Crosstalk between Oxidative Stress and Tauopathy



1
Molecular Recognition Research Center, Korea Institute of Science and Technology (KIST), Seoul 02792, Korea
2
Bio-Med Division, KIST-School UST, Seoul 02792, Korea
3
Convergence Research Center for Diagnosis, Treatment and Care System of Dementia, Brain Science Institute (BSI), Korea Institute of Science and Technology (KIST), Seoul 02792, Korea
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Mol. Sci. 2019, 20(8), 1959; https://doi.org/10.3390/ijms20081959
Submission received: 2 April 2019
/
Revised: 16 April 2019
/
Accepted: 19 April 2019
/
Published: 22 April 2019
(This article belongs to the Special Issue Oxidative Stress in Neurodegenerative Diseases: Amyloid, Extracellular Vesicles and Biomarkers)
Abstract
:Tauopathy is a collective term for neurodegenerative diseases associated with pathological modifications of tau protein. Tau modifications are mediated by many factors. Recently, reactive oxygen species (ROS) have attracted attention due to their upstream and downstream effects on tauopathy. In physiological conditions, healthy cells generate a moderate level of ROS for self-defense against foreign invaders. Imbalances between ROS and the anti-oxidation pathway cause an accumulation of excessive ROS. There is clear evidence that ROS directly promotes tau modifications in tauopathy. ROS is also highly upregulated in the patients’ brain of tauopathies, and anti-oxidants are currently prescribed as potential therapeutic agents for tauopathy. Thus, there is a clear connection between oxidative stress (OS) and tauopathies that needs to be studied in more detail. In this review, we will describe the chemical nature of ROS and their roles in tauopathy.
1. Tau Protein and its Pathogenicity
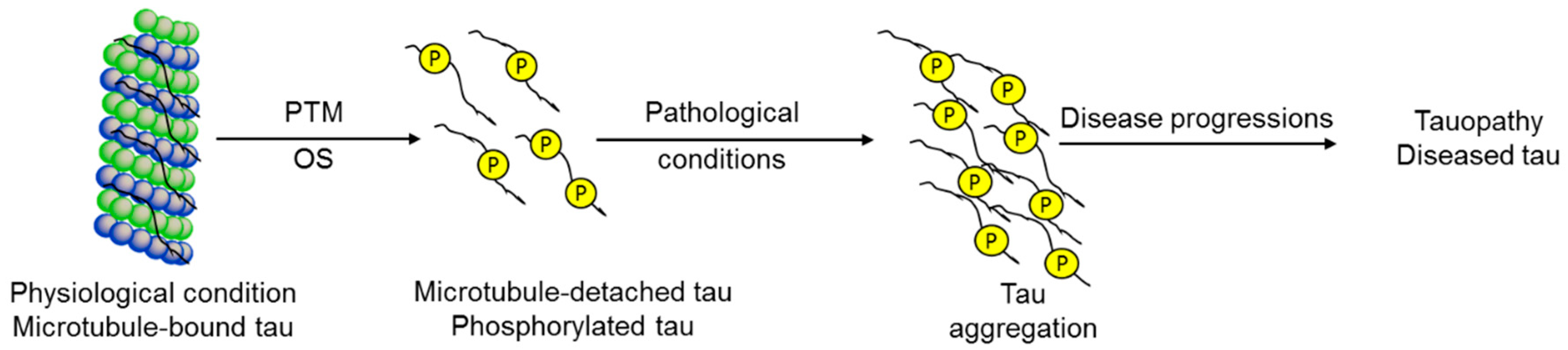
Tau protein is expressed abundantly in neurons as well as sparsely in non-neuronal cells like astrocytes and oligodendrocytes [1]. It is a microtubule-binding protein that gives microtubules’ integrity, which is critical for neuronal outgrowth [2,3,4]. It helps microtubules to anchor with other cytoskeletal filaments and organelles for structural support [5,6]. Microtubules are continuously assembled and disassembled in cells in a dynamic fashion, and this is maintained by the interaction between tau and the microtubule, which is tightly controlled by several factors. Modification of tau affects microtubule stabilization and other processes related to this protein [7]. Tau modification is promoted by post-translational modifications, conformational changes and the misfolding structure of tau. These modifications lead to the abnormal aggregation of tau into neurofibrillary tangle (NFT) structures. These NFTs accumulate in neurons, causing neuronal degeneration. Therefore, the formation of NFTs represents the significant pathological signatures in many neurodegenerative diseases classified as tauopathies [8]. The level of NFTs and tau modifications are correlated to the severity of the tauopathies, including Alzheimer’s disease (AD), Parkinson’s disease (PD), frontotemporal dementia (FTD), FTD with parkinsonism linked to chromosome-17 (FTDP-17), frontotemporal lobar degeneration (FTLD), Pick’s disease (PiD), progressive supranuclear palsy (PSP), corticobasal neurodegeneration (CBD), dementia pugilistica, etc. [9,10,11,12].
2. Causes of Tauopathies
Consequently, researchers have been studying the mechanism of tau pathogenesis. Tau is naturally a highly soluble protein, and it undergoes several modifications to become an aggregate [13,14]. The mechanisms for NFT formation from tau are still in debate today. Among them, aberrant posttranslational modifications (PTM) are the leading cause of this failure. In this regard, hyperphosphorylation, oxidation, proteolytic cleavage (truncation), acetylation, glycation, nitration, and conformational changes have been suggested to cause the neuro-pathogenicity of tau [13,15,16,17,18]. Apart from these hypotheses, imbalances in oxido-redox homeostasis, which produce reactive oxygen species (ROS), play significant roles in tauopathies.
3. Oxidative Stress and Its Relation to Tauopathies
ROS are oxygen-containing reactive molecules that are generated by oxidative stress (OS). A moderate level of ROS is critical in cellular defense mechanisms to fight against foreign subjects, and it triggers mitogen-activated protein kinase (MAPK) pathways to modulate cellular signaling (cell cycle, gene expression, cell survival and apoptosis) [19,20]. In normal physiological conditions, cells produce small amounts of ROS, and the levels of ROS are balanced by several antioxidant systems [21]. The imbalance between ROS generation and antioxidant defense causes the excessive accumulation of ROS, giving OS to the cells [21,22]. Thus, OS poses a significant threat to the brain, one of the most metabolically active organs, which is vulnerable to OS due to its high oxygen demand [23], abundance of the redox-active metals (iron or copper) [23], polyunsaturated fatty acids (substrates for lipid peroxidation) [24], and deficiency of the glutathione (GSH, an antioxidant to eliminate ROS) levels [25].
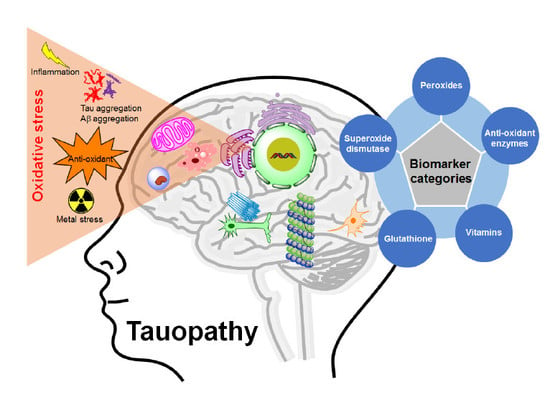
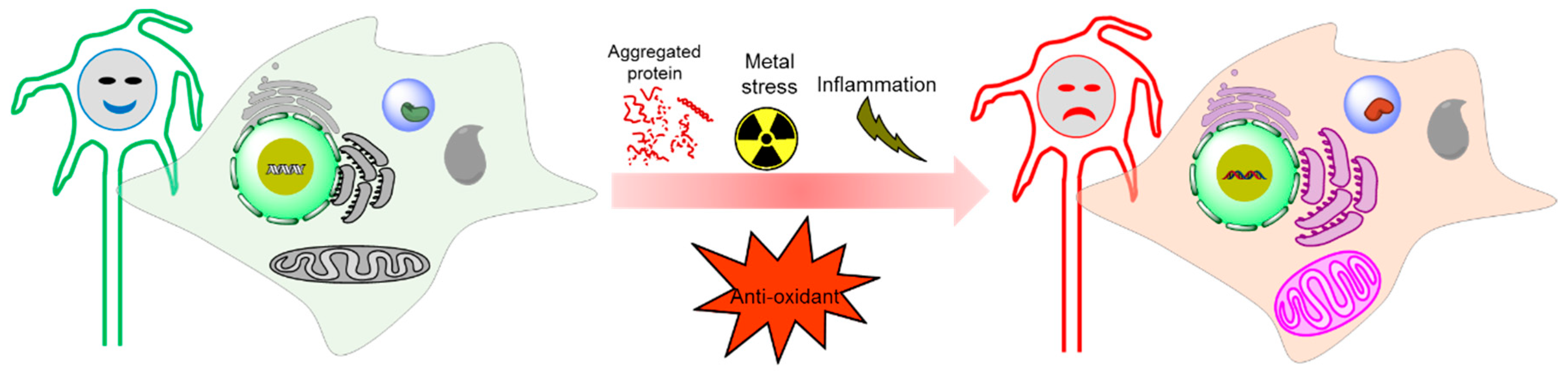
In age-related neurodegenerative diseases, balances between OS and antioxidant enzymes are distorted, resulting in various brain damages and neuronal death. There is increasing evidence that OS is one of the leading pathophysiological markers of tauopathies, and all of these findings suggests that there is a clear relationship between OS and the pathophysiology of tauopathies (Figure 1). Moreover, a series of studies have been focused on the elucidation of the mechanisms underlying ROS linked to tauopathies. However, it has not yet been fully understood whether OS is an early causal factor or a result of the cell injuries induced by tau modifications. Therefore, OS creates a scope for the development of therapeutic strategies for tauopathies. Here, we will discuss the cellular origin, reaction mechanism, and relation of ROS in tauopathies.
There is clear evidence that OS contributes to neurological deterioration, as well as the oxidative destruction of nucleic acids, proteins, or lipids in the central nervous system (CNS) in tauopathies. OS mediated ROS production is involved in protein oxidation (glycoxidation) or lipid oxidation (lipoxidation) and forms stable advanced end products. These protein products are evident in NFTs in AD, whereas lipid products are present in neurofibrillary pathologies [26]. OS damages nucleic acid (DNA or RNA), and the guanine base is the most susceptible base for oxidative modifications (8-hydroxy-2’-deoxyguanosine, 8-OHdG or 8-hydroxy-2’-guanosine, 8-OHG). These damages by OS are another phenomenon that is associated with tauopathies [27].
AD is the best-known tauopathy, among others, with increasing prevalence. Accumulation of OS is directly linked to aging, and intimately related to AD [28]. In fact, OS is one of the first observable markers in AD progression, even before the appearance of amyloid beta (Aβ) accumulation [29]. Tau overexpressed cells show increased vulnerability to OS [30,31]. Furthermore, mice (P301S and P301L) with AD showed mitochondrial dysfunction, which is associated with increased ROS [32,33]. Several studies indicate that Aβ induces OS, where Aβ serves as a source of ROS to initiate lipid peroxidation [34]. Inversely, Aβ level is increased upon the stimulus of OS but diminishes with time [35]. These phenomena indicate that not only ROS modulates Aβ production, but also Aβ generates excessive ROS in cases of AD. Another report states that free radicals affected the nature and function of neural cells, both in AD and PD [28]. PD is another well-known tauopathy of which OS is the leading contributor. OS is believed to be the main etiology that leads to idiopathic and genetic causes of PD [28]. Alterations in the antioxidant molecules were evident in the early stages of PD [36].
4. Oxidative Stress and Reactive Oxygen Species
ROS are generated upon the reduction of O2, and they consist of free radicals and non-radical species. In particular, ROS that play critical roles in the biological system include superoxide anion radical (O2− or O2•−), hydroperoxyl radical (HO2), hydrogen peroxide (H2O2), hydroxyl radical (•OH), ozone (O3), and singlet oxygen (1O2). Further, ROS also includes peroxyl (ROO•), alkoxyl (RO•), semiquinone (SQ•−), and carbonate (CO3•−) radicals, along with hypochlorous (HOCl), hypobromous (HOBr), and hypoiodous acids (HOI). Some of the reactive nitrogen species (RNS), which contain an oxygen atom, are also considered as one of these kinds, like the nitric oxide radical (NO or NO•), nitrogen dioxide radical (NO2•), nitrite (NO2−), and peroxynitrite (ONOO−) [37,38].
5. Major Sources of Oxidative Stress (OS)
The endogenous sources of OS are: (i) Hypoxia, NO and ONOO, which endorse the generation of ROS in mitochondrial electron transport chain (ETC) [37,39,40]; (ii) an increased level of misfolded proteins [41]; and (iii) intracellular enzymes that produce ROS as metabolic products of their enzymatic processes. These enzymes include NADPH oxidases, flavoenzyme ERO1, cytochrome p450, lipoxygenases, xanthine oxidase, nitric oxide synthases, etc. [37,38,40] Lastly, (iv) free metal cations (such as copper and iron ions) can convert superoxide or hydrogen peroxide into hydroxyl radicals through the Fenton reaction or the Haber–Weiss reaction [42]. A high level of intracellular Ca2+ is also one of the endogenous sources of OS [43,44]. The exogenous sources are ultraviolet light (1O2 generation), γ-irradiation (•OH generation) [45], chemical pollutants, including quinones, nitroaromatics, etc. These sources generate superoxides. It is also reported that smoke and air pollutants also contain ROS which potentially uptake into the body during the respiration process and induce OS [37,46].
6. Reactive Oxygen Species (ROS) Production in the Body
6.1. Mitochondria
Mitochondria are the major sub-cellular organelles for ROS production. ROS are produced as a byproduct of the electron transport chain reaction for adenosine triphosphate (ATP) generation. Loss of some mitochondrial proteins (cytochrome oxidase, COX; pyruvate dehydrogenase complex, PDHC; and α-ketoglutarate dehydrogenase complex, KGDHC) elevate the formation of ROS [47]. OS induces mitochondrial dysfunction by accumulating excessive ROS [30,48]. Increased oxidative damage may lead to mitochondrial DNA mutations, which has been reported to inhibit ATP production in AD patients [30]. In addition to mitochondria, endoplasmic reticulum, peroxisomes, and microglial cells are also potential sources of ROS.
6.2. Endoplasmic Reticulum
The endoplasmic reticulum (ER) is mainly responsible for protein folding and lipid biosynthesis. In the ER, ROS generation is induced by misfolded protein processing, depletion of the GSH, or through the breakage and formation of disulfide bonds [49].
6.3. Peroxisomes
Peroxisomes participate in many metabolic pathways that include fatty acid oxidation, amino acid catabolism, and phospholipid biosynthesis. They generate the majority of the H2O2 inside the body, which is balanced by catalases. This H2O2 level is increased when catalase is not working, or when peroxisomes are damaged. Peroxisomes have been shown to be depleted or damaged by tau aggregation in the rat primary hippocampal neurons, and N2a cells induce OS [31].
6.4. Microglia
Microglial cells express a high level of the glutathione peroxidase (GPx) antioxidant enzyme. They also produce ROS, either to eliminate pathogens or during neuroinflammatory processes. Recently, microglial cells were reported as ROS producers in tauopathies [11].
7. Chemical Properties of ROS
To gain a clearer perception of the ROS associated biological impact, one needs to understand the chemical properties of ROS.
7.1. Electronic Configuration of ROS
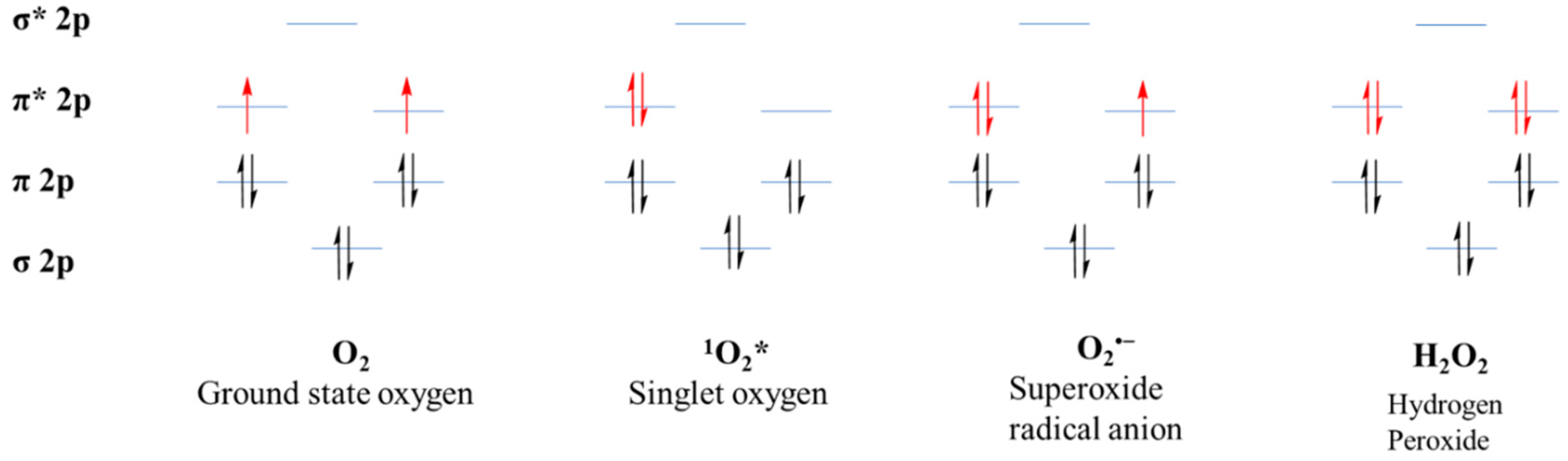
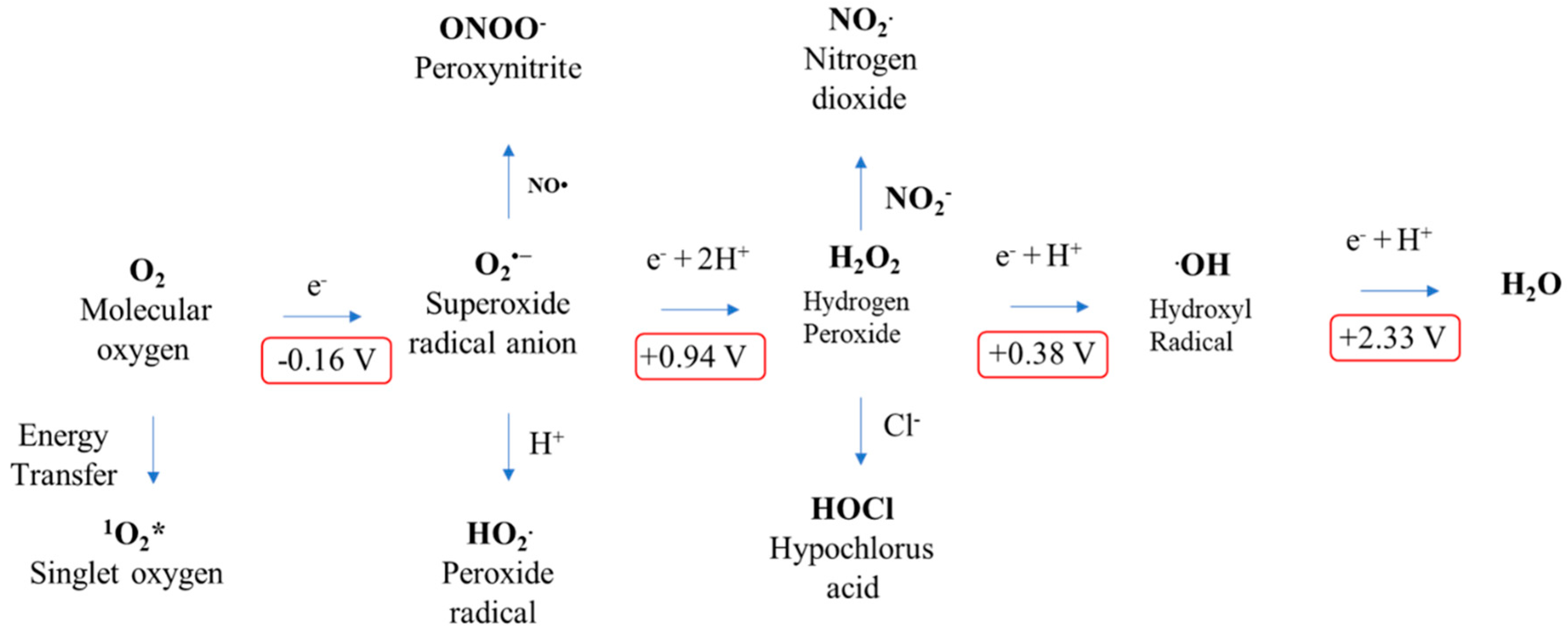
Molecular oxygen is paramagnetic species, having two electrons with a parallel spin in the π* orbital (Figure 2). Due to this kind of parallel spin, restriction is applied to an oxygen atom to take part in redox reactions. This kind of electronic configuration enables the oxygen atom to accept electrons during the redox reactions, making it unable to oxidize the biomolecules [38]. The molecular oxygen can be converted quite easily into ROS by either energy transfer or electron transfer. Among the ROS, energy transfer from oxygen results in the generation of singlet oxygen. As a result, singlet oxygen has paired electrons with opposite spin, which enhance the oxidizing ability when compared with molecular oxygen. By a one-electron reduction, O2 is converted into superoxide. Further, superoxide, with another one-electron reduction, converts into hydrogen peroxide which later converts into the hydroxyl radical by a one-electron reduction and finally the hydroxyl radical, upon reduction, converts into water. Hence, molecular oxygen can generate ROS by electron or energy transfer, making them more reactive than itself [50,51,52].
7.2. Reactivity Trends of ROS
To gain more in-depth knowledge into the reactivity of ROS, one should understand the thermodynamics of free radicals. To compare the trend of reactivity of the ROS, the role of reduction potential is vital. To plot the reactivity trend of ROS, one should consider the reduction potential of the one-electron reduction of molecular oxygen [38,50]. A few of the main features about the role of reduction potential and reactivity are listed as follows: (a) Reduction potential is the ability of an atom or molecule to acquire the electron; (b) oxidation is the loss of electron, whereas reduction is the gain of electron; and (c) from the above points, it is evident that atoms or molecules with high reduction potential have high reactivity as will be easy to reduce. Based on the above assumption, one can easily understand why molecular oxygen is a weak oxidant when compared with another ROS (Figure 3). Here, O2, with the reduction potential of −0.33 V, is a poor electron acceptor [53]. The superoxide radical also has limited reactivity when compared with its anionic charge, which makes it an electron-rich center [44]. Hydrogen peroxide is generated by the one-electron reduction of superoxide, which is also stable under physiological conditions, even though it has a +0.38 V reduction potential [52]. Among the ROS, the hydroxyl radical is the utmost powerful oxidizing agent, with a one-electron reduction potential of +2.33 V. According to the reduction potential, the ROS reactivity trend can be written as •OH > O2•− > H2O2 > O2 [50,52].
8. ROS Generating Agents in Tauopathies
8.1. Tau Aggregation
Tau aggregate is one of the primary culprits in tauopathies. There is clear evidence among OS with tau hyperphosphorylation, polymerization, and toxicity in both animal models and patients [11]. OS produces oxidized fatty acids that stimulate in vitro tau polymerization [54]. Also, the overexpressed tau protein showed increased OS in N2a cultured cells [31]. Besides, rat cortical neurons from truncated tau expressing transgenic rats showed an increased level of ROS [55]. This evidence suggests that OS directly promotes tau aggregation, and inversely toxic tau species stimulate OS conditions in tauopathies.
8.2. Amyloid-Beta Aggregation
Amyloid-beta (Aβ) aggregation is another pathogenic event in tauopathies. Numerous studies have shown that OS promotes Aβ production by diminishing α-secretase activity and increasing β and γ-secretase activity [56]. The AD mouse model (Tg2576-APP-PS1) has enhanced OS due to increasing H2O2 levels and the peroxidation of proteins and lipids [57]. Oppositely, the accumulation of Aβ increases OS and leads to memory dysfunction in the AD mouse (Tg2576-APP) [58] and mitochondrial failure in the early stages of AD [30].
8.3. Metals
Copper (Cu), zinc (Zn) and iron (Fe) are the three most abundant metals in mammalian brains, which regulate many synaptic functions. Aberrant homeostasis of these metals, like their levels, mislocalization, and dysregulation, was evidenced in the hippocampus and amygdala of AD patients [59]. In addition, aluminum (Al3+) is also associated with oxidative stress leading to AD [60].
8.4. Inflammation
Neuro-inflammation is expected in sites of metal deposition. Neuro-inflammation is followed by the production of reactive compounds by microglial cells. The microglia can be activated by several factors, including Aβ. Activated microglia play a role in AD onset by increased ROS burden and oxidative stress [61].
8.5. Anti-Oxidant
9. Evaluation of ROS in Tauopathies
Since OS is related to the pathophysiological mechanism in tauopathies (Figure 4), several surrogate markers for ROS have been assessed. The measuring parameters mainly cover peroxides including nucleic acid (8-OGH and 8-OHdG, protein or lipid (malondialdehyde, MDA or 4-hydroxynonenal, 4-HNE), antioxidant enzymes, superoxide dismutase (cytosolic SOD1 or mitochondrial SOD2), glutathione levels, glutathione S transferase (GST), heme Oxygenase-1 (HO-1), homocysteine (Hcy) levels, F2-isoprostanes (F2-IsoPs) and vitamins (A, C, or E). These markers are elevated or depleted in particular brain regions of the same patient in tauopathies. A table (Table 1) is provided here based on the available literature for potential OS markers in assessing tauopathies.
10. Fluorescence Probes for ROS Detection
Fluorescent probes have advantages for high sensitivity and selectivity over other techniques. Combined with fluorescent microscopy, the fluorescent probes can be used to visualize analytes in living cells spatiotemporally. Also, this technique is fast and straightforward, without a unique facility or high cost-efficiency. Fluorescent sensors (organic fluorophores), when recognizing the target molecule, tend to change their fluorescence emission intensity or color [94,95,96,97]. Here we would like to emphasize some of the basic concepts of the ROS probing based on the fluorescent technique (Table 2).
Fluorescent probes for hydrogen peroxide are mainly based on boronate esters via the H2O2-induced oxidation of arylboronate ester to phenols [98,99,100]. It is known that under mild alkaline conditions H2O2 reacts with arylboronic acids and arylboronate ester to produce phenols [101].
Most of the probes for the superoxide radical are based on reaction-based mechanisms, where the superoxide will react with a probe to generate a new molecule with a different fluorescence intensity or color of the probe. These probes are based on superoxide-induced oxidation reactions and reactions with nitroxide [102]. Another essential feature of the superoxide is strong nucleophilicity, where it is known as a super nucleophile. Due to this inherent property, fluorescent probes for superoxide are mainly based on nucleophilic substitution reaction [103]. In this case, most of the time, the leaving group is from the phosphinate group [104], triflate group [105], 2,4-dinitrobenzenesulfonyl [106], nitro-ethers [107], and related groups. Recently, a sulfinate-based chemosensor for superoxide sensing was reported to reduce the molecular size, with good solubility and a cLogP value of 3.8 [108].
Detection of the hydroxyl radical is very challenging due to its low concentration. It is the most reactive among the various classes of the ROS. The hydroxyl radical can oxidize most of the biomolecules like carbohydrates, proteins, and nucleic acids. One of the main classes for hydroxyl radical detection is the oxidation of leuco forms of fluorescent dyes (cyanine, 2,7-dichlorodihydrofluorescein, and dihydrorhodamine) mediated by hydroxyl radical. These probes become highly fluorescent upon reaction with a hydroxyl radical by recovering the π-conjugation networks through hydrogen atom abstraction, and subsequently one-electron oxidation [109,110,111,112]. Some of the nitroxides have been exploited as fluorescent probes for hydroxyl radicals. Here, the diamagnetization reaction between nitroxide and radical restores the fluorescence intensity was exploited, where the CH3 radical (generated from hydroxyl radical and dimethyl sulfoxide, DMSO) reacts with the nitroxide moiety to produce the diamagnetic adduct [95,113].
Singlet oxygen is produced by energy transfer from O2. As a result, singlet oxygen has paired electrons with opposite spin (spin-restriction state) which enhance oxidizing ability when compared with molecular oxygen. The fluorescent probes for singlet oxygen have been developed based on [2 + 4] cycloaddition, where singlet oxygen acts as a strong dienophile [109,114,115].
Most of the probes for peroxynitrite are based on the organic reactions of probes with peroxynitrite. These reactions include the oxidation of chalcogenides, boronic acids or boronates, hydrazides, the cleavage of C-C double bonds, and oxidative N-dearylation [116,117]. Among these, the most common reaction is the oxidation of chalcogenides (S, Se, and Te) [118,119].
Different types of functional groups have been studied for the detection of NO, which are o-diamino aromatic compounds, luminescent lanthanide complexes, transition-metal complexes, quantum dots, and carbon nanotube sensors. The most commonly studied method for the detection of NO is based on the reaction of a O-diaminophenyl group with NO to generate triazole [120,121].
The fluorescent probes for hypochlorite are mostly based on xanthene probes. The mechanism behind this sensing is the spiro-ring opening of the xanthene probes, which react with hypochlorite [95]. Also, some of the probes are based on the oxidation of catechol to benzoquinone [122].
Apart from these probes, some of the fluorescent probes for the detection of ROS are reported in Alzheimer’s disease. An oxalate-curcumin-based probe was reported for the imaging of reactive oxygen species in AD. Here, the oxalate moiety was utilized, which reacts with H2O2 to produce the fluorescence signal with a shift in wavelength [123,124,125,126]. A bifunctional fluorescent probe based on the benzothiazole core has been reported for H2O2 and amyloid aggregate detection. The probe is a combination of benzothiazole and a boronate ester [127].
11. Controlling of ROS as Therapeutic Approaches of Tauopathies
Despite the unmet needs for the treatment of tauopathies, FDA-approved treatments are limited to alleviate memory deficits and behavioral changes. Despite the fact that extensive drug discovery programs have been conducted over previous decades, there are still no convincing drugs. Some drugs have adverse side effects or exhibit a lack of cognitive improvement in trial participants. For these reasons, new strategies for therapeutic pathways are being considered. Potent pieces of evidence for the imbalance phenomena of oxidants/antioxidants in AD led to the hypothesis that compounds scavenging free radicals or upregulating the OS defense mechanism might provide therapeutic approaches for AD (Table 3).
11.1. Antioxidant Pathway
By reviewing the OS theories in tauopathies, several therapeutic approaches have been conducted in different tauopathies. Curcumin, found in turmeric, is an antioxidant that decreases Aβ-induced tau hyperphosphorylation in PC12 cells. It also protects PC12 cells through the inhibition of OS [131]. Another class of antioxidant, methylene blue (methylthioninium chloride, MB), which can penetrate the blood brain barrier, diminished oxidized nucleic acids and tau hyperphosphorylation in tauP301S transgenic mice [132]. Administration of coenzyme Q10 (CoQ10; a critical member in electron transport chain), which has antioxidant effects, can significantly reduce tau phosphorylation, lipid peroxidation and OS, while ameliorating behavioral deficits and the survival rate of tauP301S transgenic mice [133]. The overexpression of thioredoxin peroxidase, an antioxidant enzyme, worsens disease phenotypes of tauR406W transgenic drosophila [66]. In PD mice, sulforaphane (found in cruciferous vegetables) were found to be effective at protecting dopaminergic neurons and enhancing GSH levels [134]. Brilliant blue G, an antagonist of the P2X7R pathway of several neurodegeneration, can cross the blood brain barrier to ameliorate neuropathology in AD and PD mouse models [135].
11.2. Catalase
Catalase, located in peroxisomes, cytoplasm, and mitochondria, is responsible for H2O2 conversion into water and oxygen by using iron or manganese as a cofactor. The administration of SOD/catalase mimetic EUK-207 deteriorates disease phenotypes by reducing phosphorylated tau and lipid peroxidation in 3xTg-AD mice [136].
11.3. Vitamin
Cells and animal studies, as well as clinical studies, have shown a particular connection between vitamins and tauopathies. The antioxidant activities of vitamins may be useful for the treatment of tauopathies. Proper supplementation of vitamins can reduce the tauopathy incidence in the general population and improve the state of patients. Vitamin C is a water-soluble antioxidant that is abundant in vegetables, fruits, and animal livers. It involves in the inhibition of OS, reducing lipid peroxidation, the exclusion of free radicals, and acts as a cofactor for antioxidant enzymes [137]. Vitamin E is a lipid-soluble high antioxidant which can diminish the effects of peroxide, also protecting against lipid peroxidation in cell membranes. Vitamin C (ascorbic acid), when used to treat primary corticohippocampal neurons from tau transgenic rats (Tau151–391 4R), decreased ROS levels in the neurons, tau inclusions in the spinal cord, and improved behavior [138]. Supplementation of vitamin E (α-tocopherol) delayed tau pathology by attenuating motor weakness in the tau mouse model (B6D2/F1-Tau44) [139]. A recent study has shown that omega-3 fatty acids plus vitamin E supplementation can improve total antioxidant capacity and GSH levels in PD patients [140].
11.4. Metal Chelator
Alterations in metallostasis (mainly Zn, Cu and Fe) are responsible for AD. So, ionophore addition would be another treatment choice by restoring ion balances. Clioquinol (5-chloro-7-iodo-quinoline-8-ol) is a moderate chelator for Cu, Zn, and Fe that rescued memory impairment in the TgCRND8-AD mouse after oral administration [141]. The second generation of clioquinol is PBT2, which is now in a clinical trial (phase II) [142]. Both of their modes of action probably rely on copper ionophore activity. Based on this mechanism, another copper containing chelator, CuIIGTSM [bis(thiosemicarbazone)], was developed, which could change GSK-3β activity, tau phosphorylation, and restore cognitive impairment in APP/PS1 transgenic mice [143]. One Fe chelator, desferrioxamine, was intramuscularly injected into AD patients, slowing the clinical progression of dementia [144].
12. Conclusions
Oxidative stress contributes to the development of tauopathies. It forms vicious pathophysiology inducing mitochondrial dysfunctions, neuronal damages and promotes metal toxicity. The complex nature, genesis, and responses of ROS in tauopathies are still actively being investigated. At the cellular level, individual cells generate excessive levels of ROS, followed by a dysfunctional state of mitochondria that interacts with redox metals and oxidative stress-responsive elements. Although the formation of ROS poses a threat to tauopathies, compensatory responses provoked by ROS removal or the prevention of the ROS generation pathway may interrupt the onset or slow down the progression of tauopathies through multiple mechanisms. These mechanisms include the reduction of oxidative stress-mediated neuronal toxicity, a decrease of tau phosphorylation and aggregation, restoration of mitochondrial function, and metal homeostasis. Therefore, treatment with antioxidants could be an alternative approach to target molecular events implicated in tauopathies. However, evidence on antioxidants as potential therapeutic agents for tauopathies has not been carried out to a significant level. Hence, a more in-depth understanding of OS in tauopathies is in high demanded.
Funding
This work was supported by the National Research Foundation (NRF-2018M3A9H4079286), Brain Research Program (2016M3C7A1913845), and the National Research Council of Science and Technology (NST) (No. CRC-15-04-KIST) funded by the Ministry of Science, ICT & Future Planning. Dr. Haque is a recipient of the Korea Research Fellowship Program through the National Research Foundation of Korea (NRF) (NRF-2018H1D3A1A02074556).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| •OH | Hydroxyl radical |
| 1O2 | Singlet oxygen |
| 8-OHdG | 8-Hydroxy-2’-deoxyguanosine |
| 8-OHG | 8-Hydroxy-2’-guanosine |
| Aβ | Amyloid beta’ Abeta |
| AD | Alzheimer’s disease |
| Al | Aluminium |
| ATP | Adenosine triphosphate |
| CBD | Corticobasal degeneration |
| CNS | Central nervous system |
| CO3•− | Carbonate radical |
| CoQ10 | Coenzyme Q10 |
| COX | Cytochrome oxidase |
| Cu | Copper |
| DMSO | Dimethyl sulfoxide |
| ER | Endoplasmic reticulum |
| ETC | Electron transport chain |
| FDA | US Food and Drug Administration |
| Fe | Iron |
| FTD | Frontotemporal dementia |
| FTDP-17 | FTD with parkinsonism linked to chromosome-17 |
| FTLD | Frontotemporal lobar degeneration |
| GPx | Glutathione peroxidase |
| GSH | Glutathione |
| GST | Glutathione S transferase |
| H2O2 | Hydrogen peroxide |
| Hcy | Homocysteine |
| 4-HNE | 4-Hydroxynonenal |
| HO-1 | Heme oxygenase-1 |
| HO2 | Hydroperoxyl radical |
| HOBr | Hypobromous |
| HOCl | Hypochlorous |
| HOI | Hypoiodous acids |
| Hr | Hour |
| KGDHC | α-Ketogluterate dehydrogenase complex |
| MAPK | Mitogen-activated protein kinase |
| MB | Methylene blue; methylthioninium chloride |
| NFT | Neurofibrillary tangle |
| NO or NO• | Nitric oxide radical |
| NO2− | Nitrite |
| NO2• | Nitrogen dioxide radical |
| O2− or O2•− | Superoxide anion radical |
| O3 | Ozone |
| ONOO− | Peroxynitrite |
| OS | Oxidative stress |
| PD | Parkinson’s disease |
| PDHC | Pyruvate dehydrogenase complex |
| PiD | Pick’s disease |
| PSP | Progressive supranuclear palsy |
| PTM | Posttranslational modification |
| RNS | Reactive nitrogen species |
| RO• | Alkoxyl radical |
| ROI | Reactive oxygen intermediate |
| ROO• | Peroxyl radical |
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
| SQ•− | Semiquinone radical |
| Zn | Zinc |
References
- Shin, R.W.; Iwaki, T.; Kitamoto, T.; Tateishi, J. Hydrated autoclave pretreatment enhances tau immunoreactivity in formalin-fixed normal and Alzheimer’s disease brain tissues. Lab. Investig. 1991, 64, 693–702. [Google Scholar]
- Cleveland, D.W.; Hwo, S.Y.; Kirschner, M.W. Physical and chemical properties of purified tau factor and the role of tau in microtubule assembly. J. Mol. Biol. 1977, 116, 227–247. [Google Scholar] [CrossRef]
- Kolarova, M.; Garcia-Sierra, F.; Bartos, A.; Ricny, J.; Ripova, D. Structure and pathology of tau protein in Alzheimer disease. Int. J. Alzheimers Dis. 2012, 2012, 731526. [Google Scholar] [CrossRef]
- Mazanetz, M.P.; Fischer, P.M. Untangling tau hyperphosphorylation in drug design for neurodegenerative diseases. Nat. Rev. Drug Discov. 2007, 6, 464–479. [Google Scholar] [CrossRef] [PubMed]
- Miyata, Y.; Hoshi, M.; Nishida, E.; Minami, Y.; Sakai, H. Binding of microtubule-associated protein 2 and tau to the intermediate filament reassembled from neurofilament 70-kDa subunit protein. Its regulation by calmodulin. J. Biol. Chem. 1986, 261, 13026–13030. [Google Scholar]
- Jung, D.; Filliol, D.; Miehe, M.; Rendon, A. Interaction of brain mitochondria with microtubules reconstituted from brain tubulin and MAP2 or TAU. Cell Motil. Cytoskeleton 1993, 24, 245–255. [Google Scholar] [CrossRef]
- Mandelkow, E.M.; Stamer, K.; Vogel, R.; Thies, E.; Mandelkow, E. Clogging of axons by tau, inhibition of axonal traffic and starvation of synapses. Neurobiol. Aging 2003, 24, 1079–1085. [Google Scholar] [CrossRef] [PubMed]
- Obulesu, M.; Venu, R.; Somashekhar, R. Tau mediated neurodegeneration: An insight into Alzheimer’s disease pathology. Neurochem. Res. 2011, 36, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Buee, L.; Bussiere, T.; Buee-Scherrer, V.; Delacourte, A.; Hof, P.R. Tau protein isoforms, phosphorylation and role in neurodegenerative disorders. Brain Res. Brain Res. Rev. 2000, 33, 95–130. [Google Scholar] [CrossRef]
- Spillantini, M.G.; Goedert, M. Tau pathology and neurodegeneration. Lancet Neurol. 2013, 12, 609–622. [Google Scholar] [CrossRef]
- Alavi Naini, S.M.; Soussi-Yanicostas, N. Tau Hyperphosphorylation and Oxidative Stress, a Critical Vicious Circle in Neurodegenerative Tauopathies? Oxid. Med. Cell Longev. 2015, 2015, 151979. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, F.; Wang, D.; Li, C.; Fu, Y.; He, W.; Zhang, J. Tau Pathology in Parkinson’s Disease. Front. Neurol. 2018, 9, 809. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.M.; Kim, D.; Yu, Y.H.; Lim, S.; Kim, D.J.; Chang, Y.T.; Ha, H.H.; Kim, Y.K. Inhibition of tau aggregation by a rosamine derivative that blocks tau intermolecular disulfide cross-linking. Amyloid 2014, 21, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.; Haque, M.M.; Kim, D.; Kim, D.J.; Kim, Y.K. Cell-based Models To Investigate Tau Aggregation. Comput. Struct. Biotechnol. J. 2014, 12, 7–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrell, R.W.; Gooptu, B. Conformational changes and disease—Serpins, prions and Alzheimer’s. Curr. Opin. Struct. Biol. 1998, 8, 799–809. [Google Scholar] [CrossRef]
- Binder, L.I.; Guillozet-Bongaarts, A.L.; Garcia-Sierra, F.; Berry, R.W. Tau, tangles, and Alzheimer’s disease. Biochim. Biophys. Acta 2005, 1739, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Schweers, O.; Mandelkow, E.M.; Biernat, J.; Mandelkow, E. Oxidation of cysteine-322 in the repeat domain of microtubule-associated protein tau controls the in vitro assembly of paired helical filaments. Proc. Natl. Acad. Sci. USA 1995, 92, 8463–8467. [Google Scholar] [CrossRef] [PubMed]
- Wischik, C.M.; Novak, M.; Edwards, P.C.; Klug, A.; Tichelaar, W.; Crowther, R.A. Structural characterization of the core of the paired helical filament of Alzheimer disease. Proc. Natl. Acad. Sci. USA 1988, 85, 4884–4888. [Google Scholar] [CrossRef]
- Groeger, G.; Quiney, C.; Cotter, T.G. Hydrogen peroxide as a cell-survival signaling molecule. Antioxid. Redox Signal. 2009, 11, 2655–2671. [Google Scholar] [CrossRef]
- Patten, D.A.; Germain, M.; Kelly, M.A.; Slack, R.S. Reactive oxygen species: Stuck in the middle of neurodegeneration. J. Alzheimers Dis. 2010, 20 (Suppl. 2), S357–S367. [Google Scholar] [CrossRef]
- Dasuri, K.; Zhang, L.; Keller, J.N. Oxidative stress, neurodegeneration, and the balance of protein degradation and protein synthesis. Free Radic. Biol. Med. 2013, 62, 170–185. [Google Scholar] [CrossRef]
- Ray, P.D.; Huang, B.W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.H.; Kim, J.E.; Rhie, S.J.; Yoon, S. The Role of Oxidative Stress in Neurodegenerative Diseases. Exp. Neurobiol. 2015, 24, 325–340. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Michaelis, E.K. Selective neuronal vulnerability to oxidative stress in the brain. Front. Aging Neurosci. 2010, 2, 12. [Google Scholar] [CrossRef]
- Ferreira, M.E.; de Vasconcelos, A.S.; da Costa Vilhena, T.; da Silva, T.L.; da Silva Barbosa, A.; Gomes, A.R.; Dolabela, M.F.; Percario, S. Oxidative Stress in Alzheimer’s Disease: Should We Keep Trying Antioxidant Therapies? Cell Mol. Neurobiol. 2015, 35, 595–614. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Lee, H.G.; Casadesus, G.; Avila, J.; Drew, K.; Perry, G.; Smith, M.A. Oxidative imbalance in Alzheimer’s disease. Mol. Neurobiol. 2005, 31, 205–217. [Google Scholar] [CrossRef]
- Chen, Z.; Zhong, C. Oxidative stress in Alzheimer’s disease. Neurosci. Bull. 2014, 30, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Sun, Q.; Chen, S. Oxidative stress: A major pathogenesis and potential therapeutic target of antioxidative agents in Parkinson’s disease and Alzheimer’s disease. Prog. Neurobiol. 2016, 147, 1–19. [Google Scholar] [CrossRef]
- Resende, R.; Moreira, P.I.; Proenca, T.; Deshpande, A.; Busciglio, J.; Pereira, C.; Oliveira, C.R. Brain oxidative stress in a triple-transgenic mouse model of Alzheimer disease. Free Radic. Biol. Med. 2008, 44, 2051–2057. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zhao, B. Oxidative stress and the pathogenesis of Alzheimer’s disease. Oxid. Med. Cell Longev. 2013, 2013, 316523. [Google Scholar] [CrossRef]
- Stamer, K.; Vogel, R.; Thies, E.; Mandelkow, E.; Mandelkow, E.M. Tau blocks traffic of organelles, neurofilaments, and APP vesicles in neurons and enhances oxidative stress. J. Cell Biol. 2002, 156, 1051–1063. [Google Scholar] [CrossRef] [Green Version]
- Yoshiyama, Y.; Higuchi, M.; Zhang, B.; Huang, S.M.; Iwata, N.; Saido, T.C.; Maeda, J.; Suhara, T.; Trojanowski, J.Q.; Lee, V.M. Synapse loss and microglial activation precede tangles in a P301S tauopathy mouse model. Neuron 2007, 53, 337–351. [Google Scholar] [CrossRef]
- David, D.C.; Hauptmann, S.; Scherping, I.; Schuessel, K.; Keil, U.; Rizzu, P.; Ravid, R.; Drose, S.; Brandt, U.; Muller, W.E.; et al. Proteomic and functional analyses reveal a mitochondrial dysfunction in P301L tau transgenic mice. J. Biol. Chem. 2005, 280, 23802–23814. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Swomley, A.M.; Sultana, R. Amyloid beta-peptide (1-42)-induced oxidative stress in Alzheimer disease: Importance in disease pathogenesis and progression. Antioxid. Redox Signal. 2013, 19, 823–835. [Google Scholar] [CrossRef]
- Ashabi, G.; Ahmadiani, A.; Abdi, A.; Abraki, S.B.; Khodagholi, F. Time course study of Abeta formation and neurite outgrowth disruption in differentiated human neuroblastoma cells exposed to H2O2: Protective role of autophagy. Toxicol. In Vitro 2013, 27, 1780–1788. [Google Scholar] [CrossRef]
- Hauser, D.N.; Hastings, T.G. Mitochondrial dysfunction and oxidative stress in Parkinson’s disease and monogenic parkinsonism. Neurobiol. Dis. 2013, 51, 35–42. [Google Scholar] [CrossRef]
- Nathan, C.; Ding, A. SnapShot: Reactive oxygen intermediates (ROI). Cell 2010, 140, 951–951.e2. [Google Scholar] [CrossRef] [PubMed]
- Krumova, K.; Cosa, G. Overview of reactive oxygen species. Singlet Oxygen: Appl. Biosci. Nanosci. 2016, 1, 1–21. [Google Scholar]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, B.C.; Chang, C.J. Chemistry and biology of reactive oxygen species in signaling or stress responses. Nat. Chem. Biol. 2011, 7, 504. [Google Scholar] [CrossRef]
- Chong, W.; Shastri, M.; Eri, R. Endoplasmic reticulum stress and oxidative stress: A vicious nexus implicated in bowel disease pathophysiology. Int. J. Mol. Sci. 2017, 18, 771. [Google Scholar] [CrossRef] [PubMed]
- Walling, C. Fenton’s reagent revisited. Acc. Chem. Res. 1975, 8, 125–131. [Google Scholar] [CrossRef]
- Imlay, J.A. Pathways of oxidative damage. Annu. Rev. Microbiol. 2003, 57, 395–418. [Google Scholar] [CrossRef] [PubMed]
- D’Autréaux, B.; Toledano, M.B. ROS as signalling molecules: Mechanisms that generate specificity in ROS homeostasis. Nat. Rev. Mol. Cell Biol. 2007, 8, 813. [Google Scholar] [CrossRef]
- Leach, J.K.; Van Tuyle, G.; Lin, P.-S.; Schmidt-Ullrich, R.; Mikkelsen, R.B. Ionizing radiation-induced, mitochondria-dependent generation of reactive oxygen/nitrogen. Cancer Res. 2001, 61, 3894–3901. [Google Scholar] [PubMed]
- Winterbourn, C.C. Reconciling the chemistry and biology of reactive oxygen species. Nat. Chem. Biol. 2008, 4, 278. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Raina, A.K.; Lee, H.G.; Casadesus, G.; Smith, M.A.; Perry, G. Oxidative stress signalling in Alzheimer’s disease. Brain Res 2004, 1000, 32–39. [Google Scholar] [CrossRef]
- Radi, E.; Formichi, P.; Battisti, C.; Federico, A. Apoptosis and oxidative stress in neurodegenerative diseases. J. Alzheimers Dis. 2014, 42 (Suppl. 3), S125–S152. [Google Scholar] [CrossRef]
- Bhandary, B.; Marahatta, A.; Kim, H.R.; Chae, H.J. An involvement of oxidative stress in endoplasmic reticulum stress and its associated diseases. Int. J. Mol. Sci. 2012, 14, 434–456. [Google Scholar] [CrossRef]
- Buettner, G.R. The pecking order of free radicals and antioxidants: Lipid peroxidation, α-tocopherol, and ascorbate. Arch. Biochem. Biophys. 1993, 300, 535–543. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Valko, M.; Izakovic, M.; Mazur, M.; Rhodes, C.J.; Telser, J. Role of oxygen radicals in DNA damage and cancer incidence. Mol. Cell. Biochem. 2004, 266, 37–56. [Google Scholar] [CrossRef]
- Wood, P.M. The potential diagram for oxygen at pH 7. Biochem. J. 1988, 253, 287–289. [Google Scholar] [CrossRef] [Green Version]
- Gamblin, T.C.; King, M.E.; Kuret, J.; Berry, R.W.; Binder, L.I. Oxidative regulation of fatty acid-induced tau polymerization. Biochemistry 2000, 39, 14203–14210. [Google Scholar] [CrossRef]
- Cente, M.; Filipcik, P.; Pevalova, M.; Novak, M. Expression of a truncated tau protein induces oxidative stress in a rodent model of tauopathy. Eur. J. Neurosci. 2006, 24, 1085–1090. [Google Scholar] [CrossRef]
- Chen, L.; Na, R.; Gu, M.; Richardson, A.; Ran, Q. Lipid peroxidation up-regulates BACE1 expression in vivo: A possible early event of amyloidogenesis in Alzheimer’s disease. J. Neurochem. 2008, 107, 197–207. [Google Scholar] [CrossRef]
- Matsuoka, Y.; Picciano, M.; La Francois, J.; Duff, K. Fibrillar beta-amyloid evokes oxidative damage in a transgenic mouse model of Alzheimer’s disease. Neuroscience 2001, 104, 609–613. [Google Scholar] [CrossRef]
- Murakami, K.; Murata, N.; Noda, Y.; Tahara, S.; Kaneko, T.; Kinoshita, N.; Hatsuta, H.; Murayama, S.; Barnham, K.J.; Irie, K.; et al. SOD1 (copper/zinc superoxide dismutase) deficiency drives amyloid beta protein oligomerization and memory loss in mouse model of Alzheimer disease. J. Biol. Chem. 2011, 286, 44557–44568. [Google Scholar] [CrossRef]
- Deibel, M.A.; Ehmann, W.D.; Markesbery, W.R. Copper, iron, and zinc imbalances in severely degenerated brain regions in Alzheimer’s disease: Possible relation to oxidative stress. J. Neurol. Sci. 1996, 143, 137–142. [Google Scholar] [CrossRef]
- Crapper, D.R.; Krishnan, S.S.; Dalton, A.J. Brain aluminum distribution in Alzheimer’s disease and experimental neurofibrillary degeneration. Science 1973, 180, 511–513. [Google Scholar] [CrossRef]
- Mandrekar-Colucci, S.; Landreth, G.E. Microglia and inflammation in Alzheimer’s disease. CNS Neurol. Disord. Drug Targets 2010, 9, 156–167. [Google Scholar] [CrossRef]
- Wang, X.; Wang, W.; Li, L.; Perry, G.; Lee, H.G.; Zhu, X. Oxidative stress and mitochondrial dysfunction in Alzheimer’s disease. Biochim. Biophys. Acta 2014, 1842, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Rees, H.D.; Weintraub, S.T.; Levey, A.I.; Chin, L.S.; Li, L. Oxidative modifications and aggregation of Cu,Zn-superoxide dismutase associated with Alzheimer and Parkinson diseases. J. Biol. Chem. 2005, 280, 11648–11655. [Google Scholar] [CrossRef]
- Martinez, A.; Carmona, M.; Portero-Otin, M.; Naudi, A.; Pamplona, R.; Ferrer, I. Type-dependent oxidative damage in frontotemporal lobar degeneration: Cortical astrocytes are targets of oxidative damage. J. Neuropathol. Exp. Neurol. 2008, 67, 1122–1136. [Google Scholar] [CrossRef] [PubMed]
- De Leo, M.E.; Borrello, S.; Passantino, M.; Palazzotti, B.; Mordente, A.; Daniele, A.; Filippini, V.; Galeotti, T.; Masullo, C. Oxidative stress and overexpression of manganese superoxide dismutase in patients with Alzheimer’s disease. Neurosci. Lett. 1998, 250, 173–176. [Google Scholar] [CrossRef]
- Dias-Santagata, D.; Fulga, T.A.; Duttaroy, A.; Feany, M.B. Oxidative stress mediates tau-induced neurodegeneration in Drosophila. J. Clin. Investig. 2007, 117, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Jenner, P.; Olanow, C.W. Understanding cell death in Parkinson’s disease. Ann. Neurol. 1998, 44, S72–S84. [Google Scholar] [CrossRef] [PubMed]
- Rossi, L.; Squitti, R.; Pasqualetti, P.; Marchese, E.; Cassetta, E.; Forastiere, E.; Rotilio, G.; Rossini, P.M.; Finazzi-Agro, A. Red blood cell copper, zinc superoxide dismutase activity is higher in Alzheimer’s disease and is decreased by D-penicillamine. Neurosci. Lett 2002, 329, 137–140. [Google Scholar] [CrossRef]
- Perry, T.L.; Hansen, S.; Jones, K. Brain amino acids and glutathione in progressive supranuclear palsy. Neurology 1988, 38, 943–946. [Google Scholar] [CrossRef]
- Fitzmaurice, P.S.; Ang, L.; Guttman, M.; Rajput, A.H.; Furukawa, Y.; Kish, S.J. Nigral glutathione deficiency is not specific for idiopathic Parkinson’s disease. Mov. Disord. 2003, 18, 969–976. [Google Scholar] [CrossRef]
- Wojsiat, J.; Zoltowska, K.M.; Laskowska-Kaszub, K.; Wojda, U. Oxidant/Antioxidant Imbalance in Alzheimer’s Disease: Therapeutic and Diagnostic Prospects. Oxid. Med. Cell Longev. 2018, 2018, 6435861. [Google Scholar] [CrossRef]
- Youssef, P.; Chami, B.; Lim, J.; Middleton, T.; Sutherland, G.T.; Witting, P.K. Evidence supporting oxidative stress in a moderately affected area of the brain in Alzheimer’s disease. Sci. Rep. 2018, 8, 11553. [Google Scholar] [CrossRef] [PubMed]
- Nikam, S.; Nikam, P.; Ahaley, S.K.; Sontakke, A.V. Oxidative stress in Parkinson’s disease. Indian J. Clin. Biochem. 2009, 24, 98–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, L.L.; Quaglio, N.B.; de Souza, G.T.; Garcia, R.T.; Dati, L.M.; Moreira, W.L.; Loureiro, A.P.; de Souza-Talarico, J.N.; Smid, J.; Porto, C.S.; et al. Peripheral oxidative stress biomarkers in mild cognitive impairment and Alzheimer’s disease. J. Alzheimers Dis. 2011, 26, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Aoyama, K.; Matsubara, K.; Kobayashi, S. Aging and oxidative stress in progressive supranuclear palsy. Eur. J. Neurol. 2006, 13, 89–92. [Google Scholar] [CrossRef]
- Damier, P.; Hirsch, E.C.; Zhang, P.; Agid, Y.; Javoy-Agid, F. Glutathione peroxidase, glial cells and Parkinson’s disease. Neuroscience 1993, 52, 1–6. [Google Scholar] [CrossRef]
- Odetti, P.; Garibaldi, S.; Norese, R.; Angelini, G.; Marinelli, L.; Valentini, S.; Menini, S.; Traverso, N.; Zaccheo, D.; Siedlak, S.; et al. Lipoperoxidation is selectively involved in progressive supranuclear palsy. J. Neuropathol. Exp. Neurol. 2000, 59, 393–397. [Google Scholar] [CrossRef]
- Gerst, J.L.; Siedlak, S.L.; Nunomura, A.; Castellani, R.; Perry, G.; Smith, M.A. Role of oxidative stress in frontotemporal dementia. Dement. Geriatr. Cogn. Disord. 1999, 10 (Suppl. 1), 85–87. [Google Scholar] [CrossRef]
- Sudha, K.; Rao, A.V.; Rao, S.; Rao, A. Free radical toxicity and antioxidants in Parkinson’s disease. Neurol. India 2003, 51, 60–62. [Google Scholar] [PubMed]
- Albers, D.S.; Augood, S.J.; Martin, D.M.; Standaert, D.G.; Vonsattel, J.P.; Beal, M.F. Evidence for oxidative stress in the subthalamic nucleus in progressive supranuclear palsy. J. Neurochem. 1999, 73, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Zarkovic, K. 4-Hydroxynonenal and neurodegenerative diseases. Mol. Asp. Med. 2003, 24, 293–303. [Google Scholar] [CrossRef]
- Yoritaka, A.; Hattori, N.; Uchida, K.; Tanaka, M.; Stadtman, E.R.; Mizuno, Y. Immunohistochemical detection of 4-hydroxynonenal protein adducts in Parkinson disease. Proc. Natl. Acad. Sci. USA 1996, 93, 2696–2701. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Yamada, M. Vitamin A and Alzheimer’s disease. Geriatr. Gerontol. Int. 2012, 12, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Kook, S.Y.; Lee, K.M.; Kim, Y.; Cha, M.Y.; Kang, S.; Baik, S.H.; Lee, H.; Park, R.; Mook-Jung, I. High-dose of vitamin C supplementation reduces amyloid plaque burden and ameliorates pathological changes in the brain of 5XFAD mice. Cell Death Dis. 2014, 5, e1083. [Google Scholar] [CrossRef] [PubMed]
- Lloret, A.; Esteve, D.; Monllor, P.; Cervera-Ferri, A.; Lloret, A. The Effectiveness of Vitamin E Treatment in Alzheimer’s Disease. Int. J. Mol. Sci. 2019, 20, 879. [Google Scholar] [CrossRef]
- Castellani, R.; Smith, M.A.; Richey, P.L.; Kalaria, R.; Gambetti, P.; Perry, G. Evidence for oxidative stress in Pick disease and corticobasal degeneration. Brain Res 1995, 696, 268–271. [Google Scholar] [CrossRef]
- Ihara, Y.; Chuda, M.; Kuroda, S.; Hayabara, T. Hydroxyl radical and superoxide dismutase in blood of patients with Parkinson’s disease: Relationship to clinical data. J. Neurol. Sci. 1999, 170, 90–95. [Google Scholar] [CrossRef]
- Forsyth, C.B.; Shannon, K.M.; Kordower, J.H.; Voigt, R.M.; Shaikh, M.; Jaglin, J.A.; Estes, J.D.; Dodiya, H.B.; Keshavarzian, A. Increased intestinal permeability correlates with sigmoid mucosa alpha-synuclein staining and endotoxin exposure markers in early Parkinson’s disease. PLoS ONE 2011, 6, e28032. [Google Scholar] [CrossRef]
- Kikuchi, A.; Takeda, A.; Onodera, H.; Kimpara, T.; Hisanaga, K.; Sato, N.; Nunomura, A.; Castellani, R.J.; Perry, G.; Smith, M.A.; et al. Systemic increase of oxidative nucleic acid damage in Parkinson’s disease and multiple system atrophy. Neurobiol. Dis. 2002, 9, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Roostaei, T.; Felsky, D.; Nazeri, A.; de Jager, P.L.; Schneider, J.A.; Bennett, D.A.; Voineskos, A.N.; Alzheimer’s Disease Neuroimaging Initiative. Genetic influence of plasma homocysteine on Alzheimer’s disease. Neurobiol. Aging 2018, 62, 243.e7–243.e14. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Guan, P.P.; Wang, T.; Yu, X.; Guo, J.J.; Wang, Z.Y. Aggravation of Alzheimer’s disease due to the COX-2-mediated reciprocal regulation of IL-1beta and Abeta between glial and neuron cells. Aging Cell 2014, 13, 605–615. [Google Scholar] [CrossRef]
- Martin, E.; Rosenthal, R.E.; Fiskum, G. Pyruvate dehydrogenase complex: Metabolic link to ischemic brain injury and target of oxidative stress. J. Neurosci. Res. 2005, 79, 240–247. [Google Scholar] [CrossRef]
- Shi, Q.; Xu, H.; Kleinman, W.A.; Gibson, G.E. Novel functions of the alpha-ketoglutarate dehydrogenase complex may mediate diverse oxidant-induced changes in mitochondrial enzymes associated with Alzheimer’s disease. Biochim. Biophys. Acta 2008, 1782, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dai, M.; Yuan, Z. Methods for the detection of reactive oxygen species. Anal. Methods 2018, 10, 4625–4638. [Google Scholar] [CrossRef]
- Chen, X.; Wang, F.; Hyun, J.Y.; Wei, T.; Qiang, J.; Ren, X.; Shin, I.; Yoon, J. Recent progress in the development of fluorescent, luminescent and colorimetric probes for detection of reactive oxygen and nitrogen species. Chem. Soc. Rev. 2016, 45, 2976–3016. [Google Scholar] [CrossRef]
- Andina, D.; Leroux, J.C.; Luciani, P. Ratiometric fluorescent probes for the detection of reactive oxygen species. Chemistry 2017, 23, 13549–13573. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.; Fernandes, E.; Lima, J.L. Fluorescence probes used for detection of reactive oxygen species. J. Biochem. Biophys. Methods 2005, 65, 45–80. [Google Scholar] [CrossRef] [PubMed]
- Lo, L.-C.; Chu, C.-Y. Development of highly selective and sensitive probes for hydrogen peroxide. Chem. Commun. 2003, 7, 2728–2729. [Google Scholar] [CrossRef]
- Chang, M.C.; Pralle, A.; Isacoff, E.Y.; Chang, C.J. A selective, cell-permeable optical probe for hydrogen peroxide in living cells. J. Am. Chem. Soc. 2004, 126, 15392–15393. [Google Scholar] [CrossRef] [PubMed]
- Lippert, A.R.; van de Bittner, G.C.; Chang, C.J. Boronate oxidation as a bioorthogonal reaction approach for studying the chemistry of hydrogen peroxide in living systems. Acc. Chem. Res. 2011, 44, 793–804. [Google Scholar] [CrossRef]
- Kuivila, H.G.; Armour, A.G. Electrophilic displacement reactions. IX. Effects of substituents on rates of reactions between hydrogen peroxide and benzeneboronic acid1-3. J. Am. Chem. Soc. 1957, 79, 5659–5662. [Google Scholar] [CrossRef]
- Medvedeva, N.; Martin, V.V.; Weis, A.L.; Likhtenshten, G.I. Dual fluorophore-nitronyl probe for investigation of superoxide dynamics and antioxidant status of biological systems. J. Photochem. Photobiol. A Chem. 2004, 163, 45–51. [Google Scholar] [CrossRef]
- Afanasev, I. On mechanism of superoxide signaling under physiological and pathophysiological conditions. Med. Hypotheses 2005, 64, 127–129. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Liu, X.; Tang, B. A phosphinate-based red fluorescent probe for imaging the superoxide radical anion generated by RAW264. 7 macrophages. ChemBioChem 2007, 8, 453–458. [Google Scholar] [CrossRef]
- Hu, J.J.; Wong, N.-K.; Ye, S.; Chen, X.; Lu, M.-Y.; Zhao, A.Q.; Guo, Y.; Ma, A.C.-H.; Leung, A.Y.-H.; Shen, J. Fluorescent probe HKSOX-1 for imaging and detection of endogenous superoxide in live cells and in vivo. J. Am. Chem. Soc. 2015, 137, 6837–6843. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Yamamoto, K.; Nomura, Y.; Kohno, I.; Hafsi, L.; Ueda, N.; Yoshida, S.; Fukuda, M.; Fukuyasu, Y.; Yamauchi, Y. A design of fluorescent probes for superoxide based on a nonredox mechanism. J. Am. Chem. Soc. 2005, 127, 68–69. [Google Scholar] [CrossRef]
- Murale, D.P.; Kim, H.; Choi, W.S.; Churchill, D.G. Highly selective excited state intramolecular proton transfer (ESIPT)-based superoxide probing. Organ. Lett. 2013, 15, 3946–3949. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.C.; Murale, D.P.; Jang, S.Y.; Haque, M.M.; Seo, M.; Lee, S.; Woo, D.H.; Kwon, J.; Song, C.S.; Kim, Y.K. Discrimination of Avian Influenza Virus Subtypes using Host-Cell Infection Fingerprinting by a Sulfinate-based Fluorescence Superoxide Probe. Angew. Chem. Int. Ed. 2018, 57, 9716–9721. [Google Scholar] [CrossRef]
- You, Y.; Nam, W. Designing photoluminescent molecular probes for singlet oxygen, hydroxyl radical, and iron–oxygen species. Chem. Sci. 2014, 5, 4123–4135. [Google Scholar] [CrossRef]
- Ohashi, T.; Mizutani, A.; Murakami, A.; Kojo, S.; Ishii, T.; Taketani, S. Rapid oxidation of dichlorodihydrofluorescin with heme and hemoproteins: Formation of the fluorescein is independent of the generation of reactive oxygen species. FEBS Lett. 2002, 511, 21–27. [Google Scholar] [CrossRef]
- Kundu, K.; Knight, S.F.; Willett, N.; Lee, S.; Taylor, W.R.; Murthy, N. Hydrocyanines: A class of fluorescent sensors that can image reactive oxygen species in cell culture, tissue, and in vivo. Angew. Chem. Int. Ed. 2009, 48, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Lin, W.; Song, J. Ratiometric fluorescent detection of intracellular hydroxyl radicals based on a hybrid coumarin–cyanine platform. Chem. Commun. 2010, 46, 7930–7932. [Google Scholar] [CrossRef]
- Aliaga, C.; Fuentealba, P.; Rezende, M.C.; Cárdenas, C. Mechanism of fluorophore quenching in a pre-fluorescent nitroxide probe: A theoretical illustration. Chem. Phys. Lett. 2014, 593, 89–92. [Google Scholar] [CrossRef]
- Kang, P.; Foote, C.S. Formation of transient intermediates in low-temperature photosensitized oxidation of an 8-13C-guanosine derivative. J. Am. Chem. Soc. 2002, 124, 4865–4873. [Google Scholar] [CrossRef]
- Umezawa, N.; Tanaka, K.; Urano, Y.; Kikuchi, K.; Higuchi, T.; Nagano, T. Novel fluorescent probes for singlet oxygen. Angew. Chem. Int. Ed. 1999, 38, 2899–2901. [Google Scholar] [CrossRef]
- Wang, S.; Chen, L.; Jangili, P.; Sharma, A.; Li, W.; Hou, J.-T.; Qin, C.; Yoon, J.; Kim, J.S. Design and applications of fluorescent detectors for peroxynitrite. Coord. Chem. Rev. 2018, 374, 36–54. [Google Scholar] [CrossRef]
- Wang, B.; Yu, F.; Li, P.; Sun, X.; Han, K. A BODIPY fluorescence probe modulated by selenoxide spirocyclization reaction for peroxynitrite detection and imaging in living cells. Dyes Pigment. 2013, 96, 383–390. [Google Scholar] [CrossRef] [Green Version]
- Manjare, S.T.; Kim, Y.; Churchill, D.G. Selenium-and tellurium-containing fluorescent molecular probes for the detection of biologically important analytes. Acc. Chem. Res. 2014, 47, 2985–2998. [Google Scholar] [CrossRef]
- Lou, Z.; Li, P.; Han, K. Redox-responsive fluorescent probes with different design strategies. Acc. Chem. Res. 2015, 48, 1358–1368. [Google Scholar] [CrossRef] [PubMed]
- Iverson, N.; Hofferber, E.; Stapleton, J. Nitric Oxide Sensors for Biological Applications. Chemosensors 2018, 6, 8. [Google Scholar] [CrossRef]
- Vegesna, G.K.; Sripathi, S.R.; Zhang, J.; Zhu, S.; He, W.; Luo, F.-T.; Jahng, W.J.; Frost, M.; Liu, H. Highly water-soluble BODIPY-based fluorescent probe for sensitive and selective detection of nitric oxide in living cells. ACS Appl. Mater. Interfaces 2013, 5, 4107–4112. [Google Scholar] [CrossRef]
- Kim, J.; Kim, Y. A water-soluble sulfonate-BODIPY based fluorescent probe for selective detection of HOCl/OCl− in aqueous media. Analyst 2014, 139, 2986–2989. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, X.; Yuan, P.; Yang, J.; Xu, Y.; Grutzendler, J.; Shao, Y.; Moore, A.; Ran, C. Oxalate-curcumin-based probe for micro- and macroimaging of reactive oxygen species in Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2017, 114, 12384–12389. [Google Scholar] [CrossRef]
- Yang, J.; Yang, J.; Liang, S.H.; Xu, Y.; Moore, A.; Ran, C. Imaging hydrogen peroxide in Alzheimer’s disease via cascade signal amplification. Sci. Rep. 2016, 6, 35613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Yang, J.; Li, Y.; Xu, Y.; Ran, C. Near-infrared Fluorescence Ocular Imaging (NIRFOI) of Alzheimer’s Disease. Mol. Imaging Biol. 2019, 21, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Mu, X.Y.; Yang, J.; Liang, Y.Y.; Zhang, X.D.; Ming, D. Brain imaging with near-infrared fluorophores. Coord. Chem. Rev. 2019, 380, 550–571. [Google Scholar] [CrossRef]
- Needham, L.M.; Weber, J.; Fyfe, J.W.B.; Kabia, O.M.; Do, D.T.; Klimont, E.; Zhang, Y.; Rodrigues, M.; Dobson, C.M.; Ghandi, S.; et al. Bifunctional fluorescent probes for detection of amyloid aggregates and reactive oxygen species. R. Soc. Open Sci. 2018, 5, 171399. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Liu, X.; Tang, B.; Yang, G.; Yang, Y.; An, L. Design of a phosphinate-based fluorescent probe for superoxide detection in mouse peritoneal macrophages. Chemistry 2007, 13, 1411–1416. [Google Scholar] [CrossRef]
- Yang, X.F.; Guo, X.Q. Investigation of the anthracene-nitroxide hybrid molecule as a probe for hydroxyl radicals. Analyst 2001, 126, 1800–1804. [Google Scholar] [CrossRef]
- Leikert, J.F.; Rathel, T.R.; Muller, C.; Vollmar, A.M.; Dirsch, V.M. Reliable in vitro measurement of nitric oxide released from endothelial cells using low concentrations of the fluorescent probe 4,5-diaminofluorescein. FEBS Lett. 2001, 506, 131–134. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Kim, H.S.; Cho, E.K.; Kwon, B.Y.; Phark, S.; Hwang, K.W.; Sul, D. Curcumin protected PC12 cells against beta-amyloid-induced toxicity through the inhibition of oxidative damage and tau hyperphosphorylation. Food Chem. Toxicol. 2008, 46, 2881–2887. [Google Scholar] [CrossRef]
- Stack, C.; Jainuddin, S.; Elipenahli, C.; Gerges, M.; Starkova, N.; Starkov, A.A.; Jove, M.; Portero-Otin, M.; Launay, N.; Pujol, A.; et al. Methylene blue upregulates Nrf2/ARE genes and prevents tau-related neurotoxicity. Hum. Mol. Genet. 2014, 23, 3716–3732. [Google Scholar] [CrossRef] [Green Version]
- Elipenahli, C.; Stack, C.; Jainuddin, S.; Gerges, M.; Yang, L.; Starkov, A.; Beal, M.F.; Dumont, M. Behavioral improvement after chronic administration of coenzyme Q10 in P301S transgenic mice. J. Alzheimers Dis. 2012, 28, 173–182. [Google Scholar] [CrossRef]
- Morroni, F.; Tarozzi, A.; Sita, G.; Bolondi, C.; Zolezzi Moraga, J.M.; Cantelli-Forti, G.; Hrelia, P. Neuroprotective effect of sulforaphane in 6-hydroxydopamine-lesioned mouse model of Parkinson’s disease. Neurotoxicology 2013, 36, 63–71. [Google Scholar] [CrossRef]
- Takenouchi, T.; Sekiyama, K.; Sekigawa, A.; Fujita, M.; Waragai, M.; Sugama, S.; Iwamaru, Y.; Kitani, H.; Hashimoto, M. P2X7 receptor signaling pathway as a therapeutic target for neurodegenerative diseases. Arch. Immunol. Ther. Exp. (Warsz.) 2010, 58, 91–96. [Google Scholar] [CrossRef]
- Clausen, A.; Xu, X.; Bi, X.; Baudry, M. Effects of the superoxide dismutase/catalase mimetic EUK-207 in a mouse model of Alzheimer’s disease: Protection against and interruption of progression of amyloid and tau pathology and cognitive decline. J. Alzheimers Dis. 2012, 30, 183–208. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, M.; Li, C.; Jiang, X.; Su, Y.; Zhang, Y. Benefits of Vitamins in the Treatment of Parkinson’s Disease. Oxid. Med. Cell Longev. 2019, 2019, 9426867. [Google Scholar] [CrossRef]
- Cente, M.; Filipcik, P.; Mandakova, S.; Zilka, N.; Krajciova, G.; Novak, M. Expression of a truncated human tau protein induces aqueous-phase free radicals in a rat model of tauopathy: Implications for targeted antioxidative therapy. J. Alzheimers Dis. 2009, 17, 913–920. [Google Scholar] [CrossRef]
- Nakashima, H.; Ishihara, T.; Yokota, O.; Terada, S.; Trojanowski, J.Q.; Lee, V.M.; Kuroda, S. Effects of alpha-tocopherol on an animal model of tauopathies. Free Radic. Biol. Med. 2004, 37, 176–186. [Google Scholar] [CrossRef]
- Taghizadeh, M.; Tamtaji, O.R.; Dadgostar, E.; Daneshvar Kakhaki, R.; Bahmani, F.; Abolhassani, J.; Aarabi, M.H.; Kouchaki, E.; Memarzadeh, M.R.; Asemi, Z. The effects of omega-3 fatty acids and vitamin E co-supplementation on clinical and metabolic status in patients with Parkinson’s disease: A randomized, double-blind, placebo-controlled trial. Neurochem. Int. 2017, 108, 183–189. [Google Scholar] [CrossRef]
- Grossi, C.; Francese, S.; Casini, A.; Rosi, M.C.; Luccarini, I.; Fiorentini, A.; Gabbiani, C.; Messori, L.; Moneti, G.; Casamenti, F. Clioquinol decreases amyloid-beta burden and reduces working memory impairment in a transgenic mouse model of Alzheimer’s disease. J. Alzheimers Dis. 2009, 17, 423–440. [Google Scholar] [CrossRef]
- Faux, N.G.; Ritchie, C.W.; Gunn, A.; Rembach, A.; Tsatsanis, A.; Bedo, J.; Harrison, J.; Lannfelt, L.; Blennow, K.; et al. PBT2 rapidly improves cognition in Alzheimer’s Disease: Additional phase II analyses. J. Alzheimers Dis. 2010, 20, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Crouch, P.J.; Hung, L.W.; Adlard, P.A.; Cortes, M.; Lal, V.; Filiz, G.; Perez, K.A.; Nurjono, M.; Caragounis, A.; Du, T.; et al. Increasing Cu bioavailability inhibits Abeta oligomers and tau phosphorylation. Proc. Natl. Acad. Sci. USA 2009, 106, 381–386. [Google Scholar] [CrossRef]
- Crapper McLachlan, D.R.; Dalton, A.J.; Kruck, T.P.; Bell, M.Y.; Smith, W.L.; Kalow, W.; Andrews, D.F. Intramuscular desferrioxamine in patients with Alzheimer’s disease. Lancet 1991, 337, 1304–1308. [Google Scholar] [CrossRef]
- Lejri, I.; Grimm, A.; Miesch, M.; Geoffroy, P.; Eckert, A.; Mensah-Nyagan, A.G. Allopregnanolone and its analog BR 297 rescue neuronal cells from oxidative stress-induced death through bioenergetic improvement. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Perez-Gonzalez, R.; Pascual, C.; Antequera, D.; Bolos, M.; Redondo, M.; Perez, D.I.; Perez-Grijalba, V.; Krzyzanowska, A.; Sarasa, M.; Gil, C.; et al. Phosphodiesterase 7 inhibitor reduced cognitive impairment and pathological hallmarks in a mouse model of Alzheimer’s disease. Neurobiol. Aging 2013, 34, 2133–2145. [Google Scholar] [CrossRef] [PubMed]
- Turner, R.S.; Thomas, R.G.; Craft, S.; van Dyck, C.H.; Mintzer, J.; Reynolds, B.A.; Brewer, J.B.; Rissman, R.A.; Raman, R.; Aisen, P.S. A randomized, double-blind, placebo-controlled trial of resveratrol for Alzheimer disease. Neurology 2015, 85, 1383–1391. [Google Scholar] [CrossRef] [Green Version]
- Dysken, M.W.; Sano, M.; Asthana, S.; Vertrees, J.E.; Pallaki, M.; Llorente, M.; Love, S.; Schellenberg, G.D.; McCarten, J.R.; Malphurs, J.; et al. Effect of vitamin E and memantine on functional decline in Alzheimer disease: The TEAM-AD VA cooperative randomized trial. JAMA 2014, 311, 33–44. [Google Scholar] [CrossRef]
- Ibrahim, N.F.; Yanagisawa, D.; Durani, L.W.; Hamezah, H.S.; Damanhuri, H.A.; Wan Ngah, W.Z.; Tsuji, M.; Kiuchi, Y.; Ono, K.; Tooyama, I. Tocotrienol-Rich Fraction Modulates Amyloid Pathology and Improves Cognitive Function in AbetaPP/PS1 Mice. J. Alzheimers Dis. 2017, 55, 597–612. [Google Scholar] [CrossRef]
Figure 1.
Oxidative stress-mediated tauopathy.

Figure 2.
Electronic configuration of reactive oxygen species (ROS).

Figure 3.
Standard reduction potentials of ROS formation via energy or electron transfer reactions of molecular oxygen.
Figure 3.
Standard reduction potentials of ROS formation via energy or electron transfer reactions of molecular oxygen.

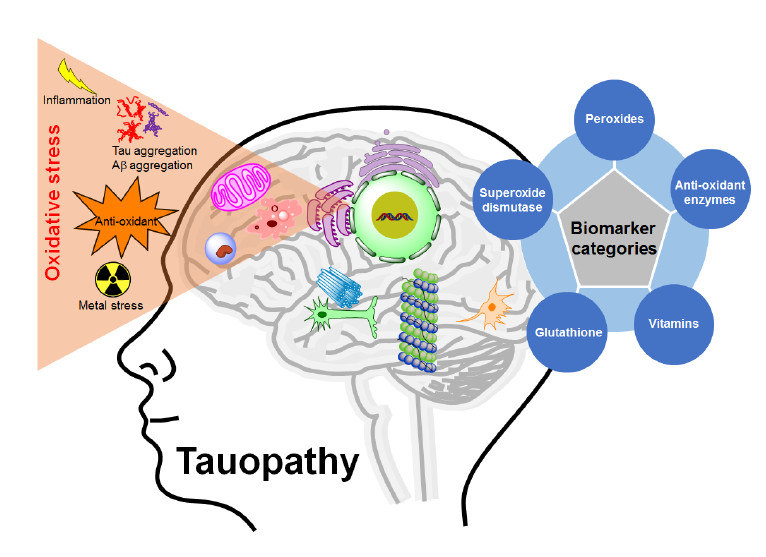
Figure 4.
Physiological condition versus tauopathy condition by oxidative stress generation.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Potential oxidative markers in tauopathies (up-regulation is labeled as up-arrow, and down-regulation is labeled as down-arrow).
Table 1.
Potential oxidative markers in tauopathies (up-regulation is labeled as up-arrow, and down-regulation is labeled as down-arrow).
| OS Markers | AD | PSP | PiD | CBD | FTD | FTDP-17 | FTLD | PD |
|---|---|---|---|---|---|---|---|---|
| SOD1 | ↓ [27] ↑ [63] | ↑ [11] | ↑ [64] | ↑ [63] | ||||
| SOD2 | ↓ [27] ↑ [65] | ↑ [11] | ↓ [66] | ↑ [64] | ↑ [67] | |||
| Cu/Zn-SOD | ↓ [27] ↑ [68] | ↑ [11] | ↑ [63] | |||||
| GSH | ↓ [27] | ↑ [69] ↓ [70] | ↓ [64] | ↓ [69] | ||||
| GST | ↓ [71] | ↓ [11] | ||||||
| Catalase | ↓ [72] | ↓ [73] | ||||||
| GPx | ↓ [72] ↑ [74] | ↑ [75] | ↑ [11] ↓ [11] | ↑ [76] ↓ [67] | ||||
| Lipid peroxidase | ↑ [27] | ↑ [77] | ↑ [78] | ↑ [79] | ||||
| MDA | ↑ [27] | ↑ [80] | ↑ [64] | ↑ [81] | ||||
| 4-HNE | ↑ [27] | ↑ [75] | ↑ [81] | ↑ [64] | ↑ [82] | |||
| Vitamin A | ↓ [83] | |||||||
| Vitamin C | ↓ [84] | ↓ [73] | ||||||
| Vitamin E | ↓ [85] | ↓ [73] | ||||||
| HO-1 | ↑ [86] | ↑ [86] | ↑ [86] | ↑ [86] | ↑ [78] | ↑ [87] | ||
| Protein carbonyl | ↑ [27] | ↑ [78] | ↑ [63] | |||||
| 3-nitrotyrosine | ↑ [27] | ↑ [88] | ||||||
| 8-OHG | ↑ [27] | ↑ [89] | ||||||
| 8-OHdG | ↑ [27] | ↑ [89] | ||||||
| F2-IsoPs | ↑ [27] | |||||||
| Hcy | ↑ [90] | |||||||
| COX | ↑ [91] | |||||||
| PDHC | ↓ [92] | |||||||
| KGDHC | ↓ [93] |
Table 2.
ROS and their mode of detection using fluorescent probes.
| ROS | Fluorescent Probe | Reaction | Examples |
|---|---|---|---|
| Hydrogen peroxide | H2O2-induced oxidation of arylboronate ester to phenols. |  |  (Ref: 1 [98], 2 [99]) |
| Superoxide | Nucleophilic substitution reaction of superoxide with the probes where the leaving group is phosphinate group [104], triflate group [105], 2,4-dinitrobenzenesulfonyl [106], nitro-ethers [107] and the related groups. |  |  (Ref: 3 [128],4 [108]) |
| Hydroxyl radical | Hydroxyl radical-mediated oxidation of leuco forms of fluorescent dyes and nitroxide moiety. |  |  (Ref: 5 [112], 6 [129]) |
| Singlet oxygen | The fluorescent probes for singlet oxygen have been developed based on the [2 + 4] cycloaddition where singlet oxygen acts as a strong dienophile [109,114,115]. |  |  (Ref: 7 [115]) |
| Peroxynitrite | Peroxynitrite-mediated oxidation of chalcogenides (S, Se, and Te), boronic acids or boronates, hydrazides, cleavage of C–C double bonds and oxidative N-dearylation. |  |  (Ref: 8 [122]) |
| Nitric oxide | NO-mediated reaction of O-diaminophenyl group with NO to generate the triazole. |  |  (Ref: 9 [130]) |
| Hypochlorite | Hypochlorite mediated spiro-ring opening of the xanthene probes and oxidation of catechol to benzoquinone [122]. |  |  (Ref: 10 [117]) |
Table 3.
Therapeutic approaches of tauopathies related to ROS.
| Therapeutic Approaches | Chemical Agents | Treating Route and Doses | Experimental Model | Reference |
|---|---|---|---|---|
| Antioxidant pathway | Curcumin | 10 μg/mL for 1 h | Aβ treated PC12 cell | [131] |
| Methylene blue | 4 mg/kg in diet | TauP301S mouse | [132] | |
| CoQ10 | 0.5% in diet | TauP301S mouse | [133] | |
| Paraquant | 30 mM in diet for 48 h | TauR406W drosophila | [66] | |
| Sulforaphane | 5 mg/kg twice a week by intraperitoneally | C57Bl/66-OHDA mouse | [134] | |
| BR 297 | 500 nM for 24 h | APP treated SH-SY5Y cell | [145] | |
| S14 | 5 mg/Kg/daily for 4 weeks by intraperitoneally | Tg2576APP/PS1 AD mouse | [146] | |
| Resveratrol | 500–1000 mg/daily for 26 months by orally | AD patients | [147] | |
| Catalase | EUK-207 | 3.41 mM/day for 28 days by micro-osmotic pump | C57BL/6/129S-3xTg-AD mouse | [136] |
| Vitamin | Ascorbic acid | 250–500 μM for 24 h | Neurons form Tau151–391 rat | [138] |
| α-tocopherol | 0.5–1.5 mM in diet for 10 days | tauR406W drosophila | [66] | |
| 49 IU/Kg in diet | B6D2/F1-tau44 mouse | [139] | ||
| 2000 IU/day for 6 months | AD patient | [148] | ||
| Tocotrienol | 5 mg/Kg/day for 15 months by orally | APPswe/PS1dE9 mouse | [149] | |
| Metal chelator | Clioquinol | 30 mg/kg/day for 5 weeks by orally | TgCRND8-AD mouse | [141] |
| PBT2 | 250 mg/day for 12 weeks by orally | AD patient | [142] | |
| CuIIGTSM | 10 mg/kg/daily by orally | APP/PS1 AD mouse | [143] | |
| Desferrioxamine | 125 mg twice daily/5 days per week for 24 months by intramuscularly | AD patient | [144] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Haque, M.M.; Murale, D.P.; Kim, Y.K.; Lee, J.-S. Crosstalk between Oxidative Stress and Tauopathy. Int. J. Mol. Sci. 2019, 20, 1959. https://doi.org/10.3390/ijms20081959
AMA Style
Haque MM, Murale DP, Kim YK, Lee J-S. Crosstalk between Oxidative Stress and Tauopathy. International Journal of Molecular Sciences. 2019; 20(8):1959. https://doi.org/10.3390/ijms20081959
Chicago/Turabian StyleHaque, Md. Mamunul, Dhiraj P. Murale, Yun Kyung Kim, and Jun-Seok Lee. 2019. "Crosstalk between Oxidative Stress and Tauopathy" International Journal of Molecular Sciences 20, no. 8: 1959. https://doi.org/10.3390/ijms20081959
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.