Investigation of the AQP Family in Soybean and the Promoter Activity of TIP2;6 in Heat Stress and Hormone Responses

Abstract
:1. Introduction
2. Results
2.1. Identification of the Soybean AQP Family
2.2. Key Structural Features of the AQP Proteins
2.3. Chromosome Distribution of the AQP Genes
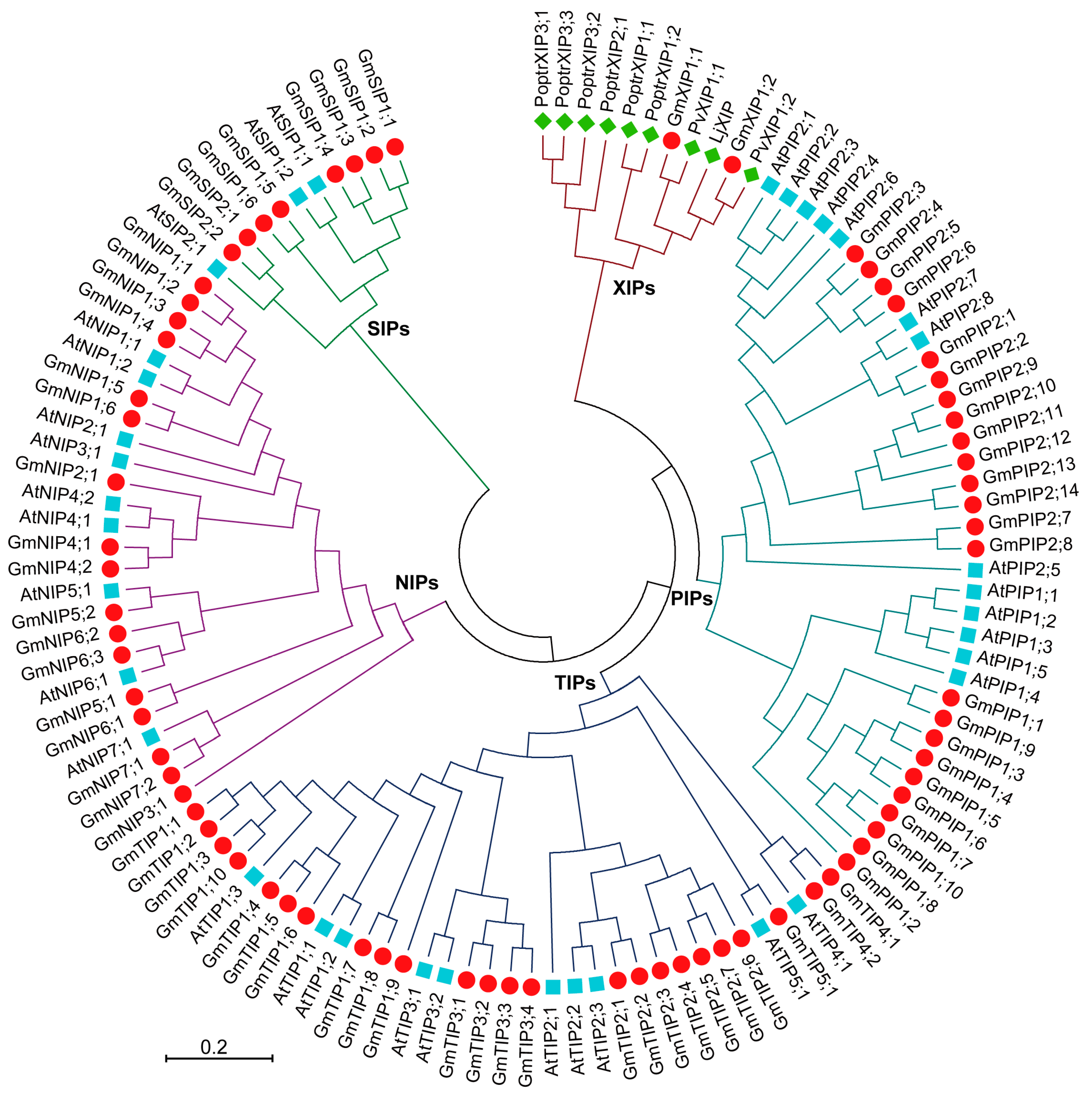
2.4. Evolutionary Characterization of the AQP Genes
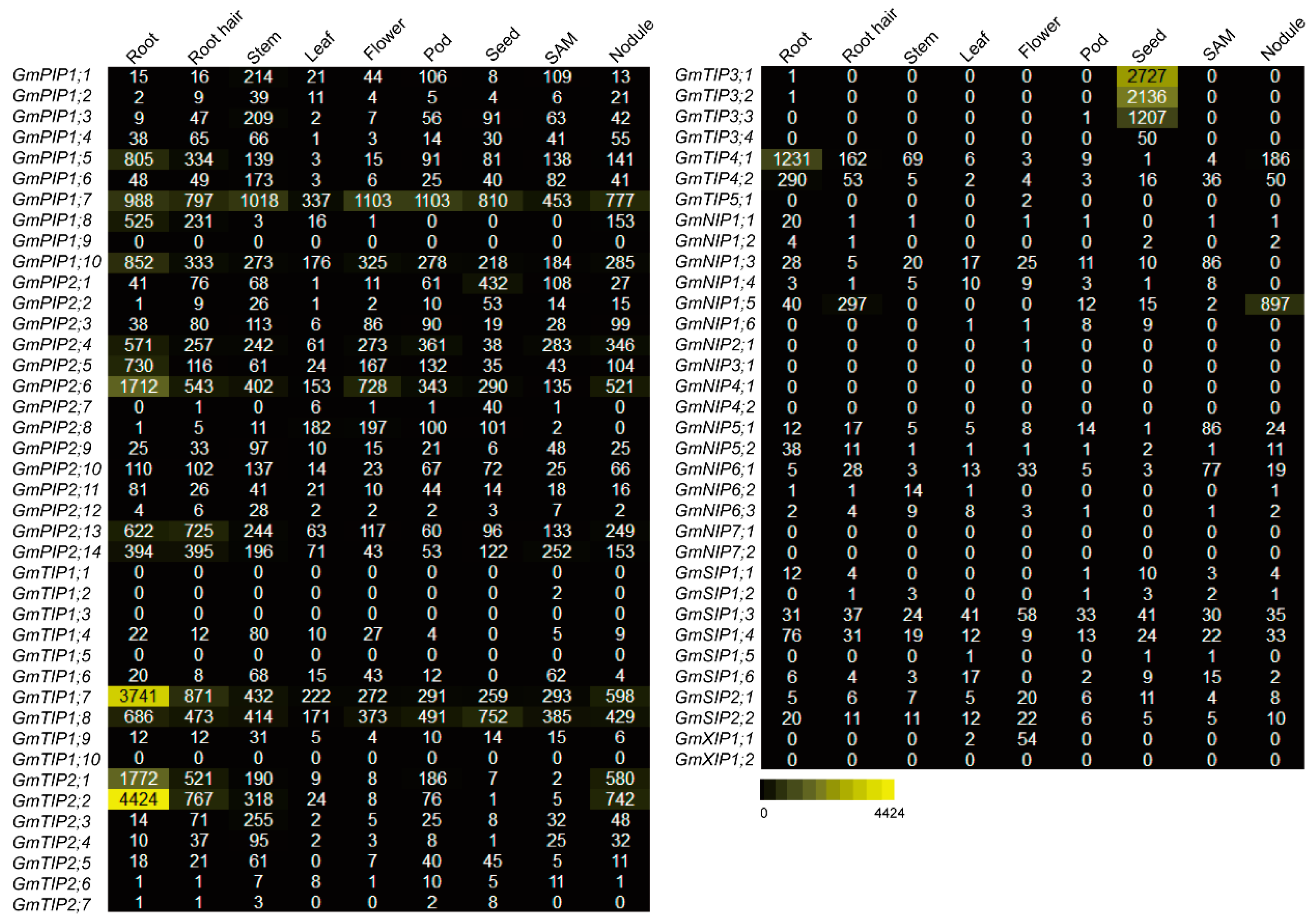
2.5. Expression Profiles of the AQP Genes in Different Tissues
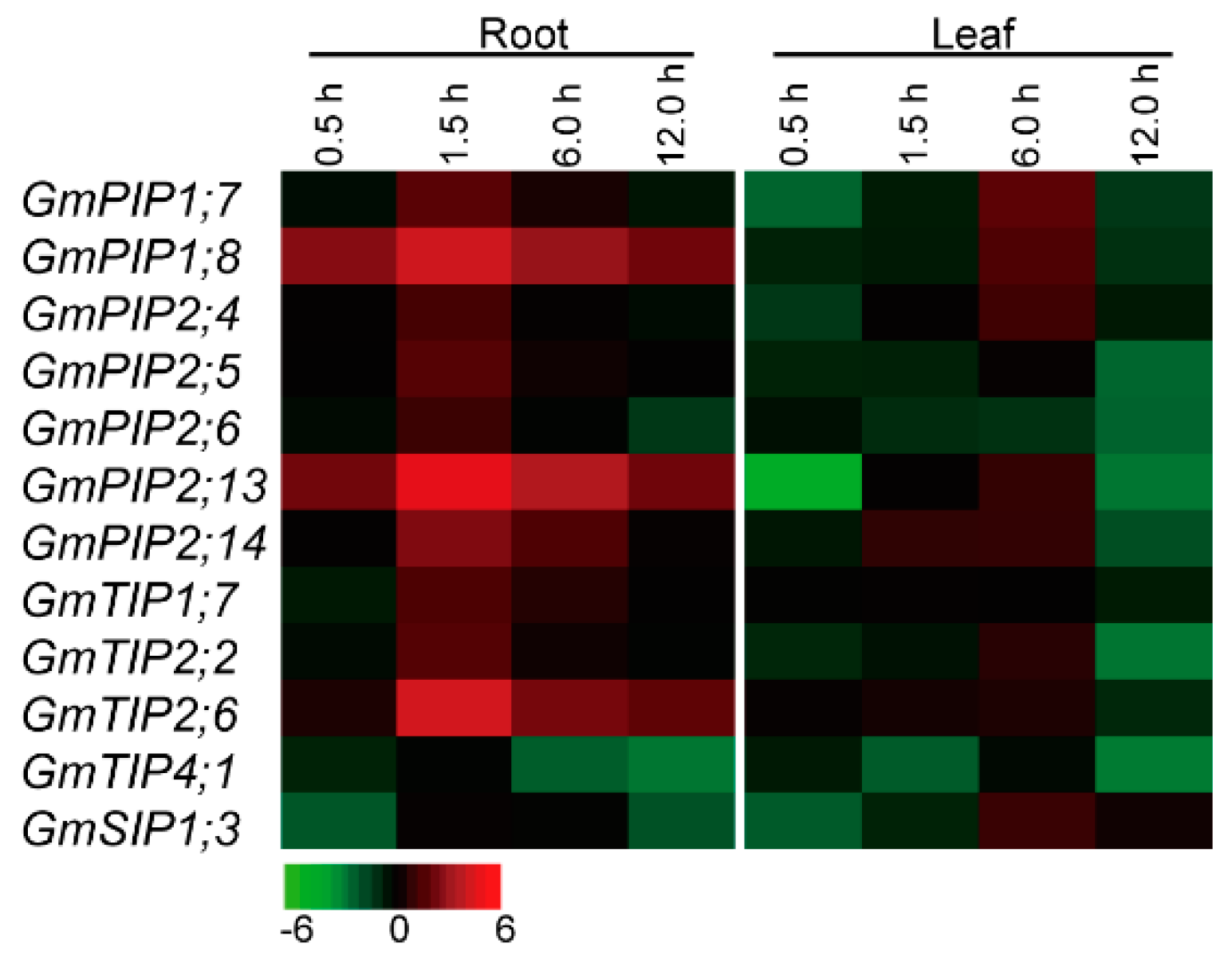
2.6. Expression Profiles of the Candidate AQP Genes in Response to Heat Stress
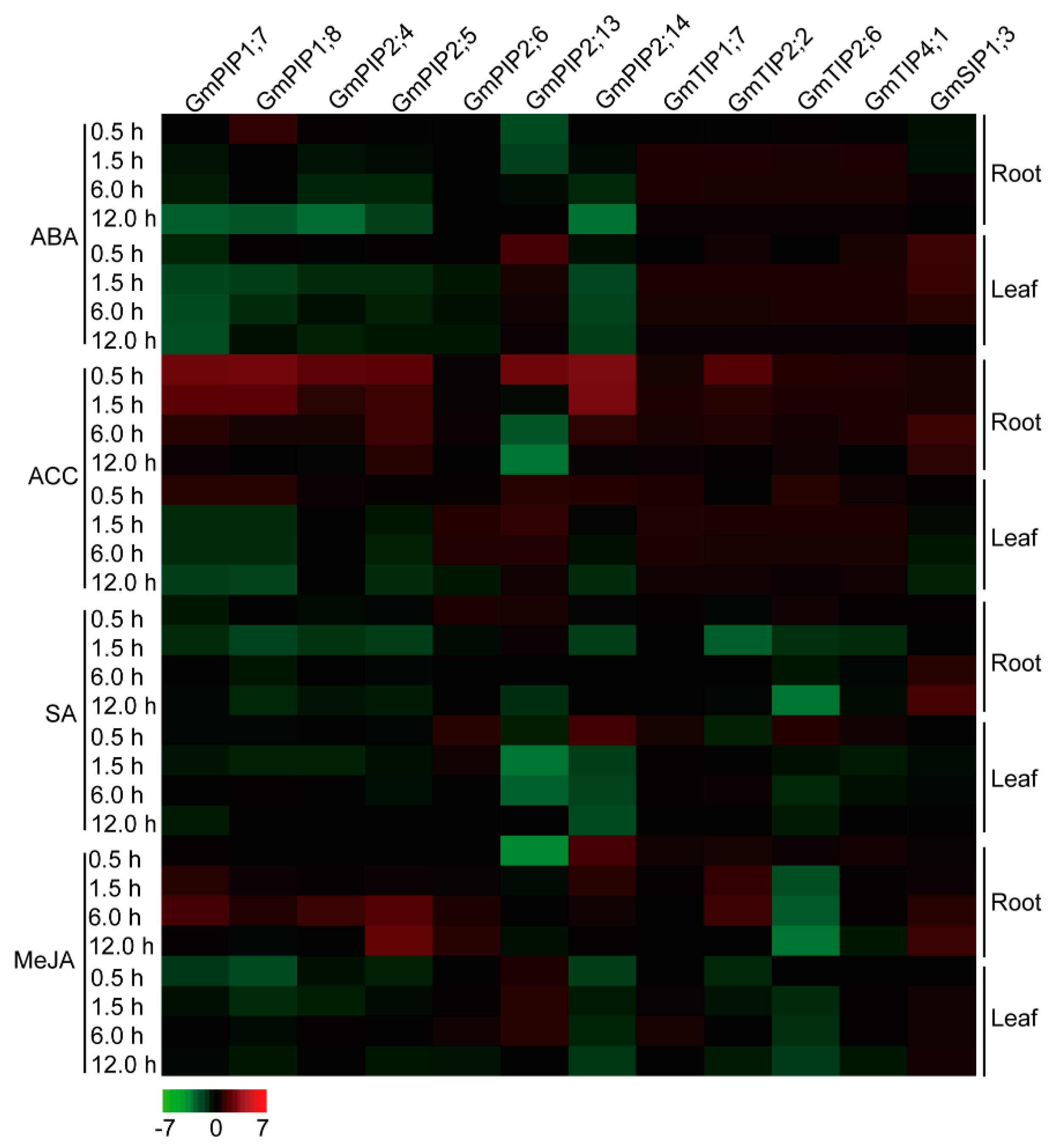
2.7. Expression Profiles of the Candidate AQP Genes in Response to ABA, ACC, SA, and MeJA Signals
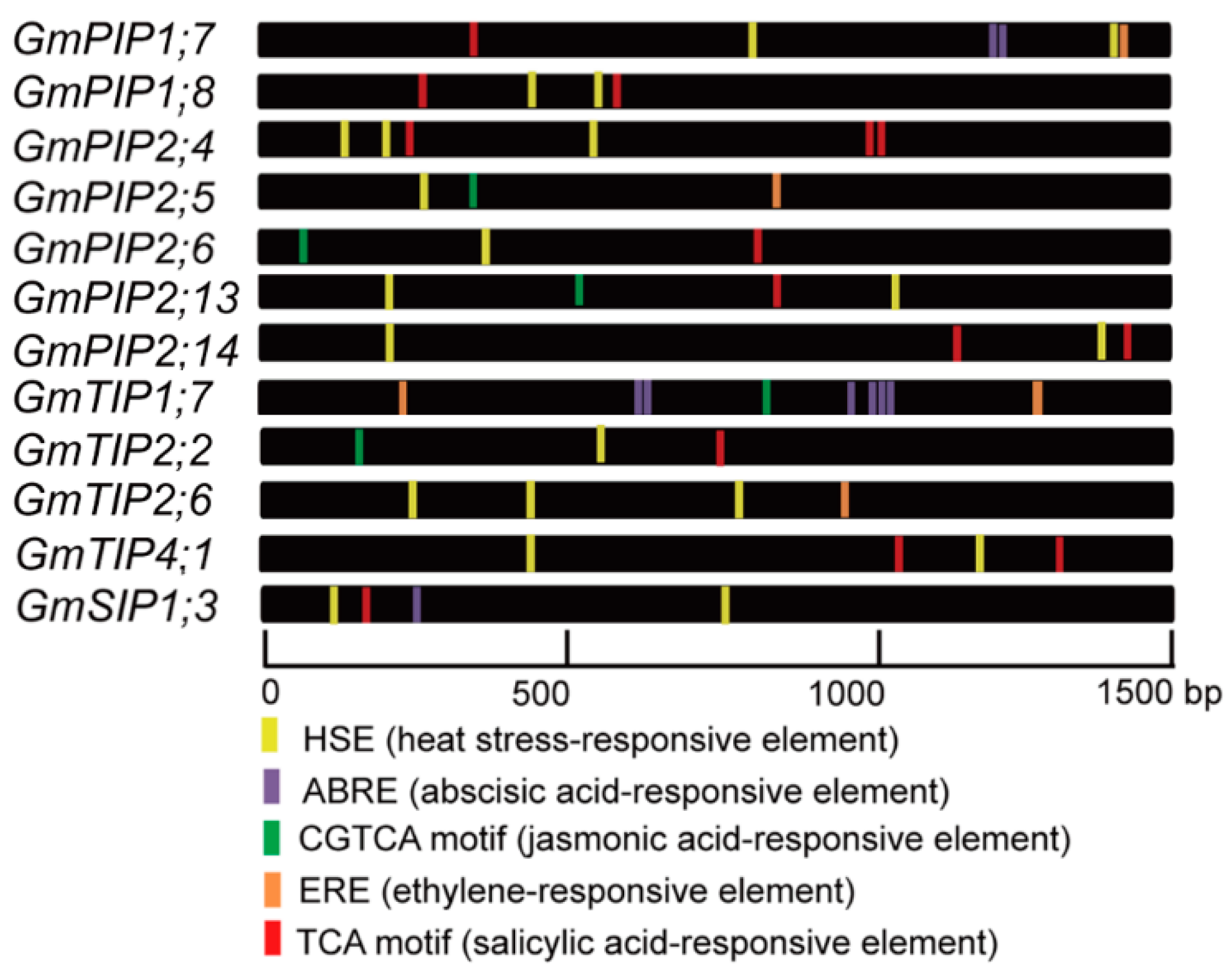
2.8. Promoter Regulatory Elements of the Candidate AQP Genes
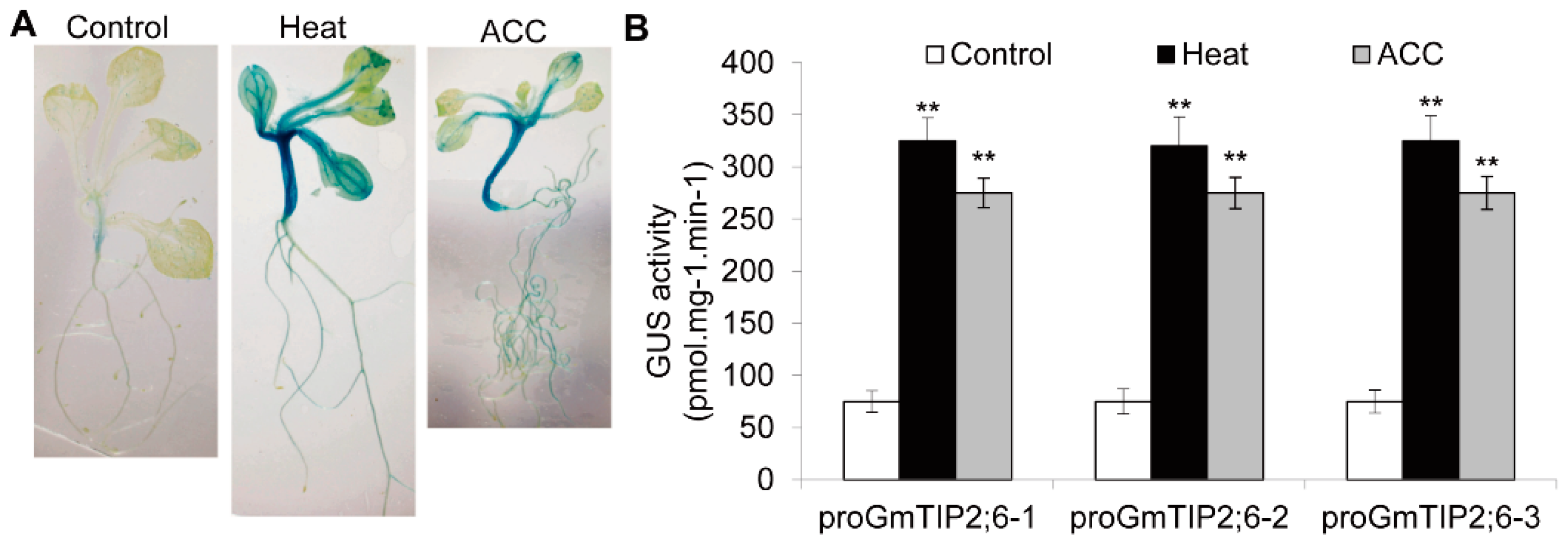
2.9. GUS Activity of the GmTIP2;6 Promoter
3. Discussion
4. Materials and Methods
4.1. Categorization of Soybean AQP Genes
4.2. Phylogenetic Tree
4.3. Tissular Expression Profile Analyses
4.4. Heat Stress and Hormone Treatments
4.5. qRT-PCR
4.6. Promoter Element Prediction
4.7. Promoter Cloning and Arabidopsis Transformation
4.8. GUS Activity Detection
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| AQP | aquaporin |
| ABA | abscisic acid |
| ACC | l-aminocyclopropane-l-carboxylic acid |
| SA | salicylic acid |
| MeJA | methyl jasmonate |
| qRT-PCR | quantitative real-time PCR |
| GUS | beta-glucuronidase |
References
- Fox, A.R.; Maistriaux, L.C.; Chaumont, F. Toward understanding of the high number of plant aquaporin isoforms and multiple regulation mechanisms. Plant Sci. 2017, 264, 179–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaumont, F.; Barrieu, F.; Wojcik, E.; Chrispeels, M.J.; Jung, R. Aquaporins constitute a large and highly divergent protein family in maize. Plant Physiol. 2001, 125, 1206–1215. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.Y.; Kim, D.G.; Kim, Y.O.; Kim, J.S.; Kang, H. An expression analysis of a gene family encoding plasma membrane aquaporins in response to abiotic stresses in Arabidopsis thaliana. Plant Mol. Biol. 2004, 54, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, J.; Ishikawa, F.; Yamaguchi, T.; Uemura, M.; Maeshima, M. Identification of 33 rice aquaporin genes and analysis of their expression and function. Plant Cell Physiol. 2005, 46, 1568–1577. [Google Scholar] [CrossRef] [PubMed]
- Park, W.; Scheffler, B.E.; Bauer, P.J.; Campbell, B.T. Identification of the family of aquaporin genes and their expression in upland cotton (Gossypium hirsutum L.). BMC Plant Biol. 2010, 10, 142–158. [Google Scholar] [CrossRef] [PubMed]
- Reuscher, S.; Akiyama, M.; Mori, C.; Aoki, K.; Shibata, D.; Shiratake, K. Genome-wide identification and expression analysis of aquaporins in tomato. PLoS ONE 2013, 8, e79052. [Google Scholar] [CrossRef] [PubMed]
- Yue, C.; Cao, H.; Wang, L.; Zhou, Y.; Hao, X.; Zeng, J.; Wang, X.; Yang, Y. Molecular cloning and expression analysis of tea plant aquaporin (AQP) gene family. Plant Physiol. Biochem. 2014, 83, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Abascal, F.; Irisarri, I.; Zardoya, R. Diversity and evolution of membrane intrinsic proteins. Biochim. Biophys. Acta 2014, 1840, 1468–1481. [Google Scholar] [CrossRef] [Green Version]
- Afzal, Z.; Howton, T.C.; Sun, Y.L.; Shahid Mukhtar, M. The roles of aquaporins in plant stress responses. J. Dev. Biol. 2016, 4, 9. [Google Scholar] [CrossRef]
- Maurel, C.; Verdoucq, L.; Luu, D.T.; Santoni, V. Plant aquaporins: Membrane channels with multiple integrated functions. Annu. Rev. Plant Biol. 2008, 59, 595–624. [Google Scholar] [CrossRef]
- Alexandersson, E.; Fraysse, L.; Sjövall-Larsen, S.; Gustavsson, S.; Fellert, M.; Karlsson, M.; Johanson, U.; Kjellbom, P. Whole gene family expression and drought stress regulation of aquaporins. Plant Mol. Biol. 2005, 59, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Boursiac, Y.; Chen, S.; Luu, D.T.; Sorieul, M.; van den Dries, N.; Maurel, C. Early effects of salinity on water transport in Arabidopsis roots molecular and cellular features of aquaporin expression. Plant Physiol. 2005, 139, 790–805. [Google Scholar] [CrossRef] [PubMed]
- Putpeerawit, P.; Sojikul, P.; Thitamadee, S.; Narangajavana, J. Genome-wide analysis of aquaporin gene family and their responses to water-deficit stress conditions in cassava. Plant Physiol. Biochem. 2017, 121, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Kadam, S.; Abril, A.; Dhanapal, A.P.; Koester, R.P.; Vermerris, W.; Jose, S.; Fritschi, F.B. Characterization and regulation of aquaporin genes of sorghum [Sorghum bicolor (L.) Moench] in response to waterlogging stress. Front. Plant Sci. 2017, 8, 862. [Google Scholar] [CrossRef] [PubMed]
- Hove, R.M.; Bhave, M. Plant aquaporins with non-aqua functions: Deciphering the signature sequences. Plant Mol. Biol. 2011, 75, 413–430. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.J.; Xu, S.C.; Liu, N.; Zhang, G.W.; Hu, Q.Z.; Xu, Z.S.; Gong, Y.M. Identification of the AQP members involved in abiotic stress responses from Arabidopsis. Gene 2018, 646, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, J.; Yu, J.W.; Park, S.W. Genome-wide analysis and expression profiling of the Solanum tuberosum aquaporins. Plant Physiol. Biochem. 2013, 73, 392–404. [Google Scholar] [CrossRef]
- Szucs, A.; Jäger, K.; Jurca, M.E.; Fábián, A.; Bottka, S.; Zvara, A.; Barnabás, B.; Fehér, A. Histological and microarray analysis of the direct effect of water shortage alone or combined with heat on early grain development in wheat (Triticum aestivum). Physiol. Plant 2010, 140, 174–188. [Google Scholar] [CrossRef]
- Chen, K.; Wang, X.; Fessehaie, A.; Yin, Y.; Wang, X.; Arora, R. Is expression of aquaporins (plasma membrane intrinsic protein 2s, PIP2s) associated with thermonasty (leaf-curling) in Rhododendron. J. Plant Physiol. 2013, 170, 1447–1454. [Google Scholar] [CrossRef]
- Christou, A.; Filippou, P.; Manganaris, G.A.; Fotopoulos, V. Sodium hydrosulfide induces systemic thermotolerance to strawberry plants through transcriptional regulation of heat shock proteins and aquaporin. BMC Plant Biol. 2014, 14, 42. [Google Scholar] [CrossRef]
- Saha, P.; Sade, N.; Arzani, A.; Wilhelmi, M.D.M.R.; Coe, K.M.; Li, B.; Blumwald, E. Effects of abiotic stress on physiological plasticity and water use of Setaria viridis (L.). Plant Sci. 2016, 251, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Obaid, A.Y.; Sabir, J.S.; Atef, A.; Liu, X.; Edris, S.; El-Domyati, F.M.; Mutwakil, M.Z.; Gadalla, N.O.; Hajrah, N.H.; Al-Kordy, M.A.; et al. Analysis of transcriptional response to heat stress in Rhazya stricta. BMC Plant Biol. 2016, 16, 252. [Google Scholar] [CrossRef] [PubMed]
- Georgii, E.; Jin, M.; Zhao, J.; Kanawati, B.; Schmitt-Kopplin, P.; Albert, A.; Winkler, J.B.; Schäffner, A.R. Relationships between drought, heat and air humidity responses revealed by transcriptome-metabolome co-analysis. BMC Plant Biol. 2017, 17, 120. [Google Scholar] [CrossRef]
- Maruyama, K.; Ogata, T.; Kanamori, N.; Yoshiwara, K.; Goto, S.; Yamamoto, Y.Y.; Tokoro, Y.; Noda, C.; Takaki, Y.; Urawa, H.; et al. Design of an optimal promoter involved in the heat-induced transcriptional pathway in Arabidopsis, soybean, rice and maize. Plant J. 2017, 89, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Eldakak, M.; Paudel, B.; Kim, D.W.; Hemmati, H.; Basu, C.; Rohila, J.S. Leaf proteome analysis reveals prospective drought and heat stress response mechanisms in soybean. BioMed Res. Int. 2016, 2016, 6021047. [Google Scholar] [CrossRef] [PubMed]
- Valdés-López, O.; Batek, J.; Gomez-Hernandez, N.; Nguyen, C.T.; Isidra-Arellano, M.C.; Zhang, N.; Joshi, T.; Xu, D.; Hixson, K.K.; Weitz, K.K.; et al. Soybean roots grown under heat stress show global changes in their transcriptional and proteomic Profiles. Front. Plant Sci. 2016, 7, 517. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, N.; Donnart, T.; Nouri, M.Z.; Komatsu, S. Tissue-specific defense and thermo-adaptive mechanisms of soybean seedlings under heat stress revealed by proteomic approach. J. Proteome Res. 2010, 9, 4189–4204. [Google Scholar] [CrossRef]
- Zhang, D.Y.; Ali, Z.; Wang, C.B.; Xu, L.; Yi, J.X.; Xu, Z.L.; Liu, X.Q.; He, X.L.; Huang, Y.H.; Khan, I.A.; et al. Genome-wide sequence characterization and expression analysis of major intrinsic proteins in soybean (Glycine max L.). PLoS ONE 2013, 8, e56312. [Google Scholar] [CrossRef]
- Deshmukh, R.K.; Vivancos, J.; Guérin, V.; Sonah, H.; Labbé, C.; Belzile, F.; Bélanger, R.R. Identification and functional characterization of silicon transporters in soybean using comparative genomics of major intrinsic proteins in Arabidopsis and rice. Plant Mol. Biol. 2013, 3, 303–315. [Google Scholar] [CrossRef]
- Beitz, E.; Wu, B.; Holm, L.M.; Schultz, J.E.; Zeuthen, T. Point mutations in the aromatic/arginine region in aquaporin 1 allow passage of urea, glycerol, ammonia, and protons. Proc. Natl. Acad. Sci. USA. 2006, 103, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Hub, J.S.; de Groot, B.L. Mechanism of selectivity in aquaporins and aquaglyceroporins. Proc. Natl. Acad. Sci. USA. 2008, 105, 1198–1203. [Google Scholar] [CrossRef] [PubMed]
- Mitani-Ueno, N.; Yamaji, N.; Zhao, F.J.; Ma, J.F. The aromatic/arginine selectivity filter of NIP aquaporins plays a critical role in substrate selectivity for silicon, boron, and arsenic. J. Exp. Bot. 2011, 62, 4391–4398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prak, S.; Hem, S.; Boudet, J.; Viennois, G.; Sommerer, N.; Rossignol, M.; Maurel, C.; Santoni, V. Multiple phosphorylations in the C-terminal tail of plant plasma membrane aquaporins: Role in subcellular trafficking of AtPIP2;1 in response to salt stress. Mol. Cell. Proteom. 2008, 7, 1019–1030. [Google Scholar] [CrossRef] [PubMed]
- Daniels, M.J.; Yeager, M. Phosphorylation of aquaporin PvTIP3; 1 defined by mass spectrometry and molecular modeling. Biochemistry 2005, 44, 14443–14454. [Google Scholar] [CrossRef] [PubMed]
- Guenther, J.F.; Chanmanivone, N.; Galetovic, M.P.; Wallace, I.S.; Cobb, J.A.; Roberts, D.M. Phosphorylation of soybean nodulin 26 on serine 262 enhances water permeability and is regulated developmentally and by osmotic signals. Plant Cell 2003, 15, 981–991. [Google Scholar] [CrossRef]
- Pou, A.; Jeanguenin, L.; Milhiet, T.; Batoko, H.; Chaumont, F.; Hachez, C. Salinity-mediated transcriptional and post-translational regulation of the Arabidopsis aquaporin PIP2;7. Plant Mol. Biol. 2016, 92, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Soto, G.; Alleva, K.; Mazzella, M.A.; Amodeo, G.; Muschietti, J.P. AtTIP1;3 and AtTIP5;1, the only highly expressed Arabidopsis pollen-specific aquaporins, transport water and urea. FEBS Lett. 2008, 582, 4077–4082. [Google Scholar] [CrossRef]
- Pang, Y.; Li, L.; Ren, F.; Lu, P.; Wei, P.; Cai, J.; Xin, L.; Zhang, J.; Chen, J.; Wang, X. Overexpression of the tonoplast aquaporin AtTIP5;1 conferred tolerance to boron toxicity in Arabidopsis. J. Genet. Genom. 2010, 37, 389–397. [Google Scholar] [CrossRef]
- Lindsay, E.R.; Maathuis, F.J. Arabidopsis thaliana NIP7;1 is involved in tissue arsenic distribution and tolerance in response to arsenate. FEBS Lett. 2016, 590, 779–786. [Google Scholar] [CrossRef]
- Hachez, C.; Zelazny, E.F. Chaumont modulating the expression of aquaporin genes in planta: A key to understand their physiological functions. Biochem. Biophys. Acta. 2006, 1758, 1142–1156. [Google Scholar] [CrossRef]
- Kapilan, R.; Vaziri, M.; Zwiazek, J.J. Regulation of aquaporins in plants under stress. Biol. Res. 2018, 51, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawłowicz, I.; Masajada, K. Aquaporins as a link between water relations and photosynthetic pathway in abiotic stress tolerance in plants. Gene 2019, 687, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Winter, D.; Vinegar, B.; Nahal, H.; Ammar, R.; Wilson, G.V.; Provart, N.J. An “Electronic Fluorescent Pictograph” browser for exploring and analyzing large-scale biological data sets. PLoS ONE 2007, 2, e718. [Google Scholar] [CrossRef] [PubMed]
- Lamaoui, M.; Jemo, M.; Datla, R.; Bekkaoui, F. Heat and drought stresses in crops and approaches for their mitigation. Front. Chem. 2018, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Umeda, M.; Uchimiya, H. Isolation and expression analysis of two rice genes encoding the major intrinsic protein. Plant Mol. Biol. 1994, 26, 2003–2007. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.P.; Young, L.; Bonham-Smith, P.; Gusta, L.V. Characterization and expression of plasma and tonoplast membrane aquaporins in primed seed of Brassica napus during germination under stress conditions. Plant Mol. Biol. 1999, 40, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Mariaux, J.B.; Bockel, C.; Salamini, F.; Bartels, D. Dessication- and abscisic acid-responsive genes encoding major intrinsic proteins (MIPs) from the resurrection plant Craterostigma plantagineum. Plant Mol. Biol. 1998, 38, 1089–1099. [Google Scholar] [CrossRef]
- Hu, W.; Yuan, Q.; Wang, Y.; Cai, R.; Deng, X.; Wang, J.; Zhou, S.; Chen, M.; Chen, L.; Huang, C.; et al. Overexpression of a wheat aquaporin gene, TaAQP8, enhances salt stress tolerance in transgenic tobacco. Plant Cell Physiol. 2012, 53, 2127–2141. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.Q.; Yang, F.; Gao, J.P. Isolation of RhTIP1;1, an aquaporin gene and its expression in rose flowers in response to ethylene and water deficit. Postharvest Biol. Tec. 2009, 51, 407–413. [Google Scholar] [CrossRef]
- Phillips, A.L.; Huttly, A.K. Cloning of two gibberellin-regulated cDNAs from Arabidopsis thaliana by subtractive hybridization: Expression of the tonoplast water channel, gamma-TIP, is increased by GA3. Plant Mol. Biol. 1994, 24, 603–615. [Google Scholar] [CrossRef]
- Morillon, R.; Catterou, M.; Sangwan, R.S.; Sangwan, B.S.; Lassalles, J.P. Brassinolide may control aquaporin activities in Arabidopsis thaliana. Planta 2001, 212, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Gattolin, S.; Sorieul, M.; Hunter, P.R.; Khonsari, R.H.; Frigerio, L. In vivo imaging of the tonoplast intrinsic protein family in Arabidopsis roots. BMC Plant Biol. 2009, 9, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Santoni, V.; Maurel, C. Plant aquaporins: Roles in plant physiology. Biochim. Biophys. Acta. 2014, 1840, 1574–1582. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, D.; Zou, D.; Luo, F.; Wang, X.; Zheng, Y.; Li, X. A cotton gene encoding a plasma membrane aquaporin is involved in seedling development and in response to drought stress. Acta Biochim. Biophys. Sin. 2013, 45, 104–114. [Google Scholar] [CrossRef]
- Xu, W.; Dai, W.; Yan, H.; Li, S.; Shen, H.; Chen, Y.; Xu, H.; Sun, Y.; He, Z.; Ma, M. Arabidopsis NIP3;1 Plays an important role in arsenic uptake and root-to-shoot translocation under arsenite stress conditions. Mol. Plant 2015, 8, 722–733. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Tong, J.; He, X.; Xu, Z.; Xu, L.; Wei, P.; Huang, Y.; Brestic, M.; Ma, H.; Shao, H. A novel soybean intrinsic protein gene, GmTIP2;3, involved in responding to osmotic Stress. Front. Plant Sci. 2016, 6, 1237. [Google Scholar] [CrossRef] [PubMed]
- De Paula Santos Martins, C.; Pedrosa, A.M.; Du, D.; Gonçalves, L.P.; Yu, Q.; Gmitter, F.G., Jr.; Costa, M.G. Genome-wide characterization and expression analysis of major intrinsic proteins during abiotic and biotic stresses in sweet orange (Citrus sinensis L. Osb.). PLoS ONE 2015, 10, e0138786. [Google Scholar] [CrossRef]
- Johanson, U.; Karlsson, M.; Johansson, I.; Gustavsson, S.; Sjövall, S.; Fraysse, L.; Weig, A.R.; Kjellbom, P. The complete set of genes encoding major intrinsic proteins in Arabidopsis provides a framework for a new nomenclature for major intrinsic proteins in plants. Plant Physiol. 2001, 126, 1358–1369. [Google Scholar] [CrossRef]
- Gupta, A.B.; Sankararamakrishnan, R. Genome-wide analysis of major intrinsic proteins in the tree plant Populus trichocarpa: Characterization of XIP subfamily of aquaporins from evolutionary perspective. BMC Plant. Biol. 2009, 9, 134. [Google Scholar] [CrossRef]
- Ariani, A.; Gepts, P. Genome-wide identification and characterization of aquaporin gene family in common bean (Phaseolus vulgaris L.). Mol. Genet. Genom. 2015, 290, 1771–1785. [Google Scholar] [CrossRef]
- Giovannetti, M.; Balestrini, R.; Volpe, V.; Guether, M.; Straub, D.; Costa, A.; Ludewig, U.; Bonfante, P. Two putative-aquaporin genes are differentially expressed during arbuscular mycorrhizal symbiosis in Lotus japonicus. BMC Plant Biol. 2012, 12, 186. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.J.; He, G.H.; Zheng, W.J.; Lu, P.P.; Chen, M.; Gong, Y.M.; Ma, Y.Z.; Xu, Z.S. Foxtail millet NF-Y families: Genome-wide survey and evolution analyses identified two functional genes important in abiotic stresses. Front. Plant Sci. 2015, 6, 1142. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4, molecular evolutionary genetics analysis MEGA software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Libault, M.; Farmer, A.; Joshi, T.; Takahashi, K.; Langley, R.J.; Franklin, L.D.; He, J.; Xu, D.; May, G.; Stacey, G. An integrated transcriptome atlas of the crop model Glycine max, and its use in comparative analyses in plants. Plant J. 2010, 63, 86–99. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wan, Q.; He, X.; Ning, L.; Huang, Y.; Xu, Z.; Liu, J.; Shao, H. Genome-wide characterization of the ankyrin repeats gene family under salt stress in soybean. Sci. Total Environ. 2016, 568, 899–909. [Google Scholar] [CrossRef] [Green Version]
- Quach, T.N.; Nguyen, H.T.; Valliyodan, B.; Joshi, T.; Xu, D.; Nguyen, H.T. Genome-wide expression analysis of soybean NF-Y genes reveals potential function in development and drought response. Mol. Genet. Genom. 2015, 290, 1095–1115. [Google Scholar] [CrossRef]
- Liu, H.; Che, Z.; Zeng, X.; Zhou, X.; Sitoe, H.M.; Wang, H.; Yu, D. Genome-wide analysis of calcium-dependent protein kinases and their expression patterns in response to herbivore and wounding stresses in soybean. Funct. Integr. Genom. 2016, 16, 481–493. [Google Scholar] [CrossRef]
- Zhou, F.; Guo, Y.; Qiu, L.J. Genome-wide identification and evolutionary analysis of leucine-rich repeat receptor-like protein kinase genes in soybean. BMC Plant Biol. 2016, 16, 58–70. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCт method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Jefferson, R.A. Assaying chimeric genes in plants: The GUS gene fusion system. Plant Mol. Biol. Rep. 1987, 5, 387–405. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Jing, R.; Mao, X. Functional characterization of TaSnRK2.8 promoter in response to abiotic stresses by deletion analysis in transgenic Arabidopsis. Front. Plant Sci. 2017, 8, 1198. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Xu, Y.; Huang, D.; Miao, H.; Liu, J.; Jia, C.; Hu, W.; Valarezo, A.V.; Xu, B.; Jin, Z. Identification of a novel promoter from banana aquaporin family gene (MaTIP1;2) which responses to drought and salt-stress in transgenic Arabidopsis thaliana. Plant Physiol. Biochem. 2018, 128, 163–169. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Number | Gene Name | Gene Symbol | Chromosome Location | CDS Length (bp) | Protein Length (aa) | pI | MW (kDa) | Numbers of Phosphorylation Sites |
|---|---|---|---|---|---|---|---|---|
| GmAQP1 | GmPIP1;1 | Glyma.03G078700 | 3:18018230..18021565 | 855 | 284 | 9.10 | 30.41 | Ser: 7 Thr: 1 Tyr: 2 |
| GmAQP2 | GmPIP1;2 | Glyma.18G198300 | 18:51879812..51881980 | 864 | 287 | 9.26 | 30.89 | Ser: 8 Thr: 3 Tyr: 2 |
| GmAQP3 | GmPIP1;3 | Glyma.01G220600 | 1:54066066..54068057 | 861 | 286 | 9.13 | 30.79 | Ser: 7 Thr: 1 Tyr: 1 |
| GmAQP4 | GmPIP1;4 | Glyma.11G023200 | 11:1656129..1658174 | 861 | 286 | 8.84 | 30.74 | Ser: 6 Thr: 1 Tyr: 1 |
| GmAQP5 | GmPIP1;5 | Glyma.05G208700 | 5:41267148..41268807 | 864 | 287 | 9.00 | 30.89 | Ser: 9 Thr: 1 Tyr: 2 |
| GmAQP6 | GmPIP1;6 | Glyma.08G015300 | 8:1202356..1204135 | 870 | 289 | 8.61 | 30.89 | Ser: 9 Thr: 1 Tyr: 2 |
| GmAQP7 | GmPIP1;7 | Glyma.14G061500 | 14:4894197..4896207 | 870 | 289 | 8.60 | 30.61 | Ser: 9 Thr: 3 Tyr: 3 |
| GmAQP8 | GmPIP1;8 | Glyma.11G228000 | 11:36767510..36769078 | 870 | 289 | 7.01 | 30.77 | Ser: 11 Thr: 2 Tyr: 3 |
| GmAQP9 | GmPIP1;9 | Glyma.01G113400 | 1:1138834287..38837846 | 1092 | 363 | 9.31 | 39.41 | Ser: 14 Thr: 11 Tyr: 7 |
| GmAQP10 | GmPIP1;10 | Glyma.02G255000 | 2:44207467..44209844 | 960 | 319 | 9.38 | 34.19 | Ser: 12 Thr: 8 Tyr: 4 |
| GmAQP11 | GmPIP2;1 | Glyma.04G003200 | 4:227991..229365 | 828 | 275 | 9.45 | 29.30 | Ser: 9 Thr: 0 Tyr: 2 |
| GmAQP12 | GmPIP2;2 | Glyma.06G003200 | 6:264336..265850 | 837 | 278 | 9.35 | 29.30 | Ser: 9 Thr: 0 Tyr: 2 |
| GmAQP13 | GmPIP2;3 | Glyma.11G146500 | 11:11300751..11303007 | 861 | 286 | 6.95 | 29.55 | Ser: 8 Thr: 7 Tyr: 4 |
| GmAQP14 | GmPIP2;4 | Glyma.12G075400 | 12:5747587..5750039 | 861 | 286 | 6.19 | 30.38 | Ser: 7 Thr: 9 Tyr: 3 |
| GmAQP15 | GmPIP2;5 | Glyma.12G172500 | 12:32929324..32931027 | 864 | 287 | 8.25 | 30.67 | Ser: 9 Thr: 2 Tyr: 4 |
| GmAQP16 | GmPIP2;6 | Glyma.13G325900 | 13:40664607..40666361 | 864 | 287 | 8.26 | 30.84 | Ser: 8 Thr: 2 Tyr: 3 |
| GmAQP17 | GmPIP2;7 | Glyma.03G180900 | 3:41279731..41281496 | 861 | 286 | 8.98 | 30.79 | Ser: 12 Thr: 8 Tyr: 4 |
| GmAQP18 | GmPIP2;8 | Glyma.19G181300 | 19:44007407..44009765 | 858 | 285 | 9.15 | 30.61 | Ser: 13 Thr: 9 Tyr: 4 |
| GmAQP19 | GmPIP2;9 | Glyma.02G073600 | 2:6421649..6424849 | 858 | 285 | 8.29 | 30.56 | Ser: 10 Thr: 6 Tyr: 1 |
| GmAQP20 | GmPIP2;10 | Glyma.16G155000 | 16:31513389..31517035 | 858 | 285 | 8.29 | 30.68 | Ser: 10 Thr: 6 Tyr: 3 |
| GmAQP21 | GmPIP2;11 | Glyma.16G155100 | 16:31522994..31524889 | 858 | 285 | 8.29 | 30.41 | Ser: 10 Thr: 6 Tyr: 3 |
| GmAQP22 | GmPIP2;12 | Glyma.02G073700 | 2:6434383..6437873 | 858 | 285 | 8.59 | 30.37 | Ser: 11 Thr: 6 Tyr: 3 |
| GmAQP23 | GmPIP2;13 | Glyma.10G211000 | 10:44343751..44346957 | 891 | 296 | 7.70 | 30.44 | Ser: 9 Thr: 6 Tyr: 3 |
| GmAQP24 | GmPIP2;14 | Glyma.20G179700 | 20:41738693..41741581 | 855 | 284 | 8.29 | 31.73 | Ser: 10 Thr: 7 Tyr: 4 |
| GmAQP25 | GmTIP1;1 | Glyma.02G094700 | 2:8409966..8411440 | 759 | 252 | 5.12 | 25.96 | Ser: 3 Thr: 3 Tyr: 0 |
| GmAQP26 | GmTIP1;2 | Glyma.18G286700 | 18:60989768..60991401 | 759 | 252 | 5.49 | 26.04 | Ser: 4 Thr: 2 Tyr: 0 |
| GmAQP27 | GmTIP1;3 | Glyma.10G290600 | 10:50271428..50272965 | 759 | 252 | 6.01 | 26.02 | Ser: 3 Thr: 2 Tyr: 0 |
| GmAQP28 | GmTIP1;4 | Glyma.11G143100 | 11:10892421..10894109 | 759 | 252 | 5.37 | 25.79 | Ser: 5 Thr: 1 Tyr: 1 |
| GmAQP29 | GmTIP1;5 | Glyma.12G066200 | 12:4870480..4871652 | 738 | 245 | 6.02 | 25.03 | Ser: 5 Thr: 1 Tyr: 1 |
| GmAQP30 | GmTIP1;6 | Glyma.13G333100 | 13:41270585..41271998 | 759 | 252 | 5.16 | 26.01 | Ser: 2 Thr: 1 Tyr: 0 |
| GmAQP31 | GmTIP1;7 | Glyma.03G185900 | 3:41779243..41780564 | 753 | 250 | 6.01 | 25.45 | Ser: 0 Thr: 3 Tyr: 1 |
| GmAQP32 | GmTIP1;8 | Glyma.19G186100 | 19:44258426..44259853 | 753 | 250 | 6.01 | 25.53 | Ser: 0 Thr: 3 Tyr: 1 |
| GmAQP33 | GmTIP1;9 | Glyma.13G146300 | 13:24436182..24438466 | 753 | 250 | 10.01 | 26.54 | Ser: 8 Thr: 2 Tyr: 0 |
| GmAQP34 | GmTIP1;10 | Glyma.20G098600 | 20:34184591..34191923 | 732 | 243 | 6.17 | 25.52 | Ser: 6 Thr: 3 Tyr: 0 |
| GmAQP35 | GmTIP2;1 | Glyma.01G208200 | 1:53110677..53113455 | 750 | 249 | 5.08 | 25.28 | Ser: 4 Thr: 2 Tyr: 1 |
| GmAQP36 | GmTIP2;2 | Glyma.11G034000 | 11:2476012..2478825 | 750 | 249 | 5.08 | 25.32 | Ser: 2 Thr: 1 Tyr: 2 |
| GmAQP37 | GmTIP2;3 | Glyma.07G018000 | 7:1435523..1437651 | 747 | 248 | 5.69 | 25.23 | Ser: 6 Thr: 1 Tyr: 1 |
| GmAQP38 | GmTIP2;4 | Glyma.08G203000 | 8:16535219..16537122 | 747 | 248 | 5.69 | 25.27 | Ser: 3 Thr: 2 Tyr: 2 |
| GmAQP39 | GmTIP2;5 | Glyma.13G356000 | 13:43018922..43020336 | 744 | 247 | 5.51 | 25.07 | Ser: 2 Thr: 2 Tyr: 2 |
| GmAQP40 | GmTIP2;6 | Glyma.15G018100 | 15:1393557..1395809 | 744 | 247 | 5.50 | 25.07 | Ser: 3 Thr: 3 Tyr: 1 |
| GmAQP41 | GmTIP2;7 | Glyma.19G035400 | 19:4625496..4626575 | 714 | 237 | 5.57 | 24.08 | Ser: 4 Thr: 1 Tyr: 1 |
| GmAQP42 | GmTIP3;1 | Glyma.09G160500 | 9:35913523..35915582 | 768 | 255 | 6.54 | 27.03 | Ser: 5 Thr: 1 Tyr: 1 |
| GmAQP43 | GmTIP3;2 | Glyma.16G210000 | 16:36421819..36424304 | 768 | 255 | 6.54 | 27.11 | Ser: 5 Thr: 0 Tyr: 1 |
| GmAQP44 | GmTIP3;3 | Glyma.10G174400 | 10:40238530..40240337 | 765 | 254 | 7.13 | 27.08 | Ser: 5 Thr: 0 Tyr: 0 |
| GmAQP45 | GmTIP3;4 | Glyma.20G216100 | 20:44068541..44070258 | 765 | 254 | 7.88 | 27.07 | Ser: 5 Thr: 3 Tyr: 1 |
| GmAQP46 | GmTIP4;1 | Glyma.04G083200 | 4:7019276..7020984 | 741 | 246 | 5.71 | 25.65 | Ser: 3 Thr: 2 Tyr: 0 |
| GmAQP47 | GmTIP4;2 | Glyma.06G084600 | 6:6498818..6500103 | 741 | 246 | 5.71 | 25.63 | Ser: 5 Thr: 2 Tyr: 2 |
| GmAQP48 | GmTIP5;1 | Glyma.09G224700 | 9:41742635..41743884 | 759 | 252 | 7.82 | 26.30 | Ser: 9 Thr: 2 Tyr: 2 |
| GmAQP49 | GmNIP1;1 | Glyma.05G162600 | 5:35105884..35108185 | 813 | 270 | 9.67 | 28.68 | Ser: 8 Thr: 5 Tyr: 1 |
| GmAQP50 | GmNIP1;2 | Glyma.08G120200 | 8:9268559..9270946 | 825 | 274 | 9.48 | 29.27 | Ser: 9 Thr: 3 Tyr: 1 |
| GmAQP51 | GmNIP1;3 | Glyma.13G224900 | 13:32551102..32553703 | 822 | 273 | 7.76 | 28.93 | Ser: 6 Thr: 4 Tyr: 2 |
| GmAQP52 | GmNIP1;4 | Glyma.15G087300 | 15:6704209..6706791 | 822 | 273 | 7.74 | 28.83 | Ser: 6 Thr: 4 Tyr: 2 |
| GmAQP53 | GmNIP1;5 | Glyma.08G120100 | 8:9262302..9265834 | 816 | 271 | 6.41 | 28.91 | Ser: 9 Thr: 3 Tyr: 5 |
| GmAQP54 | GmNIP1;6 | Glyma.05G162500 | 5:35371190..35375992 | 816 | 271 | 8.87 | 28.66 | Ser: 11 Thr: 8 Tyr: 1 |
| GmAQP55 | GmNIP2;1 | Glyma.07G217700 | 7:39062920..39065820 | 789 | 262 | 8.14 | 28.13 | Ser: 4 Thr: 2 Tyr: 1 |
| GmAQP56 | GmNIP3;1 | Glyma.14G174300 | 14:43721841..43723560 | 813 | 270 | 8.23 | 28.66 | Ser: 15 Thr: 11 Tyr: 3 |
| GmAQP57 | GmNIP4;1 | Glyma.02G246700 | 2:46541265..46543675 | 786 | 261 | 7.61 | 27.61 | Ser: 23 Thr: 5 Tyr: 4 |
| GmAQP58 | GmNIP4;2 | Glyma.14G069500 | 14:5711153..5714115 | 786 | 261 | 8.25 | 27.59 | Ser: 17 Thr: 4 Tyr: 4 |
| GmAQP59 | GmNIP5;1 | Glyma.09G238200 | 9:42824943..42829709 | 882 | 293 | 8.55 | 30.44 | Ser: 28 Thr: 9 Tyr: 1 |
| GmAQP60 | GmNIP5;2 | Glyma.10G221100 | 10:44670892..44676555 | 900 | 299 | 7.68 | 31.15 | Ser: 6 Thr: 5 Tyr: 0 |
| GmAQP61 | GmNIP6;1 | Glyma.18G259500 | 18:58816436..58821548 | 888 | 295 | 6.96 | 30.55 | Ser: 19 Thr: 5 Tyr: 0 |
| GmAQP62 | GmNIP6;2 | Glyma.08G217400 | 8:17701761..17706495 | 921 | 306 | 9.13 | 31.74 | Ser: 8 Thr: 5 Tyr: 0 |
| GmAQP63 | GmNIP6;3 | Glyma.15G003900 | 15:355676..359967 | 915 | 304 | 8.25 | 31.28 | Ser: 7 Thr: 2 Tyr: 0 |
| GmAQP64 | GmNIP7;1 | Glyma.02G140500 | 2:14348789..14351092 | 891 | 296 | 8.46 | 31.42 | Ser: 10 Thr: 1 Tyr: 3 |
| GmAQP65 | GmNIP7;2 | Glyma.10G033600 | 10:2898411..2900795 | 870 | 289 | 8.69 | 30.82 | Ser: 7 Thr: 1 Tyr: 1 |
| GmAQP66 | GmSIP1;1 | Glyma.02G069800 | 2:6061309..6065568 | 921 | 306 | 9.27 | 26.64 | Ser: 0 Thr: 2 Tyr: 0 |
| GmAQP67 | GmSIP1;2 | Glyma.16G151300 | 16:30813218..30817735 | 738 | 245 | 9.27 | 26.43 | Ser: 0 Thr: 2 Tyr: 0 |
| GmAQP68 | GmSIP1;3 | Glyma.19G108400 | 19:35912781..35923174 | 747 | 248 | 9.12 | 26.56 | Ser: 1 Thr: 2 Tyr: 3 |
| GmAQP69 | GmSIP1;4 | Glyma.16G043800 | 16:4096288..4102424 | 747 | 248 | 9.10 | 26.52 | Ser: 2 Thr: 2 Tyr: 2 |
| GmAQP70 | GmSIP1;5 | Glyma.12G097800 | 12:8369034..8369846 | 720 | 239 | 9.99 | 26.02 | Ser: 4 Thr: 3 Tyr: 1 |
| GmAQP71 | GmSIP1;6 | Glyma.06G307000 | 6:48987251..48988278 | 720 | 239 | 9.91 | 25.87 | Ser: 4 Thr: 4 Tyr: 1 |
| GmAQP72 | GmSIP2;1 | Glyma.03G119300 | 3:35075328..35078322 | 693 | 230 | 9.45 | 25.24 | Ser: 6 Thr: 7 Tyr: 0 |
| GmAQP73 | GmSIP2;2 | Glyma.19G123600 | 19:37949307..37951724 | 711 | 236 | 9.45 | 25.97 | Ser: 10 Thr: 6 Tyr: 0 |
| GmAQP74 | GmXIP1;1 | Glyma.12G023600 | 12:1729006..1730580 | 939 | 312 | 7.02 | 33.75 | Ser: 6 Thr: 2 Tyr: 3 |
| GmAQP75 | GmXIP1;2 | Glyma.11G097800 | 11:7449914..7450765 | 852 | 283 | 6.50 | 30.12 | Ser: 9 Thr: 5 Tyr: 3 |
| Gene Name | Gene Symbol | TM Number | NPA Motifs | ar/R Selectivity Filters | FPs | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LB | LE | H2 | H5 | LE1 | LE2 | P1 | P2 | P3 | P4 | P5 | |||
| GmPIP1;1 | Glyma.03G078700 | 6 | NPA | NPA | F | H | T | R | E | S | A | F | W |
| GmPIP1;2 | Glyma.18G198300 | 6 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| GmPIP1;3 | Glyma.01G220600 | 6 | NPA | NPA | F | H | T | R | E | S | A | F | W |
| GmPIP1;4 | Glyma.11G023200 | 6 | NPA | NPA | F | H | T | R | E | S | A | F | W |
| GmPIP1;5 | Glyma.05G208700 | 6 | NPA | NPA | F | H | T | R | E | S | A | F | W |
| GmPIP1;6 | Glyma.08G015300 | 6 | NPA | NPA | F | H | T | R | E | S | A | F | W |
| GmPIP1;7 | Glyma.14G061500 | 6 | NPA | NPA | F | H | T | R | E | S | A | F | W |
| GmPIP1;8 | Glyma.11G228000 | 6 | NPA | NPA | F | H | T | R | E | S | A | F | W |
| GmPIP1;9 | Glyma.01G113400 | 6 | NPA | NPA | F | H | T | R | E | S | A | F | W |
| GmPIP1;10 | Glyma.02G255000 | 6 | NPA | NPA | F | H | T | R | E | S | A | F | W |
| GmPIP2;1 | Glyma.04G003200 | 6 | NPA | NPA | F | H | T | R | M | S | A | F | W |
| GmPIP2;2 | Glyma.06G003200 | 6 | NPA | NPA | F | H | T | R | M | S | A | F | W |
| GmPIP2;3 | Glyma.11G146500 | 6 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| GmPIP2;4 | Glyma.12G075400 | 6 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| GmPIP2;5 | Glyma.12G172500 | 6 | NPA | NPA | F | H | T | R | Q | S | A | Y | W |
| GmPIP2;6 | Glyma.13G325900 | 6 | NPA | NPA | F | H | T | R | Q | S | A | Y | W |
| GmPIP2;7 | Glyma.03G180900 | 6 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| GmPIP2;8 | Glyma.19G181300 | 6 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| GmPIP2;9 | Glyma.02G073600 | 6 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| GmPIP2;10 | Glyma.16G155000 | 6 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| GmPIP2;11 | Glyma.16G155100 | 6 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| GmPIP2;12 | Glyma.02G073700 | 6 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| GmPIP2;13 | Glyma.10G211000 | 6 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| GmPIP2;14 | Glyma.20G179700 | 6 | NPA | NPA | F | H | T | R | Q | S | A | F | W |
| GmTIP1;1 | Glyma.02G094700 | 6 | NPA | NPA | H | I | A | V | T | S | A | Y | W |
| GmTIP1;2 | Glyma.18G286700 | 6 | NPA | NPA | H | I | A | V | T | S | A | Y | W |
| GmTIP1;3 | Glyma.10G290600 | 6 | NPA | NPA | H | I | A | V | T | C | A | Y | W |
| GmTIP1;4 | Glyma.11G143100 | 6 | NPA | NPA | H | I | A | V | T | S | A | Y | W |
| GmTIP1;5 | Glyma.12G066200 | 6 | NPA | NPA | H | I | A | V | T | S | A | Y | W |
| GmTIP1;6 | Glyma.13G333100 | 6 | NPA | NPA | H | I | A | V | T | S | A | Y | W |
| GmTIP1;7 | Glyma.03G185900 | 6 | NPA | NPA | H | I | A | V | T | T | A | Y | W |
| GmTIP1;8 | Glyma.19G186100 | 6 | NPA | NPA | H | I | A | V | T | T | A | Y | W |
| GmTIP1;9 | Glyma.13G146300 | 6 | NPA | NPA | H | I | A | A | T | S | A | Y | W |
| GmTIP1;10 | Glyma.20G098600 | 6 | NPA | NPA | H | I | A | V | T | S | A | Y | W |
| GmTIP2;1 | Glyma.01G208200 | 6 | NPA | NPA | H | I | G | R | T | S | A | Y | W |
| GmTIP2;2 | Glyma.11G034000 | 6 | NPA | NPA | H | I | G | R | T | S | A | Y | W |
| GmTIP2;3 | Glyma.07G018000 | 6 | NPA | NPA | H | I | G | R | T | S | A | Y | W |
| GmTIP2;4 | Glyma.08G203000 | 6 | NPA | NPA | H | I | G | R | T | S | A | Y | W |
| GmTIP2;5 | Glyma.13G356000 | 6 | NPA | NPA | H | I | G | R | T | S | A | Y | W |
| GmTIP2;6 | Glyma.15G018100 | 6 | NPA | NPA | H | I | G | R | T | S | A | Y | W |
| GmTIP2;7 | Glyma.19G035400 | 6 | NPA | NPA | H | I | G | R | T | S | A | Y | W |
| GmTIP3;1 | Glyma.09G160500 | 6 | NPA | NPA | H | I | A | L | T | A | S | F | W |
| GmTIP3;2 | Glyma.16G210000 | 6 | NPA | NPA | H | I | A | L | T | A | S | F | W |
| GmTIP3;3 | Glyma.10G174400 | 6 | NPA | NPA | H | I | A | R | T | A | A | F | W |
| GmTIP3;4 | Glyma.20G216100 | 6 | NPA | NPA | H | I | A | R | T | A | A | F | W |
| GmTIP4;1 | Glyma.04G083200 | 6 | NPA | NPA | H | I | A | R | S | S | A | Y | W |
| GmTIP4;2 | Glyma.06G084600 | 6 | NPA | NPA | H | I | A | R | S | S | A | Y | W |
| GmTIP5;1 | Glyma.09G224700 | 6 | NPA | NPA | S | V | G | C | V | A | A | Y | W |
| GmNIP1;1 | Glyma.05G162600 | 6 | NPA | NPA | W | V | A | R | F | S | A | Y | V |
| GmNIP1;2 | Glyma.08G120200 | 6 | NPA | NPA | W | V | A | R | F | S | A | Y | V |
| GmNIP1;3 | Glyma.13G224900 | 6 | NPA | NPA | W | V | A | R | F | S | A | Y | V |
| GmNIP1;4 | Glyma.15G087300 | 6 | NPA | NPA | W | V | A | R | F | S | A | Y | V |
| GmNIP1;5 | Glyma.08G120100 | 6 | NPA | NPA | W | V | A | R | F | S | A | Y | L |
| GmNIP1;6 | Glyma.05G162500 | 6 | NPA | NPV | W | V | A | R | F | S | A | Y | L |
| GmNIP2;1 | Glyma.07G217700 | 6 | NPA | NPA | W | V | A | R | F | S | A | Y | V |
| GmNIP3;1 | Glyma.14G174300 | 6 | NPA | NPA | S | V | A | R | Y | S | A | Y | I |
| GmNIP4;1 | Glyma.02G246700 | 6 | NPA | NPA | W | V | A | R | L | S | A | Y | V |
| GmNIP4;2 | Glyma.14G069500 | 6 | NPA | NPA | W | V | A | R | L | S | A | Y | V |
| GmNIP5;1 | Glyma.09G238200 | 6 | NPA | NPA | G | S | G | R | L | T | A | Y | F |
| GmNIP5;2 | Glyma.10G221100 | 6 | NPS | NPV | A | I | G | R | Y | T | A | Y | L |
| GmNIP6;1 | Glyma.18G259500 | 6 | NPA | NPA | G | S | G | R | L | T | A | Y | F |
| GmNIP6;2 | Glyma.08G217400 | 6 | NPA | NPV | N | I | S | R | F | T | A | Y | L |
| GmNIP6;3 | Glyma.15G003900 | 6 | NPA | NPV | T | I | G | R | Y | T | A | Y | L |
| GmNIP7;1 | Glyma.02G140500 | 6 | NPA | NPA | A | V | G | R | Y | S | A | Y | M |
| GmNIP7;2 | Glyma.10G033600 | 6 | NPA | NPA | A | V | G | R | Y | S | A | Y | M |
| GmSIP1;1 | Glyma.02G069800 | 6 | NPT | NPA | I | I | P | F | M | A | A | Y | W |
| GmSIP1;2 | Glyma.16G151300 | 6 | NPT | NPA | I | I | P | F | M | A | A | Y | W |
| GmSIP1;3 | Glyma.19G108400 | 6 | NPT | NPA | V | V | P | N | M | A | A | Y | W |
| GmSIP1;4 | Glyma.16G043800 | 6 | NPT | NPA | V | V | P | N | M | A | A | Y | W |
| GmSIP1;5 | Glyma.12G097800 | 6 | NPS | NPA | N | A | P | N | L | A | A | Y | W |
| GmSIP1;6 | Glyma.06G307000 | 6 | NPS | NPA | N | A | P | N | L | A | A | Y | W |
| GmSIP2;1 | Glyma.03G119300 | 6 | NP_ | NPA | S | H | G | S | I | V | A | Y | W |
| GmSIP2;2 | Glyma.19G123600 | 6 | NP_ | NPA | S | H | G | S | I | V | A | Y | W |
| GmXIP1;1 | Glyma.12G023600 | 6 | NPI | SPA | V | V | A | R | E | C | A | F | W |
| GmXIP1;2 | Glyma.11G097800 | 6 | SPV | NPA | V | V | V | R | D | C | A | F | W |
| Gene Name | Gene Symbol | HSE | ABRE | CGTCA | ERE | TCA |
|---|---|---|---|---|---|---|
| GmPIP1;7 | Glyma.14G061500 | 2 | 2 | 0 | 1 | 1 |
| GmPIP1;8 | Glyma.11G228000 | 2 | 0 | 0 | 0 | 2 |
| GmPIP2;4 | Glyma.12G075400 | 3 | 0 | 0 | 0 | 3 |
| GmPIP2;5 | Glyma.12G172500 | 1 | 0 | 1 | 1 | 0 |
| GmPIP2;6 | Glyma.13G325900 | 1 | 0 | 1 | 0 | 1 |
| GmPIP2;13 | Glyma.10G211000 | 2 | 0 | 1 | 0 | 1 |
| GmPIP2;14 | Glyma.20G179700 | 2 | 0 | 0 | 0 | 2 |
| GmTIP1;7 | Glyma.03G185900 | 0 | 6 | 1 | 2 | 0 |
| GmTIP2;2 | Glyma.11G034000 | 1 | 0 | 1 | 0 | 1 |
| GmTIP2;6 | Glyma.15G018100 | 3 | 0 | 0 | 1 | 0 |
| GmTIP4;1 | Glyma.04G083200 | 2 | 0 | 0 | 0 | 2 |
| GmSIP1;3 | Glyma.19G108400 | 2 | 1 | 0 | 0 | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, Z.-J.; Liu, N.; Zhang, G.-W.; Niu, F.-G.; Xu, S.-C.; Gong, Y.-M. Investigation of the AQP Family in Soybean and the Promoter Activity of TIP2;6 in Heat Stress and Hormone Responses. Int. J. Mol. Sci. 2019, 20, 262. https://doi.org/10.3390/ijms20020262
Feng Z-J, Liu N, Zhang G-W, Niu F-G, Xu S-C, Gong Y-M. Investigation of the AQP Family in Soybean and the Promoter Activity of TIP2;6 in Heat Stress and Hormone Responses. International Journal of Molecular Sciences. 2019; 20(2):262. https://doi.org/10.3390/ijms20020262
Chicago/Turabian StyleFeng, Zhi-Juan, Na Liu, Gu-Wen Zhang, Fu-Ge Niu, Sheng-Chun Xu, and Ya-Ming Gong. 2019. "Investigation of the AQP Family in Soybean and the Promoter Activity of TIP2;6 in Heat Stress and Hormone Responses" International Journal of Molecular Sciences 20, no. 2: 262. https://doi.org/10.3390/ijms20020262