Molecular Mechanisms Driving Progression of Liver Cirrhosis towards Hepatocellular Carcinoma in Chronic Hepatitis B and C Infections: A Review



Abstract
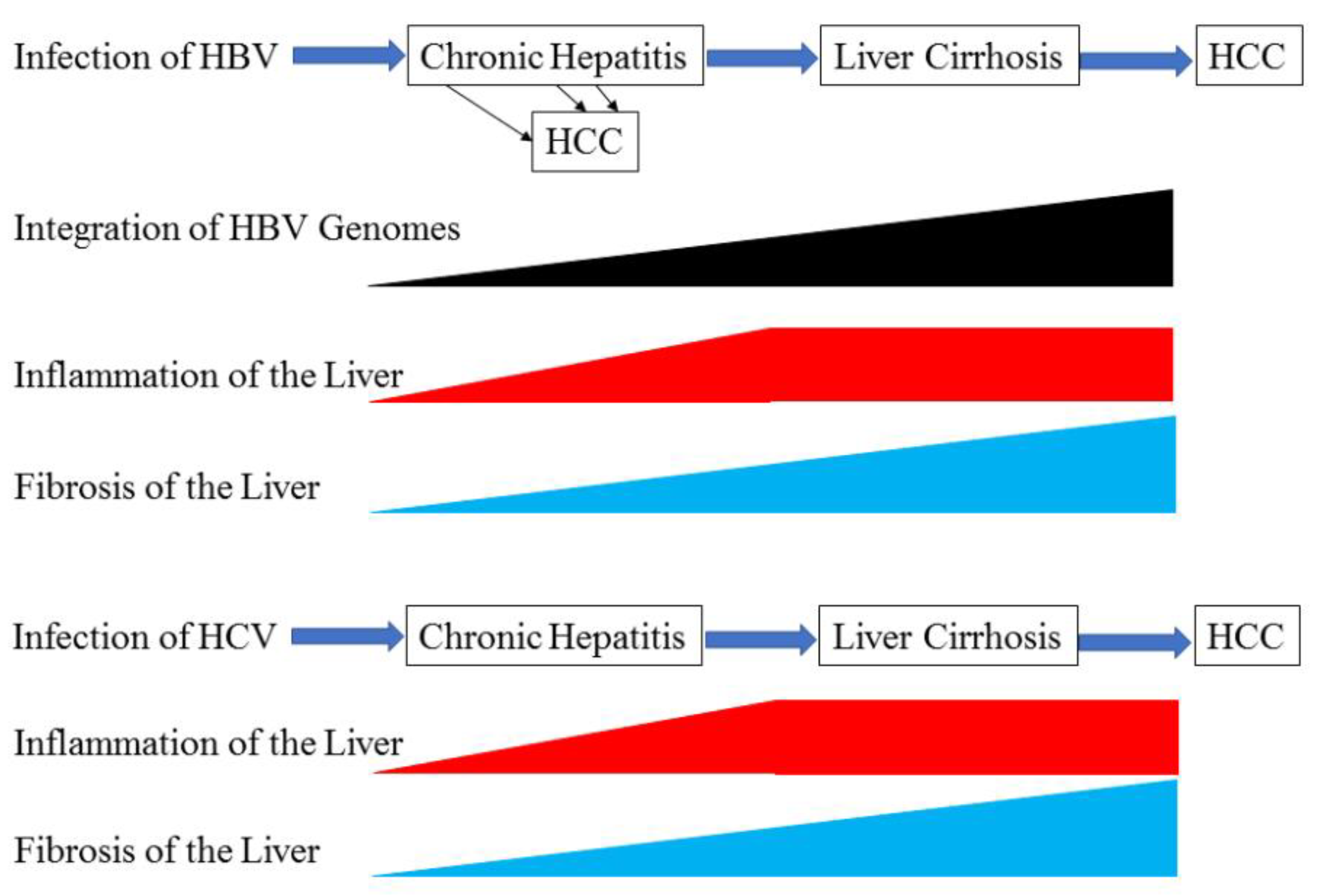
:1. Introduction
2. Liver Cirrhosis and Its Progression to HCC with HBV Infection
2.1. Development of Liver Cirrhosis in Patients with Chronic Hepatitis B Infection
2.2. Development of HCC in Patients with HBV-related Liver Cirrhosis
3. Liver Cirrhosis and Its Progression to HCC with HCV Infection
4. Molecular Mechanisms of Liver Cirrhosis and Its Progression to HCC
4.1. Driver-Gene Candidates in HCC
4.2. The p53-RB Pathway
4.3. The β-catenin Pathway (WNT Pathway)
4.4. Chromatin and Transcription Modulators
4.5. Other Pathways
5. Molecular Mechanisms of HBV-Associated HCC
5.1. HBV Genome Integration Promotes HCC
5.2. Inflammation Promotes HBV-Associated HCC
5.3. Epigenetic Mechanisms Involved in HBV-Associated HCC
5.4. Roles of HBV-encoded Proteins
6. Molecular Mechanisms of HCV-Associated HCC
6.1. Inflammation Promotes HCV-Associated HCC
6.2. Epigenetic Mechanisms Involved in HCV-Associated HCC
6.3. Roles of HCV-coding Proteins
7. HCC-specific Molecular Mechanisms from a Therapeutic Point of View
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Takano, S.; Yokosuka, O.; Imazeki, F.; Tagawa, M.; Omata, M. Incidence of hepatocellular carcinoma in chronic hepatitis B and C: A prospective study of 251 patients. Hepatology 1995, 21, 650–655. [Google Scholar] [CrossRef] [PubMed]
- Takayama, T.; Makuuchi, M. Segmental liver resections, present and future-caudate lobe resection for liver tumours. Hepatogastroenterology 1998, 45, 20–23. [Google Scholar]
- Llovet, J.M.; Ricci, S.; Mazzaferro, V.; Hilgard, P.; Gane, E.; Blanc, J.F.; de Oliveira, A.C.; Santoro, A.; Raoul, J.L.; Forner, A.; et al. Sorafenib in advanced hepatocellular carcinoma. N. Engl. J. Med. 2008, 359, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.L.; Kang, Y.K.; Chen, Z.; Tsao, C.J.; Qin, S.; Kim, J.S.; Luo, R.; Feng, J.; Ye, S.; Yang, T.S.; et al. Efficacy and safety of sorafenib in patients in the Asia-Pacific region with advanced hepatocellular carcinoma: A phase III randomised, double-blind, placebo-controlled trial. Lancet Oncol. 2009, 10, 25–34. [Google Scholar] [CrossRef]
- Bruix, J.; Qin, S.; Merle, P.; Granito, A.; Huang, Y.H.; Bodoky, G.; Pracht, M.; Yokosuka, O.; Rosmorduc, O.; Breder, V.; et al. Regorafenib for patients with hepatocellular carcinoma who progressed on sorafenib treatment (RESORCE): A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet 2017, 389, 56–66. [Google Scholar] [CrossRef]
- Kudo, M.; Finn, R.S.; Qin, S.; Han, K.H.; Ikeda, K.; Piscaglia, F.; Baron, A.; Park, J.W.; Han, G.; Jassem, J.; et al. Lenvatinib versus sorafenib in first-line treatment of patients with unresectable hepatocellular carcinoma: A randomised phase 3 non-inferiority trial. Lancet 2018, 391, 1163–1173. [Google Scholar] [CrossRef]
- Schachter, J.; Ribas, A.; Long, G.V.; Arance, A.; Grob, J.J.; Mortier, L.; Daud, A.; Carlino, M.S.; McNeil, C.; Lotem, M.; et al. Pembrolizumab versus ipilimumab for advanced melanoma: Final overall survival results of a multicentre, randomised, open-label phase 3 study (KEYNOTE-006). Lancet 2017, 390, 1853–1862. [Google Scholar] [CrossRef]
- Bruix, J.; Reig, M.; Sherman, M. Evidence-Based Diagnosis, Staging, and Treatment of Patients With Hepatocellular Carcinoma. Gastroenterology 2016, 150, 835–853. [Google Scholar] [CrossRef]
- Obi, S.; Yoshida, H.; Toune, R.; Unuma, T.; Kanda, M.; Sato, S.; Tateishi, R.; Teratani, T.; Shiina, S.; Omata, M. Combination therapy of intraarterial 5-fluorouracil and systemic interferon-alpha for advanced hepatocellular carcinoma with portal venous invasion. Cancer 2006, 106, 1990–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanda, T.; Takahashi, K.; Nakamura, M.; Nakamoto, S.; Wu, S.; Haga, Y.; Sasaki, R.; Jiang, X.; Yokosuka, O. Androgen Receptor Could Be a Potential Therapeutic Target in Patients with Advanced Hepatocellular Carcinoma. Cancers 2017, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- Fattovich, G.; Stroffolini, T.; Zagni, I.; Donato, F. Hepatocellular carcinoma in cirrhosis: Incidence and risk factors. Gastroenterology 2004, 127 (Suppl. 1), S35–S50. [Google Scholar] [CrossRef]
- Bataller, R.; Brenner, D.A. Liver fibrosis. J. Clin. Investig. 2005, 115, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.; Bedossa, P.; Castera, L. Diagnosis of liver fibrosis: Present and future. Semin. Liver Dis. 2015, 35, 166–183. [Google Scholar] [CrossRef]
- Xu, L.; Hui, A.Y.; Albanis, E.; Arthur, M.J.; O’Byrne, S.M.; Blaner, W.S.; Mukherjee, P.; Friedman, S.L.; Eng, F.J. Human hepatic stellate cell lines, LX-1 and LX-2: New tools for analysis of hepatic fibrosis. Gut 2005, 54, 142–151. [Google Scholar] [CrossRef]
- Okuda, K.; Nakashima, T.; Kojiro, M.; Kondo, Y.; Wada, K. Hepatocellular carcinoma without cirrhosis in Japanese patients. Gastroenterology 1989, 97, 140–146. [Google Scholar] [CrossRef]
- Clouston, A.D.; Powell, E.E.; Walsh, M.J.; Richardson, M.M.; Demetris, A.J.; Jonsson, J.R. Fibrosis correlates with a ductular reaction in hepatitis C: Roles of impaired replication, progenitor cells and steatosis. Hepatology 2005, 41, 809–818. [Google Scholar] [CrossRef] [Green Version]
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abraham, J.; Adair, T.; Aggarwal, R.; Ahn, S.Y.; et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- Liaw, Y.F.; Tai, D.I.; Chu, C.M.; Chen, T.J. The development of cirrhosis in patients with chronic type B hepatitis: A prospective study. Hepatology 1988, 8, 493–496. [Google Scholar] [CrossRef]
- Fattovich, G.; Brollo, L.; Giustina, G.; Noventa, F.; Pontisso, P.; Alberti, A.; Realdi, G.; Ruol, A. Natural history and prognostic factors for chronic hepatitis type B. Gut 1991, 32, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Sumi, H.; Yokosuka, O.; Seki, N.; Arai, M.; Imazeki, F.; Kurihara, T.; Kanda, T.; Fukai, K.; Kato, M.; Saisho, H. Influence of hepatitis B virus genotypes on the progression of chronic type B liver disease. Hepatology 2003, 37, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Liaw, Y.F.; Chen, Y.C.; Sheen, I.S.; Chien, R.N.; Yeh, C.T.; Chu, C.M. Impact of acute hepatitis C virus superinfection in patients with chronic hepatitis B virus infection. Gastroenterology 2004, 126, 1024–1029. [Google Scholar] [CrossRef]
- Lacombe, K.; Massari, V.; Girard, P.M.; Serfaty, L.; Gozlan, J.; Pialoux, G.; Mialhes, P.; Molina, J.M.; Lascoux-Combe, C.; Wendum, D.; et al. Major role of hepatitis B genotypes in liver fibrosis during coinfection with HIV. AIDS 2006, 20, 419–427. [Google Scholar] [CrossRef]
- Stroffolini, T.; Esvan, R.; Biliotti, E.; Sagnelli, E.; Gaeta, G.B.; Almasio, P.L. Gender differences in chronic HBsAg carriers in Italy: Evidence for the independent role of male sex in severity of liver disease. J. Med. Virol. 2015, 87, 1899–1903. [Google Scholar] [CrossRef]
- Al-Qahtani, A.; Khalak, H.G.; Alkuraya, F.S.; Al-hamoudi, W.; Alswat, K.; Al Balwi, M.A.; Al Abdulkareem, I.; Sanai, F.M.; Abdo, A.A. Genome-wide association study of chronic hepatitis B virus infection reveals a novel candidate risk allele on 11q22.3. J. Med. Genet. 2013, 50, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.W.; Fann, C.S.; Su, W.H.; Wang, Y.C.; Weng, C.C.; Yu, C.J.; Hsu, C.L.; Hsieh, A.R.; Chien, R.N.; Chu, C.M.; et al. A genome-wide association study on chronic HBV infection and its clinical progression in male Han-Taiwanese. PLoS ONE 2014, 9, e99724. [Google Scholar] [CrossRef]
- Chu, C.M.; Liaw, Y.F. Hepatitis B virus-related cirrhosis: Natural history and treatment. Semin. Liver Dis. 2006, 26, 142–152. [Google Scholar] [CrossRef]
- Tawada, A.; Kanda, T.; Imazeki, F.; Yokosuka, O. Prevention of hepatitis B virus-associated liver diseases by antiviral therapy. Hepatol. Int. 2016, 10, 574–593. [Google Scholar] [CrossRef]
- World Health Organization. Global Hepatitis Report. 2017. Available online: www.who.int/hepatitis/publications/global-hepatitis-report2017/en/ (accessed on 27 December 2018).
- Yang, Y.H.; Chen, W.C.; Tsan, Y.T.; Chen, M.J.; Shih, W.T.; Tsai, Y.H.; Chen, P.C. Statin use and the risk of cirrhosis development in patients with hepatitis C virus infection. J. Hepatol. 2015, 63, 1111–1117. [Google Scholar] [CrossRef]
- Forns, X.; Ampurdanès, S.; Sanchez-Tapias, J.M.; Guilera, M.; Sans, M.; Sánchez-Fueyo, A.; Quintó, L.; Joya, P.; Bruguera, M.; Rodés, J. Long-term follow-up of chronic hepatitis C in patients diagnosed at a tertiary-care center. J. Hepatol. 2001, 35, 265–271. [Google Scholar] [CrossRef]
- Westin, J.; Nordlinder, H.; Lagging, M.; Norkrans, G.; Wejstål, R. Steatosis accelerates fibrosis development over time in hepatitis C virus genotype 3 infected patients. J. Hepatol. 2002, 37, 837–842. [Google Scholar] [CrossRef]
- Rubbia-Brandt, L.; Fabris, P.; Paganin, S.; Leandro, G.; Male, P.J.; Giostra, E.; Carlotto, A.; Bozzola, L.; Smedile, A.; Negro, F. Steatosis affects chronic hepatitis C progression in a genotype specific way. Gut 2004, 53, 406–412. [Google Scholar] [CrossRef] [Green Version]
- Locasciulli, A.; Testa, M.; Pontisso, P.; Benvegnù, L.; Fraschini, D.; Corbetta, A.; Noventa, F.; Masera, G.; Alberti, A. Prevalence and natural history of hepatitis C infection in patients cured of childhood leukemia. Blood 1997, 90, 4628–4633. [Google Scholar] [PubMed]
- Kenny-Walsh, E. Clinical outcomes after hpatitis C infection from contaminated anti-D immune globulin. Irish Hepatology Research Group. N. Engl. J. Med. 1999, 340, 1228–1233. [Google Scholar] [CrossRef]
- Poynard, T.; Bedossa, P.; Opolon, P. Natural history of liver fibrosis progression in patients with chronic hepatitis C. The OBSVIRC, METAVIR, CLINIVIR, and DOSVIRC groups. Lancet 1997, 349, 825–832. [Google Scholar] [CrossRef]
- Poynard, T.; Ratziu, V.; Charlotte, F.; Goodman, Z.; McHutchison, J.; Albrecht, J. Rates and risk factors of liver fibrosis progression in patients with chronic hepatitis c. J. Hepatol. 2001, 34, 730–739. [Google Scholar] [CrossRef]
- Shiffman, M.L. Natural history and risk factors for progression of hepatitis C virus disease and development of hepatocellular cancer before liver transplantation. Liver Transpl. 2003, 9, S14–S20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuster, D.; Planas, R.; Muga, R.; Ballesteros, A.L.; Santos, J.; Tor, J.; Sirera, G.; Guardiola, H.; Salas, A.; Cabré, E.; et al. Advanced liver fibrosis in HIV/HCV-coinfected patients on antiretroviral therapy. AIDS Res. Hum. Retrovir. 2004, 20, 1293–1297. [Google Scholar] [CrossRef] [PubMed]
- Ribes, J.; Clèries, R.; Rubió, A.; Hernández, J.M.; Mazzara, R.; Madoz, P.; Casanovas, T.; Casanova, A.; Gallen, M.; Rodríguez, C.; et al. Cofactors associated with liver disease mortality in an HBsAg-positive Mediterranean cohort: 20 years of follow-up. Int. J. Cancer 2006, 119, 687–694. [Google Scholar] [CrossRef] [Green Version]
- Wilson, L.E.; Torbenson, M.; Astemborski, J.; Faruki, H.; Spoler, C.; Rai, R.; Mehta, S.; Kirk, G.D.; Nelson, K.; Afdhal, N.; et al. Progression of liver fibrosis among injection drug users with chronic hepatitis C. Hepatology 2006, 43, 788–795. [Google Scholar] [CrossRef] [Green Version]
- Petta, S.; Cammà, C.; Di Marco, V.; Alessi, N.; Cabibi, D.; Caldarella, R.; Licata, A.; Massenti, F.; Tarantino, G.; Marchesini, G.; et al. Insulin resistance and diabetes increase fibrosis in the liver of patients with genotype 1 HCV infection. Am. J. Gastroenterol. 2008, 103, 1136–1144. [Google Scholar] [CrossRef]
- Dyal, H.K.; Aguilar, M.; Bhuket, T.; Liu, B.; Holt, E.W.; Torres, S.; Cheung, R.; Wong, R.J. Concurrent Obesity, Diabetes, and Steatosis Increase Risk of Advanced Fibrosis Among HCV Patients: A Systematic Review. Dig. Dis. Sci. 2015, 60, 2813–2824. [Google Scholar] [CrossRef] [PubMed]
- Shiratori, Y.; Imazeki, F.; Moriyama, M.; Yano, M.; Arakawa, Y.; Yokosuka, O.; Kuroki, T.; Nishiguchi, S.; Sata, M.; Yamada, G.; et al. Histologic improvement of fibrosis in patients with hepatitis C who have sustained response to interferon therapy. Ann. Intern. Med. 2000, 132, 517–524. [Google Scholar] [CrossRef]
- Caporaso, N.; Romano, M.; Marmo, R.; de Sio, I.; Morisco, F.; Minerva, A.; Coltorti, M. Hepatitis C virus infection is an additive risk factor for development of hepatocellular carcinoma in patients with cirrhosis. J. Hepatol. 1991, 12, 367–371. [Google Scholar] [CrossRef]
- Benvegnù, L.; Fattovich, G.; Noventa, F.; Tremolada, F.; Chemello, L.; Cecchetto, A.; Alberti, A. Concurrent hepatitis B and C virus infection and risk of hepatocellular carcinoma in cirrhosis. A prospective study. Cancer 1994, 74, 2442–2448. [Google Scholar] [CrossRef]
- Ishikawa, T.; Ichida, T.; Yamagiwa, S.; Sugahara, S.; Uehara, K.; Okoshi, S.; Asakura, H. High viral loads, serum alanine aminotransferase and gender are predictive factors for the development of hepatocellular carcinoma from viral compensated liver cirrhosis. J. Gastroenterol. Hepatol. 2001, 16, 1274–1281. [Google Scholar] [CrossRef]
- Ohata, K.; Hamasaki, K.; Toriyama, K.; Matsumoto, K.; Saeki, A.; Yanagi, K.; Abiru, S.; Nakagawa, Y.; Shigeno, M.; Miyazoe, S.; et al. Hepatic steatosis is a risk factor for hepatocellular carcinoma in patients with chronic hepatitis C virus infection. Cancer 2003, 97, 3036–3043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, H.; Shiratori, Y.; Moriyama, M.; Arakawa, Y.; Ide, T.; Sata, M.; Inoue, O.; Yano, M.; Tanaka, M.; Fujiyama, S.; et al. Interferon therapy reduces the risk for hepatocellular carcinoma: National surveillance program of cirrhotic and noncirrhotic patients with chronic hepatitis C in Japan. IHIT Study Group. Inhibition of Hepatocarcinogenesis by Interferon Therapy. Ann. Intern. Med. 1999, 131, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Alberti, A.; Piovesan, S. Increased incidence of liver cancer after successful DAA treatment of chronic hepatitis C: Fact or fiction? Liver Int. 2017, 37, 802–808. [Google Scholar] [CrossRef] [PubMed]
- El Kassas, M.; Funk, A.L.; Salaheldin, M.; Shimakawa, Y.; Eltabbakh, M.; Jean, K.; El Tahan, A.; Sweedy, A.T.; Afify, S.; Youssef, N.F.; et al. Increased recurrence rates of hepatocellular carcinoma after DAA therapy in a hepatitis C-infected Egyptian cohort: A comparative analysis. J. Viral Hepat. 2018, 25, 623–630. [Google Scholar] [CrossRef]
- Sasaki, R.; Kanda, T.; Kato, N.; Yokosuka, O.; Moriyama, M. Hepatitis C virus-associated hepatocellular carcinoma after sustained virologic response. World J. Hepatol. 2018, 10, 898–906. [Google Scholar] [CrossRef]
- Totoki, Y.; Tatsuno, K.; Covington, K.R.; Ueda, H.; Creighton, C.J.; Kato, M.; Tsuji, S.; Donehower, L.A.; Slagle, B.L.; Nakamura, H.; et al. Trans-ancestry mutational landscape of hepatocellular carcinoma genomes. Nat. Genet. 2014, 46, 1267–1273. [Google Scholar] [CrossRef]
- Fujimoto, A.; Totoki, Y.; Abe, T.; Boroevich, K.A.; Hosoda, F.; Nguyen, H.H.; Aoki, M.; Hosono, N.; Kubo, M.; Miya, F.; et al. Whole-genome sequencing of liver cancers identifies etiological influences on mutation patterns and recurrent mutations in chromatin regulators. Nat. Genet. 2012, 44, 760–764. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Deng, Q.; Wang, Q.; Li, K.Y.; Dai, J.H.; Li, N.; Zhu, Z.D.; Zhou, B.; Liu, X.Y.; Liu, R.F.; et al. Exome sequencing of hepatitis B virus-associated hepatocellular carcinoma. Nat. Genet. 2012, 44, 1117–1121. [Google Scholar] [CrossRef]
- Schulze, K.; Imbeaud, S.; Letouzé, E.; Alexandrov, L.B.; Calderaro, J.; Rebouissou, S.; Couchy, G.; Meiller, C.; Shinde, J.; Soysouvanh, F.; et al. Exome sequencing of hepatocellular carcinomas identifies new mutational signatures and potential therapeutic targets. Nat. Genet. 2015, 47, 505–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nault, J.C.; Mallet, M.; Pilati, C.; Calderaro, J.; Bioulac-Sage, P.; Laurent, C.; Laurent, A.; Cherqui, D.; Balabaud, C.; Zucman-Rossi, J. High frequency of telomerase reverse-transcriptase promoter somatic mutations in hepatocellular carcinoma and preneoplastic lesions. Nat. Commun. 2013, 4, 2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Zhao, H.; Zhang, X.; Wood, L.D.; Anders, R.A.; Choti, M.A.; Pawlik, T.M.; Daniel, H.D.; Kannangai, R.; Offerhaus, G.J.; et al. Inactivating mutations of the chromatin remodeling gene ARID2 in hepatocellular carcinoma. Nat. Genet. 2011, 43, 828–829. [Google Scholar] [CrossRef] [Green Version]
- Moore, A.; Wu, L.; Chuang, J.C.; Sun, X.; Luo, X.; Gopal, P.; Li, L.; Celen, C.; Zimmer, M.; Zhu, H. Arid1a loss drives non-alcoholic steatohepatitis in mice via epigenetic dysregulation of hepatic lipogenesis and fatty acid oxidation. Hepatology 2018. [Google Scholar] [CrossRef]
- Nose, H.; Imazeki, F.; Ohto, M.; Omata, M. p53 gene mutations and 17p allelic deletions in hepatocellular carcinoma from Japan. Cancer 1993, 72, 355–3560. [Google Scholar] [CrossRef]
- Imazeki, F.; Yokosuka, O.; Ohto, M.; Omata, M. Aflatoxin and p53 abnormality in duck hepatocellular carcinoma. J. Gastroenterol. Hepatol. 1995, 10, 646–649. [Google Scholar] [CrossRef]
- Ray, R.B.; Meyer, K.; Ray, R. Suppression of apoptotic cell death by hepatitis C virus core protein. Virology 1996, 226, 176–182. [Google Scholar] [CrossRef]
- Ray, R.B.; Steele, R.; Meyer, K.; Ray, R. Hepatitis C virus core protein represses p21WAF1/Cip1/Sid1 promoter activity. Gene 1998, 208, 331–336. [Google Scholar] [CrossRef]
- Fei, Q.; Shang, K.; Zhang, J.; Chuai, S.; Kong, D.; Zhou, T.; Fu, S.; Liang, Y.; Li, C.; Chen, Z.; et al. Histone methyltransferase SETDB1 regulates liver cancer cell growth through methylation of p53. Nat. Commun. 2015, 6, 8651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, C.M.; Wei, L.; Law, C.T.; Ho, D.W.; Tsang, F.H.; Au, S.L.; Sze, K.M.; Lee, J.M.; Wong, C.C.; Ng, I.O. Up-regulation of histone methyltransferase SETDB1 by multiple mechanisms in hepatocellular carcinoma promotes cancer metastasis. Hepatology 2016, 63, 474–487. [Google Scholar] [CrossRef] [PubMed]
- Kodama, T.; Newberg, J.Y.; Kodama, M.; Rangel, R.; Yoshihara, K.; Tien, J.C.; Parsons, P.H.; Wu, H.; Finegold, M.J.; Copeland, N.G.; et al. Transposon mutagenesis identifies genes and cellular processes driving epithelial-mesenchymal transition in hepatocellular carcinoma. Proc. Natl. Acad. Sci. USA 2016, 113, E3384–E3393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harding, J.J.; Nandakumar, S.; Armenia, J.; Khalil, D.N.; Albano, M.; Ly, M.; Shia, J.; Hechtman, J.F.; Kundra, R.; El Dika, I.; et al. Prospective Genotyping of Hepatocellular Carcinoma: Clinical Implications of Next Generation Sequencing for Matching Patients to Targeted and Immune Therapies. Clin. Cancer Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Inami, Y.; Waguri, S.; Sakamoto, A.; Kouno, T.; Nakada, K.; Hino, O.; Watanabe, S.; Ando, J.; Iwadate, M.; Yamamoto, M.; et al. Persistent activation of Nrf2 through p62 in hepatocellular carcinoma cells. J. Cell Biol. 2011, 193, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Zavattari, P.; Perra, A.; Menegon, S.; Kowalik, M.A.; Petrelli, A.; Angioni, M.M.; Follenzi, A.; Quagliata, L.; Ledda-Columbano, G.M.; Terracciano, L.; et al. Nrf2, but not β-catenin, mutation represents an early event in rat hepatocarcinogenesis. Hepatology 2015, 62, 851–862. [Google Scholar] [CrossRef] [Green Version]
- Bartolini, D.; Dallaglio, K.; Torquato, P.; Piroddi, M.; Galli, F. Nrf2-p62 autophagy pathway and its response to oxidative stress in hepatocellular carcinoma. Transl. Res. 2018, 193, 54–71. [Google Scholar] [CrossRef]
- Fujimoto, A.; Furuta, M.; Totoki, Y.; Tsunoda, T.; Kato, M.; Shiraishi, Y.; Tanaka, H.; Taniguchi, H.; Kawakami, Y.; Ueno, M.; et al. Whole-genome mutational landscape and characterization of noncoding and structural mutations in liver cancer. Nat. Genet. 2016, 48, 500–509. [Google Scholar] [CrossRef]
- Kojima, H.; Yokosuka, O.; Imazeki, F.; Saisho, H.; Omata, M. Telomerase activity and telomere length in hepatocellular carcinoma and chronic liver disease. Gastroenterology 1997, 112, 493–500. [Google Scholar] [CrossRef]
- Tokino, T.; Matsubara, K. Chromosomal sites for hepatitis B virus integration in human hepatocellular carcinoma. J. Virol. 1991, 65, 6761–6764. [Google Scholar]
- Nagaya, T.; Nakamura, T.; Tokino, T.; Tsurimoto, T.; Imai, M.; Mayumi, T.; Kamino, K.; Yamamura, K.; Matsubara, K. The mode of hepatitis B virus DNA integration in chromosomes of human hepatocellular carcinoma. Genes Dev. 1987, 1, 773–782. [Google Scholar] [CrossRef]
- Imazeki, F.; Omata, M.; Yokosuka, O.; Okuda, K. Integration of hepatitis B virus DNA in hepatocellular carcinoma. Cancer 1986, 58, 1055–1060. [Google Scholar] [CrossRef]
- Wang, M.; Xi, D.; Ning, Q. Virus-induced hepatocellular carcinoma with special emphasis on HBV. Hepatol. Int. 2017, 11, 171–180. [Google Scholar] [CrossRef]
- Meyerson, M.; Counter, C.M.; Eaton, E.N.; Ellisen, L.W.; Steiner, P.; Caddle, S.D.; Ziaugra, L.; Beijersbergen, R.L.; Davidoff, M.J.; Liu, Q.; et al. hEST2, the putative human telomerase catalytic subunit gene, is up-regulated in tumor cells and during immortalization. Cell 1997, 90, 785–795. [Google Scholar] [CrossRef]
- Tokino, T.; Fukushige, S.; Nakamura, T.; Nagaya, T.; Murotsu, T.; Shiga, K.; Aoki, N.; Matsubara, K. Chromosomal translocation and inverted duplication associated with integrated hepatitis B virus in hepatocellular carcinomas. J. Virol. 1987, 61, 3848–3854. [Google Scholar]
- Koike, K.; Shirakata, Y.; Yaginuma, K.; Arii, M.; Takada, S.; Nakamura, I.; Hayashi, Y.; Kawada, M.; Kobayashi, M. Oncogenic potential of hepatitis B virus. Mol. Biol. Med. 1989, 6, 151–160. [Google Scholar]
- Kanda, T.; Yokosuka, O.; Imazeki, F.; Yamada, Y.; Imamura, T.; Fukai, K.; Nagao, K.; Saisho, H. Hepatitis B virus X protein (HBx)-induced apoptosis in HuH-7 cells: Influence of HBV genotype and basal core promoter mutations. Scand. J. Gastroenterol. 2004, 39, 478–485. [Google Scholar] [CrossRef]
- Liu, H.; Shi, W.; Luan, F.; Xu, S.; Yang, F.; Sun, W.; Liu, J.; Ma, C. Hepatitis B virus X protein upregulates transcriptional activation of human telomerase reverse transcriptase. Virus Genes. 2010, 40, 174–182. [Google Scholar] [CrossRef]
- Zou, S.Q.; Qu, Z.L.; Li, Z.F.; Wang, X. Hepatitis B virus X gene induces human telomerase reverse transcriptase mRNA expression in cultured normal human cholangiocytes. World J. Gastroenterol. 2004, 10, 2259–2262. [Google Scholar] [CrossRef] [Green Version]
- Kojima, H.; Kaita, K.D.; Xu, Z.; Ou, J.H.; Gong, Y.; Zhang, M.; Minuk, G.Y. The absence of up-regulation of telomerase activity during regeneration after partial hepatectomy in hepatitis B virus X gene transgenic mice. J. Hepatol. 2003, 39, 262–268. [Google Scholar] [CrossRef]
- Sung, W.K.; Zheng, H.; Li, S.; Chen, R.; Liu, X.; Li, Y.; Lee, N.P.; Lee, W.H.; Ariyaratne, P.N.; Tennakoon, C.; et al. Genome-wide survey of recurrent HBV integration in hepatocellular carcinoma. Nat. Genet. 2012, 44, 765–769. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.; Kawaguchi, T.; Nakamoto, S.; Kawaguchi, A.; Kanda, T.; Imazeki, F.; Kuromatsu, R.; Sumie, S.; Satani, M.; Yamada, S.; et al. Effect of occult hepatitis B virus infection on the early-onset of hepatocellular carcinoma in patients with hepatitis C virus infection. Oncol. Rep. 2013, 30, 2049–2055. [Google Scholar] [CrossRef] [PubMed]
- Mason, W.S.; Gill, U.S.; Litwin, S.; Zhou, Y.; Peri, S.; Pop, O.; Hong, M.L.; Naik, S.; Quaglia, A.; Bertoletti, A.; et al. HBV DNA Integration and Clonal Hepatocyte Expansion in Chronic Hepatitis B Patients Considered Immune Tolerant. Gastroenterology 2016, 151, 986–998. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.T.F.; Litwin, S.; Dolman, G.E.; Bertoletti, A.; Mason, W.S. Immune Tolerant Chronic Hepatitis B: The Unrecognized Risks. Viruses 2017, 9, 96. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Kanda, T.; Nakamoto, S.; Jiang, X.; Nakamura, M.; Sasaki, R.; Haga, Y.; Shirasawa, H.; Yokosuka, O. Cooperative effects of hepatitis B virus and TNF may play important roles in the activation of metabolic pathways through the activation of NF-κB. Int. J. Mol. Med. 2016, 38, 475–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y. Hepatitis B Virus-Associated Hepatocellular Carcinoma. Adv. Exp. Med. Biol. 2017, 1018, 11–21. [Google Scholar] [CrossRef]
- Jiang, X.; Kanda, T.; Wu, S.; Nakamura, M.; Miyamura, T.; Nakamoto, S.; Banerjee, A.; Yokosuka, O. Regulation of microRNA by hepatitis B virus infection and their possible association with control of innate immunity. World J. Gastroenterol. 2014, 20, 7197–7206. [Google Scholar] [CrossRef]
- Sarkar, N.; Panigrahi, R.; Pal, A.; Biswas, A.; Singh, S.P.; Kar, S.K.; Bandopadhyay, M.; Das, D.; Saha, D.; Kanda, T.; et al. Expression of microRNA-155 correlates positively with the expression of Toll-like receptor 7 and modulates hepatitis B virus via C/EBP-β in hepatocytes. J. Viral Hepat. 2015, 22, 817–827. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Bian, H.; Li, F.; Li, X.; Zhang, D.; Sun, S.; Song, S.; Zhu, Q.; Ren, W.; Qin, C.; et al. HBeAg induces the expression of macrophage miR-155 to accelerate liver injury via promoting production of inflammatory cytokines. Cell. Mol. Life Sci. 2018, 75, 2627–2641. [Google Scholar] [CrossRef] [PubMed]
- Wong, Q.W.; Lung, R.W.; Law, P.T.; Lai, P.B.; Chan, K.Y.; To, K.F.; Wong, N. MicroRNA-223 is commonly repressed in hepatocellular carcinoma and potentiates expression of Stathmin1. Gastroenterology 2008, 135, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, S.; Hu, T.; Liu, S.; He, Y.; Sun, S. Up-regulated microRNA-143 transcribed by nuclear factor kappa B enhances hepatocarcinoma metastasis by repressing fibronectin expression. Hepatology 2009, 50, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Ma, Z.; Wang, D.; Zhao, W.; Chen, L.; Wang, G. MicroRNA-602 regulating tumor suppressive gene RASSF1A is overexpressed in hepatitis B virus-infected liver and hepatocellular carcinoma. Cancer Biol. Ther. 2010, 9, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lu, Y.; Toh, S.T.; Sung, W.K.; Tan, P.; Chow, P.; Chung, A.Y.; Jooi, L.L.; Lee, C.G. Lethal-7 is down-regulated by the hepatitis B virus x protein and targets signal transducer and activator of transcription 3. J. Hepatol. 2010, 53, 57–66. [Google Scholar] [CrossRef]
- Huang, J.; Wang, Y.; Guo, Y.; Sun, S. Down-regulated microRNA-152 induces aberrant DNA methylation in hepatitis B virus-related hepatocellular carcinoma by targeting DNA methyltransferase 1. Hepatology 2010, 52, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Li, L.M.; Hu, Z.B.; Zhou, Z.X.; Chen, X.; Liu, F.Y.; Zhang, J.F.; Shen, H.B.; Zhang, C.Y.; Zen, K. Serum microRNA profiles serve as novel biomarkers for HBV infection and diagnosis of HBV-positive hepatocarcinoma. Cancer Res. 2010, 70, 9798–9807. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Wong, C.C.; Tung, E.K.; Lee, J.M.; Wong, C.M.; Ng, I.O. Deregulation of microRNA expression occurs early and accumulates in early stages of HBV-associated multistep hepatocarcinogenesis. J. Hepatol. 2011, 54, 1177–1184. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Deng, L.; Zhao, L.; Li, X.; Zhang, F.; Xia, Y.; Gao, Y.; Wang, X.; Sun, B. miR-22 promotes HBV-related hepatocellular carcinoma development in males. Clin. Cancer Res. 2011, 17, 5593–5603. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.M.; Wang, Y.; Fan, C.G.; Xu, F.F.; Sun, W.S.; Liu, Y.G.; Jia, J.H. miR-29c targets TNFAIP3, inhibits cell proliferation and induces apoptosis in hepatitis B virus-related hepatocellular carcinoma. Biochem. Biophys. Res. Commun. 2011, 411, 586–592. [Google Scholar] [CrossRef]
- Qi, P.; Cheng, S.Q.; Wang, H.; Li, N.; Chen, Y.F.; Gao, C.F. Serum microRNAs as biomarkers for hepatocellular carcinoma in Chinese patients with chronic hepatitis B virus infection. PLoS ONE 2011, 6, e28486. [Google Scholar] [CrossRef]
- Cardin, R.; Piciocchi, M.; Sinigaglia, A.; Lavezzo, E.; Bortolami, M.; Kotsafti, A.; Cillo, U.; Zanus, G.; Mescoli, C.; Rugge, M.; et al. Oxidative DNA damage correlates with cell immortalization and mir-92 expression in hepatocellular carcinoma. BMC Cancer 2012, 12, 177. [Google Scholar] [CrossRef]
- Li, L.; Guo, Z.; Wang, J.; Mao, Y.; Gao, Q. Serum miR-18a: A potential marker for hepatitis B virus-related hepatocellular carcinoma screening. Dig. Dis. Sci. 2012, 57, 2910–2916. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Tan, D.; Hou, Z.; Hu, Z.; Liu, G. miR-338-3p is down-regulated by hepatitis B virus X and inhibits cell proliferation by targeting the 3’-UTR region of CyclinD1. Int. J. Mol. Sci. 2012, 13, 8514–8539. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Li, Q.J.; Feng, Y.; Zhang, Y.; Markowitz, G.J.; Ning, S.; Deng, Y.; Zhao, J.; Jiang, S.; Yuan, Y.; et al. TGF-β-miR-34a-CCL22 signaling-induced Treg cell recruitment promotes venous metastases of HBV-positive hepatocellular carcinoma. Cancer Cell 2012, 22, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Xiang, T.; Ren, G.; Tan, C.; Liu, R.; Xu, X.; Wu, Z. miR-101 is down-regulated by the hepatitis B virus x protein and induces aberrant DNA methylation by targeting DNA methyltransferase 3A. Cell. Signal. 2013, 25, 439–446. [Google Scholar] [CrossRef]
- Li, C.; Wang, Y.; Wang, S.; Wu, B.; Hao, J.; Fan, H.; Ju, Y.; Ding, Y.; Chen, L.; Chu, X.; et al. Hepatitis B virus mRNA-mediated miR-122 inhibition upregulates PTTG1-binding protein, which promotes hepatocellular carcinoma tumor growth and cell invasion. J. Virol. 2013, 87, 2193–2205. [Google Scholar] [CrossRef]
- Xu, X.; Fan, Z.; Kang, L.; Han, J.; Jiang, C.; Zheng, X.; Zhu, Z.; Jiao, H.; Lin, J.; Jiang, K.; et al. Hepatitis B virus X protein represses miRNA-148a to enhance tumorigenesis. J. Clin. Investig. 2013, 123, 630–645. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Xu, X. MicroRNA-22 is down-regulated in hepatitis B virus-related hepatocellular carcinoma. Biomed. Pharmacother. 2013, 67, 375–380. [Google Scholar] [CrossRef]
- Lan, S.H.; Wu, S.Y.; Zuchini, R.; Lin, X.Z.; Su, I.J.; Tsai, T.F.; Lin, Y.J.; Wu, C.T.; Liu, H.S. Autophagy suppresses tumorigenesis of hepatitis B virus-associated hepatocellular carcinoma through degradation of microRNA-224. Hepatology 2014, 59, 505–517. [Google Scholar] [CrossRef]
- Fu, Y.; Wei, X.; Tang, C.; Li, J.; Liu, R.; Shen, A.; Wu, Z. Circulating microRNA-101 as a potential biomarker for hepatitis B virus-related hepatocellular carcinoma. Oncol. Lett. 2013, 6, 1811–1815. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Shi, W.; Gao, Y.; Yang, B.; Jing, X.; Shan, S.; Wang, Y.; Du, Z. Analysis of microRNA expression profiles in human hepatitis B virus-related hepatocellular carcinoma. Clin. Lab. 2013, 59, 1009–1015. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhang, J.; Cui, M.; Liu, F.; You, X.; Du, Y.; Gao, Y.; Zhang, S.; Lu, Z.; Ye, L.; et al. Hepatitis B virus X protein inhibits tumor suppressor miR-205 through inducing hypermethylation of miR-205 promoter to enhance carcinogenesis. Neoplasia 2013, 15, 1282–1291. [Google Scholar] [CrossRef]
- Sheng, Y.; Li, J.; Zou, C.; Wang, S.; Cao, Y.; Zhang, J.; Huang, A.; Tang, H. Downregulation of miR-101-3p by hepatitis B virus promotes proliferation and migration of hepatocellular carcinoma cells by targeting Rab5a. Arch. Virol. 2014, 159, 2397–2410. [Google Scholar] [CrossRef] [PubMed]
- Dang, Y.W.; Zeng, J.; He, R.Q.; Rong, M.H.; Luo, D.Z.; Chen, G. Effects of miR-152 on cell growth inhibition, motility suppression and apoptosis induction in hepatocellular carcinoma cells. Asian Pac. J. Cancer Prev. 2014, 15, 4969–4976. [Google Scholar] [CrossRef] [PubMed]
- Lan, S.H.; Wu, S.Y.; Zuchini, R.; Lin, X.Z.; Su, I.J.; Tsai, T.F.; Lin, Y.J.; Wu, C.T.; Liu, H.S. Autophagy-preferential degradation of MIR224 participates in hepatocellular carcinoma tumorigenesis. Autophagy 2014, 10, 1687–1689. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.L.; Wang, W.; Jia, W.D. Diagnostic and prognostic significance of serum miR-24-3p in HBV-related hepatocellular carcinoma. Med. Oncol. 2014, 31, 177. [Google Scholar] [CrossRef]
- Bandopadhyay, M.; Banerjee, A.; Sarkar, N.; Panigrahi, R.; Datta, S.; Pal, A.; Singh, S.P.; Biswas, A.; Chakrabarti, S.; Chakravarty, R. Tumor suppressor micro RNA miR-145 and onco micro RNAs miR-21 and miR-222 expressions are differentially modulated by hepatitis B virus X protein in malignant hepatocytes. BMC Cancer 2014, 14, 721. [Google Scholar] [CrossRef] [PubMed]
- Xing, T.J.; Jiang, D.F.; Huang, J.X.; Xu, Z.L. Expression and clinical significance of miR-122 and miR-29 in hepatitis B virus-related liver disease. Genet. Mol. Res. 2014, 13, 7912–7918. [Google Scholar] [CrossRef]
- Zhao, Q.; Li, T.; Qi, J.; Liu, J.; Qin, C. The miR-545/374a cluster encoded in the Ftx lncRNA is overexpressed in HBV-related hepatocellular carcinoma and promotes tumorigenesis and tumor progression. PLoS ONE 2014, 9, e109782. [Google Scholar] [CrossRef]
- Gao, H.; Liu, C. miR-429 represses cell proliferation and induces apoptosis in HBV-related HCC. Biomed. Pharmacother. 2014, 68, 943–949. [Google Scholar] [CrossRef]
- Liu, F.Y.; Zhou, S.J.; Deng, Y.L.; Zhang, Z.Y.; Zhang, E.L.; Wu, Z.B.; Huang, Z.Y.; Chen, X.P. MiR-216b is involved in pathogenesis and progression of hepatocellular carcinoma through HBx-miR-216b-IGF2BP2 signaling pathway. Cell Death Dis. 2015, 6, e1670. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Lu, Z.; Chen, B.; Dong, P.; Zheng, J. microRNA-150: A promising novel biomarker for hepatitis B virus-related hepatocellular carcinoma. Diagn. Pathol. 2015, 10, 129. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Chen, J.; Wang, D.; Peng, H.; Tan, X.; Xiong, D.; Huang, A.; Tang, H. Upregulated in Hepatitis B virus-associated hepatocellular carcinoma cells, miR-331-3p promotes proliferation of hepatocellular carcinoma cells by targeting ING5. Oncotarget 2015, 6, 38093–38106. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Sun, X.; Wang, L.; Tang, S.; Yan, C. Downregulation of MicroRNA-145 Caused by Hepatitis B Virus X Protein Promotes Expression of CUL5 and Contributes to Pathogenesis of Hepatitis B Virus-Associated Hepatocellular Carcinoma. Cell. Physiol. Biochem. 2015, 37, 1547–1559. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Cao, J.; Liu, Y.; Mei, H.; Zhang, Y.; Xu, W. MicroRNA-519a promotes proliferation and inhibits apoptosis of hepatocellular carcinoma cells by targeting FOXF2. FEBS Open Bio 2015, 5, 893–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, C.M.; Jiang, Z.Z.; Yu, X.J.; Fan, C.G.; Xu, F.F.; Zhang, Q.; Li, L.I.; Li, R.F.; Sun, W.S.; et al. MicroRNA-34c targets TGFB-induced factor homeobox 2, represses cell proliferation and induces apoptosis in hepatitis B virus-related hepatocellular carcinoma. Oncol. Lett. 2015, 10, 3095–3102. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A.; Ghosh, A.; Datta, S.; Dasgupta, D.; Das, S.; Ray, S.; Gupta, S.; Datta, S.; Chowdhury, A.; Chatterjee, R.; et al. Hepatic miR-126 is a potential plasma biomarker for detection of hepatitis B virus infected hepatocellular carcinoma. Int. J. Cancer 2016, 138, 2732–2744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Dong, X.; Yu, D.; Wang, X. Serum miR-96 is a promising biomarker for hepatocellular carcinoma in patients with chronic hepatitis B virus infection. Int. J. Clin. Exp. Med. 2015, 8, 18462–18468. [Google Scholar] [PubMed]
- Chen, S.; Chen, H.; Gao, S.; Qiu, S.; Zhou, H.; Yu, M.; Tu, J. Differential expression of plasma microRNA-125b in hepatitis B virus-related liver diseases and diagnostic potential for hepatitis B virus-induced hepatocellular carcinoma. Hepatol. Res. 2017, 47, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Yen, C.S.; Su, Z.R.; Lee, Y.P.; Liu, I.T.; Yen, C.J. miR-106b promotes cancer progression in hepatitis B virus-associated hepatocellular carcinoma. World J. Gastroenterol. 2016, 22, 5183–5192. [Google Scholar] [CrossRef]
- Kong, X.X.; Lv, Y.R.; Shao, L.P.; Nong, X.Y.; Zhang, G.L.; Zhang, Y.; Fan, H.X.; Liu, M.; Li, X.; Tang, H. HBx-induced MiR-1269b in NF-κB dependent manner upregulates cell division cycle 40 homolog (CDC40) to promote proliferation and migration in hepatoma cells. J. Transl. Med. 2016, 14, 189. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhang, Y.; Wang, P.; Wang, H.; Su, H.; Zhou, X.; Zhang, L. HBX Protein-Induced Downregulation of microRNA-18a is Responsible for Upregulation of Connective Tissue Growth Factor in HBV Infection-Associated Hepatocarcinoma. Med. Sci. Monit. 2016, 22, 2492–2500. [Google Scholar] [CrossRef] [Green Version]
- Qiao, D.D.; Yang, J.; Lei, X.F.; Mi, G.L.; Li, S.L.; Li, K.; Xu, C.Q.; Yang, H.L. Expression of microRNA-122 and microRNA-22 in HBV-related liver cancer and the correlation with clinical features. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 742–747. [Google Scholar] [PubMed]
- Qin, X.; Chen, J.; Wu, L.; Liu, Z. MiR-30b-5p acts as a tumor suppressor, repressing cell proliferation and cell cycle in human hepatocellular carcinoma. Biomed. Pharmacother. 2017, 89, 742–750. [Google Scholar] [CrossRef] [PubMed]
- Bai, P.S.; Xia, N.; Sun, H.; Kong, Y. Pleiotrophin, a target of miR-384, promotes proliferation, metastasis and lipogenesis in HBV-related hepatocellular carcinoma. J. Cell. Mol. Med. 2017, 21, 3023–3043. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Zhang, W.; Gong, L.; Huang, X. MicroRNA-125a-5p Inhibits Cell Proliferation and Induces Apoptosis in Hepatitis B Virus-Related Hepatocellular Carcinoma by Downregulation of ErbB3. Oncol. Res. 2017. [Google Scholar] [CrossRef]
- Zhao, Q.; Sun, X.; Liu, C.; Li, T.; Cui, J.; Qin, C. Expression of the microRNA-143/145 cluster is decreased in hepatitis B virus-associated hepatocellular carcinoma and may serve as a biomarker for tumorigenesis in patients with chronic hepatitis B. Oncol. Lett. 2018, 15, 6115–6122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quoc, N.B.; Phuong, N.D.N.; Ngan, T.K.; Linh, N.T.M.; Cuong, P.H.; Chau, N.N.B. Expression of Plasma hsa-miR122 in HBV-Related Hepatocellular Carcinoma (HCC) in Vietnamese Patients. Microrna 2018, 7, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.R.; Nabinger, S.C.; Lee, S.; Sahu, S.S.; Althouse, S.; Saxena, R.; Johnson, M.S.; Chalasani, N.; Gawrieh, S.; Kota, J. Lower expression of tumor microRNA-26a is associated with higher recurrence in patients with hepatocellular carcinoma undergoing surgical treatment. J. Surg. Oncol. 2018, 118, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Guo, Y.; Liu, X.; Wang, T.; Tong, X.; Lei, K.; Wang, J.; Huang, D.; Xu, Q. The tumor suppressive miR-302c-3p inhibits migration and invasion of hepatocellular carcinoma cells by targeting TRAF4. J. Cancer 2018, 9, 2693–2701. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, Z.X.; Huang, F.; Yuan, X.W.; Deng, L.; Tang, D. MicroRNA-1271 functions as a potential tumor suppressor in hepatitis B virus-associated hepatocellular carcinoma through the AMPK signaling pathway by binding to CCNA1. J. Cell. Physiol. 2018. [Google Scholar] [CrossRef]
- Qiu, L.; Wang, T.; Xu, X.; Wu, Y.; Tang, Q.; Chen, K. Long Non-Coding RNAs in Hepatitis B Virus-Related Hepatocellular Carcinoma: Regulation, Functions, and Underlying Mechanisms. Int. J. Mol. Sci. 2017, 18, 2505. [Google Scholar] [CrossRef]
- Cui, S.; Qian, Z.; Chen, Y.; Li, L.; Li, P.; Ding, H. Screening of up- and downregulation of circRNAs in HBV-related hepatocellular carcinoma by microarray. Oncol. Lett. 2018, 15, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Cui, S.; Zhao, W.; Qian, Z.; Liu, H.; Chen, Y.; Lv, F.; Ding, H.G. Screening and bioinformatics analysis of circular RNA expression profiles in hepatitis B-related hepatocellular carcinoma. Cancer Biomark. 2018, 22, 631–640. [Google Scholar] [CrossRef]
- Matsuda, Y.; Ichida, T.; Genda, T.; Yamagiwa, S.; Aoyagi, Y.; Asakura, H. Loss of p16 contributes to p27 sequestration by cyclin D(1)-cyclin-dependent kinase 4 complexes and poor prognosis in hepatocellular carcinoma. Clin. Cancer Res. 2003, 9, 3389–3396. [Google Scholar] [PubMed]
- Pezzuto, F.; Buonaguro, L.; Buonaguro, F.M.; Tornesello, M.L. The Role of Circulating Free DNA and MicroRNA in Non-Invasive Diagnosis of HBV- and HCV-Related Hepatocellular Carcinoma. Int. J. Mol. Sci. 2018, 19, 1007. [Google Scholar] [CrossRef]
- Tian, Y.; Ou, J.H. Genetic and epigenetic alterations in hepatitis B virus-associated hepatocellular carcinoma. Virol. Sin. 2015, 30, 85–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Kanda, T.; Imazeki, F.; Arai, M.; Yonemitsu, Y.; Nakamoto, S.; Fujiwara, K.; Fukai, K.; Nomura, F.; Yokosuka, O. Hepatitis B virus e antigen downregulates cytokine production in human hepatoma cell lines. Viral Immunol. 2010, 23, 467–476. [Google Scholar] [CrossRef]
- Hadziyannis, E.; Laras, A. Viral Biomarkers in Chronic HBeAg Negative HBV Infection. Genes 2018, 9, 469. [Google Scholar] [CrossRef] [PubMed]
- Hildt, E.; Hofschneider, P.H. The PreS2 activators of the hepatitis B virus: Activators of tumour promoter pathways. Recent Results Cancer Res. 1998, 154, 315–329. [Google Scholar] [PubMed]
- Liu, P.; Zhang, H.; Liang, X.; Ma, H.; Luan, F.; Wang, B.; Bai, F.; Gao, L.; Ma, C. HBV preS2 promotes the expression of TAZ via miRNA-338-3p to enhance the tumorigenesis of hepatocellular carcinoma. Oncotarget 2015, 6, 29048–29059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.L.; Wu, D.B.; Tao, Y.C.; Chen, L.L.; Liu, C.P.; Chen, E.Q.; Tang, H. The truncated mutant HBsAg expression increases the tumorigenesis of hepatitis B virus by regulating TGF-β/Smad signaling pathway. Virol. J. 2018, 15, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Zhang, X.; Ge, Q.; Yuan, C.; Chu, L.; Liang, H.F.; Liao, Z.; Liu, Q.; Zhang, Z.; Zhang, B. CRISPR/Cas9-mediated knockout of HBsAg inhibits proliferation and tumorigenicity of HBV-positive hepatocellular carcinoma cells. J. Cell. Biochem. 2018, 119, 8419–8431. [Google Scholar] [CrossRef] [PubMed]
- Kanda, T.; Wu, S.; Sasaki, R.; Nakamura, M.; Haga, Y.; Jiang, X.; Nakamoto, S.; Yokosuka, O. HBV Core Protein Enhances Cytokine Production. Diseases 2015, 3, 213–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Kanda, T.; Imazeki, F.; Nakamoto, S.; Tanaka, T.; Arai, M.; Roger, T.; Shirasawa, H.; Nomura, F.; Yokosuka, O. Hepatitis B virus e antigen physically associates with receptor-interacting serine/threonine protein kinase 2 and regulates IL-6 gene expression. J. Infect. Dis. 2012, 206, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.T.; Billaud, J.N.; Sällberg, M.; Guidotti, L.G.; Chisari, F.V.; Jones, J.; Hughes, J.; Milich, D.R. A function of the hepatitis B virus precore protein is to regulate the immune response to the core antigen. Proc. Natl. Acad. Sci. USA 2004, 101, 14913–14918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Sällberg, M.; Hughes, J.; Jones, J.; Guidotti, L.G.; Chisari, F.V.; Billaud, J.N.; Milich, D.R. Immune tolerance split between hepatitis B virus precore and core proteins. J. Virol. 2005, 79, 3016–3027. [Google Scholar] [CrossRef]
- Yaginuma, K.; Kobayashi, H.; Kobayashi, M.; Morishima, T.; Matsuyama, K.; Koike, K. Multiple integration site of hepatitis B virus DNA in hepatocellular carcinoma and chronic active hepatitis tissues from children. J. Virol. 1987, 61, 1808–1813. [Google Scholar]
- Zhou, Y.Z.; Butel, J.S.; Li, P.J.; Finegold, M.J.; Melnick, J.L. Integrated state of subgenomic fragments of hepatitis B virus DNA in hepatocellular carcinoma from mainland China. J. Natl. Cancer Inst. 1987, 79, 223–231. [Google Scholar]
- Kim, C.M.; Koike, K.; Saito, I.; Miyamura, T.; Jay, G. HBx gene of hepatitis B virus induces liver cancer in transgenic mice. Nature 1991, 351, 317–320. [Google Scholar] [CrossRef]
- Wollersheim, M.; Debelka, U.; Hofschneider, P.H. A transactivating function encoded in the hepatitis B virus X gene is conserved in the integrated state. Oncogene 1988, 3, 545–552. [Google Scholar] [PubMed]
- Kekulé, A.S.; Lauer, U.; Weiss, L.; Luber, B.; Hofschneider, P.H. Hepatitis B virus transactivator HBx uses a tumour promoter signalling pathway. Nature 1993, 361, 742–745. [Google Scholar] [CrossRef]
- Choi, B.H.; Choi, M.; Jeon, H.Y.; Rho, H.M. Hepatitis B viral X protein overcomes inhibition of E2F1 activity by pRb on the human Rb gene promoter. DNA Cell Biol. 2001, 20, 75–80. [Google Scholar] [CrossRef]
- Staib, F.; Hussain, S.P.; Hofseth, L.J.; Wang, X.W.; Harris, C.C. TP53 and liver carcinogenesis. Hum. Mutat. 2003, 21, 201–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, M.Y.; Kim, C.M.; Park, Y.M.; Ryu, W.S. Hepatitis B virus X protein is essential for the activation of Wnt/beta-catenin signaling in hepatoma cells. Hepatology 2004, 39, 1683–1693. [Google Scholar] [CrossRef]
- Longato, L.; de la Monte, S.; Kuzushita, N.; Horimoto, M.; Rogers, A.B.; Slagle, B.L.; Wands, J.R. Overexpression of insulin receptor substrate-1 and hepatitis Bx genes causes premalignant alterations in the liver. Hepatology 2009, 49, 1935–1943. [Google Scholar] [CrossRef] [PubMed]
- Keng, V.W.; Tschida, B.R.; Bell, J.B.; Largaespada, D.A. Modeling hepatitis B virus X-induced hepatocellular carcinoma in mice with the Sleeping Beauty transposon system. Hepatology 2011, 53, 781–790. [Google Scholar] [CrossRef]
- Wang, C.; Yang, W.; Yan, H.X.; Luo, T.; Zhang, J.; Tang, L.; Wu, F.Q.; Zhang, H.L.; Yu, L.X.; Zheng, L.Y.; et al. Hepatitis B virus X (HBx) induces tumorigenicity of hepatic progenitor cells in 3,5-diethoxycarbonyl-1,4-dihydrocollidine-treated HBx transgenic mice. Hepatology 2012, 55, 108–120. [Google Scholar] [CrossRef]
- Von Olshausen, G.; Quasdorff, M.; Bester, R.; Arzberger, S.; Ko, C.; van de Klundert, M.; Zhang, K.; Odenthal, M.; Ringelhan, M.; Niessen, C.M.; et al. Hepatitis B virus promotes β-catenin-signalling and disassembly of adherens junctions in a Src kinase dependent fashion. Oncotarget 2018, 9, 33947–33960. [Google Scholar] [CrossRef]
- Singh, A.K.; Swarnalatha, M.; Kumar, V. c-ETS1 facilitates G1/S-phase transition by up-regulating cyclin E and CDK2 genes and cooperates with hepatitis B virus X protein for their deregulation. J. Biol. Chem. 2011, 286, 21961–21970. [Google Scholar] [CrossRef]
- Luo, L.; Chen, S.; Gong, Q.; Luo, N.; Lei, Y.; Guo, J.; He, S. Hepatitis B virus X protein modulates remodelling of minichromosomes related to hepatitis B virus replication in HepG2 cells. Int. J. Mol. Med. 2013, 31, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Saeed, U.; Kim, J.; Piracha, Z.Z.; Kwon, H.; Jung, J.; Chwae, Y.J.; Park, S.; Shin, H.J.; Kim, K. Parvulin 14 and parvulin 17 bind to HBx and cccDNA and upregulate HBV replication from cccDNA to virion in a HBx-dependent manner. J. Virol. 2019, 93, e01840-18. [Google Scholar] [CrossRef] [PubMed]
- Swarnalatha, M.; Singh, A.K.; Kumar, V. Promoter occupancy of MLL1 histone methyltransferase seems to specify the proliferative and apoptotic functions of E2F1 in a tumour microenvironment. J. Cell Sci. 2013, 126, 4636–4646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.H.; Tseng, Y.H.; Lin, W.R.; Hung, G.; Chen, T.C.; Wang, T.H.; Lee, W.C.; Yeh, C.T. HBV polymerase overexpression due to large core gene deletion enhances hepatoma cell growth by binding inhibition of microRNA-100. Oncotarget 2016, 7, 9448–9461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, H.J.; Chen, X.; Yu, Y.; Lee, H.K.; Song, C.H.; Choe, H.; Lee, S.; Kim, H.J.; Hong, S.T. A critical role of hepatitis B virus polymerase in cirrhosis, hepatocellular carcinoma, and steatosis. FEBS Open Bio 2017, 8, 130–145. [Google Scholar] [CrossRef] [Green Version]
- Kakumu, S.; Okumura, A.; Ishikawa, T.; Yano, M.; Enomoto, A.; Nishimura, H.; Yoshioka, K.; Yoshika, Y. Serum levels of IL-10, IL-15 and soluble tumour necrosis factor-alpha (TNF-alpha) receptors in type C chronic liver disease. Clin. Exp. Immunol. 1997, 109, 458–463. [Google Scholar] [CrossRef]
- Kanda, T.; Yokosuka, O.; Omata, M. Hepatitis C virus and hepatocellular carcinoma. Biology 2013, 2, 304–316. [Google Scholar] [CrossRef]
- Braconi, C.; Valeri, N.; Gasparini, P.; Huang, N.; Taccioli, C.; Nuovo, G.; Suzuki, T.; Croce, C.M.; Patel, T. Hepatitis C virus proteins modulate microRNA expression and chemosensitivity in malignant hepatocytes. Clin. Cancer Res. 2010, 16, 957–966. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wei, W.; Cheng, N.; Wang, K.; Li, B.; Jiang, X.; Sun, S. Hepatitis C virus-induced up-regulation of microRNA-155 promotes hepatocarcinogenesis by activating Wnt signaling. Hepatology 2012, 56, 1631–1640. [Google Scholar] [CrossRef]
- Hsu, S.H.; Wang, B.; Kota, J.; Yu, J.; Costinean, S.; Kutay, H.; Yu, L.; Bai, S.; La Perle, K.; Chivukula, R.R.; et al. Essential metabolic, anti-inflammatory, and anti-tumorigenic functions of miR-122 in liver. J. Clin. Investig. 2012, 122, 2871–2883. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Li, F.; Taylor, E.W. Can tobacco use promote HCV-induced miR-122 hijacking and hepatocarcinogenesis? Med. Hypotheses 2013, 80, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Salvi, A.; Abeni, E.; Portolani, N.; Barlati, S.; De Petro, G. Human hepatocellular carcinoma cell-specific miRNAs reveal the differential expression of miR-24 and miR-27a in cirrhotic/non-cirrhotic HCC. Int. J. Oncol. 2013, 42, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Elfimova, N.; Sievers, E.; Eischeid, H.; Kwiecinski, M.; Noetel, A.; Hunt, H.; Becker, D.; Frommolt, P.; Quasdorff, M.; Steffen, H.M.; et al. Control of mitogenic and motogenic pathways by miR-198, diminishing hepatoma cell growth and migration. Biochim. Biophys. Acta 2013, 1833, 1190–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, M.; Deiters, A. MicroRNA miR-122 as a therapeutic target for oligonucleotides and small molecules. Curr Med Chem. 2013, 20, 3629–3640. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Xie, Y.; Yang, P.; Chen, P.; Zhang, L. HCV core protein-induced down-regulation of microRNA-152 promoted aberrant proliferation by regulating Wnt1 in HepG2 cells. PLoS ONE 2014, 9, e81730. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Shrivastava, S.; Bhanja Chowdhury, J.; Ray, R.; Ray, R.B. Transcriptional suppression of miR-181c by hepatitis C virus enhances homeobox A1 expression. J. Virol. 2014, 88, 7929–7940. [Google Scholar] [CrossRef]
- Xu, G.; Yang, F.; Ding, C.L.; Wang, J.; Zhao, P.; Wang, W.; Ren, H. MiR-221 accentuates IFN׳s anti-HCV effect by downregulating SOCS1 and SOCS3. Virology 2014, 462–463, 343–350. [Google Scholar] [CrossRef]
- Pan, L.; Ren, F.; Rong, M.; Dang, Y.; Luo, Y.; Luo, D.; Chen, G. Correlation between down-expression of miR-431 and clinicopathological significance in HCC tissues. Clin. Transl. Oncol. 2015, 17, 557–563. [Google Scholar] [CrossRef]
- El-Abd, N.E.; Fawzy, N.A.; El-Sheikh, S.M.; Soliman, M.E. Circulating miRNA-122, miRNA-199a, and miRNA-16 as Biomarkers for Early Detection of Hepatocellular Carcinoma in Egyptian Patients with Chronic Hepatitis C Virus Infection. Mol. Diagn. Ther. 2015, 19, 213–220. [Google Scholar] [CrossRef]
- Devhare, P.B.; Steele, R.; Di Bisceglie, A.M.; Kaplan, D.E.; Ray, R.B. Differential Expression of MicroRNAs in Hepatitis C Virus-Mediated Liver Disease Between African Americans and Caucasians: Implications for Racial Health Disparities. Gene Expr. 2017, 17, 89–98. [Google Scholar] [CrossRef]
- Shiu, T.Y.; Shih, Y.L.; Feng, A.C.; Lin, H.H.; Huang, S.M.; Huang, T.Y.; Hsieh, C.B.; Chang, W.K.; Hsieh, T.Y. HCV core inhibits hepatocellular carcinoma cell replicative senescence through downregulating microRNA-138 expression. J. Mol. Med. 2017, 95, 629–639. [Google Scholar] [CrossRef]
- Shaker, O.; Alhelf, M.; Morcos, G.; Elsharkawy, A. miRNA-101-1 and miRNA-221 expressions and their polymorphisms as biomarkers for early diagnosis of hepatocellular carcinoma. Infect. Genet. Evol. 2017, 51, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Shehata, R.H.; Abdelmoneim, S.S.; Osman, O.A.; Hasanain, A.F.; Osama, A.; Abdelmoneim, S.S.; Toraih, E.A. Deregulation of miR-34a and Its Chaperon Hsp70 in Hepatitis C virus-Induced Liver Cirrhosis and Hepatocellular Carcinoma Patients. Asian Pac. J. Cancer Prev. 2017, 18, 2395–2401. [Google Scholar] [PubMed]
- Sur, S.; Sasaki, R.; Devhare, P.; Steele, R.; Ray, R.; Ray, R.B. Association between MicroRNA-373 and Long Noncoding RNA NORAD in Hepatitis C Virus-Infected Hepatocytes Impairs Wee1 Expression for Growth Promotion. J. Virol. 2018, 92, e01215-18. [Google Scholar] [CrossRef] [PubMed]
- Rashad, N.M.; El-Shal, A.S.; Shalaby, S.M.; Mohamed, S.Y. Serum miRNA-27a and miRNA-18b as potential predictive biomarkers of hepatitis C virus-associated hepatocellular carcinoma. Mol. Cell. Biochem. 2018, 447, 125–136. [Google Scholar] [CrossRef]
- Hou, W.; Bonkovsky, H.L. Non-coding RNAs in hepatitis C-induced hepatocellular carcinoma: Dysregulation and implications for early detection, diagnosis and therapy. World J. Gastroenterol. 2013, 19, 7836–7845. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, C.; Zhao, Y.; Li, M.; Wu, L.; Yang, X.; Wan, X.; Wang, A.; Zhang, M.Q.; Sang, X.; et al. Long non-coding RNA expression profiles of hepatitis C virus-related dysplasia and hepatocellular carcinoma. Oncotarget 2015, 6, 43770–43778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamel, M.M.; Matboli, M.; Sallam, M.; Montasser, I.F.; Saad, A.S.; El-Tawdi, A.H.F. Investigation of long noncoding RNAs expression profile as potential serum biomarkers in patients with hepatocellular carcinoma. Transl. Res. 2016, 168, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Fu, N.; Niu, X.; Wang, Y.; Du, H.; Wang, B.; Du, J.; Li, Y.; Wang, R.; Zhang, Y.; Zhao, S.; et al. Role of LncRNA-activated by transforming growth factor beta in the progression of hepatitis C virus-related liver fibrosis. Discov. Med. 2016, 22, 29–42. [Google Scholar]
- Kanda, T.; Tada, M.; Imazeki, F.; Yokosuka, O.; Nagao, K.; Saisho, H. 5-aza-2’-deoxycytidine sensitizes hepatoma and pancreatic cancer cell lines. Oncol. Rep. 2005, 14, 975–979. [Google Scholar] [CrossRef]
- Ray, R.B.; Lagging, L.M.; Meyer, K.; Ray, R. Hepatitis C virus core protein cooperates with ras and transforms primary rat embryo fibroblasts to tumorigenic phenotype. J. Virol. 1996, 70, 4438–4443. [Google Scholar] [PubMed]
- Moriya, K.; Fujie, H.; Shintani, Y.; Yotsuyanagi, H.; Tsutsumi, T.; Ishibashi, K.; Matsuura, Y.; Kimura, S.; Miyamura, T.; Koike, K. The core protein of hepatitis C virus induces hepatocellular carcinoma in transgenic mice. Nat. Med. 1998, 4, 1065–1067. [Google Scholar] [CrossRef]
- Kanda, T.; Steele, R.; Ray, R.; Ray, R.B. Hepatitis C virus core protein augments androgen receptor-mediated signaling. J. Virol. 2008, 82, 11066–11072. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.K.; Majumder, M.; Steele, R.; Meyer, K.; Ray, R.; Ray, R.B. Hepatitis C virus NS5A protein protects against TNF-alpha mediated apoptotic cell death. Virus Res. 2000, 67, 173–178. [Google Scholar] [CrossRef]
- Majumder, M.; Ghosh, A.K.; Steele, R.; Ray, R.; Ray, R.B. Hepatitis C virus NS5A physically associates with p53 and regulates p21/waf1 gene expression in a p53-dependent manner. J. Virol. 2001, 75, 1401–1407. [Google Scholar] [CrossRef] [PubMed]
- Imazeki, F.; Yokosuka, O.; Fukai, K.; Kanda, T.; Kojima, H.; Saisho, H. Prevalence of diabetes mellitus and insulin resistance in patients with chronic hepatitis C: Comparison with hepatitis B virus-infected and hepatitis C virus-cleared patients. Liver Int. 2008, 28, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Neuschwander-Tetri, B.A.; Clark, J.M.; Bass, N.M.; Van Natta, M.L.; Unalp-Arida, A.; Tonascia, J.; Zein, C.O.; Brunt, E.M.; Kleiner, D.E.; McCullough, A.J.; et al. NASH Clinical Research Network. Clinical, laboratory and histological associations in adults with nonalcoholic fatty liver disease. Hepatology 2010, 52, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Haga, Y.; Kanda, T.; Sasaki, R.; Nakamura, M.; Nakamoto, S.; Yokosuka, O. Nonalcoholic fatty liver disease and hepatic cirrhosis: Comparison with viral hepatitis-associated steatosis. World J. Gastroenterol. 2015, 21, 12989–12995. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, T.; Yoshida, T.; Harada, M.; Hisamoto, T.; Nagao, Y.; Ide, T.; Taniguchi, E.; Kumemura, H.; Hanada, S.; Maeyama, M.; et al. Hepatitis C virus down-regulates insulin receptor substrates 1 and 2 through up-regulation of suppressor of cytokine signaling 3. Am. J. Pathol. 2004, 165, 1499–1508. [Google Scholar] [CrossRef]
- Miyamoto, H.; Moriishi, K.; Moriya, K.; Murata, S.; Tanaka, K.; Suzuki, T.; Miyamura, T.; Koike, K.; Matsuura, Y. Involvement of the PA28gamma-dependent pathway in insulin resistance induced by hepatitis C virus core protein. J. Virol. 2007, 81, 1727–1735. [Google Scholar] [CrossRef]
- Banerjee, S.; Saito, K.; Ait-Goughoulte, M.; Meyer, K.; Ray, R.B.; Ray, R. Hepatitis C virus core protein upregulates serine phosphorylation of insulin receptor substrate-1 and impairs the downstream akt/protein kinase B signaling pathway for insulin resistance. J. Virol. 2008, 82, 2606–2612. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Meyer, K.; Mazumdar, B.; Ray, R.B.; Ray, R. Hepatitis C virus differentially modulates activation of forkhead transcription factors and insulin-induced metabolic gene expression. J. Virol. 2010, 84, 5936–5946. [Google Scholar] [CrossRef] [PubMed]
- Bose, S.K.; Shrivastava, S.; Meyer, K.; Ray, R.B.; Ray, R. Hepatitis C virus activates the mTOR/S6K1 signaling pathway in inhibiting IRS-1 function for insulin resistance. J. Virol. 2012, 86, 6315–6322. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Wands, J.R. Insulin receptor substrate 1 overexpression in human hepatocellular carcinoma cells prevents transforming growth factor beta1-induced apoptosis. Cancer Res. 1996, 56, 3391–3394. [Google Scholar] [PubMed]
- Qadri, I.; Choudhury, M.; Rahman, S.M.; Knotts, T.A.; Janssen, R.C.; Schaack, J.; Iwahashi, M.; Puljak, L.; Simon, F.R.; Kilic, G.; et al. Increased phosphoenolpyruvate carboxykinase gene expression and steatosis during hepatitis C virus subgenome replication: Role of nonstructural component 5A and CCAAT/enhancer-binding protein β. J. Biol. Chem. 2012, 287, 37340–37351. [Google Scholar] [CrossRef]
- Parvaiz, F.; Manzoor, S.; Iqbal, J.; McRae, S.; Javed, F.; Ahmed, Q.L.; Waris, G. Hepatitis C virus nonstructural protein 5A favors upregulation of gluconeogenic and lipogenic gene expression leading towards insulin resistance: A metabolic syndrome. Arch. Virol. 2014, 159, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Parvaiz, F.; Manzoor, S.; Iqbal, J.; Sarkar-Dutta, M.; Imran, M.; Waris, G. Hepatitis C virus NS5A promotes insulin resistance through IRS-1 serine phosphorylation and increased gluconeogenesis. World J. Gastroenterol. 2015, 21, 12361–12369. [Google Scholar] [CrossRef]
- Kwon, Y.C.; Bose, S.K.; Steele, R.; Meyer, K.; Di Bisceglie, A.M.; Ray, R.B.; Ray, R. Promotion of Cancer Stem-Like Cell Properties in Hepatitis C Virus-Infected Hepatocytes. J. Virol. 2015, 89, 11549–11556. [Google Scholar] [CrossRef] [Green Version]
- Kwon, Y.C.; Sasaki, R.; Meyer, K.; Ray, R. Hepatitis C Virus Core Protein Modulates Endoglin (CD105) Signaling Pathway for Liver Pathogenesis. J. Virol. 2017, 91, e01235-17. [Google Scholar] [CrossRef]
- Uthaya Kumar, D.B.; Chen, C.L.; Liu, J.C.; Feldman, D.E.; Sher, L.S.; French, S.; DiNorcia, J.; French, S.W.; Naini, B.V.; Junrungsee, S.; et al. TLR4 Signaling via NANOG Cooperates with STAT3 to Activate Twist1 and Promote Formation of Tumor-Initiating Stem-Like Cells in Livers of Mice. Gastroenterology 2016, 150, 707–719. [Google Scholar] [CrossRef]
- Kudo, M. Systemic Therapy for Hepatocellular Carcinoma: Latest Advances. Cancers 2018, 10, 412. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, T.; Honjo, T. PD-1 and PD-1 ligands: From discovery to clinical application. Int. Immunol. 2007, 19, 813–824. [Google Scholar] [CrossRef] [PubMed]
- Iwai, Y.; Ishida, M.; Tanaka, Y.; Okazaki, T.; Honjo, T.; Minato, N. Involvement of PD-L1 on tumor cells in the escape from host immune system and tumor immunotherapy by PD-L1 blockade. Proc. Natl. Acad. Sci. USA 2002, 99, 12293–12297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leach, D.R.; Krummel, M.F.; Allison, J.P. Enhancement of antitumor immunity by CTLA-4 blockade. Science 1996, 271, 1734–1736. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Name | Pathways | References |
|---|---|---|---|
| TERT | Telomerase reverse transcriptase | TERT | [52,53,54,55,56] |
| CTNNB1 | Catenin β1 | β-catenin | [52,53] |
| TP53 | Tumor protein p53 | p53–RB | [52,53] |
| ARID2 | AT-rich interaction domain 2 | Chromatin remodeling | [52] |
| AXIN1 | Axin 1 | β-catenin | [52] |
| TSC2 | TSC complex subunit 2 | PI3K–mTOR | [52] |
| RB1 | Retinoblastoma 1 | p53–RB | [52] |
| ACVR2A | Activin A receptor type 2A | SMAD | [52] |
| BRD7 | Bromodomain containing 7 | Chromatin remodeling | [52] |
| CDKN1A | Cyclin-dependent kinase inhibitor 1A | β-catenin | [52] |
| MEN1 | Menin 1 | (MEN1 syndrome) | [52] |
| GALN11 | Polypeptide N-acetylgalactosaminyltransferase 11 | NOTCH | [52] |
| FGF19 | Fibroblast growth factor 19 | β-catenin | [52] |
| CCND1 | Cyclin D1 | p53–RB | [52] |
| ARID1A | AT-rich interaction domain 1A | Chromatin remodeling | [52,53,54,57,58] |
| CDKN2A | Cyclin-dependent kinase inhibitor 2A | p53–RB | [52] |
| CDKN2B | Cyclin-dependent kinase inhibitor 2B | p53–RB | [52] |
| RPS6KA3 | Ribosomal protein S6 kinase, 90kDa, polypeptide 3 | p53–RB | [52] |
| NFE2L2 | Nuclear factor, erythroid 2-like 2 | NRF2–KEAP1 | [52] |
| NCOR1 | Nuclear receptor corepressor 1 | β-catenin/chromatin remodeling | [52] |
| ADH1B | Alcohol dehydrogenase 1B, β polypeptide | [52] | |
| SRCAP | Snf2-related CREB binding protein activator protein | Chromatin remodeling | [52] |
| FCRL1 | Fc receptor like 1 | [52] | |
| PTEN | Phosphatase and tensin homolog | PI3K–mTOR | [52] |
| HNRNPA2B1 | Heterogeneous nuclear ribonucleoprotein A2/B1 | MAPK | [52] |
| CYP2E1 | Cytochrome P450 family 2 subfamily E member 1 | [52] | |
| MAP2K3 | Mitogen-activated protein kinase 3 | MAPK | [52] |
| TSC1 | Tuberous sclerosis 1 | mTOR | [52] |
| TMEM99 | Transmembrane protein 99 | [52] | |
| G6PC | Glucose-6-phosphatase, catalytic subunit | FoxO | [52] |
| miRs | Upregulation/Downregulation | Target Genes | References |
|---|---|---|---|
| Hepatic miRs | |||
| miR-223 | Upregulation | Stathmin 1 (STMN1) | [92] |
| miR-143 | Upregulation | Fibronectin type III domain containing 3B (FNDC3B) | [93] |
| miR-602 | Upregulation | Ras-association domain family member 1 (RASSF1A) | [94] |
| miR-224 | Upregulation | N/A | [98,110,116] |
| miR-22 | Upregulation in male HCC- adjacent tissue | Estrogen receptor alpha (ERα) | [99] |
| miR-96 | Upregulation | N/A | [112] |
| miR-183 | Upregulation | N/A | [112] |
| miR-196a | Upregulation | N/A | [112] |
| miR-545/374a | Upregulation | Estrogen-related receptor gamma (ESRRG) | [120] |
| miR-331-3p | Upregulation | Inhibitor of growth family member 5 (ING5) | [124] |
| miR-519a | Upregulation | Forkhead box F2 (FOXF2) | [124] |
| miR-106b | Upregulation | N/A | [131] |
| miR-1269b | Upregulation | Cell division cycle 40 homolog (CDC40) | [132] |
| let-7a | Downregulation | Signal transducer and activator of transcription 3 (STAT3) | [95] |
| miR-152 | Downregulation | DNA methyltransferase 1 (DNMT1), TNFRF6B | [96,115] |
| miR-145 | Downregulation | Cullin 5 (CUL5) | [98,125] |
| miR-199b | Downregulation | N/A | [98] |
| miR-29c | Downregulation | TNF-α-induced protein 3 (TNFAIP3) | [100] |
| miR-92 | Downregulation | N/A | [102] |
| miR-338-3p | Downregulation | 3′ UTR region of CCND1 | [104] |
| miR-34a | Downregulation | C-C motif chemokine ligand 22 (CCL22) | [115] |
| miR-101 | Downregulation | DNA methyltransferase (DNMT)3A | [116] |
| miR-122 | Downregulation | Pituitary tumor-transforming gene 1 (PTTG1) binding factor (PBF) | [117,134,139] |
| miR-148a | Downregulation | Hematopoietic pre-B cell leukemia transcription factor-interacting protein (HPIP) | [108] |
| miR-22 | Downregulation | CDKN1A | [109,134] |
| let-7c | Downregulation | N/A | [112] |
| miR-138 | Downregulation | N/A | [112] |
| miR-205 | Downregulation | N/A | [113] |
| miR-101-3p | Downregulation | RAB GTPase 5A (RAB5A) | [114] |
| miR-429 | Downregulation | NOTCH1 | [121] |
| miR-216 | Downregulation | Insulin-like growth factor 2 mRNA-binding protein 2 (IGF2BP2) | [122] |
| miR-34c | Downregulation | Transforming growth factor-β-induced factor homeobox 2 (TGIF2) | [127] |
| miR-18a | Downregulation | Connective tissue growth factor (CTGF) | [133] |
| miR-30b-5p | Downregulation | DNMT3α, USP37 | [135] |
| miR-384 | Downregulation | Pleiotrophin (PTN) | [136] |
| miR-125a-5p | Downregulation | erb-b2 receptor tyrosine kinase 3 (ERBB3) | [137] |
| miR-26a | Downregulation | N/A | [140] |
| miR-302c-3p | Downregulation | TNF receptor-associated factor 4 (TRAF4) | [141] |
| miR-1271 | Downregulation | CCNA1 | [142] |
| Serum miRs | |||
| miR-25 | Upregulation | N/A | [97] |
| miR-375 | Upregulation | N/A | [97] |
| let-7f | Upregulation | N/A | [97] |
| miR-122 | Upregulation | N/A | [99] |
| miR-18a | Upregulation | N/A | [103] |
| miR-101 | Upregulation | N/A | [111] |
| miR-24-3p | Upregulation | N/A | [117] |
| miR-545/374a | Upregulation | ERR gamma (ERRG) | [120] |
| miR-96 | Upregulation | N/A | [129] |
| miR-21 | Downregulation | N/A | [118] |
| miR-222 | Downregulation | N/A | [118] |
| miR-29 | Downregulation | N/A | [119] |
| miR-150 | Downregulation | N/A | [123] |
| miR-126 | Downregulation | N/A | [128] |
| miR-125b | Downregulation | N/A | [130] |
| miR-143/145 | Downregulation | N/A | [138] |
| miRs | Upregulation/Downregulation | Target Genes | References |
|---|---|---|---|
| Hepatic miRs | |||
| miR-193b | Upregulation | STMN1 | [179] |
| miR-155 | Upregulation | N/A | [180] |
| miR-122 | Upregulation | Cyclin G1 | [181,182,186] |
| miR-373 | Upregulation | Wee1 | [195] |
| miR-24 | Downregulation | N/A | [183] |
| miR-27a | Downregulation | N/A | [183] |
| miR-198 | Downregulation | N/A | [184] |
| miR-152 | Downregulation | N/A | [186] |
| miR-181c | Downregulation | Homeobox A1 | [187] |
| miR-431 | Downregulation | N/A | [189] |
| miR-138 | Downregulation | TERT | [192] |
| Serum miRs | |||
| miR-150 | Upregulation | N/A | [191] |
| miR-221 | Upregulation | N/A | [193] |
| miR-101-1 | Upregulation | N/A | [193] |
| miR-27a | Upregulation | N/A | [194] |
| miR-221 | Downregulation | Suppressor of cytokine signaling (SOCS)1 and SOCS3 | [188] |
| miR-16 | Downregulation | N/A | [190] |
| miR-34a | Downregulation | Heat-shock protein 70 | [194] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanda, T.; Goto, T.; Hirotsu, Y.; Moriyama, M.; Omata, M. Molecular Mechanisms Driving Progression of Liver Cirrhosis towards Hepatocellular Carcinoma in Chronic Hepatitis B and C Infections: A Review. Int. J. Mol. Sci. 2019, 20, 1358. https://doi.org/10.3390/ijms20061358
Kanda T, Goto T, Hirotsu Y, Moriyama M, Omata M. Molecular Mechanisms Driving Progression of Liver Cirrhosis towards Hepatocellular Carcinoma in Chronic Hepatitis B and C Infections: A Review. International Journal of Molecular Sciences. 2019; 20(6):1358. https://doi.org/10.3390/ijms20061358
Chicago/Turabian StyleKanda, Tatsuo, Taichiro Goto, Yosuke Hirotsu, Mitsuhiko Moriyama, and Masao Omata. 2019. "Molecular Mechanisms Driving Progression of Liver Cirrhosis towards Hepatocellular Carcinoma in Chronic Hepatitis B and C Infections: A Review" International Journal of Molecular Sciences 20, no. 6: 1358. https://doi.org/10.3390/ijms20061358