Metabolic and Immunological Shifts during Mid-to-Late Gestation Influence Maternal Blood Methylation of CPT1A and SREBF1

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Pregnancy-Induced Adiposity Trait
2.2. Mean Methylation Levels of Variably Methylated Sites in CPT1A and SREBF1 during Pregnancy
2.3. Confirmative Study of the Association between Adiposity Phenotype and Methylation Levels
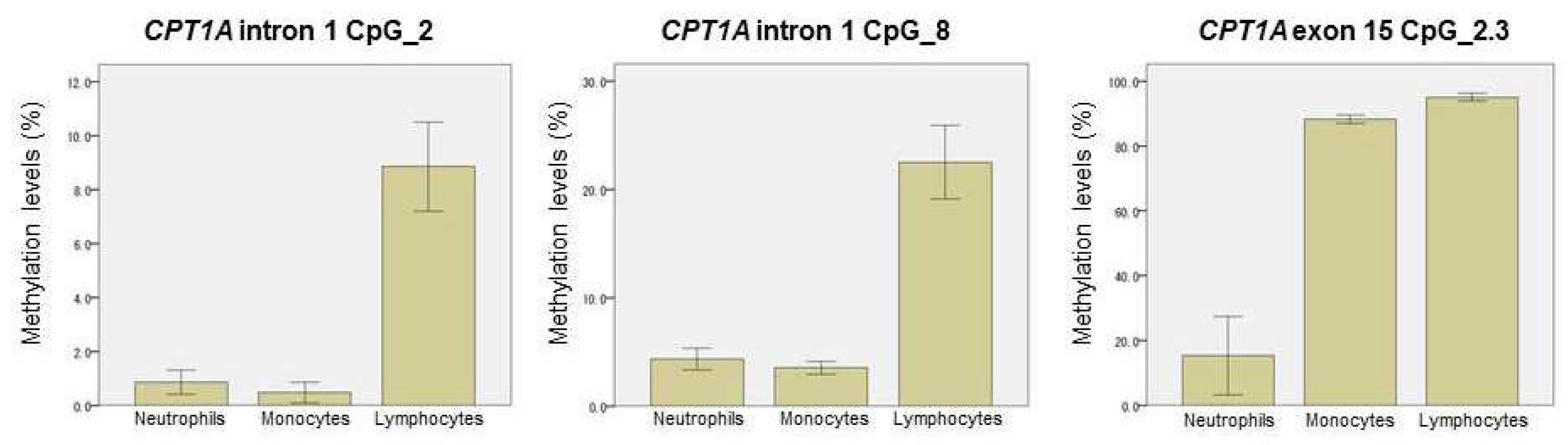
2.4. Methylation-Based Estimation of Cell Type Composition
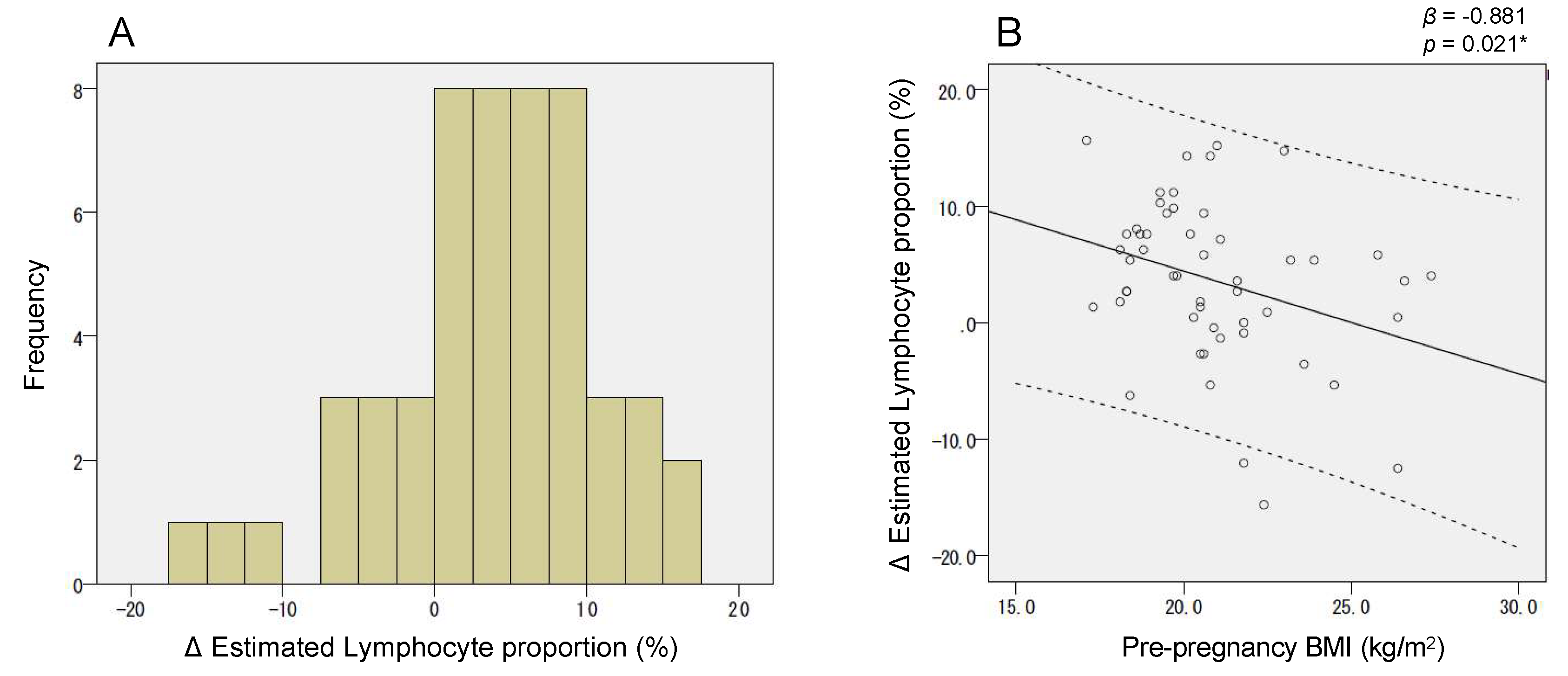
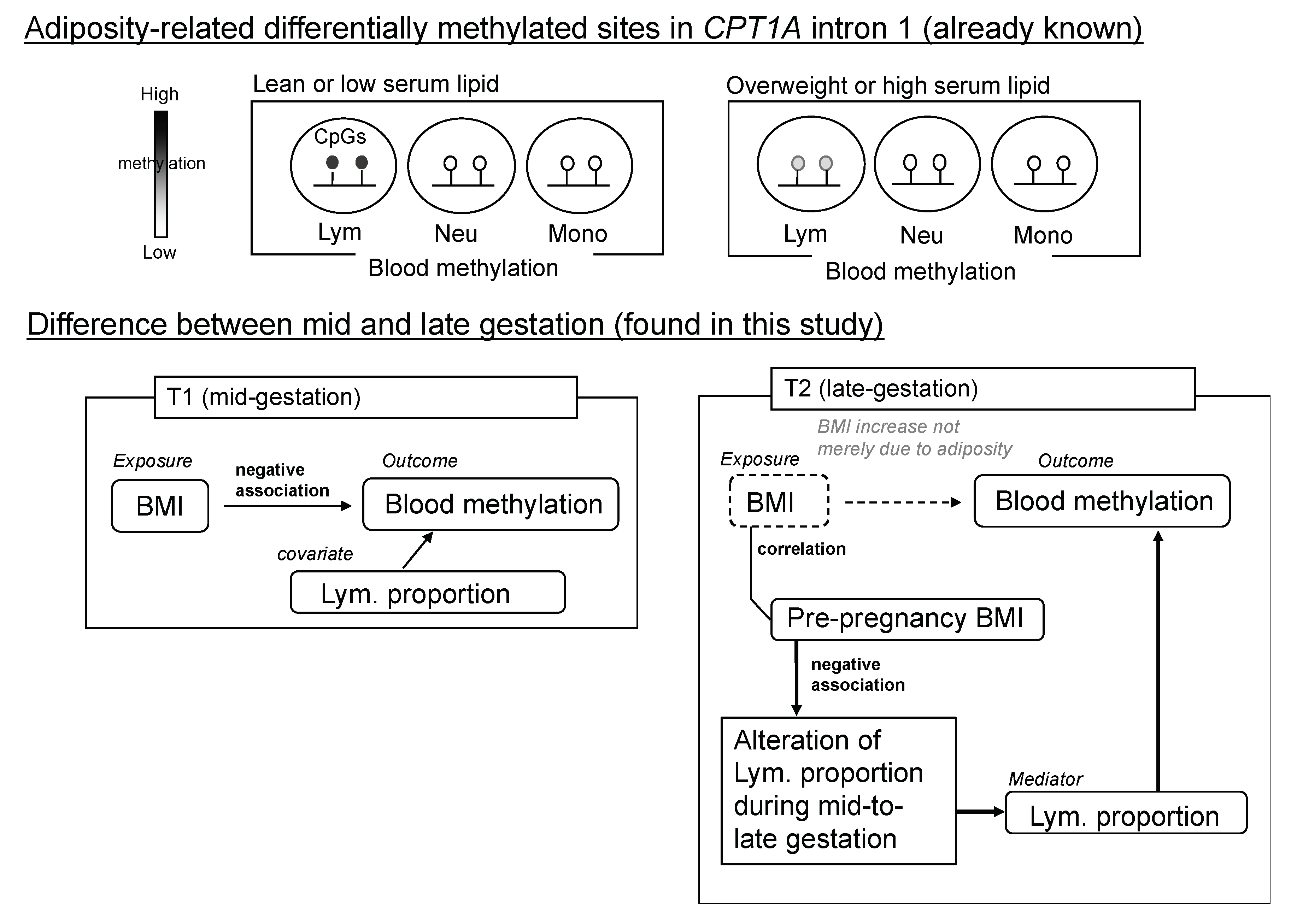
2.5. Dependency of the Lymphocyte Proportion Change on Pre-Pregnancy BMI Influenced the Association between CPT1A Intron1 Methylation and BMI at Late Gestation (T2)
3. Discussion
4. Materials and Methods
4.1. Study Population and Design
4.2. Maternal Blood Sample Collection and Serum LDL-C Quantification
4.3. Collection of Control Blood Samples for the Cell Fractionation and Cell Counts
4.4. Methylation Analysis Using EpiTYPER
4.5. Construction of a Standard Curve for Specific Cell Type Proportion vs. Marker Site Methylation
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CpG | 5′-Cytosine-phosphate-guanine-3′ |
| EWAS | Epigenome-wide association study |
| CPT1A | Carnitine palmitoyltransferase 1A |
| SREBF1 | Sterol regulatory element-binding transcription factor 1 |
| STK24 | Serine/threonine kinase 24 |
| BMI | Body mass index |
| LDL-C | Low-density lipoprotein cholesterol |
| BC-GENIST | Birth Cohort Gene and ENvironment Interaction Study of TMDU |
References
- Nelson, S.M.; Matthews, P.; Poston, L. Maternal Metabolism and Obesity: Modifiable Determinants of Pregnancy Outcome. Hum. Reprod. Updat. 2010, 16, 255–275. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Liu, F.; Li, S. Metabolic Adaptations in Pregnancy: A Review. Ann. Nutr. Metab. 2017, 70, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Farias, D.; Franco-sena, A.B.; Vilela, A.A.F.; Lepsch, J.; Mendes, R.H.; Kac, G. Lipid Changes throughout Pregnancy According to Pre-Pregnancy BMI: Results from a Prospective Cohort. Epidemiology 2016, 123, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Ahlsson, F.; Diderholm, B.; Ewald, U.; Jonsson, B.; Forslund, A.; Stridsberg, M.; Gustafsson, J. Adipokines and Their Relation to Maternal Energy Substrate Production, Insulin Resistance and Fetal Size. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 168, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Pugh, S.J.; Albert, P.S.; Kim, S.; Grobman, W.; Hinkle, S.N.; Newman, R.B.; Wing, D.A.; Katherine, L. Patterns of Gestational Weight Gain and Birth Weight Outcomes in the NICHD Fetal Growth Study—Singletons: A Prospective Study. Am. J. Obs. Gynecol. 2017, 217, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Stubert, J.; Reister, F.; Hartmann, S.; Janni, W. The Risks Associated with Obesity in Pregnancy. Dtsch. Arztebl. Int. 2018, 115, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Mendelson, M.M.; Marioni, R.E.; Joehanes, R.; Liu, C.; Hedman, Å.K.; Aslibekyan, S.; Demerath, E.W.; Guan, W.; Zhi, D.; Yao, C.; et al. Association of Body Mass Index with DNA Methylation and Gene Expression in Blood Cells and Relations to Cardiometabolic Disease: A Mendelian Randomization Approach. PLoS Med. 2017, 14, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Das, M.; Sha, J.; Hidalgo, B.; Aslibekyan, S.; Do, A.N.; Zhi, D.; Sun, D.; Zhang, T.; Li, S.; Chen, W.; et al. Association of DNA Methylation at CPT1A Locus with Metabolic Syndrome in the Genetics of Lipid Lowering Drugs and Diet Network (GOLDN) Study. PLoS ONE 2016, 11, e0145789. [Google Scholar] [CrossRef] [PubMed]
- Demerath, E.W.; Guan, W.; Grove, M.L.; Aslibekyan, S.; Mendelson, M.; Zhou, Y.; Hedman, Å.K.; Johanna, K.; Li, L.; Irvin, M.R.; et al. Epigenome-Wide Association Study (EWAS) of BMI, BMI Change and Waist Circumference in African American Adults Identifies Multiple Replicated Loci. Hum. Mol. Genet. 2015, 24, 4464–4479. [Google Scholar] [CrossRef] [PubMed]
- Wahl, S.; Drong, A.; Lehne, B.; Loh, M.; Scott, W.R.; Kunze, S.; Tsai, P.C.; Ried, J.S.; Zhang, W.; Yang, Y.; et al. Epigenome-Wide Association Study of Body Mass Index, and the Adverse Outcomes of Adiposity. Nature 2017, 541, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Aslibekyan, S.; Demerath, E.W.; Mendelson, M.; Zhi, D.; Guan, W.; Liang, L.; Sha, J.; Pankow, J.S.; Liu, C.; Irvin, M.R.; et al. Epigenome-Wide Study Identifies Novel Methylation Loci Associated with Body Mass Index and Waist Circumference. Obesity 2015, 23, 1493–1501. [Google Scholar] [CrossRef] [PubMed]
- Meeks, K.A.C.; Henneman, P.; Venema, A.; Burr, T.; Galbete, C.; Danquah, I.; Schulze, M.B.; Mockenhaupt, F.P.; Owusu-Dabo, E.; Rotimi, C.N.; et al. An Epigenome-Wide Association Study in Whole Blood of Measures of Adiposity among Ghanaians: The RODAM Study. Clin. Epigenet. 2017, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, K.F.; van Iterson, M.; Slieker, R.C.; Moed, M.H.; Bonder, M.J.; van Galen, M.; Mei, H.; Zhernakova, D.V.; van den Berg, L.H.; Deelen, J.; et al. Blood Lipids Influence DNA Methylation in Circulating Cells. Genome Biol. 2016, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Truong, V.; Huang, S.; Dennis, J.; Lemire, M.; Zwingerman, N.; Aïssi, D.; Kassam, I.; Perret, C.; Wells, P.; Morange, P.E.; et al. Blood Triglyceride Levels Are Associated with DNA Methylation at the Serine Metabolism Gene PHGDH. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, L.; Wahl, S.; Pilling, L.C.; Reischl, E.; Sandling, J.K.; Kunze, S.; Holdt, L.M.; Kretschmer, A.; Schramm, K.; Adamski, J.; et al. DNA Methylation of Lipid-Related Genes Affects Blood Lipid Levels. Circ. Cardiovasc. Genet. 2015, 8, 334–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irvin, M.R.; Zhi, D.; Joehanes, R.; Mendelson, M.; Aslibekyan, S.; Claas, S.A.; Thibeault, K.S.; Patel, N.; Day, K.; Jones, L.W.; et al. Epigenome-Wide Association Study of Fasting Blood Lipids in the Genetics of Lipid Lowering Drugs and Diet Network Study. Circulation 2014, 130, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Frazier-Wood, A.C.; Aslibekyan, S.; Absher, D.M.; Hopkins, P.N.; Sha, J.; Tsai, M.Y.; Tiwari, H.K.; Waite, L.L.; Zhi, D.; Arnett, D.K. Methylation at CPT1A Locus Is Associated with Lipoprotein Subfraction Profiles. J. Lipid Res. 2014, 55, 1324–1330. [Google Scholar] [CrossRef] [PubMed]
- Sayols-Baixeras, S.; Subirana, I.; Civeira, F.; Roquer, J.; Soriano-t, C.; Jime, J.; Do, A.N.; Absher, D.; Cenarro, A.; Mu, D.; et al. Identification and Validation of Seven New Loci Showing Differential DNA Methylation Related to Serum Lipid Profile: An Epigenome-Wide Approach. The REGICOR Study. Hum. Mol. Genet. 2016, 25, 4556–4565. [Google Scholar] [CrossRef] [PubMed]
- Hedman, Å.K.; Mendelson, M.M.; Marioni, R.E.; Gustafsson, S.; Joehanes, R.; Irvin, M.R.; Zhi, D.; Sandling, J.K.; Yao, C.; Liu, C.; et al. Epigenetic Patterns in Blood Associated with Lipid Traits Predict Incident Coronary Heart Disease Events and Are Enriched for Results from Genome-Wide Association Studies. Circ. Cardiovasc. Genet. 2017, 10, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, F.; Aïssi, D.; Carrié, A.; Morange, P. Robust Validation of Methylation Levels Association at CPT1A Locus with Lipid Plasma Levels 1. J. Lipid Res. 2014, 55, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- McGarry, J.D.; Brown, N.F. The mitochondrial carnitine palmitoyltransferase system. From concept to molecular analysis. Eur. J. Biochem. 1997, 244, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Shimano, H.; Sato, R. SREBP-Regulated Lipid Metabolism: Convergent Physiology—Divergent Pathophysiology. Nat. Publ. Gr. 2017, 13, 710–730. [Google Scholar] [CrossRef] [PubMed]
- Tobi, E.W.; Goeman, J.J.; Monajemi, R.; Gu, H.; Putter, H.; Zhang, Y.; Slieker, R.C.; Stok, A.P.; Thijssen, P.E.; Müller, F.; et al. DNA Methylation Signatures Link Prenatal Famine Exposure to Growth and Metabolism. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Pitkin, R.M. Nutritional support in obstetrics and gynecology. Clin. Obstet. Gynecol. 1976, 3, 489–513. [Google Scholar] [CrossRef]
- Ecker, S.; Chen, L.; Pancaldi, V.; Bagger, F.O.; Fernández, J.M.; Carrillo de Santa Pau, E.; Juan, D.; Mann, A.L.; Watt, S.; Casale, F.P.; et al. Genome-Wide Analysis of Differential Transcriptional and Epigenetic Variability across Human Immune Cell Types. Genome Biol. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Luppi, P.; Haluszczak, C.; Betters, D.; Richard, C.A.H.; Trucco, M.; Deloia, J.A. Monocytes Are Progressively Activated in the Circulation of Pregnant Women. J. Leukoc. Biol. 2002, 72, 874–884. [Google Scholar] [PubMed]
- Keung, W.; Ussher, J.R.; Jaswal, J.S.; Raubenheimer, M.; Lam, V.H.M.; Wagg, C.S.; Lopaschuk, G.D. Inhibition of Carnitine Palmitoyltransferase-1 Activity Alleviates Insulin Resistance in Diet-Induced Obese Mice. Diabetes 2013, 62, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Strable, M.S.J.M.N. Genetic Control of de Novo Lipogenesis: Role in Diet-Induced Obesity. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 199–214. [Google Scholar] [CrossRef] [PubMed]
- Herrera, E. Metabolic Adaptations in Pregnancy and Their Implications for Availability of Substrates to the Fetus. Eur. J. Clin. Nutr. 2000, 54, S47–S51. [Google Scholar] [CrossRef] [PubMed]
- Aghaeepour, N.; Ganio, E.A.; Mcilwain, D.; Tsai, A.S.; Tingle, M.; Van Gassen, S.; Gaudilliere, D.K.; Baca, Q.; McNeil, L.; Okada, R.; et al. An immune clock of human pregnancy. Sci. Immunol. 2017, 2, eaan2946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takimoto, H.; Mitsuishi, C.; Kato, N. Attitudes toward Pregnancy Related Changes and Self-Judged Dieting Behavior. Asia Pac. J. Clin. Nutr. 2011, 20, 212–219. [Google Scholar] [PubMed]
- Harald Theml, M.D.; Heinz Diem, M.D.; Torsten Haferlach, M.D. Practical Microscopic and Clinical Diagnosis, 2nd ed.; Thieme Verlag: Stuttgart, Germany, 2004. [Google Scholar]
- Suchiman, H.E.D.; Slieker, R.C.; Kremer, D.; Slagboom, P.E.; Heijmans, B.T.; Tobi, E.W. Design, Measurement and Processing of Region-Specific DNA Methylation Assays: The Mass Spectrometry-Based Method EpiTYPER. Front. Genet. 2015, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, A.E.; Irizarry, R.A. Accounting for Cellular Heterogeneity Is Critical in Epigenome-Wide Association Studies. Genome Biol. 2014, 15, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Salas, L.A.; Koestler, D.C.; Butler, R.A.; Hansen, H.M.; Wiencke, J.K.; Kelsey, K.T.; Christensen, B.C. An Optimized Library for Reference-Based Deconvolution of Whole-Blood Biospecimens Assayed Using the Illumina HumanMethylationEPIC BeadArray. Genome Biol. 2018, 19, 1–14. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Genomic Context | EpiTYPER CpG Unit | Mean Methylation (%) (SD) | p-Value * | |
|---|---|---|---|---|---|
| T1 | T2 | ||||
| CPT1A | Intron 1 | CpG_2 a | 1.9 (0.7) | 1.9 (0.6) | 8.2 × 10−1 |
| CpG_3 | 8.9 (1.3) | 9.1 (1.0) | 2.4 × 10−1 | ||
| CpG_4 | 4.5 (1.1) | 4.7 (0.9) | 7.3 × 10−2 | ||
| CpG_6 | 11.9 (3.8) | 12.5 (4.0) | 2.8 × 10−1 | ||
| CpG_7 | 4.8 (1.3) | 4.9 (1.1) | 3.6 × 10−1 | ||
| CpG_8 b | 6.5 (1.5) | 6.8 (1.3) | 1.4 × 10−1 | ||
| SREBF1 | Intron 1 | CpG_1 | 46.6 (2.8) | 47.2 (2.9) | 5.0 × 10−3 |
| CpG_3 | 36.9 (4.0) | 36.7 (4.2) | 7.5 × 10−1 | ||
| CpG_4 c | 71.3 (2.5) | 72.7 (2.3) | 1.6 × 10−7 | ||
| CPT1A | Exon 15 | CpG_2.3 d | 26.0 (13.3) | 29.1 (13.0) | 2.9 × 10−5 |
| CpG_5.6 | 48.5 (10.5) | 52.0 (10.1) | 2.3 × 10−6 | ||
| CpG_8.9 | 73.5 (5.7) | 75.2 (5.7) | 3.0 × 10−4 | ||
| CpG_10 | 38.7 (10.3) | 41.7 (9.9) | 3.3 × 10−5 | ||
| CpG_12 | 19.5 (5.8) | 21.9 (5.6) | 6.4 × 10−4 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavethynath, S.; Imai, C.; Jin, X.; Hichiwa, N.; Takimoto, H.; Okamitsu, M.; Tarui, I.; Aoyama, T.; Yago, S.; Fudono, A.; et al. Metabolic and Immunological Shifts during Mid-to-Late Gestation Influence Maternal Blood Methylation of CPT1A and SREBF1. Int. J. Mol. Sci. 2019, 20, 1066. https://doi.org/10.3390/ijms20051066
Pavethynath S, Imai C, Jin X, Hichiwa N, Takimoto H, Okamitsu M, Tarui I, Aoyama T, Yago S, Fudono A, et al. Metabolic and Immunological Shifts during Mid-to-Late Gestation Influence Maternal Blood Methylation of CPT1A and SREBF1. International Journal of Molecular Sciences. 2019; 20(5):1066. https://doi.org/10.3390/ijms20051066
Chicago/Turabian StylePavethynath, Shilpa, Chihiro Imai, Xin Jin, Naomi Hichiwa, Hidemi Takimoto, Motoko Okamitsu, Iori Tarui, Tomoko Aoyama, Satoshi Yago, Ayako Fudono, and et al. 2019. "Metabolic and Immunological Shifts during Mid-to-Late Gestation Influence Maternal Blood Methylation of CPT1A and SREBF1" International Journal of Molecular Sciences 20, no. 5: 1066. https://doi.org/10.3390/ijms20051066