and (2) cytonemes oriented toward the MF and populated with EGFR-containing puncta (light blue dots).

Figure 2.
Schematic representation of air sac primordium (ASP), wing disc epithelial cells, and myoblasts cells with specific Delta- or Frizzled-cytonemes.
Figure 2.
Schematic representation of air sac primordium (ASP), wing disc epithelial cells, and myoblasts cells with specific Delta- or Frizzled-cytonemes.

Figure 3.
Cytonemes in tumorigenesis. Glioma cells produce a network of tumor microtubes that grow to reach and enwrap neighboring neurons. Increased glia-neuron membrane contacts facilitate neuronal Wg sequestering mediated by glioma Frizzled1 receptor accumulated in the tumor microtubes.
Figure 3.
Cytonemes in tumorigenesis. Glioma cells produce a network of tumor microtubes that grow to reach and enwrap neighboring neurons. Increased glia-neuron membrane contacts facilitate neuronal Wg sequestering mediated by glioma Frizzled1 receptor accumulated in the tumor microtubes.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Casas-Tintó, S.; Portela, M. Cytonemes, Their Formation, Regulation, and Roles in Signaling and Communication in Tumorigenesis. Int. J. Mol. Sci. 2019, 20, 5641. https://doi.org/10.3390/ijms20225641
AMA Style
Casas-Tintó S, Portela M. Cytonemes, Their Formation, Regulation, and Roles in Signaling and Communication in Tumorigenesis. International Journal of Molecular Sciences. 2019; 20(22):5641. https://doi.org/10.3390/ijms20225641
Chicago/Turabian StyleCasas-Tintó, Sergio, and Marta Portela. 2019. "Cytonemes, Their Formation, Regulation, and Roles in Signaling and Communication in Tumorigenesis" International Journal of Molecular Sciences 20, no. 22: 5641. https://doi.org/10.3390/ijms20225641
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.