Conflicting Roles of 20-HETE in Hypertension and Stroke
by
 , , and
, , and
Shashank Shekhar
1,† ,
,
Kevin Varghese
2,†,
Man Li
2,
Letao Fan
2,
George W. Booz
2,
Richard J. Roman
2 and
Fan Fan
2,* 1
Department of Neurology, University of Mississippi Medical Center, Jackson, MS 39216, USA
2
Department of Pharmacology and Toxicology, University of Mississippi Medical Center, Jackson, MS 39216, USA
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Mol. Sci. 2019, 20(18), 4500; https://doi.org/10.3390/ijms20184500
Submission received: 17 July 2019
/
Revised: 28 August 2019
/
Accepted: 8 September 2019
/
Published: 11 September 2019
(This article belongs to the Special Issue Molecular Advances in Hypertension)
{kind=link}
{kind=link}
Abstract
:Hypertension is the most common modifiable risk factor for stroke, and understanding the underlying mechanisms of hypertension and hypertension-related stroke is crucial. 20-hydroxy-5, 8, 11, 14-eicosatetraenoic acid (20-HETE), which plays an important role in vasoconstriction, autoregulation, endothelial dysfunction, angiogenesis, inflammation, and blood-brain barrier integrity, has been linked to hypertension and stroke. 20-HETE can promote hypertension by potentiating the vascular response to vasoconstrictors; it also can reduce blood pressure by inhibition of sodium transport in the kidney. The production of 20-HETE is elevated after the onset of both ischemic and hemorrhagic strokes; on the other hand, subjects with genetic variants in CYP4F2 and CYP4A11 that reduce 20-HETE production are more susceptible to stroke. This review summarizes recent genetic variants in CYP4F2, and CYP4A11 influencing 20-HETE production and discusses the role of 20-HETE in hypertension and the susceptibility to the onset, progression, and prognosis of ischemic and hemorrhagic strokes.
1. Introduction
Hypertension is a prevalent chronic disease that affects approximately 75 million people in the U.S. and an estimated cost of $48.6 billion annually [1]. Unlike hypertension, which takes years to manifest clinically; stroke presents with sudden and devastating neurological consequences. Stroke is one of the major risk factors of mortality and causes more survivors with long-term disability than other risk factors [2]. Hypertension is the most common modifiable risk factor of stroke. It acts through various mechanisms affecting vascular smooth muscle cells (VSMCs), endothelial dysfunction, oxidative stress, changes in cerebral blood flow, and inflammation [3,4].
Numerous animal and clinical studies have indicated the importance of genetic and environmental factors to the pathogenesis of stroke. A genome-wide association study (GWAS) demonstrated that genetic variation in the Notch3 receptor is linked to cerebral autosomal dominant arteriopathy with subcortical infarcts and leukoencephalopathy (CADASIL), or cerebral autosomal-recessive arteriopathy with subcortical infarcts and leukoencephalopathy (CARASIL) [5]. Single nucleotide polymorphisms (SNPs) or mutations have been linked to increased risk of ischemic (i.e., COL3A1, GLA) or hemorrhagic stroke (i.e., APP, COL4A1) [6]. Many of these same SNPs also are associated with hypertension. For example, a recent clinical study with 18,000 ischemic stroke patients and 70,000 controls identified SNPs in SH2B3 and ALDH2 linked to ischemic stroke, which were also associated with blood pressure [6]. Variants in CYP4F2 and CYP4A11 have been repeatedly linked to hypertension, stroke, and cardiovascular disease [7,8,9,10,11].
CYP4F2 and CYP4A11 are the two most potent enzymes to catalyze the metabolism of arachidonic acid (AA) to 20-hydroxy-5, 8, 11, 14-eicosatetraenoic acid (20-HETE) in humans [12]. A number of SNPs in CYP4F2 and CYP4A11 have been reported to play an essential role in the development of hypertension and stroke in several populations, as summarized in Figure 1. These include rs2108622 (V433M) and rs1558139 in CYP4F2 in Japanese, Australian, Swedish, Chinese, South Indian, and American subjects [11,13,14,15,16,17,18,19] and rs1126742 (F434S), rs9333025, and rs389011 in CYP4A11 in Chinese, American, Japanese, and African subjects [11,20,21,22,23,24].
Among these SNPs, rs2108622 (V433M, CYP4F2) was confirmed to reduce 20-HETE production in vitro [25,26], and increase urinary 20-HETE excretion [14], and rs1126742 (F434S, CYP4A11) was first reported to reduce 20-HETE synthase activity [27]. Saito et al, reported that five (rs12564525, rs2056900, rs2056899, rs4926581, and rs2405599) of 10 variants in CYP4A11 or CYP4A22 identified in the Pharmacogene Variation Consortium (https://www.pharmvar.org/) markedly reduced activity when expressed in COS-7 cells [28]. However, the functional significance of these observations has not been identified. In animal studies, Dahl salt-sensitive (SS) rats rapidly develop severe hypertension when fed a high salt diet. This strain has a deficiency in the expression of CYP4A enzymes and the formation of 20-HETE. They displayed cerebral vascular dysfunction and enhanced blood–brain barrier (BBB) leakage, which may increase the susceptibility to the development of stroke [29,30]. The available evidence seems to suggest that reduced 20-HETE levels may predispose to the development of stroke, although alterations in 20-HETE production associated with many of the human CYP4F2 and CYP4A11 SNPs have not been defined. On the other hand, numerous studies in human and animal models indicate that 20-HETE production is enhanced following traumatic brain injury (TBI) and ischemic and hemorrhagic stroke [31,32,33]. Elevations in 20-HETE contribute to an acute fall in cerebral blood flow (CBF) and delayed vasospasm following subarachnoid hemorrhage (SAH). Inhibitors of the formation of 20-HETE reduce infarct size following ischemic stroke and reverse vasospasm following SAH [7,33]. Similarly, inhibition of 20-HETE prevents BBB leakage, reduces cerebral edema, and is beneficial to the outcomes following TBI [31,32]. Ischemic stroke patients who have both rs9333025 GG (CYP4A11) and rs2108622 GG (CYP4F2) variants have higher plasma 20-HETE levels [34]. Spontaneously hypertensive rats (SHR) that have elevated endogenous 20-HETE production are more susceptible to ischemic stroke. Elevated 20-HETE levels are also associated with worse prognosis in SAH patients [17,33]. This evidence suggests that elevated 20-HETE participates in the severity of ischemic and hemorrhagic strokes.
Indeed, the link between 20-HETE and hypertension is complicated as 20-HETE promotes the effects of vasoconstrictors to increase blood pressure in some models but reduces blood pressure by inhibition of sodium transport in the kidney in other models [12,33]. The complicated relationship between 20-HETE and stroke is thus expected, not only because of the conflicting roles of 20-HETE, but also because of the complexity of the onset, development, and prognosis in different types of stroke. This review addresses the conflicting roles of 20-HETE in hypertension and stroke by dissecting the molecular mechanisms of 20-HETE and its contribution at various stages of ischemic and hemorrhagic stroke.
2. 20-HETE in Hypertension
2.1. 20-HETE
20-HETE is an AA metabolite catalyzed by enzymes of the CYP4A and CYP4F families. In humans, CYP4A11, CYP4A22, CYP4F2, and CYP4F3 are responsible for generating 20-HETE through ω-hydroxylation. CYP4F2 and CYP4A11 are the two most potent isoforms. [35]. The major isoforms of these enzymes in rats, CYP4A1, CYP4A2, CYP4A3, CYP4A8, CYP4F1, and CYP4F4 are found in the liver, kidney, brain, blood vessels, and other tissues. Among these, CYPA1 is the most active isoform in the production of 20-HETE [7,33]. In mice, there are several isoforms of CYP4A expressed, including CYP4A10, CYP4A12A, CYP4A12B, and CYP4A14, but only CYP12A can produce 20-HETE [12]. 20-HETE can be inactivated by cyclooxygenase-2 (COX2) after conversion to 20-hydroxy-PGE. It can also be metabolized and inactivated by alcohol dehydrogenase (ADH) to 20-carboxy-eicosatetraenoic acid (20-COOH-HETE) which can be further converted to the 18-dicarboxylic acid and 16-dicarboxylic acid through beta-oxidation. The actions of 20-HETE can also be terminated by metabolism to a glucuronide by uridine 5’-diphosphoglucuronosyltransferase (UGT) [7].
20-HETE exhibits a wide variety of functions essential to the physiology and pathogenesis of many diseases. It acts as a natriuretic agent in the kidney, serves as an autocrine factor, and potentiates the effects of vasoconstrictive agents. It also promotes renal oxidative stress, endothelial dysfunction, vascular inflammation, and activates the renin-angiotensin-aldosterone system (RAAS), especially in the vascular endothelium [12]. Moreover, 20-HETE plays a role in angiogenesis, cell proliferation, vascular remodeling, and restenosis [33,36]. In addition, by suppressing thromboxane formation, 20-HETE inhibits platelet aggregation [37]. Administration of angiotensin II (ANG II) stimulates phospholipase A2 (PLA2) and releases AA resulting in enhanced 20-HETE production [38]. Induction of heme oxygenase (HO) reduces, and administration of dihydrotestosterone (DHT) elevates, 20-HETE production in the kidney and vasculature [12]. Carbon monoxide (CO) and nitric oxide (NO) act as inhibitors of 20-HETE [8].
Two 20-HETE receptors have been discovered recently. A chemokine CCL5 (RANTES) receptor, G-protein-coupled receptor GPR75 (Gq), was identified to bind to 20-HETE leading to endothelial dysfunction [39]. We reported that GPR75 is highly expressed in the brain and cerebral vasculature at protein levels in rats [40], and it is highly expressed at mRNA levels in the brain in humans (Human Protein Atlas: https://www.proteinatlas.org/ENSG00000119737-GPR75/tissue); however, the exact cellular localization and roles of GPR75 in the brain are yet to be determined [41]. It is interesting that CCL5 is highly expressed in the cerebral vasculature in Alzheimer’s patients [42], which prevents amyloid β-induced neuronal cell death via activation of GPR75 [43]. High doses of 20-HETE also activate GPR40 in the pancreatic islet cells to regulate insulin secretion [44], but epoxyeicosatrienoic acids (EETs) activate GPR40 in endothelial, smooth muscle cells and arteries resulting in vasodilation [45]. Nevertheless, whether GPR75 and GPR40 play a role in the vascular effects of 20-HETE in stroke in unknown.
2.2. 20-HETE in Hypertension
20-HETE has both pro- and anti-hypertensive actions. Here, we mainly discuss the conflicting roles of 20-HETE in the kidney and vasculature in the control of blood pressure.
2.2.1. 20-HETE and the Kidney
20-HETE reduces sodium transport by inhibition of Na+/K+-ATPase activity and internalization of the sodium–hydrogen exchanger 3 (NHE3) in the proximal tubule (PT). It also blocks Na+/K+-ATPase, and potassium channels and limits sodium and potassium uptake via the Na–K–Cl cotransporter (NKCC2) in the thick ascending loop of Henle (TALH) [8,46]. 20-HETE also interacts with ANG II, dopamine, endothelin, and parathyroid hormone, and contributes to their natriuretic effects in the PT. The production of 20-HETE was elevated when renal perfusion pressure was increased, indicating its role in pressure natriuretic response [8].
2.2.2. 20-HETE and the Vasculature
In the vasculature, 20-HETE constricts VSMC and contributes to the responses to stretch and vasoconstrictors such as ANG II, serotonin (5-HT), and endothelin by blocking calcium-activated potassium (BK) channel activity and enhancing calcium influx through voltage-gated L-type calcium channels. It promotes cell proliferation and endothelial dysfunction and is associated with vascular inflammation [7]. Moreover, a 20-HETE receptor, GPR75, has been linked to endothelial dysfunction [39]. 20-HETE augments constriction of VSMCs by enhancing calcium entry via activation of mitogen-activated protein kinases (MAPKs), protein kinase C (PKC), Rho-associated protein kinase (ROCK), and tyrosine-kinases [33]. 20-HETE enhances the myogenic reactivity and autoregulation of CBF and renal blood flow [7,29]. In the kidney, 20-HETE increases constriction of the afferent arteriole (Af-art) in response to elevations in perfusion pressure [47]. Blockade of 20-HETE formation diminished the vasoconstrictor response to both elevations in transmural pressure and is thought to be the mediator of tubuloglomerular feedback responses (TGF) [47]. Indeed, inhibitors of 20-HETE have been shown to block TGF-mediated constriction of the Af-art of isolated perfused glomeruli with an intact macula densa to elevations in sodium concentration in the tubular perfusate. 20-HETE inhibitors also blocked TGF in vivo [7,47].
20-HETE has been reported to increase the expression of angiotensin-converting enzyme (ACE) in the endothelium. Upregulation of the formation of 20-HETE in the vasculature in response to administration of androgens or Cyp4a12 transgenic and Cyp4a14 knockout (KO) mice produces 20-HETE dependent hypertension that is mediated in part through increases in the expression of ACE in the vascular endothelium. Indeed, 20-HETE induced hypertension and vascular hypertrophy can be blocked by administration of ANG II receptor type 1 (AT1) receptor antagonists [7].
Previous studies have indicated that the expression of CYP4A enzymes and the production of 20-HETE in the kidney and vasculature are reduced in SS rats [29,30]. SNPs in Cyp4a on chromosome 5 cosegregate with salt-sensitive hypertension in an F2 cross of SS and Lewis rats [48]. Moreover, a deficiency in 20-HETE is thought to reset the pressure-natriuretic relationship and enhance sodium transport in the PT and TALH. Induction of the formation of 20-HETE with fibrates or transfer of wild type (WT) Cyp4a genes in congenic strains improves sodium excretion and reduces blood pressure [8,49]. In contrast, the expression of CYP4A and production of 20-HETE are elevated in the vasculature of the SHR. Inhibitors of the formation of 20-HETE markedly reduce blood pressure in this strain [8,49].
3. 20-HETE in Stroke
Each year around 750,000 strokes occur in the USA, with 85% of that comprised of the ischemic type and 15% of the hemorrhagic type. Stroke results in a large number of patients with moderate to severe disability. Hypertension and diabetes are the most common factors that predispose patients to stroke [1,2].
Ischemic stroke occurs when a propagating clot, inherent plaque, or a damaged vessel suddenly occludes the blood supply in the specific region of the brain. Various ischemic stroke rodent models have been established to study different pathophysiologies and the immediate effect of occlusion [50]. Following occlusion of a cerebral vessel, there is a failure of ATP-dependent pump resulting in accumulation of intracellular sodium and calcium ions and depletion of intracellular potassium ions in the ischemic core. These changes cause cellular acidification and cell swelling, damaging of organelles including mitochondria, the release of proteolytic enzymes and inflammatory mediators, and eventually promote neuronal cell death. The degree of cerebral ischemic damage is augmented by the release of other factors such as glycine, glutamate, dopamine, 5-HT, and free fatty acids (FAA). FAA are used as substrates for the formation of neuronal lipid peroxides, which affects the metabolism and oxygen utilization of cells. Ischemia also promotes the release of other fatty acids such as the 20-HETE substrate AA, leukotrienes, and eicosanoids, which produce free radicals and further enhance leukocyte recruitment and cellular damage [7].
Subarachnoid hemorrhage (SAH), the most common type of hemorrhagic stroke, results from bleeding in the arachnoid space and clot formation. The cause of non-traumatic SAH and intracerebral hemorrhage (ICH) is an aneurysmal rupture. Hypertension has been implicated as one of the main risk factors for aneurysmal formation and rupture [51,52]. Acute cerebral ischemia (ACI) occurs within hours after intracranial bleeding occurs. There is also increased intracranial pressure dependent on the volume of bleed that further compromises cerebral perfusion during the ACI period, followed by delayed cerebral ischemia (DCI) due to a second peak if vasospasm, which can extend up to 21 days in some patients. Almost a third of patients who develop DCI suffer from permanent neurological damage [53,54,55].
3.1. 20-HETE in the Susceptibility of the Onset of Stroke
Emerging evidence demonstrates that several SNPs in CYP4F2 and CYP4A11 are associated with hypertension and stroke in several populations. A large urban-based population study in Sweden suggested there may be an increased risk of stroke, especially in males in the presence of the CYP4F2 rs2108622 (V433M) variant [15]. In Japanese men, the presence of the CYP4F2 rs2108622 G allele was associated with an increased risk of ischemic stroke [13]. Genetic polymorphisms in CYP4F2 were also found in Chinese [18,19], South India [16], Australian [14], and American [17] patients and were associated with an increased incidence of hypertension and both ischemic and hemorrhagic strokes. The SNPs of rs1126742 (F434S), rs9333025, and rs389011 in CYP4A11 were found to be associated with hypertension and stroke in American [24,27], Chinese [20,56], and Japanese [21,22] populations. These reports suggest that 20-HETE plays an important role in increasing susceptibility to the onset of stroke, especially in hypertensive patients. However, it remains to be determined whether this increased susceptibility is induced by lower or higher 20-HETE levels.
Two clinically relevant SNPs have been confirmed to reduce the formation of 20-HETE: CYP4F2 rs2108622 (V433M) and CYP4A11 rs1126742 (F434S) [25,26,27]. The observation that these SNPs are associated with an increased incidence of stroke could be explained by the fact that reduced 20-HETE diminishes the vasoconstrictive capability of cerebral vessels, which causes impaired myogenic response and autoregulation of cerebral blood flow. Such cerebral vascular dysregulation could transmit higher perfusion pressure to downstream capillaries, especially in association with hypertension. Enhanced pressure could distend the capillaries and damage BBB integrity resulting in increased permeability and bleeding. Further, with time, elevated perfusion pressure might increase capillary rarefaction and focal ischemia. This hypothesis is consistent with the results of our recent study. We found that SS rats with a 20-HETE deficiency exhibited impaired cerebral vascular autoregulation and BBB leakage, which was rescued in a CYP4A1 transgenic rat on the SS genetic background that restored vascular production of 20-HETE [29].
In contrast, ischemic stroke patients with both CYP4A11 (rs9333025) and CYP4F2 (rs2108622) SNPs exhibit elevated plasma 20-HETE levels [56]. Subjects of 322 minor ischemic stroke in a multicenter prospective observational study in China had levels of 20-HETE as high as 1,675 pmol/L [57]. These findings suggest that elevated 20-HETE production may promote the development of stroke, which at first glance seems contradictory to a role of reduced 20-HETE levels in stroke. However, animal studies in the SHR rat model are consistent with these reports since this hypertensive rat model exhibits higher 20-HETE levels and enhanced infarct size following transient ischemia and reperfusion injury. Moreover, since 20-HETE promotes oxidative stress, endothelial dysfunction and vascular inflammation, the hypothesis that elevated 20-HETE increases the susceptibility to the onset of stroke also seems logical. Although GPR75 has been identified as a 20-HETE receptor and contributes to endothelial dysfunction, it remains to be determined if the receptor plays a role in 20-HETE-related cerebrovascular function or in stroke [39,41].
Indeed, there is not enough evidence to determine whether 20-HETE is beneficial or detrimental to the onset of stroke. In fact, the controversial results between the available human and animal studies cannot be solely explained by the function of 20-HETE. The pathogenesis of hypertension and stroke are multi-factorial, including genetic and environmental. It is also possible that alterations in 20-HETE production caused by SNPs in CYP4A11 and CYP4F2 interplays with other molecules in the AA-CYP4A/4F-20-HETE pathway and changes the production of other AA metabolites catalyzed by cyclooxygenase (COX) and lipoxygenase (LOX).
3.2. 20-HETE in the Progression of Stroke
3.2.1. 20-HETE in the Progression of Ischemic Stroke
The production of 20-HETE is elevated in the plasma of ischemic stroke patients [58]. This observation is consistent with results obtained from animal studies using the middle cerebral artery occlusion (MCAo) model in rodents [7,33]. The levels of 20-HETE in MCAo rats are not only elevated in plasma but also cerebral tissues. As discussed above, the enhanced 20-HETE production after ischemic stimulation could be multifactorial, including accumulation of intracellular sodium and membrane depolarization to trigger calcium influx, and the release of AA. Elevated levels of 5-HT and glutamate also stimulate phospholipase and the release of AA following reperfusion. This view was confirmed in animal studies that revealed that inhibition of 20-HETE significantly reduced infarct volume and improved the neurological outcomes by reducing oxidative stress and apoptosis in neurons, rather than through a reduction of blood flow in the penumbral regions by 20-HETE [7,33,59,60,61]. In addition to increasing infarct size, elevated 20-HETE levels in mild ischemic stroke patients were shown to be associated with neurological dysfunction (ND). In this study, 72% of patients were found to have ND within the first 24h of admission [57]. Chronic administration of 20-HETE inhibitors in MCAo rats reduced neurological deficits [44]. 20-HETE synthase was elevated on the ipsilateral side 7 days after brain ischemic injury in a positron emission tomography (PET)-based study [62] suggesting upregulation of CYP4A and CYP4F isoforms might be involved in angiogenesis and neuroinflammation after stroke. Moreover, in a study analyzing carotid plaque characteristics, 20-HETE was found to be significantly higher in a patient with atheroma plaques compared to other metabolites [63]. Whether the 20-HETE content in the atheromatous plaque has any consequence on stroke outcome is still unclear, however, genetic variations in CYP450 isoforms have also been associated with plaque stability in patients with ischemic stroke [64].
3.2.2. 20-HETE in the Progression of Hemorrhagic Stroke
After the aneurysmal rupture, blood spreads into the arachnoid space and the perivascular spaces. The direct effect of blood and its components results in the sentinel headache and the initial cascade of pathological effects which causes the early neurological changes [65]. The severe headache may proceed with acute vasospasm contributing to ND on the first two days [54]. In most cases, it will be followed by delayed vasospasm, which takes days to weeks and has the most devastating consequences for SAH patients [7,17].
The acute injury in SAH results from the direct mechanical damage of bleeding [65]. The initial drop in the cerebral blood flow is also due to acute vasospasm and is correlated with the amount of the hemoglobin (Hb) in the cerebrospinal fluid (CSF) that was validated by the injection of clotted blood or oxyhemoglobin into the CSF [66]. The acute vasospasm is induced by the release of vasoconstrictors such as 5-HT and ATP from the clots, scavenging of the potent vasodilator NO by Hb, as well as the release of AA stimulated by 5-HT-activated PLA2 that produces 20-HETE [7,67]. The decrease in NO levels also stimulates CYP4A expression and in turn, 20-HETE production. Blocking the synthesis of 20-HETE with HET0016 prevents the fall in rCBF, and reduces the vasoconstrictor response of isolated MCA by >60% after SAH [68]. The elevated 20-HETE levels contribute to delayed vasospasm. DCI occurs in 30 to 40% of SAH patients [54]. The pathophysiologic processes responsible for DCI include impaired cerebral autoregulation, cortical spreading ischemia (CSI), microvascular thrombosis, and BBB disruption [33,54]. In animal SAH models, including rats and dogs, DCI was associated with increased levels of 20-HETE in CSF, and 20-HETE inhibitors reversed delayed vasospasm in these models [69,70].
Similarly, elevated levels of 20-HETE in CSF after SAH in human has been reported [17]. SAH is associated with activated PKC, MAPK, and ROCK, as well as diminished potassium channel activity, all of which results in depolarization and activation of VSMC. Both acute and delayed vasospasm following ischemic stroke could be attenuated with blockage of the actions of vasoconstrictors, including 20-HETE, 5-HT receptor, thromboxane, and ETA receptor, as well as inhibition of PKC, MAPK, and ROCK [7].
CSI, induced by the wave of neuronal depolarization or cortical spreading depolarization (CSD), is thought to play a role to brain damage in SAH patients by resulting in impaired neurovascular coupling and reduced cerebral blood flow in SAH [71]. The increased CSD has been associated with elevated 20-HETE synthesis and reduced cerebral blood flow in rat models, and 20-HETE inhibitors resulted in a neuroprotective effect and reduction in edema [71,72].
Very little is known about the relationship between 20-HETE and ICH. Administration of 20-HETE inhibitor reduced the hemorrhagic volume, the neurological deficit, and edema in ICH via reducing inflammatory response and ROS production without angiogenesis [73,74].
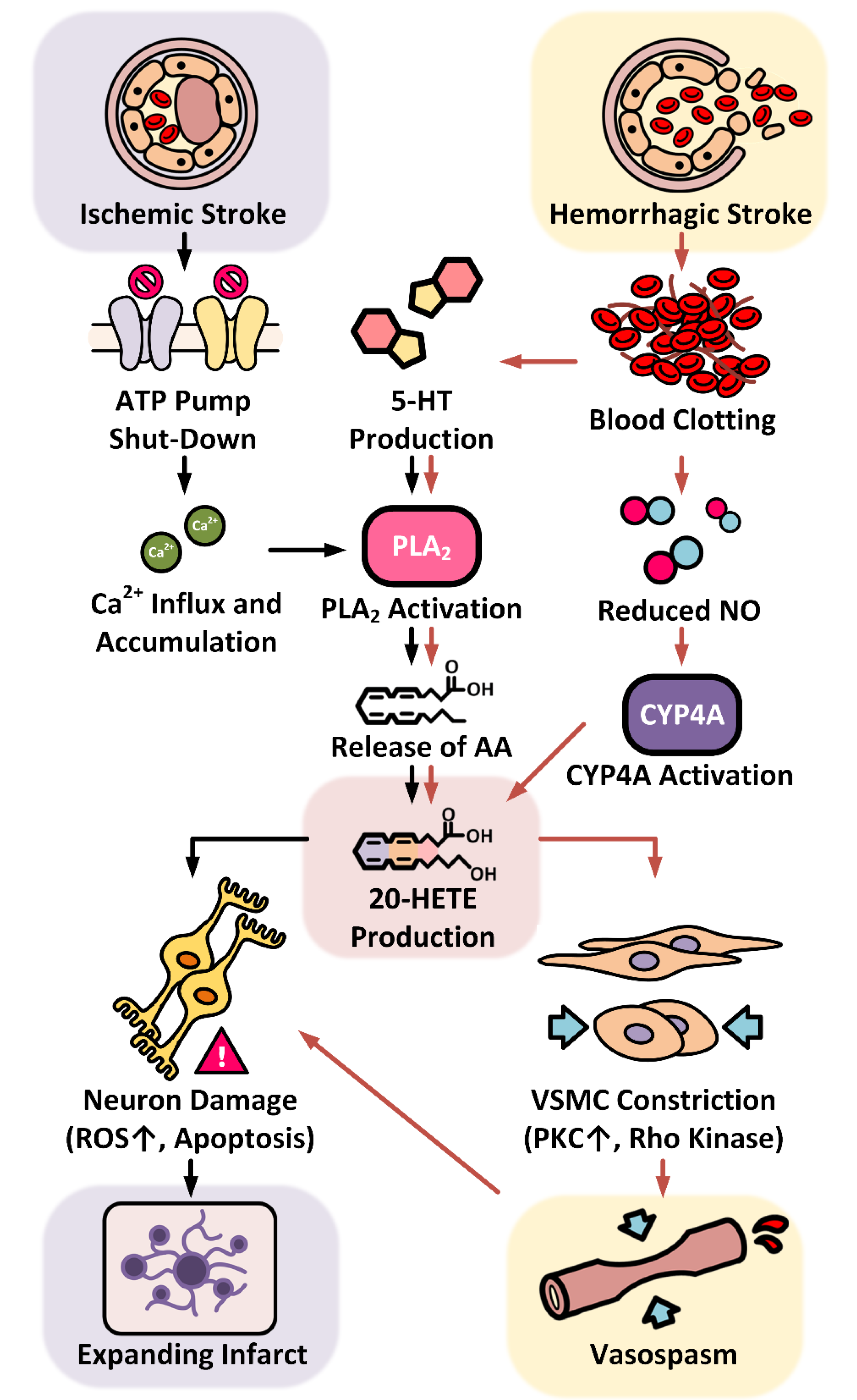
As summarized in Figure 2, it has been well established that elevated 20-HETE levels play a detrimental role in the progression of ischemic and hemorrhagic strokes and worsens neurological outcomes in human and animal SAH models.
3.3. 20-HETE in the Prognosis of Stroke
Approximately 25% of stroke patients die from this disease. Ten percent of ischemic stroke patients completely recover, but the outcomes after hemorrhagic stroke are generally worse than for ischemic stroke [1,2]. Long-term complications of stroke include disability and dementia. Prognosis of stroke not only depends on the type of stroke and the degree and duration of stroke but is also influenced by genetic and environmental factors.
BBB breakdown worsens the ischemic injury and promotes inflammation after stroke. Endothelial cells, pericytes, and astrocytes all play essential roles in maintaining the integrity of BBB. These cells all express CYP4A and produce 20-HETE [7,40,75]. A study using an MCAo obese ob/ob mouse demonstrated that increased BBB leakage enhanced ischemic brain injury by inducing astrocyte end-foot swelling and endothelial vesicles in association with vessel lumen collapse [76]. Low 20-HETE formation in SS rats has been shown to increase BBB leakage due to impaired myogenic response and CBF autoregulation, resulting in increased capillary pressure [29]. Impaired myogenic response and vascular leakage in hypertensive patients has been suggested to accelerate white matter damage and increase the risk of lacunar infarctions and later cognitive decline [77]. We also found that there is an association of SNPs in CYP4F2 and CYP4A11 with mild cognitive impairment and dementia-related endophenotypes including brain volumes and performance on cognitive tests, although only two of those SNPs were reported to reduce 20-HETE production [11]. On the other hand, inhibition of 20-HETE formation reduced BBB permeability in a rat TBI model [31]. This could be due to endothelial dysfunction, but whether and how 20-HETE acts on pericytes and astrocytes to affect BBB integrity has not been studied.
Long-term recovery from stroke also depends on the degree of angiogenesis [78]. The role of 20-HETE in angiogenesis is now well established. Sa et al. first reported that fibroblast growth factor 2 (FGF2) activates PLA2 via the p42 MAPK pathway which leads to a release of AA from membrane phospholipids which further stimulates CYP4A to produce 20-HETE [79]. Chen et al. reported that hypoxia and vascular endothelial growth factor (VEGF) induced CYP4A11 gene expression and increased 20-HETE production in endothelial progenitor cells (EPC) in vivo. In contrast, the angiogenesis was reduced by local inhibition of the 20-HETE system, suggesting 20-HETE has a vital role in EPC medicated angiogenesis [80]. 20-HETE inhibitors have been reported to block the neovascularization of the cornea and the implanted glioblastoma cell (solid tumors) [81]. Various other studies using 20-HETE inhibitors have observed similar reduced vascularization and growth of rat 9L and human U251 glioma cells implanted in the brain of nude rats [7]. The reason is likely due to upregulation of CYP4A as reported by Guo et al., which enhanced phenotypic proliferation in both in vivo and in vitro experiments [82,83]. Since then, there are reports of upregulation of CYP4A/4F enzymes in thyroid, breast, colon, and ovarian cancer, as well as pancreatic adenocarcinoma that all exhibit hyperactivated angiogenesis [7], and the therapeutic effects of 20-HETE inhibitors in these cancer types [84]. More recently, using oxygen–glucose deprivation of astrocytes and a mouse model of focal cerebral ischemia, Liu et al. reported that astrocyte-derived 20-HETE contributed to angiogenesis after the induction of ischemic stroke by inducing endothelial cell proliferation, tube formation, and migration [85].
In summary, the available data does not provide sufficient information to indicate whether 20-HETE is beneficial or detrimental to the prognosis of stroke. 20-HETE has shown a positive correlation with better recovery from stroke by promoting CBF autoregulation, reducing BBB leakage, and enhancing angiogenesis; it also possibly worsens stroke recovery by increasing endothelial dysfunction, oxidative stress, and vascular inflammation.
4. Conclusion and Perspectives
Hypertension is a common disease that puts millions at risk of developing stroke and other cardiovascular disorders. It is critical that patients are advised to address hypertension as soon as possible through lifestyle changes and pharmaceutical therapies so that renal damage and stroke risk can be mitigated. While hypertension is a modifiable risk factor for preventing stroke, there are definitive data showing that genetics also plays a major role. Certain SNPs are associated with hypertension and stroke in humans. In the kidney, 20-HETE protects against hypertensive damage through the constriction of Af-art. However, increased levels of 20-HETE are associated with fibrosis and renal damage. On the other hand, prolonged exposure to 20-HETE also has a pro-hypertensive role in the vasculature. Interestingly, 20-HETE also has a contradictory role in the brain. In the vasculature, reduced 20-HETE is associated with an impaired myogenic response, autoregulation, and BBB integrity. However, high amounts of 20-HETE have been found to cause endothelial dysfunction and activation as well as disruption of BBB, increasing the risk of stroke. 20-HETE is normally increased after ischemic and hemorrhagic strokes, and the use of 20-HETE inhibitors such as HET0016 during the early stages of strokes has been found to reduce infarct size, prevent vasospasm, and improve neurological outcomes. Because of the rise in 20-HETE post-stroke, the best outcomes are achieved using 20-HETE inhibitors in the early stages of stroke recovery. Moreover, 20-HETE plays an important role in neovascularization and angiogenesis that are necessary to stroke recovery; thus, more research needs to be done on the timing of therapy with 20-HETE inhibitors.
Author Contributions
Conceptualization, F.F.; writing—original draft preparation, S.S., and K.V. (equally contributed) and F.F.; writing—review and editing, F.F., R.J.R., S.S., and G.W.B.; visualization, M.L. and L.F.; funding acquisition, F.F. and R.J.R.
Funding
This work was supported by the National Institutes of Health (AG050049, AG057842, P20GM104357, DK104184, and HL138685) and the American Heart Association (16GRNT31200036).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| 20-COOH-HETE | 20-carboxy-eicosatetraenoic acid |
| 20-HETE | 20-hydroxyeicosatetraenoic acid |
| 5-HT | serotonin |
| GWAS | Genome-wide association study |
| AA | arachidonic acid |
| ACE | angiotensin-converting enzyme |
| ACI | acute cerebral ischemia |
| ADH | alcohol dehydrogenase |
| Af-art | afferent arteriole |
| ANG II | angiotensin II |
| AT1 | angiotensin II receptor 1 |
| BBB | blood-brain barrier |
| BK | blocking calcium-activated potassium channel |
| CADASIL | cerebral autosomal dominant arteriopathy with subcortical infarcts and leukoencephalopathy |
| CARASIL | cerebral autosomal-recessive arteriopathy with subcortical infarcts and leukoencephalopathy |
| COX | cyclooxygenase |
| COX2 | cyclooxygenase-2 |
| CSD | cortical spreading depolarization |
| CSF | cerebrospinal fluid |
| CSI | cortical spreading ischemia |
| DCI | delayed cerebral ischemia |
| EETs | epoxyeicosatrienoic acids |
| EPC | endothelial progenitor cells |
| FAA | free fatty acids |
| FGF2 | fibroblast growth factor 2 |
| KO | knock out |
| LOX | lipoxygenase |
| MAPKs | mitogen-activated protein kinases |
| MCAo | middle cerebral artery occlusion |
| ND | neurological dysfunction |
| NHE3 | sodium-hydrogen exchanger 3 |
| NKCC2 | Na-K-Cl cotransporter |
| NO | nitric oxide |
| PKC | protein kinase C |
| PLA2 | phospholipase A2 |
| PT | proximal tubule |
| RAAS | renin-angiotensin-aldosterone system |
| ROMK2 | renal outer medullary potassium channel 2 |
| SAH | subarachnoid hemorrhage |
| SHR | spontaneously hypertensive rat |
| SNP | single nucleotide polymorphisms |
| SS | salt sensitive |
| TALH | thick ascending loop of Henle |
| TBI | traumatic brain injury |
| TGF | tubuloglomerular feedback responses |
| TRPC6 | transient receptor potential cation channel 6 |
| TRPV1 | transient receptor potential vanilloid 1 |
| UGT | uridine 5’-diphosphoglucuronosyltransferase |
| VEGF | vascular endothelial growth factor |
| VSMC | vascular smooth muscle cell |
| WT | wild type |
References
- Nwankwo, T.; Yoon, S.S.; Burt, V.; Gu, Q. Hypertension among adults in the United States: National Health and Nutrition Examination Survey, 2011-2012. NCHS Data Brief. 2013, 1–8. [Google Scholar]
- Murphy, S.L.; Xu, J.; Kochanek, K.D.; Curtin, S.C.; Arias, E. Deaths: Final Data for 2015. Natl. Vital Stat Rep. 2017, 66, 1–75. [Google Scholar] [PubMed]
- Shekhar, S.; Liu, R.; Travis, O.K.; Roman, R.J.; Fan, F. Cerebral Autoregulation in Hypertension and Ischemic Stroke: A Mini Review. J. Pharm. Sci. Exp. Pharmacol. 2017, 2017, 21–27. [Google Scholar] [PubMed]
- Shekhar, S.; Cunningham, M.W.; Pabbidi, M.R.; Wang, S.; Booz, G.W.; Fan, F. Targeting vascular inflammation in ischemic stroke: Recent developments on novel immunomodulatory approaches. Eur. J. Pharmacol. 2018, 833, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Francis, J.; Raghunathan, S.; Khanna, P. The role of genetics in stroke. Postgrad. Med. J. 2007, 83, 590–595. [Google Scholar] [CrossRef]
- Chauhan, G.; Debette, S. Genetic Risk Factors for Ischemic and Hemorrhagic Stroke. Curr. Cardiol. Rep. 2016, 18, 124. [Google Scholar] [CrossRef] [PubMed]
- Fan, F.; Ge, Y.; Lv, W.; Elliott, M.R.; Muroya, Y.; Hirata, T.; Booz, G.W.; Roman, R.J. Molecular mechanisms and cell signaling of 20-hydroxyeicosatetraenoic acid in vascular pathophysiology. Front. Biosci. (Landmark Ed.) 2016, 21, 1427–1463. [Google Scholar] [PubMed]
- Fan, F.; Roman, R.J. Effect of Cytochrome P450 Metabolites of Arachidonic Acid in Nephrology. J. Am. Soc. Nephrol. 2017, 28, 2845–2855. [Google Scholar] [CrossRef] [PubMed]
- Fava, C.; Ricci, M.; Melander, O.; Minuz, P. Hypertension, cardiovascular risk and polymorphisms in genes controlling the cytochrome P450 pathway of arachidonic acid: A sex-specific relation? Prostaglandins Other Lipid Mediat. 2012, 98, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Fava, C.; Bonafini, S. Eicosanoids via CYP450 and cardiovascular disease: Hints from genetic and nutrition studies. Prostaglandins Other Lipid Mediat. 2018, 139, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Fan, F.; Simino, J.; Auchus, A.; Knopman, D.; Boerwinkle, E.; Fornage, M.; Mosley, T.; Roman, R. Functional variants in CYP4A11 and CYP4F2 are associated with cognitive impairment and related dementia endophenotypes in the elderly. In Proceedings of the 16th International Winter Eicosanoid Conference, Baltimore, MD, USA, 13 March 2016; p. CV5. [Google Scholar]
- Zhang, C.; Booz, G.W.; Yu, Q.; He, X.; Wang, S.; Fan, F. Conflicting roles of 20-HETE in hypertension and renal end organ damage. Eur. J. Pharmacol. 2018, 833, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Nakayama, T.; Sato, N.; Izumi, Y.; Kasamaki, Y.; Shindo, A.; Ohta, M.; Soma, M.; Aoi, N.; Sato, M.; et al. A haplotype of the CYP4F2 gene is associated with cerebral infarction in Japanese men. Am. J. Hypertens. 2008, 21, 1216–1223. [Google Scholar] [CrossRef] [PubMed]
- Ward, N.C.; Tsai, I.J.; Barden, A.; van Bockxmeer, F.M.; Puddey, I.B.; Hodgson, J.M.; Croft, K.D. A single nucleotide polymorphism in the CYP4F2 but not CYP4A11 gene is associated with increased 20-HETE excretion and blood pressure. Hypertension 2008, 51, 1393–1398. [Google Scholar] [CrossRef] [PubMed]
- Fava, C.; Montagnana, M.; Almgren, P.; Rosberg, L.; Lippi, G.; Hedblad, B.; Engstrom, G.; Berglund, G.; Minuz, P.; Melander, O. The V433M variant of the CYP4F2 is associated with ischemic stroke in male Swedes beyond its effect on blood pressure. Hypertension. 2008, 52, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Munshi, A.; Sharma, V.; Kaul, S.; Al-Hazzani, A.; Alshatwi, A.A.; Shafi, G.; Koppula, R.; Mallemoggala, S.B.; Jyothy, A. Association of 1347 G/A cytochrome P450 4F2 (CYP4F2) gene variant with hypertension and stroke. Mol. Biol. Rep. 2012, 39, 1677–1682. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, M.K.; Crago, E.A.; Conley, Y.P.; Balzer, J.R.; Ren, D.; Ducruet, A.F.; Kochanek, P.M.; Sherwood, P.R.; Poloyac, S.M. 20-HETE is associated with unfavorable outcomes in subarachnoid hemorrhage patients. J. Cereb. Blood Flow Metab. 2015, 35, 1515–1522. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Zhu, G.; Liu, F.; Zhang, H.; Qin, X.; Li, L.; Zhiyi, H. CYP4F2 gene V433M polymorphism is associated with ischemic stroke in the male Northern Chinese Han population. Prog. Neuropsychopharmacol. Biol. Psychiatry. 2010, 34, 664–668. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Cui, G.; Zhang, L.; Xu, Y.; Bao, X.; Tu, Y.; Wu, B.; Wang, Q.; Hui, R.; Wang, W.; et al. Association of common variants of CYP4A11 and CYP4F2 with stroke in the Han Chinese population. Pharm. Genom. 2010, 20, 187–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Jin, L.; Mu, T.; Fan, Y.; Zhang, H.; Zhu, Y.; Mao, X.; Li, R.; Tang, S. Associations of CYP4A11 gene-gene and gene-smoking interactions with essential hypertension in the male eastern Chinese Han population. Clin. Exp. Hypertens. 2017, 39, 448–453. [Google Scholar] [CrossRef]
- Fu, Z.; Nakayama, T.; Sato, N.; Izumi, Y.; Kasamaki, Y.; Shindo, A.; Ohta, M.; Soma, M.; Aoi, N.; Sato, M.; et al. Haplotype-based case study of human CYP4A11 gene and cerebral infarction in Japanese subject. Endocrine 2008, 33, 215–222. [Google Scholar] [CrossRef]
- Fu, Z.; Nakayama, T.; Sato, N.; Izumi, Y.; Kasamaki, Y.; Shindo, A.; Ohta, M.; Soma, M.; Aoi, N.; Sato, M.; et al. A haplotype of the CYP4A11 gene associated with essential hypertension in Japanese men. J. Hypertens. 2008, 26, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Gainer, J.V.; Lipkowitz, M.S.; Yu, C.; Waterman, M.R.; Dawson, E.P.; Capdevila, J.H.; Brown, N.J.; Group, A.S. Association of a CYP4A11 variant and blood pressure in black men. J. Am. Soc. Nephrol. 2008, 19, 1606–1612. [Google Scholar] [CrossRef] [PubMed]
- Laffer, C.L.; Gainer, J.V.; Waterman, M.R.; Capdevila, J.H.; Laniado-Schwartzman, M.; Nasjletti, A.; Brown, N.J.; Elijovich, F. The T8590C polymorphism of CYP4A11 and 20-hydroxyeicosatetraenoic acid in essential hypertension. Hypertension 2008, 51, 767–772. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.Y.; Lee, S.J.; Min, J.; Oh, K.S.; Kim, D.H.; Kim, H.S.; Shin, J.G. Identification of novel CYP4F2 genetic variants exhibiting decreased catalytic activity in the conversion of arachidonic acid to 20-hydroxyeicosatetraenoic acid (20-HETE). Prostaglandins Leukot. Essent. Fatty Acids. 2018, 131, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Stec, D.E.; Roman, R.J.; Flasch, A.; Rieder, M.J. Functional polymorphism in human CYP4F2 decreases 20-HETE production. Physiol. Genom. 2007, 30, 74–81. [Google Scholar] [CrossRef] [Green Version]
- Gainer, J.V.; Bellamine, A.; Dawson, E.P.; Womble, K.E.; Grant, S.W.; Wang, Y.; Cupples, L.A.; Guo, C.Y.; Demissie, S.; O’Donnell, C.J.; et al. Functional variant of CYP4A11 20-hydroxyeicosatetraenoic acid synthase is associated with essential hypertension. Circulation 2005, 111, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Honda, M.; Takahashi, M.; Tsukada, C.; Ito, M.; Katono, Y.; Hosono, H.; Saigusa, D.; Suzuki, N.; Tomioka, Y.; et al. Functional characterization of 10 CYP4A11 allelic variants to evaluate the effect of genotype on arachidonic acid omega-hydroxylation. Drug Metab. Pharmacokinet. 2015, 30, 119–122. [Google Scholar] [CrossRef]
- Fan, F.; Geurts, A.M.; Murphy, S.R.; Pabbidi, M.R.; Jacob, H.J.; Roman, R.J. Impaired myogenic response and autoregulation of cerebral blood flow is rescued in CYP4A1 transgenic Dahl salt-sensitive rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R379–R390. [Google Scholar] [CrossRef]
- Williams, J.M.; Fan, F.; Murphy, S.; Schreck, C.; Lazar, J.; Jacob, H.J.; Roman, R.J. Role of 20-HETE in the antihypertensive effect of transfer of chromosome 5 from Brown Norway to Dahl salt-sensitive rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R1209–R1218. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Wang, M.; Wei, X.; Li, W. 20-HETE Inhibition by HET0016 Decreases the Blood-Brain Barrier Permeability and Brain Edema After Traumatic Brain Injury. Front. Aging Neurosci. 2018, 10, 207. [Google Scholar] [CrossRef]
- Lu, L.; Wang, M.; Yuan, F.; Wei, X.; Li, W. Roles of elevated 20HETE in the breakdown of blood brain barrier and the severity of brain edema in experimental traumatic brain injury. Mol. Med. Rep. 2018, 17, 7339–7345. [Google Scholar] [CrossRef] [PubMed]
- Roman, R.J.; Fan, F. 20-HETE: Hypertension and Beyond. Hypertension 2018, 72, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Liao, D.; Yi, X.; Zhang, B.; Zhou, Q.; Lin, J. Interaction Between CYP4F2 rs2108622 and CPY4A11 rs9333025 Variants Is Significantly Correlated with Susceptibility to Ischemic Stroke and 20-Hydroxyeicosatetraenoic Acid Level. Genet. Test Mol. Biomark. 2016, 20, 223–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, F.; Muroya, Y.; Roman, R.J. Cytochrome P450 eicosanoids in hypertension and renal disease. Curr. Opin. Nephrol. Hypertens. 2015, 24, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Orozco, L.D.; Liu, H.; Perkins, E.; Johnson, D.A.; Chen, B.B.; Fan, F.; Baker, R.C.; Roman, R.J. 20-Hydroxyeicosatetraenoic acid inhibition attenuates balloon injury-induced neointima formation and vascular remodeling in rat carotid arteries. J. Pharmacol. Exp. Ther. 2013, 346, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Hill, E.; Fitzpatrick, F.; Murphy, R.C. Biological activity and metabolism of 20-hydroxyeicosatetraenoic acid in the human platelet. Br. J. Pharmacol. 1992, 106, 267–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, F.; Sun, C.W.; Maier, K.G.; Williams, J.M.; Pabbidi, M.R.; Didion, S.P.; Falck, J.R.; Zhuo, J.; Roman, R.J. 20-Hydroxyeicosatetraenoic acid contributes to the inhibition of K+ channel activity and vasoconstrictor response to angiotensin II in rat renal microvessels. PLoS ONE 2013, 8, e82482. [Google Scholar] [CrossRef] [PubMed]
- Garcia, V.; Gilani, A.; Shkolnik, B.; Pandey, V.; Zhang, F.F.; Dakarapu, R.; Gandham, S.K.; Reddy, N.R.; Graves, J.P.; Gruzdev, A.; et al. 20-HETE Signals Through G Protein-Coupled Receptor GPR75 (Gq) to Affect Vascular Function and Trigger Hypertension. Circ. Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Fan, F.; He, X.; Cornelius, C.C.; Roman, R.J. The potential neuroprotective role of 20-HETE in association with its receptor GPR75 expression in hypertension. In Proceedings of the 17th International Winter Eicosanoid Conference, Baltimore, MA, USA, 11 March 2018. [Google Scholar]
- Fan, F.; Roman, R. GPR75 Identified as the First 20-HETE Receptor: A Chemokine Receptor Adopted by a New Family. Circ. Res. 2017, 12, 1696–1698. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, D.; Thirumangalakudi, L.; Grammas, P. RANTES upregulation in the Alzheimer’s disease brain: a possible neuroprotective role. Neurobiol. Aging 2010, 31, 8–16. [Google Scholar] [CrossRef]
- Dedoni, S.; Campbell, L.A.; Harvey, B.K.; Avdoshina, V.; Mocchetti, I. The orphan G-protein-coupled receptor 75 signaling is activated by the chemokine CCL5. J. Neurochem. 2018, 146, 526–539. [Google Scholar] [CrossRef]
- Tunaru, S.; Bonnavion, R.; Brandenburger, I.; Preussner, J.; Thomas, D.; Scholich, K.; Offermanns, S. 20-HETE promotes glucose-stimulated insulin secretion in an autocrine manner through FFAR1. Nat. Commun. 2018, 9, 177. [Google Scholar] [CrossRef]
- Park, S.K.; Herrnreiter, A.; Pfister, S.L.; Gauthier, K.M.; Falck, B.A.; Falck, J.R.; Campbell, W.B. GPR40 is a low-affinity epoxyeicosatrienoic acid receptor in vascular cells. J. Biol. Chem. 2018, 293, 10675–10691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Lopez, B.; Dos Santos, E.A.; Falck, J.R.; Roman, R.J. Effects of 20-HETE on Na+ transport and Na+-K+-ATPase activity in the thick ascending loop of Henle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R2400–R2405. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Murphy, S.R.; Fan, F.; Williams, J.M.; Falck, J.R.; Liu, R.; Roman, R.J. Role of 20-HETE in the impaired myogenic and TGF responses of the Af-Art of Dahl salt-sensitive rats. Am. J. Physiol. Renal. Physiol. 2014, 307, F509–F515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stec, D.E.; Deng, A.Y.; Rapp, J.P.; Roman, R.J. Cytochrome P4504A genotype cosegregates with hypertension in Dahl S rats. Hypertension 1996, 27, 564–568. [Google Scholar] [CrossRef]
- Roman, R.J. P-450 metabolites of arachidonic acid in the control of cardiovascular function. Physiol. Rev. 2002, 82, 131–185. [Google Scholar] [CrossRef]
- Carmichael, S.T. Rodent models of focal stroke: size, mechanism, and purpose. NeuroRx 2005, 2, 396–409. [Google Scholar] [CrossRef]
- Feigin, V.; Parag, V.; Lawes, C.M.; Rodgers, A.; Suh, I.; Woodward, M.; Jamrozik, K.; Ueshima, H. Asia Pacific Cohort Studies Collaboration. Smoking and elevated blood pressure are the most important risk factors for subarachnoid hemorrhage in the Asia-Pacific region: an overview of 26 cohorts involving 306,620 participants. Stroke 2005, 36, 1360–1365. [Google Scholar] [CrossRef]
- Vlak, M.H.; Rinkel, G.J.; Greebe, P.; Greving, J.P.; Algra, A. Lifetime risks for aneurysmal subarachnoid haemorrhage: multivariable risk stratification. J. Neurol. Neurosurg. Psychiatry 2013, 84, 619–623. [Google Scholar] [CrossRef]
- Cossu, G.; Messerer, M.; Oddo, M.; Daniel, R.T. To look beyond vasospasm in aneurysmal subarachnoid haemorrhage. Biomed. Res. Int. 2014, 2014, 628597. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, R.L. Delayed neurological deterioration after subarachnoid haemorrhage. Nat. Rev. Neurol. 2014, 10, 44–58. [Google Scholar] [CrossRef] [PubMed]
- Vergouwen, M.D.; Vermeulen, M.; van Gijn, J.; Rinkel, G.J.; Wijdicks, E.F.; Muizelaar, J.P.; Mendelow, A.D.; Juvela, S.; Yonas, H.; Terbrugge, K.G.; et al. Definition of delayed cerebral ischemia after aneurysmal subarachnoid hemorrhage as an outcome event in clinical trials and observational studies: proposal of a multidisciplinary research group. Stroke 2010, 41, 2391–2395. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Yi, X.; Wang, C.; Liao, D.; Lin, J.; Chi, L. Cytochrome 4A11 Genetic Polymorphisms Increase Susceptibility to Ischemic Stroke and Associate with Atherothrombotic Events After Stroke in Chinese. Genet. Test Mol. Biomark. 2015, 19, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Han, Z.; Zhou, Q.; Lin, J.; Liu, P. 20-Hydroxyeicosatetraenoic Acid as a Predictor of Neurological Deterioration in Acute Minor Ischemic Stroke. Stroke 2016, 47, 3045–3047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, N.C.; Croft, K.D.; Blacker, D.; Hankey, G.J.; Barden, A.; Mori, T.A.; Puddey, I.B.; Beer, C.D. Cytochrome P450 metabolites of arachidonic acid are elevated in stroke patients compared with healthy controls. Clin. Sci. (Lond.) 2011, 121, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Dunn, K.M.; Renic, M.; Flasch, A.K.; Harder, D.R.; Falck, J.; Roman, R.J. Elevated production of 20-HETE in the cerebral vasculature contributes to severity of ischemic stroke and oxidative stress in spontaneously hypertensive rats. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H2455–H2465. [Google Scholar] [CrossRef] [Green Version]
- Marumo, T.; Eto, K.; Wake, H.; Omura, T.; Nabekura, J. The inhibitor of 20-HETE synthesis, TS-011, improves cerebral microcirculatory autoregulation impaired by middle cerebral artery occlusion in mice. Br. J. Pharmacol. 2010, 161, 1391–1402. [Google Scholar] [CrossRef] [Green Version]
- Omura, T.; Tanaka, Y.; Miyata, N.; Koizumi, C.; Sakurai, T.; Fukasawa, M.; Hachiuma, K.; Minagawa, T.; Susumu, T.; Yoshida, S.; et al. Effect of a new inhibitor of the synthesis of 20-HETE on cerebral ischemia reperfusion injury. Stroke 2006, 37, 1307–1313. [Google Scholar] [CrossRef]
- Kawasaki, T.; Marumo, T.; Shirakami, K.; Mori, T.; Doi, H.; Suzuki, M.; Watanabe, Y.; Chaki, S.; Nakazato, A.; Ago, Y.; et al. Increase of 20-HETE synthase after brain ischemia in rats revealed by PET study with 11C-labeled 20-HETE synthase-specific inhibitor. J. Cereb. Blood Flow Metab. 2012, 32, 1737–1746. [Google Scholar] [CrossRef]
- Auguet, T.; Aragones, G.; Colom, M.; Aguilar, C.; Martin-Paredero, V.; Canela, N.; Ruyra, X.; Richart, C. Targeted metabolomic approach in men with carotid plaque. PLoS ONE 2018, 13, e0200547. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.Y.; Liao, D.X.; Wang, C.; Cheng, W.; Fu, X.Q.; Zhang, B. Cytochrome P450 Genetic Variants and Their Metabolite Levels Associated with Plaque Stability in Patients with Ischemic Stroke. J. Atheroscler. Thromb. 2016, 23, 330–338. [Google Scholar] [CrossRef] [Green Version]
- Fujii, M.; Yan, J.; Rolland, W.B.; Soejima, Y.; Caner, B.; Zhang, J.H. Early brain injury, an evolving frontier in subarachnoid hemorrhage research. Transl. Stroke Res. 2013, 4, 432–446. [Google Scholar] [CrossRef] [PubMed]
- Megyesi, J.F.; Vollrath, B.; Cook, D.A.; Findlay, J.M. In vivo animal models of cerebral vasospasm: a review. Neurosurgery 2000, 46, 448–461. [Google Scholar] [CrossRef] [PubMed]
- Harder, D.R.; Dernbach, P.; Waters, A. Possible cellular mechanism for cerebral vasospasm after experimental subarachnoid hemorrhage in the dog. J. Clin. Investig. 1987, 80, 875–880. [Google Scholar] [CrossRef] [PubMed]
- Cambj-Sapunar, L.; Yu, M.; Harder, D.R.; Roman, R.J. Contribution of 5-hydroxytryptamine1B receptors and 20-hydroxyeiscosatetraenoic acid to fall in cerebral blood flow after subarachnoid hemorrhage. Stroke 2003, 34, 1269–1275. [Google Scholar] [CrossRef] [PubMed]
- Hacein-Bey, L.; Harder, D.R.; Meier, H.T.; Varelas, P.N.; Miyata, N.; Lauer, K.K.; Cusick, J.F.; Roman, R.J. Reversal of delayed vasospasm by TS-011 in the dual hemorrhage dog model of subarachnoid hemorrhage. AJNR Am. J. Neuroradiol. 2006, 27, 1350–1354. [Google Scholar] [PubMed]
- Roman, R.J.; Renic, M.; Dunn, K.M.; Takeuchi, K.; Hacein-Bey, L. Evidence that 20-HETE contributes to the development of acute and delayed cerebral vasospasm. Neurol. Res. 2006, 28, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Fordsmann, J.C.; Ko, R.W.; Choi, H.B.; Thomsen, K.; Witgen, B.M.; Mathiesen, C.; Lonstrup, M.; Piilgaard, H.; MacVicar, B.A.; Lauritzen, M. Increased 20-HETE synthesis explains reduced cerebral blood flow but not impaired neurovascular coupling after cortical spreading depression in rat cerebral cortex. J. Neurosci. 2013, 33, 2562–2570. [Google Scholar] [CrossRef] [PubMed]
- Shaik, J.S.; Poloyac, S.M.; Kochanek, P.M.; Alexander, H.; Tudorascu, D.L.; Clark, R.S.; Manole, M.D. 20-Hydroxyeicosatetraenoic Acid Inhibition by HET0016 Offers Neuroprotection, Decreases Edema, and Increases Cortical Cerebral Blood Flow in a Pediatric Asphyxial Cardiac Arrest Model in Rats. J. Cereb. Blood Flow Metab. 2015, 35, 1757–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyata, N.; Seki, T.; Tanaka, Y.; Omura, T.; Taniguchi, K.; Doi, M.; Bandou, K.; Kametani, S.; Sato, M.; Okuyama, S.; et al. Beneficial effects of a new 20-hydroxyeicosatetraenoic acid synthesis inhibitor, TS-011 [N-(3-chloro-4-morpholin-4-yl) phenyl-N’-hydroxyimido formamide], on hemorrhagic and ischemic stroke. J. Pharmacol. Exp. Ther. 2005, 314, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Zhao, X.; Lan, X.; Li, Q.; Gao, Y.; Liu, X.; Wan, J.; Yang, Z.; Chen, X.; Zang, W.; et al. 20-HETE synthesis inhibition promotes cerebral protection after intracerebral hemorrhage without inhibiting angiogenesis. J. Cereb. Blood Flow Metab. 2019, 39, 1531–1543. [Google Scholar] [CrossRef] [PubMed]
- Gebremedhin, D.; Zhang, D.X.; Carver, K.A.; Rau, N.; Rarick, K.R.; Roman, R.J.; Harder, D.R. Expression of CYP 4A omega-hydroxylase and formation of 20-hydroxyeicosatetreanoic acid (20-HETE) in cultured rat brain astrocytes. Prostaglandins Other Lipid Mediat. 2016, 124, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Haley, M.J.; Lawrence, C.B. The blood-brain barrier after stroke: Structural studies and the role of transcytotic vesicles. J. Cereb. Blood Flow Metab. 2017, 37, 456–470. [Google Scholar] [CrossRef] [PubMed]
- Faraco, G.; Iadecola, C. Hypertension: a harbinger of stroke and dementia. Hypertension 2013, 62, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Ergul, A.; Alhusban, A.; Fagan, S.C. Angiogenesis: a harmonized target for recovery after stroke. Stroke 2012, 43, 2270–2274. [Google Scholar] [CrossRef] [PubMed]
- Sa, G.; Murugesan, G.; Jaye, M.; Ivashchenko, Y.; Fox, P.L. Activation of cytosolic phospholipase A2 by basic fibroblast growth factor via a p42 mitogen-activated protein kinase-dependent phosphorylation pathway in endothelial cells. J. Biol. Chem. 1995, 270, 2360–2366. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ackerman, R.; Saleh, M.; Gotlinger, K.H.; Kessler, M.; Mendelowitz, L.G.; Falck, J.R.; Arbab, A.S.; Scicli, A.G.; Schwartzman, M.L.; et al. 20-HETE regulates the angiogenic functions of human endothelial progenitor cells and contributes to angiogenesis in vivo. J. Pharmacol. Exp. Ther. 2014, 348, 442–451. [Google Scholar] [CrossRef]
- Chen, P.; Guo, M.; Wygle, D.; Edwards, P.A.; Falck, J.R.; Roman, R.J.; Scicli, A.G. Inhibitors of cytochrome P450 4A suppress angiogenic responses. Am. J. Pathol. 2005, 166, 615–624. [Google Scholar] [CrossRef]
- Guo, A.M.; Roman, R.J.; Falck, J.R.; Jafari, K.; Edwards, P.A.; Scicli, A.G. Overexpression of CYP4A1-20-HETE in U251 Glioma Cell Induces Hyperproliferative Phenotypes in vitro and in vivo. Proc. FASEB J. 2008, 22, 1136-13. [Google Scholar]
- Guo, A.M.; Sheng, J.; Scicli, G.M.; Arbab, A.S.; Lehman, N.L.; Edwards, P.A.; Falck, J.R.; Roman, R.J.; Scicli, A.G. Expression of CYP4A1 in U251 human glioma cell induces hyperproliferative phenotype in vitro and rapidly growing tumors in vivo. J. Pharmacol. Exp. Ther. 2008, 327, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Alexanian, A.; Sorokin, A. Targeting 20-HETE producing enzymes in cancer - rationale, pharmacology, and clinical potential. Onco Targets Ther. 2013, 6, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Y.; Zhan, M.; Liu, Y.; Li, Z.; Li, J.; Cheng, G.; Teng, G.; Lu, L. Astrocytic cytochrome P450 4A/20-hydroxyeicosatetraenoic acid contributes to angiogenesis in the experimental ischemic stroke. Brain Res. 2019, 1708, 160–170. [Google Scholar] [CrossRef] [PubMed]
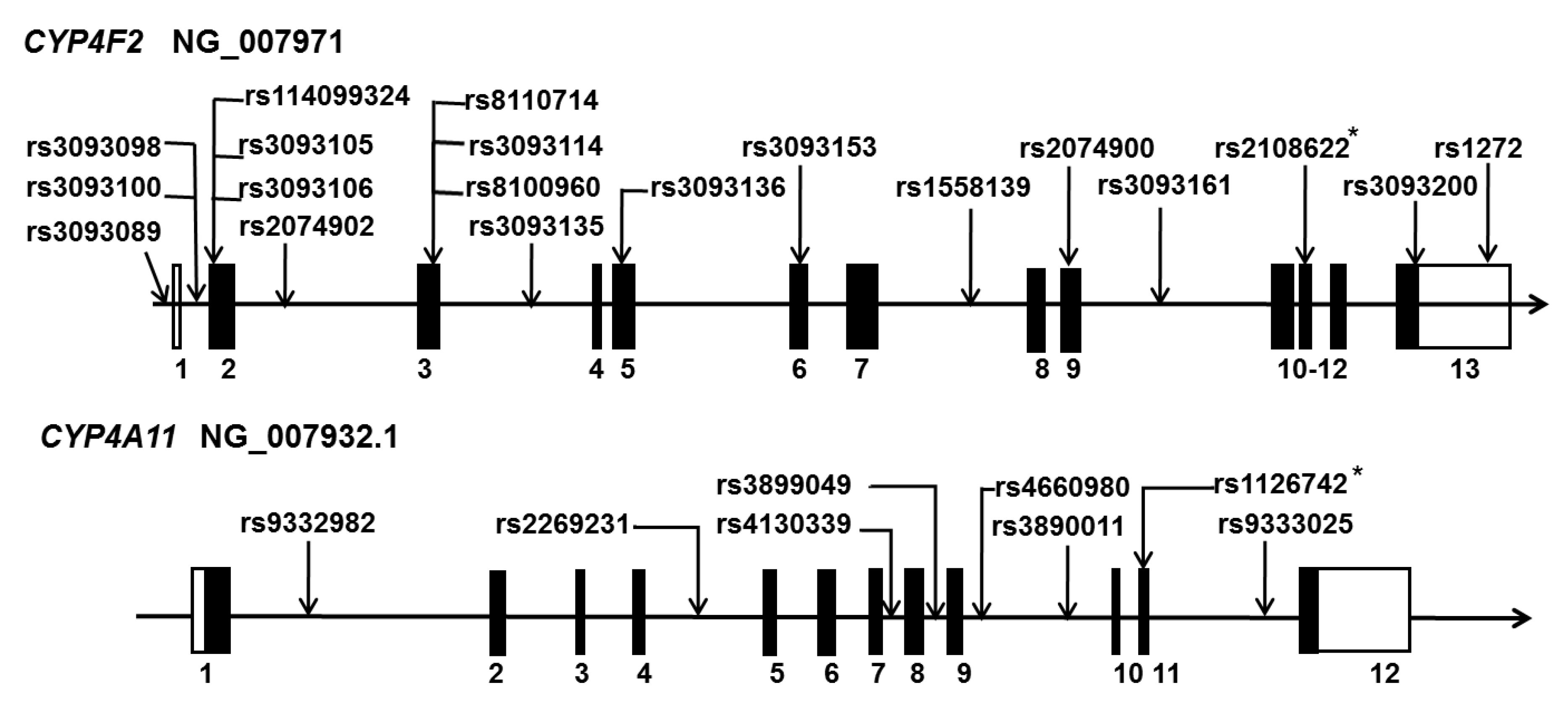
Figure 1.
Genetic polymorphisms in human CYP4F2 and CYP4A11. Localization of single nucleotide polymorphisms (SNPs) in genomic DNA of CYP4F2 (upper panel, NG_007971) and CYP4A11 (lower panel, NG_007932.1). Numbers indicate the order of exons. Solid portions of exons indicate transcription regions. Open frames on exons indicate non-transcription regions of CYP4F2 and CYP4A11. Published RefSNPs (rs) numbers linked with hypertension and stroke are listed. Asterisks (*) indicate that the SNPs are also associated with other diseases, including cardiovascular disease and kidney transplantation.
Figure 1.
Genetic polymorphisms in human CYP4F2 and CYP4A11. Localization of single nucleotide polymorphisms (SNPs) in genomic DNA of CYP4F2 (upper panel, NG_007971) and CYP4A11 (lower panel, NG_007932.1). Numbers indicate the order of exons. Solid portions of exons indicate transcription regions. Open frames on exons indicate non-transcription regions of CYP4F2 and CYP4A11. Published RefSNPs (rs) numbers linked with hypertension and stroke are listed. Asterisks (*) indicate that the SNPs are also associated with other diseases, including cardiovascular disease and kidney transplantation.

Figure 2.
Role of 20-HETE in the progression of ischemic and hemorrhagic strokes. Ischemic stroke leads to hypoxia and ATP depletion, and failure of the ATPase ion pumps within the ischemic core. These effects result in accumulation of intracellular sodium and calcium ions. Enhanced calcium influx activates phospholipase A2 (PLA2) to release arachidonic acid (AA) from the membrane, which is converted into 20-hydroxyeicosatetraenoic acid (20-HETE) by CYP4A enzymes. Clotting blood formed after hemorrhagic stroke releases free hemoglobin and serotonin (5-HT). Free hemoglobin scavenges nitric oxide (NO), and the fall in NO level promotes the activity of CYP4A enzymes. 5-HT stimulates PLA2 and releases AA to increase 20-HETE production. The rise of 20-HETE acts on vascular smooth muscle cells leading to vasospasm, further restricting the blood flow to the stroke site and inducing neuron damage. 20-HETE also affects neurons directly by increasing oxidative stress causing expansion of the infarct after ischemic stroke.
Figure 2.
Role of 20-HETE in the progression of ischemic and hemorrhagic strokes. Ischemic stroke leads to hypoxia and ATP depletion, and failure of the ATPase ion pumps within the ischemic core. These effects result in accumulation of intracellular sodium and calcium ions. Enhanced calcium influx activates phospholipase A2 (PLA2) to release arachidonic acid (AA) from the membrane, which is converted into 20-hydroxyeicosatetraenoic acid (20-HETE) by CYP4A enzymes. Clotting blood formed after hemorrhagic stroke releases free hemoglobin and serotonin (5-HT). Free hemoglobin scavenges nitric oxide (NO), and the fall in NO level promotes the activity of CYP4A enzymes. 5-HT stimulates PLA2 and releases AA to increase 20-HETE production. The rise of 20-HETE acts on vascular smooth muscle cells leading to vasospasm, further restricting the blood flow to the stroke site and inducing neuron damage. 20-HETE also affects neurons directly by increasing oxidative stress causing expansion of the infarct after ischemic stroke.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shekhar, S.; Varghese, K.; Li, M.; Fan, L.; Booz, G.W.; Roman, R.J.; Fan, F. Conflicting Roles of 20-HETE in Hypertension and Stroke. Int. J. Mol. Sci. 2019, 20, 4500. https://doi.org/10.3390/ijms20184500
AMA Style
Shekhar S, Varghese K, Li M, Fan L, Booz GW, Roman RJ, Fan F. Conflicting Roles of 20-HETE in Hypertension and Stroke. International Journal of Molecular Sciences. 2019; 20(18):4500. https://doi.org/10.3390/ijms20184500
Chicago/Turabian StyleShekhar, Shashank, Kevin Varghese, Man Li, Letao Fan, George W. Booz, Richard J. Roman, and Fan Fan. 2019. "Conflicting Roles of 20-HETE in Hypertension and Stroke" International Journal of Molecular Sciences 20, no. 18: 4500. https://doi.org/10.3390/ijms20184500
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.