Mining Late Embryogenesis Abundant (LEA) Family Genes in Cleistogenes songorica, a Xerophyte Perennial Desert Plant

 ,
,
Abstract
:1. Introduction
2. Results
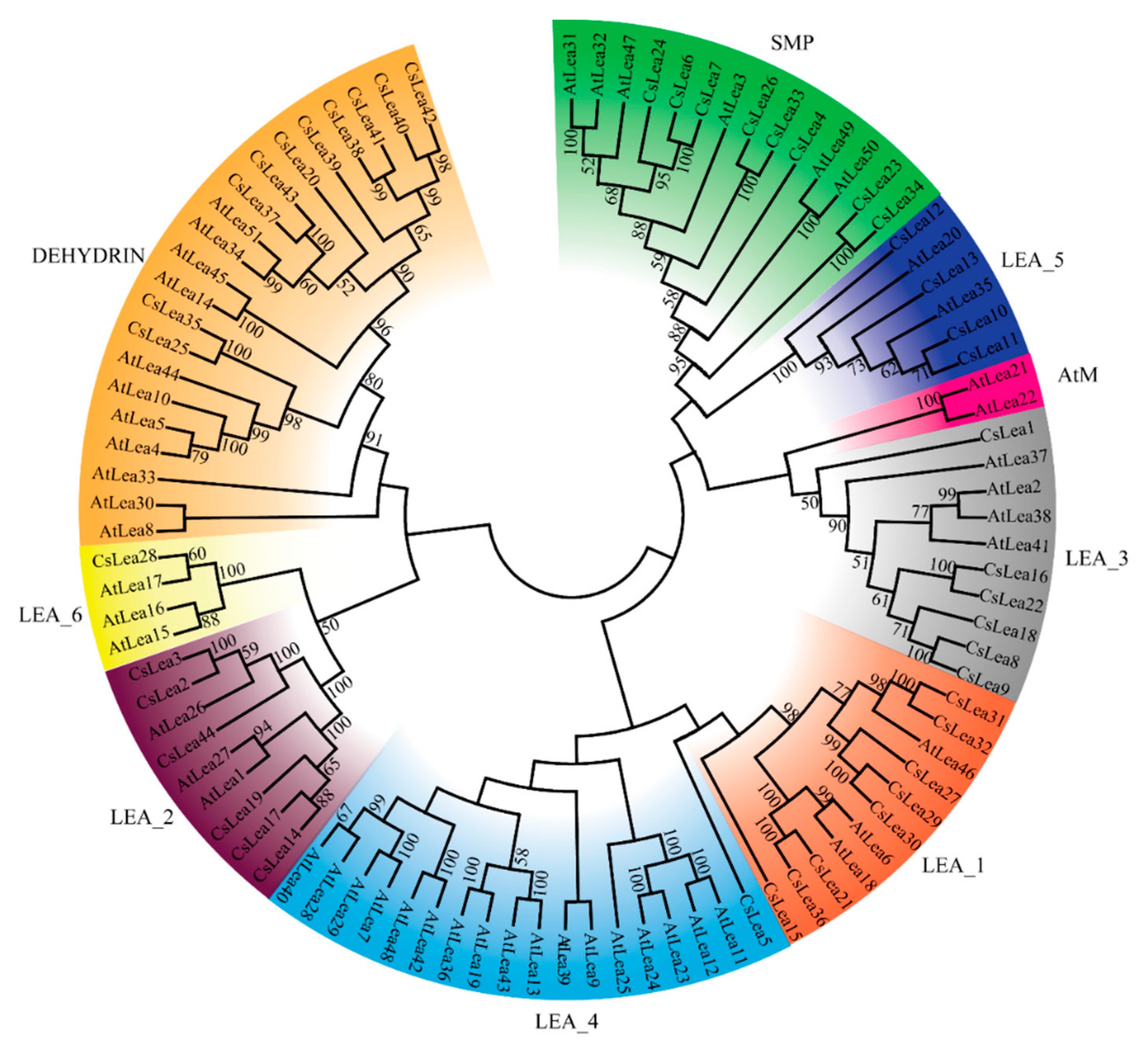
2.1. Identification of CsLEA Genes and Phylogenetic Analysis
2.2. Structures, Physiochemical Properties and Subcellular Localizations of CsLEA Proteins
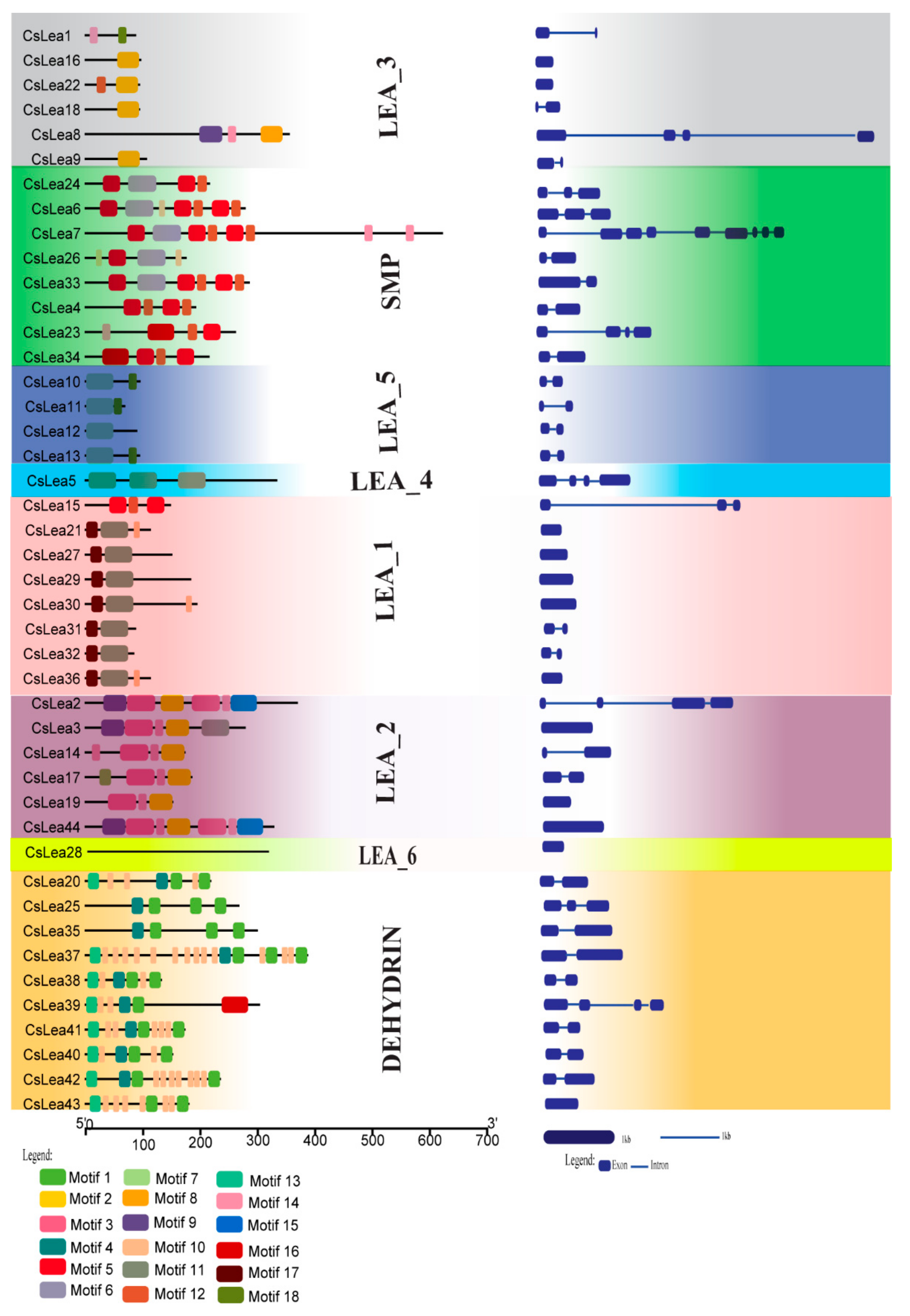
2.3. Gene and Motif Structure Analyses
2.4. Sequence Alignment of C. Songorica and Arabidopsis Dehydrin Proteins
2.5. Cis-Regulatory Element in C. Songorica LEA Gene Promoters
2.6. Chromosomal Mapping of CsLEA Genes
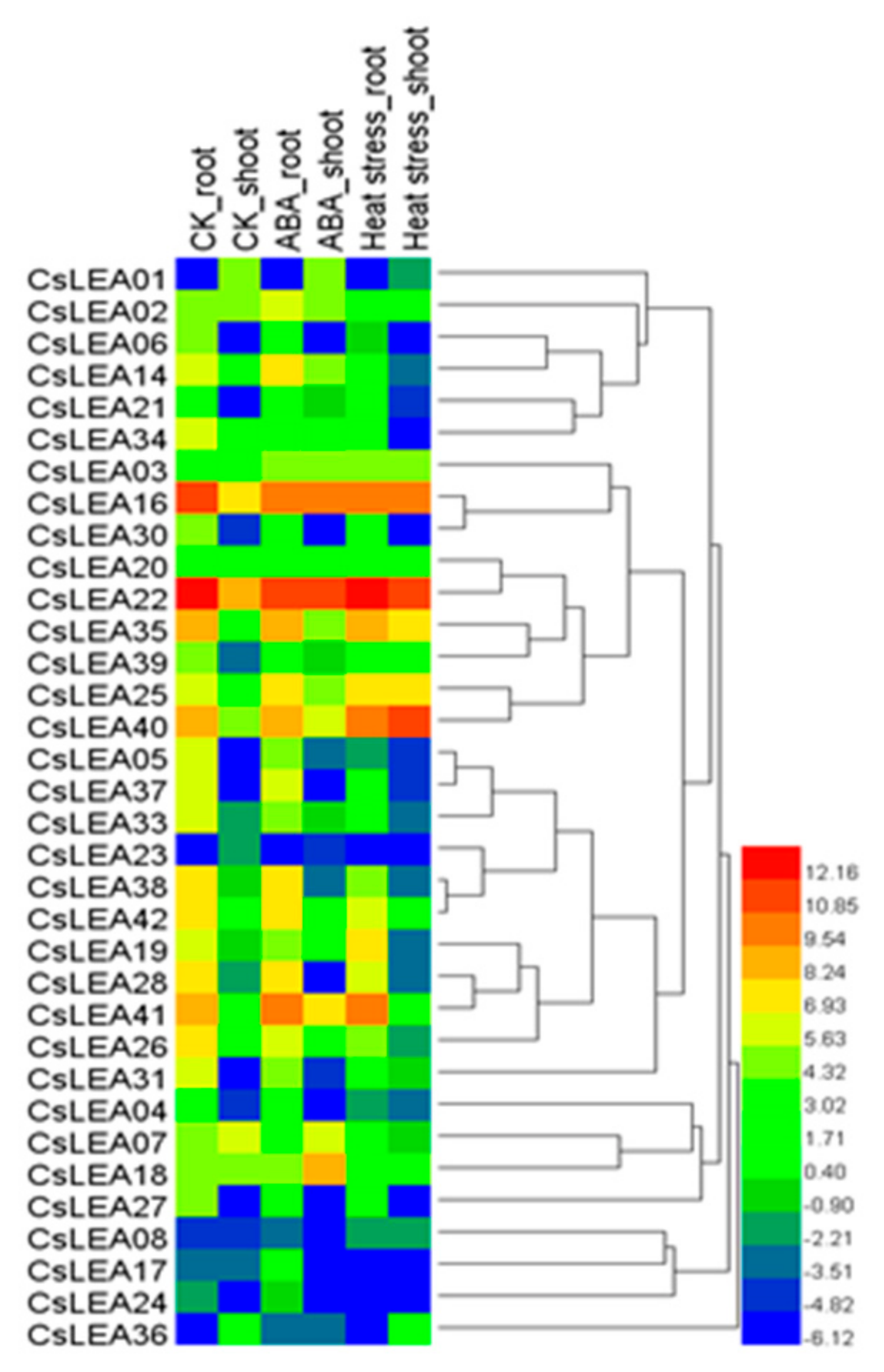
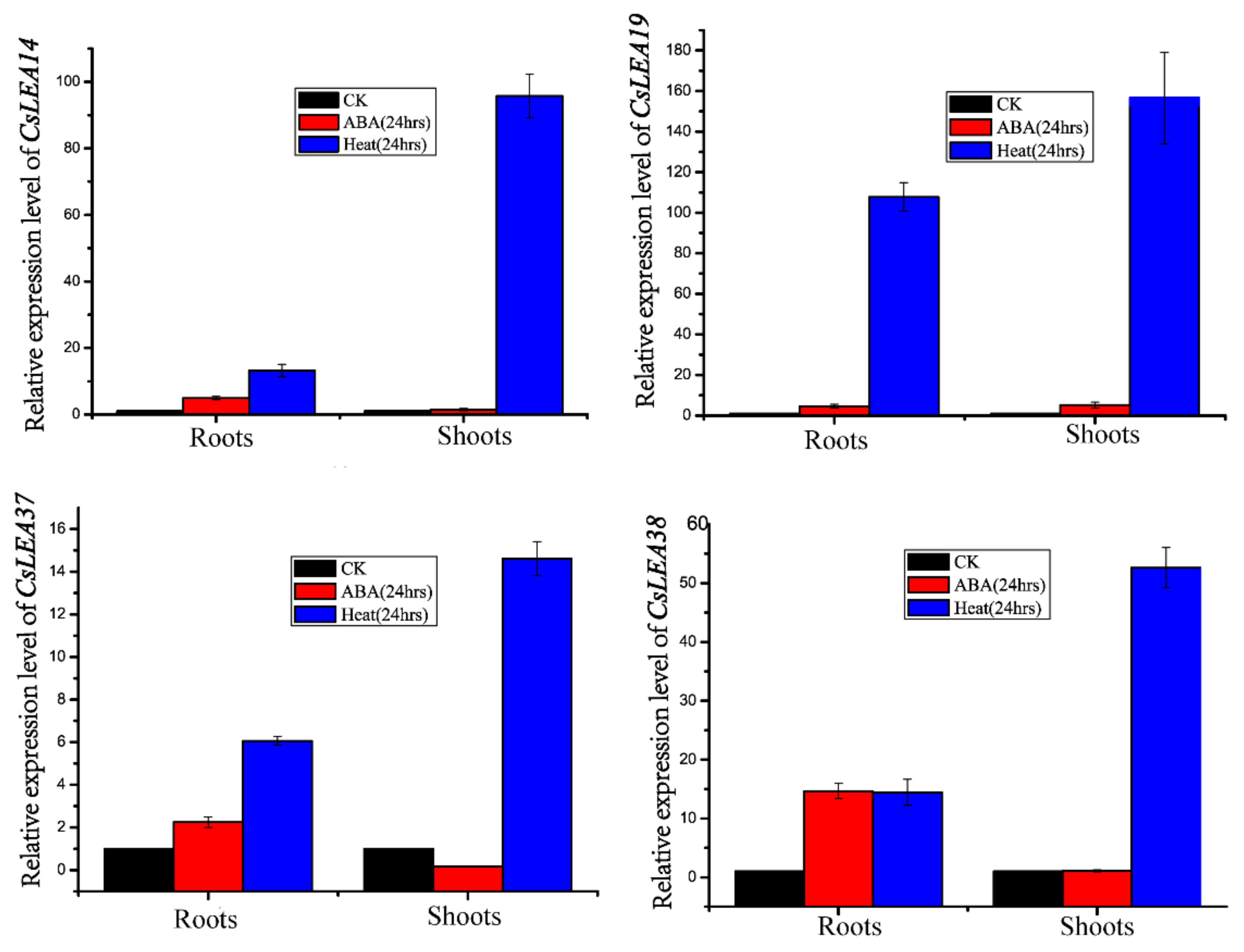
2.7. Gene Expression Analysis qRT-PCR Validation
3. Discussion
3.1. Phylogeny Analysis and Protein Sequence Analysis
3.2. Protein Domain Analysis
3.3. C. songorica Gene Promoter and Gene Expression Analysis
4. Materials and Methods
4.1. Mining LEA Genes in the C. Songorica Genome
4.2. Multiple Sequence Alignment and Phylogenetic Analysis of CsLEA Family Proteins.
4.3. In Silico Analyses of CsLEA Proteins.
4.4. Analysis of Cis-Regulatory Elements and Motifs
4.5. Plant Material Preparation and Transcriptomic Data Analysis
4.6. Gene Expressions Analysis.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shahbaz, M.; Ashraf, M. Critical reviews in plant sciences improving salinity tolerance in cereals improving salinity tolerance in cereals. Crit. Rev. Plant Sci. 2013, 32, 237–249. [Google Scholar] [CrossRef]
- Verma, G.; Dhar, Y.V.; Srivastava, D.; Kidwai, M.; Chauhan, P.S.; Bag, S.K.; Asif, M.H.; Chakrabarty, D. Genome-wide analysis of rice dehydrin gene family: Its evolutionary conservedness and expression pattern in response to peg induced dehydration stress. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Zhong, X.-J.; He, J.; Jin, S.-H.; Guo, H.-D.; Yu, X.-F.; Zhou, Y.-J.; Li, X.; Ma, M.-D.; Chen, Q.-B.; et al. Genome-wide identification, characterization, and stress-responsive expression profiling of genes encoding lea (late embryogenesis abundant) proteins in moso bamboo (phyllostachys edulis). PLoS ONE 2016, 11, e0165953. [Google Scholar] [CrossRef] [PubMed]
- Dure, L., 3rd; Greenway, S.C.; Galau, G.A. Developmental biochemistry of cottonseed embryogenesis and germination: Changing messenger ribonucleic acid populations as shown by in vitro and in vivo protein synthesis. Biochemistry 1981, 20, 4162–4168. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhu, H.; Cheng, W.; Liu, Y.; Cheng, X.; Sun, J.; Gill, S.S.; Tuteja, N. Polyamines and fabiotic stress tolerance in plants. Plant Signal Behav. 2010, 5, 26–33. [Google Scholar]
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [PubMed]
- Kikawada, T.; Nakahara, Y.; Kanamori, Y.; Iwata, K.-I.; Watanabe, M.; McGee, B.; Tunnacliffe, A.; Okuda, T. Dehydration-induced expression of lea proteins in an anhydrobiotic chironomid. Biochem. Biophys. Res. Commun. 2006, 348, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Hand, S.C.; Menze, M.A.; Toner, M.; Boswell, L.; Moore, D. Lea proteins during water stress: Not just for plants anymore. Annu. Rev. Physiol. 2011, 73, 115–134. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, M.; Olvera-Carrillo, Y.; Garciarrubio, A.; Campos, F.; Covarrubias, A.A. The enigmatic lea proteins and other hydrophilins. Plant Physiol. 2008, 148, 6–24. [Google Scholar] [CrossRef] [PubMed]
- Boucher, V.; Buitink, J.; Lin, X.; Boudet, J.; Hoekstra, F.A.; Hundertmark, M.; Renard, D.; Leprince, O. Mtpm25 is an atypical hydrophobic late embryogenesis-abundant protein that dissociates cold and desiccation-aggregated proteins. Plant Cell Environ. 2010, 33, 418–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakrabortee, S.; Tripathi, R.; Watson, M.; Schierle, G.S.; Kurniawan, D.P.; Kaminski, C.F.; Wise, M.J.; Tunnacliffe, A. Intrinsically disordered proteins as molecular shields. Mol. Biosyst. 2012, 8, 210–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olveracarrillo, Y.; Campos, F.; Reyes, J.L.; Garciarrubio, A.; Covarrubias, A.A. Functional analysis of the group 4 late embryogenesis abundant proteins reveals their relevance in the adaptive response during water deficit in arabidopsis. Plant Physiol. 2010, 154, 373–390. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed]
- Bies-Etheve, N.; Gaubier-Comella, P.; Debures, A.; Lasserre, E.; Jobet, E.; Raynal, M.; Cooke, R.; Delseny, M. Inventory, evolution and expression profiling diversity of the lea (late embryogenesis abundant) protein gene family in arabidopsis thaliana. Plant Mol. Biol. 2008, 67, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Ciccarelli, F.D.; Bork, P. The Why Domain Mediates the Response to Desiccation in Plants and Bacteria; Oxford University Press: Oxford, UK, 2005; pp. 1304–1307. [Google Scholar]
- Jaspard, E.; Hunault, G. Comparison of amino acids physico-chemical properties and usage of late embryogenesis abundant proteins, hydrophilins and why domain. PLoS ONE 2014, 9, e109570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouillon, J.-M.; Gustafsson, P.; Harryson, P. Structural investigation of disordered stress proteins. Comparison of full-length dehydrins with isolated peptides of their conserved segments. Plant Physiol. 2006, 141, 638–650. [Google Scholar] [CrossRef] [PubMed]
- Tolleter, D.; Jaquinod, M.; Mangavel, C.; Passirani, C.; Saulnier, P.; Manon, S.; Teyssier, E.; Payet, N.; Avelange-Macherel, M.H.; Macherel, D. Structure and function of a mitochondrial late embryogenesis abundant protein are revealed by desiccation. Plant Cell 2007, 19, 1580–1589. [Google Scholar] [CrossRef] [PubMed]
- Koag, M.-C.; Wilkens, S.; Fenton, R.D.; Resnik, J.; Vo, E.; Close, T.J. The k-segment of maize dhn1 mediates binding to anionic phospholipid vesicles and concomitant structural changes. Plant Physiol. 2009, 150, 1503–1514. [Google Scholar] [CrossRef] [PubMed]
- Rahman, L.N.; Chen, L.; Nazim, S.; Bamm, V.V.; Yaish, M.W.; Moffatt, B.A.; Dutcher, J.R.; Harauz, G. Interactions of intrinsically disordered thellungiella salsuginea dehydrins tsdhn-1 and tsdhn-2 with membranes—synergistic effects of lipid composition and temperature on secondary structure. Biochem. Cell Biol. 2010, 88, 791–807. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Zhou, M.; Zhou, X.; Jin, Y.; Xu, M.; Lin, J. Jclea, a novel lea-like protein from jatropha curcas, confers a high level of tolerance to dehydration and salinity in arabidopsis thaliana. PLoS ONE 2014, 8, e83056. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kong, L.; Liu, Z.; Jahufer, Z.; Duan, Z.; Huo, Y.; Di, H.; Wang, Y. Stress-induced expression in arabidopsis with a dehydrin lea protein from cleistogenes songorica, a xerophytic desert grass. Plant Omics 2015, 8, 485–492. [Google Scholar]
- Waie, B.; Rajam, M.V. Effect of increased polyamine biosynthesis on stress responses in transgenic tobacco by introduction of human s -adenosylmethionine gene. Plant Sci. 2003, 164, 727–734. [Google Scholar] [CrossRef]
- Zhang, J.; Duan, Z.; Zhang, D.; Zhang, J.; Di, H.; Wu, F.; Wang, Y. Co-transforming bar and cslea enhanced tolerance to drought and salt stress in transgenic alfalfa (medicago sativa l.). Biochem. Biophys. Res. Commun. 2016, 472, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Lan, T. Functional characterization of the late embryogenesis abundant (lea) protein gene family from pinus tabuliformis (pinaceae) in Escherichia coli. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, L.; Xing, X.; Sun, L.; Pan, J.; Kong, X.; Zhang, M.; Li, D. Zmlea3, a multifunctional group 3 lea protein from maize (zea mays l.), is involved in biotic and abiotic stresses. Plant Cell Physiol. 2013, 54, 944–959. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; John, U.P.; Wang, Y.; Li, X.; Gunawardana, D.; Polotnianka, R.M.; Spangenberg, G.C.; Nan, Z. Targeted mining of drought stress-responsive genes from est resources in Cleistogenes songorica. J. Plant Physiol. 2011, 168, 1844–1851. [Google Scholar] [CrossRef] [PubMed]
- Hundertmark, M.; Hincha, D.K. Lea (late embryogenesis abundant) proteins and their encoding genes in Arabidopsis thaliana. BMC Genomics 2008, 9. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.; Dennsteele, C.V.; Iii, L.D. Sequence and characterization of 6 lea proteins and their genes from cotton. Plant Mol. Biol. 1988, 11, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Galau, G.A.; Wang, H.Y.; Hughes, D.W. Cotton Lea5 and Lea14 encode atypical late embryogenesis-abundant proteins. Plant Physiol. 1993, 101, 695–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, S.; Tan, L.; Hu, Z.; Chen, G.; Wang, G.; Hu, T. Molecular characterization and functional analysis by heterologous expression in e. Coli under diverse abiotic stresses for oslea5, the atypical hydrophobic lea protein from oryza sativa l. Mol. Genet. Genom. 2012, 287, 39–54. [Google Scholar] [CrossRef] [PubMed]
- Lan, T.; Gao, J.; Zeng, Q.Y. Genome-wide analysis of the lea (late embryogenesis abundant) protein gene family in Populus trichocarpa. Tree Genet. Genom. 2013, 9, 253–264. [Google Scholar] [CrossRef]
- Cheng, F.; Wu, J.; Wang, X. Genome triplication drove the diversification of brassica plants. Hortic. Res. 2014, 1. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Xiong, Z.; Zheng, J.; Xu, D.; Zhu, Z.; Xiang, J.; Gan, J.; Nadia, R.; Yin, Y.; Li, M. Genome-wide identification, structural analysis and new insights into late embryogenesis abundant (lea) gene family formation pattern in Brassica napus. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Organization of cis-acting regulatory elements in osmotic- and cold-stress-responsive promoters. Trends Plant Sci. 2005, 10, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-S.; Zhu, H.-B.; Jin, G.-L.; Liu, H.-L.; Wu, W.-R.; Zhu, J. Genome-scale identification and analysis of lea genes in rice (oryza sativa l.). Plant Sci. 2007, 172, 414–420. [Google Scholar] [CrossRef]
- Li, X.; Cao, J. Late embryogenesis abundant (lea) gene family in maize: Identification, evolution, and expression profiles. Plant Mol. Biol. Rep. 2015, 34, 15–28. [Google Scholar] [CrossRef]
- Lång, V.; Palva, E.T. The expression of a rab-related gene, rab18, is induced by abscisic acid during the cold acclimation process of arabidopsis thaliana (l.) heynh. Plant Mol. Boil. 1992, 20, 951–962. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, B.; Tang, K.; Hsu, C.C.; Xie, S.; Du, H.; Yang, Y.; Tao, W.A.; Zhu, J.K. An Arabidopsis nucleoporin NUP85 modulates plant responses to ABA and salt stress. PLoS Genet. 2017, 13, e1007124. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, D.; Kalmar, E.; Torok, Z.; Tompa, P. Chaperone activity of erd10 and erd14, two disordered stress-related plant proteins. Plant Physiol. 2008, 147, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, S.K.; Kutzer, M.; Procek, J.; Grobner, G.; Harryson, P. Tunable membrane binding of the intrinsically disordered dehydrin lti30, a cold-induced plant stress protein. Plant Cell 2011, 23, 2391–2404. [Google Scholar] [CrossRef] [PubMed]
- Candat, A.; Macherel, D. The ubiquitous distribution of late embryogenesis abundant proteins across cell compartments in Arabidopsis offers tailored protection against abiotic stress. Plant Cell 2014, 26, 3148–3166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanin, M.; Brini, F.; Ebel, C.; Toda, Y.; Takeda, S.; Masmoudi, K. Plant dehydrins and stress tolerance: Versatile proteins for complex mechanisms. Plant Signal Behav. 2011, 6, 1503–1509. [Google Scholar] [CrossRef] [PubMed]
- Close, T.J. Dehydrins: A commonalty in the response of plants to dehydration and low temperature. Physiologia Plantarum 1997, 100, 291–296. [Google Scholar] [CrossRef]
- Jiang, S.; Wang, J.; Liu, X.; Liu, Y.; Guo, C.; Zhang, L.; Han, J.; Wu, X.; Xue, D.; Gomaa, A.E.; et al. Drwh, a novel why domain-containing hydrophobic lea5c protein from deinococcus radiodurans, protects enzymatic activity under oxidative stress. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Franco-Zorrilla, J.M.; López-Vidriero, I.; Carrasco, J.L.; Godoy, M.; Vera, P.; Solano, R. DNA-binding specificities of plant transcription factors and their potential to define target genes. Proc. Nati. Acad. Sci. USA 2014, 111, 2367–2372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.S.; Lee, J.H.; Kim, J.J.; Kim, C.H.; Jun, S.S.; Hong, Y.N. Molecular and functional characterization of CaLea6, the gene for a hydrophobic LEA protein from capsicum annuum. Gene 2005, 344, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-C.; Kim, Y.-H.; Jeong, J.C.; Kim, C.Y.; Lee, H.-S.; Bang, J.-W.; Kwak, S.-S. Sweetpotato late embryogenesis abundant 14 (iblea14) gene influences lignification and increases osmotic- and salt stress-tolerance of transgenic calli. Planta 2011, 233, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, P.; Li, C.; Pan, Y.; Jiang, X.; Zhu, D.; Zhao, Q.; Yu, J. SiLEA14, a novel atypical lea protein, confers abiotic stress resistance in foxtail millet. BMC Plant Biol. 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Chiappetta, A.; Muto, A.; Bruno, L.; Woloszynska, M.; Lijsebettens, M.V.; Bitonti, M.B. A dehydrin gene isolated from feral olive enhances drought tolerance in arabidopsis transgenic plants. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brini, F.; Hanin, M.; Lumbreras, V.; Amara, I.; Khoudi, H.; Hassairi, A.; Pages, M.; Masmoudi, K. Overexpression of wheat dehydrin dhn-5 enhances tolerance to salt and osmotic stress in Arabidopsis thaliana. Plant Cell Rep. 2007, 26, 2017–2026. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Mistry, J.; Tate, J.; Coggill, P.; Heger, A.; Pollington, J.E.; Gavin, O.L.; Gunasekaran, P.; Ceric, G.; Forslund, K.; et al. The pfam protein families database. Nucleic Acids Res. 2010, 38, D211–D222. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Letunic, I.; Copley, R.R.; Schmidt, S.; Ciccarelli, F.D.; Doerks, T.; Schultz, J.; Ponting, C.P.; Bork, P. Smart 4.0: Towards genomic data integration. Nucleic Acids Res. 2004, 32, D142–D144. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. Cdd: Ncbi’s conserved domain database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.-F.; Doolittle, R.F. Progressive sequence alignment as a prerequisitetto correct phylogenetic trees. J. Mol. Evol. 1987, 25, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Mega6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Higgins, D.G.; Sharp, P.M. Clustal: A package for performing multiple sequence alignment on a microcomputer. Gene 1988, 73, 237–244. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. Expasy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed]
- Emanuelsson, O.; Brunak, S.; von Heijne, G.; Nielsen, H. Locating proteins in the cell using targetp, signalp and related tools. Nat. Protoc. 2007, 2, 953–971. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. Meme suite: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Rombauts, S.; Dehais, P.; Van Montagu, M.; Rouze, P. Plantcare, a plant cis-acting regulatory element database. Nucleic Acids Res. 1999, 27, 295–296. [Google Scholar] [CrossRef] [PubMed]
- Yue, R.; Lu, C.; Sun, T.; Peng, T.; Han, X.; Qi, J.; Yan, S.; Tie, S. Identification and expression profiling analysis of calmodulin-binding transcription activator genes in maize (Zea mays L.) under abiotic and biotic stresses. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Petrov, V.; Vermeirssen, V.; De, C.I.; Van, B.F.; Minkov, I.; Vandepoele, K.; Gechev, T.S. Identification of cis-regulatory elements specific for different types of reactive oxygen species in arabidopsis thaliana. Gene 2012, 499, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Passricha, N.; Saifi, S.; Ansari, M.W.; Tuteja, N. Prediction and validation of cis-regulatory elements in 5’ upstream regulatory regions of lectin receptor-like kinase gene family in rice. Protoplasma 2017, 254, 669–684. [Google Scholar] [CrossRef] [PubMed]
- Acharya, B.R.; Jeon, B.W.; Zhang, W.; Assmann, S.M. Open stomata 1 (OST1) is limiting in abscisic acid responses of arabidopsis guard cells. New Phytol. 2013, 200, 1049–1063. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. Tophat: Discovering splice junctions with RNA-seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of rna-seq experiments with tophat and cufflinks. Nat. Protoc. 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. Degseq: An r package for identifying differentially expressed genes from rna-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Wang, Y.; Liu, Z.; Cheng, H.; Xue, Y. Hemi: A toolkit for illustrating heatmaps. PLoS ONE 2014, 9, e111988. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, M.; Takita, E.; Harada, H.; Sakurai, N.; Suzuki, H.; Ohyama, K.; Shibata, D.; Misawa, N. Pathway engineering of brassica napus seeds using multiple key enzyme genes involved in ketocarotenoid formation. J. Exp. Bot. 2009, 60, 1319–1332. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CsLEA Subfamilies | Gene Names | Functional cis-Element Names and Sequences | |||
|---|---|---|---|---|---|
| MBS. (CGGTC) | G-Box (GTGCAT/CACGAC) | ABRE (GACACGTACGT) | CGTCA Motif | ||
| Functions | |||||
| Drought Responsive (MYB Binding Site) | Light Responsive | Abscisic Acid Responsive | MeJA Responsive | ||
| LEA_1 | CsLEA29 | 3 | 1 | 1 | 0 |
| CsLEA30 | 2 | 3 | 1 | 4 | |
| CsLEA31 | 4 | 1 | 1 | 0 | |
| CsLEA32 | 2 | 2 | 1 | 0 | |
| CsLEA36 | 4 | 5 | 2 | 2 | |
| LEA_2 | CsLEA2 | 2 | 1 | 0 | 3 |
| CsLEA3 | 0 | 3 | 2 | 2 | |
| CsLEA14 | 2 | 6 | 1 | 0 | |
| CsLEA17 | 3 | 9 | 3 | 0 | |
| CsLEA19 | 5 | 0 | 0 | 1 | |
| CsLEA44 | 0 | 0 | 0 | 2 | |
| LEA_3 | CsLEA1 | 4 | 2 | 0 | 2 |
| CsLEA16 | 1 | 2 | 0 | 3 | |
| CsLEA22 | 1 | 3 | 1 | 3 | |
| CsLEA18 | 8 | 3 | 3 | 0 | |
| CsLEA8 | 0 | 7 | 1 | 2 | |
| CsLEA9 | 3 | 0 | 0 | 0 | |
| LEA_4 | CsLEA5 | 1 | 5 | 4 | 1 |
| LEA_5 | CsLEA10 | 0 | 2 | 0 | 1 |
| CsLEA11 | 0 | 3 | 0 | 3 | |
| CsLEA12 | 2 | 7 | 1 | 1 | |
| CsLEA13 | 2 | 1 | 3 | 5 | |
| LEA_6 | CsLEA28 | 0 | 9 | 5 | 3 |
| SMP | CsLEA24 | 0 | 4 | 1 | 0 |
| CsLEA6 | 2 | 0 | 2 | 0 | |
| CsLEA7 | 2 | 6 | 0 | 0 | |
| CsLEA26 | 1 | 5 | 1 | 1 | |
| CsLEA33 | 0 | 5 | 1 | 4 | |
| CsLEA4 | 2 | 1 | 0 | 0 | |
| CsLEA23 | 2 | 1 | 0 | 0 | |
| CsLEA34 | 1 | 4 | 0 | 1 | |
| CsLEA42 | 0 | 3 | 1 | 2 | |
| CsLEA43 | 0 | 0 | 1 | 5 | |
| Dehydrin | CsLEA20 | 4 | 5 | 0 | 0 |
| CsLEA25 | 4 | 2 | 0 | 1 | |
| CsLEA35 | 3 | 8 | 2 | 1 | |
| CsLEA37 | 0 | 6 | 3 | 4 | |
| CsLEA38 | 2 | 10 | 2 | 3 | |
| CsLEA39 | 3 | 1 | 0 | 1 | |
| CsLEA41 | 2 | 0 | 1 | 2 | |
| CsLEA40 | 0 | 2 | 0 | 1 | |
| CsLEA42 | 0 | 3 | 1 | 2 | |
| CsLEA43 | 0 | 0 | 1 | 5 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muvunyi, B.P.; Yan, Q.; Wu, F.; Min, X.; Yan, Z.Z.; Kanzana, G.; Wang, Y.; Zhang, J. Mining Late Embryogenesis Abundant (LEA) Family Genes in Cleistogenes songorica, a Xerophyte Perennial Desert Plant. Int. J. Mol. Sci. 2018, 19, 3430. https://doi.org/10.3390/ijms19113430
Muvunyi BP, Yan Q, Wu F, Min X, Yan ZZ, Kanzana G, Wang Y, Zhang J. Mining Late Embryogenesis Abundant (LEA) Family Genes in Cleistogenes songorica, a Xerophyte Perennial Desert Plant. International Journal of Molecular Sciences. 2018; 19(11):3430. https://doi.org/10.3390/ijms19113430
Chicago/Turabian StyleMuvunyi, Blaise Pascal, Qi Yan, Fan Wu, Xueyang Min, Zhuan Zhuan Yan, Gisele Kanzana, Yanrong Wang, and Jiyu Zhang. 2018. "Mining Late Embryogenesis Abundant (LEA) Family Genes in Cleistogenes songorica, a Xerophyte Perennial Desert Plant" International Journal of Molecular Sciences 19, no. 11: 3430. https://doi.org/10.3390/ijms19113430