C-C Motif Ligand 20 (CCL20) and C-C Motif Chemokine Receptor 6 (CCR6) in Human Peripheral Blood Mononuclear Cells: Dysregulated in Ulcerative Colitis and a Potential Role for CCL20 in IL-1β Release

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Patient Characteristics
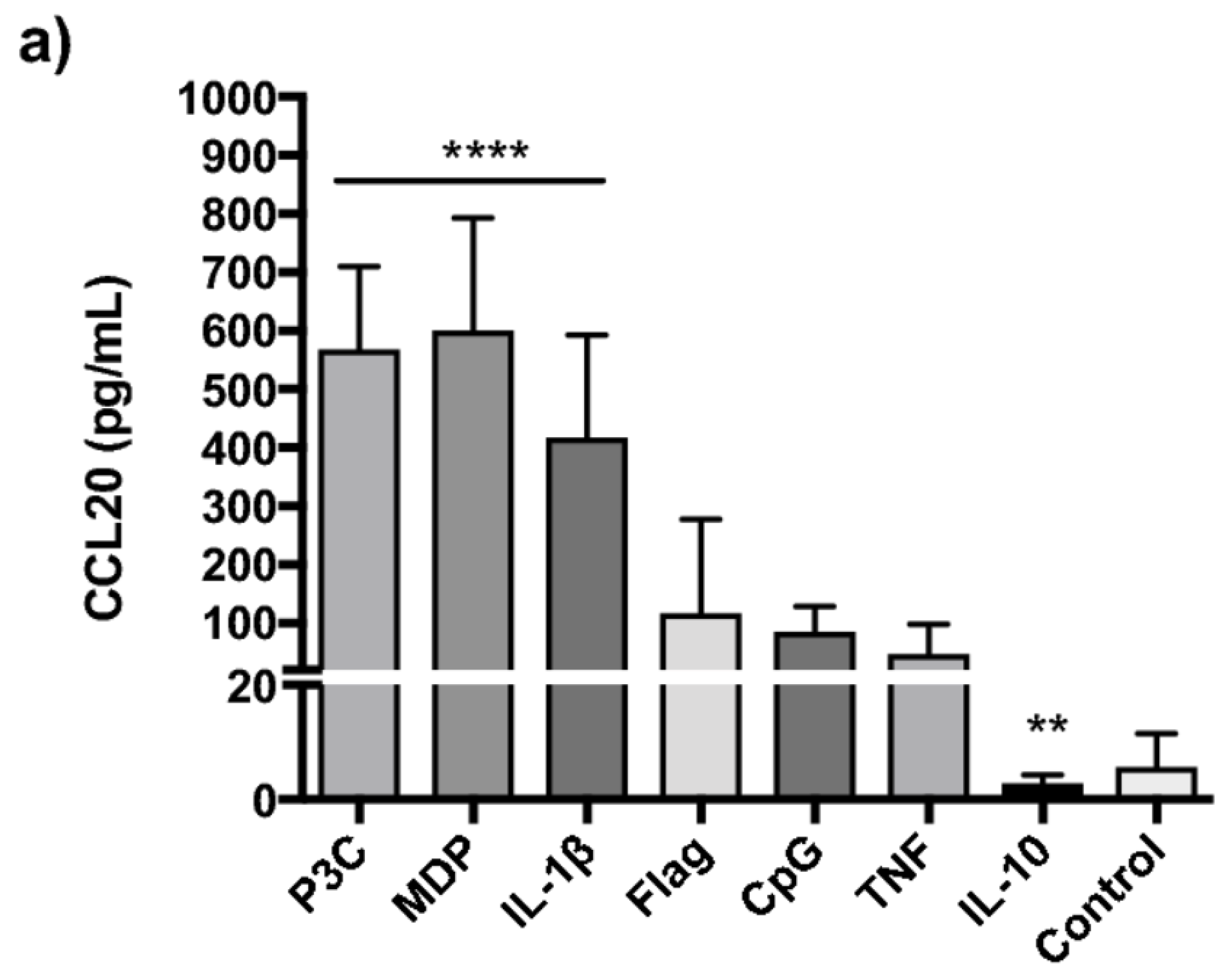
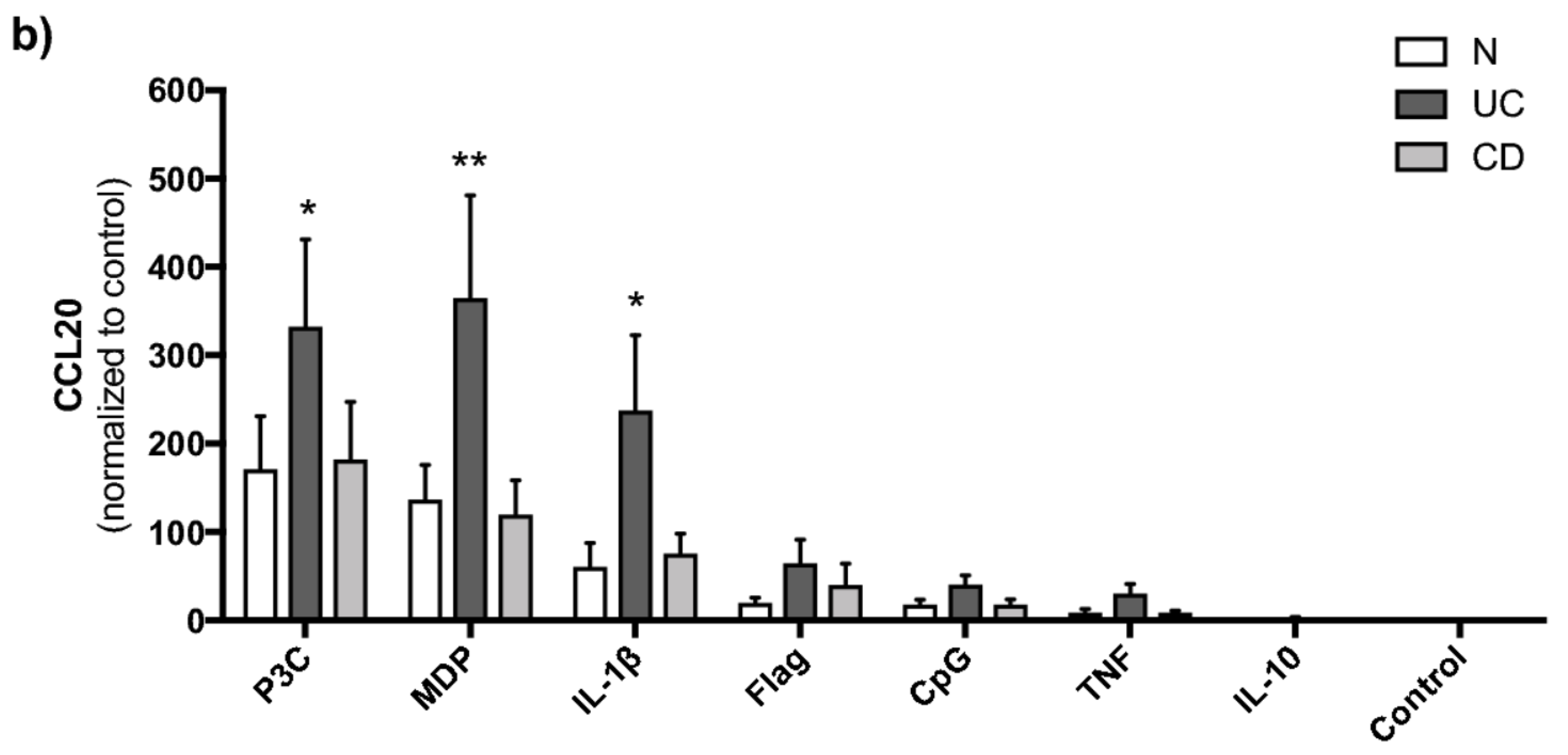
2.2. TLR2/1, NOD2 and IL-1β Strongly Induce CCL20 in PBMCs
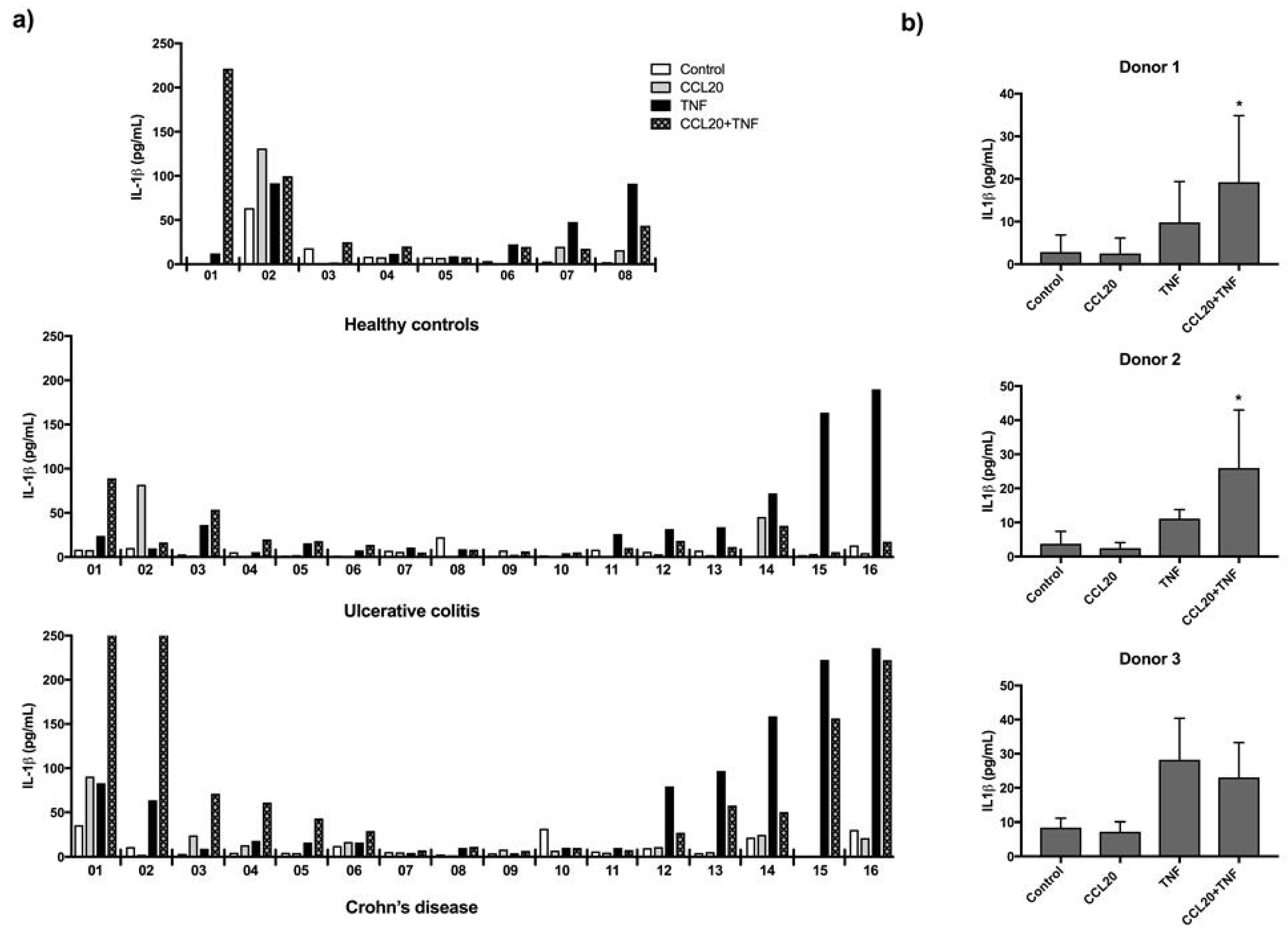
2.3. CCL20 Can Increase TNF Induced IL-1β Release
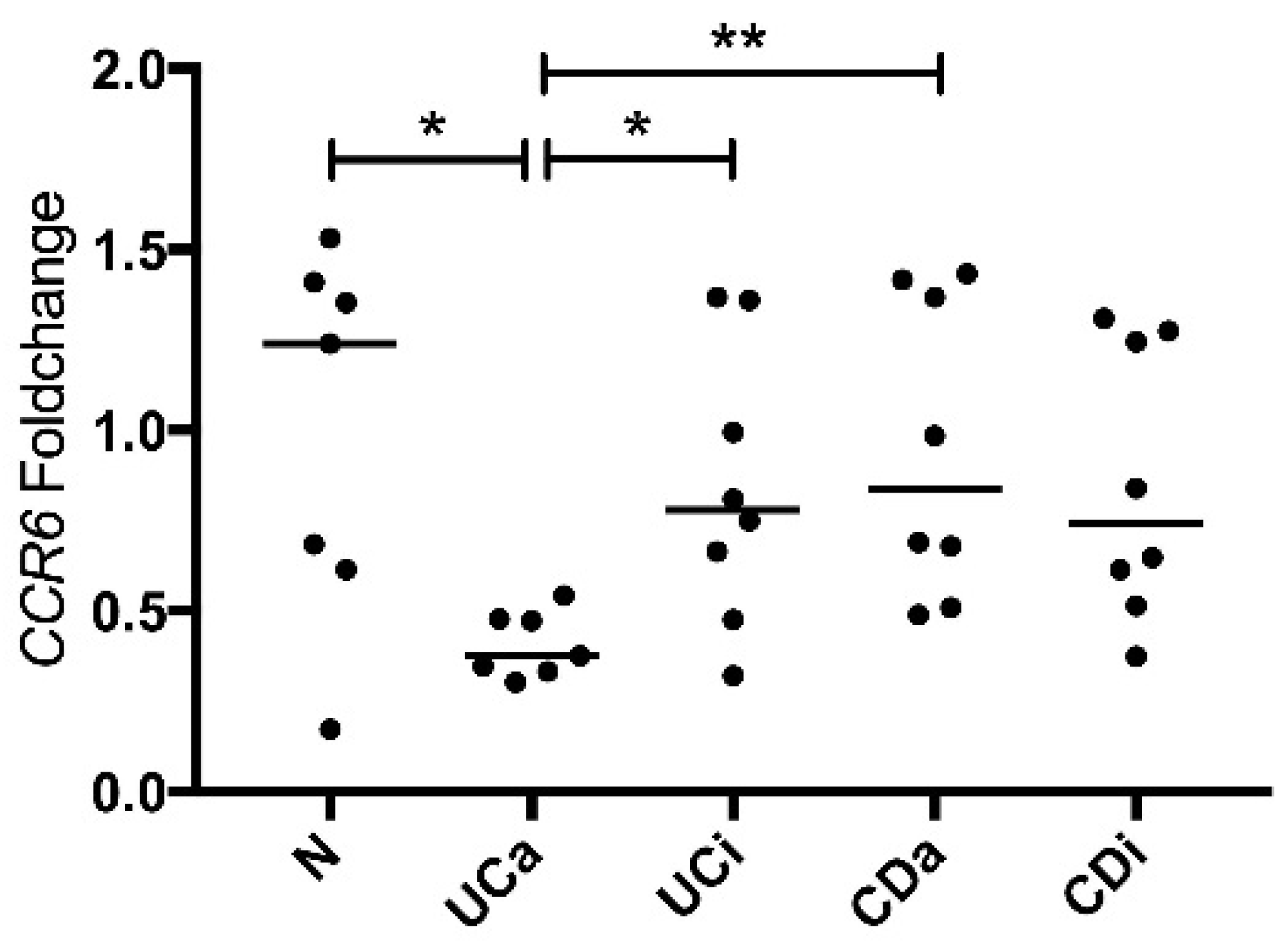
2.4. CCR6 Gene Expression Is Downregulated in PBMCs from Active Ulcerative Colitis
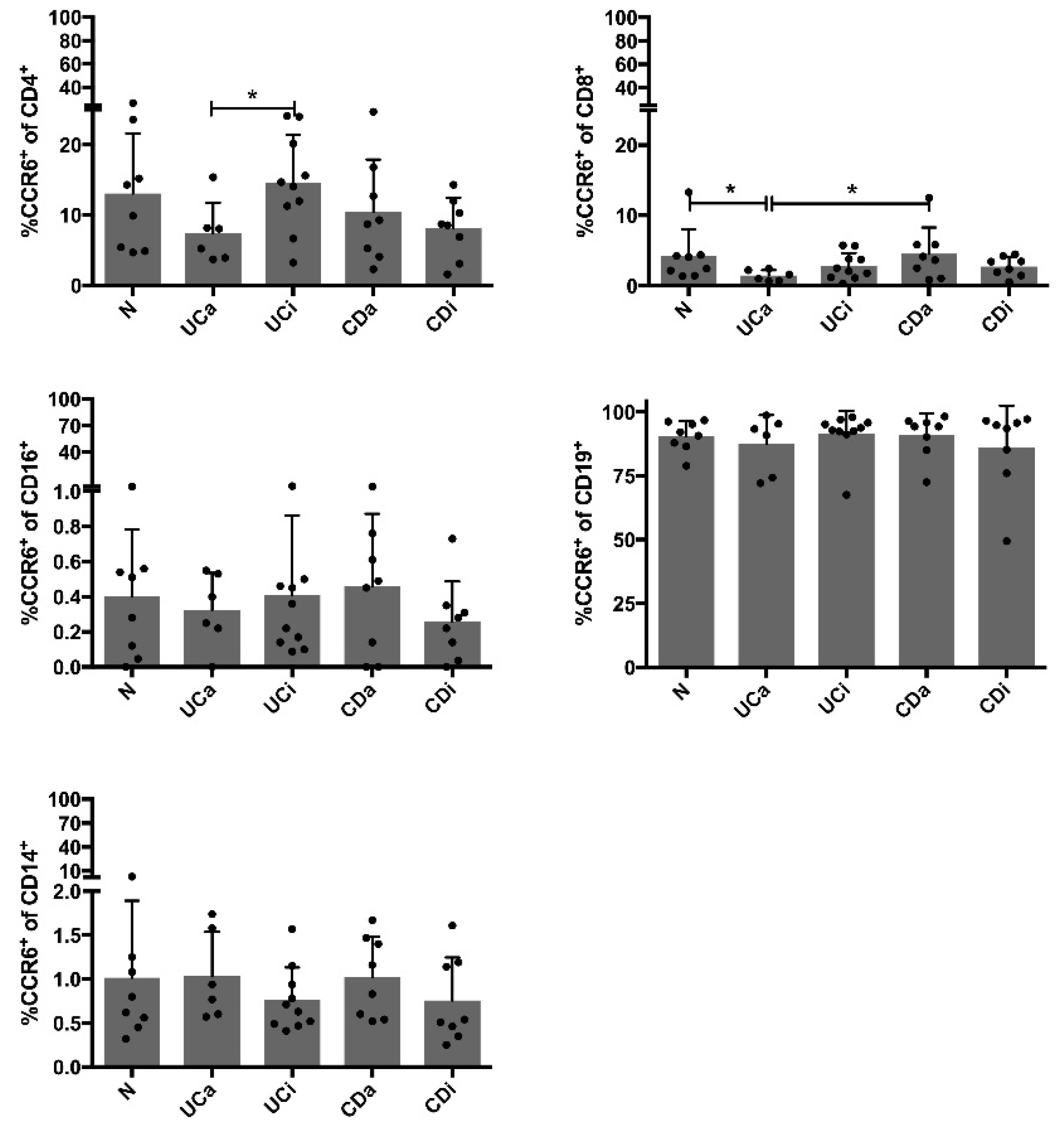
2.5. CCR6+CD4+ Cells among PBMCs Are Decreased in Active Ulcerative Colitis
3. Discussion
4. Materials and Methods
4.1. Patient Material
4.2. Ethical Considerations
4.3. Sample Preparation
4.4. Gene Expression Analyses for CCL20 and CCR6 in PBMCs
4.5. Flow Cytometry
4.6. PBMC Stimulation Assays
4.7. Measurements in Supernatant
4.8. Statistical Analysis
4.9. Data Availability
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 5-ASA | 5-aminosalicylic acid |
| S-ASA | sulphasalazine |
| CCL20 | C–C motif chemokine ligand 20 |
| CCR6 | C–C motif chemokine receptor 6 |
| CD | Crohn’s disease |
| ELISA | enzyme-linked immunosorbent assay |
| hsCRP | High sensitivity C-reactive protein |
| IBD | inflammatory bowel disease |
| IL-1β | interleukin 1β |
| IL-10 | interleukin 10 |
| mRNA | messenger ribonucleic acid |
| MDP | muramyl dipeptide |
| P3C | pam3CysSK4 |
| PRR | pattern recognition receptor |
| PBMCs | Peripheral blood mononuclear cells |
| qRT-PCR | quantitative real time polymerase chain reaction |
| TLR | toll-like receptor |
| TNF | Tumour necrosis factor |
| UC | ulcerative colitis |
References
- Ni, J.; Wu, G.D.; Albenberg, L.; Tomov, V.T. Gut microbiota and IBD: Causation or correlation? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Mazmanian, S.K. Has the Microbiota Played a Critical Role in the Evolution of the Adaptive Immune System? Science 2010, 330, 1768–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mowat, A.M. Anatomical basis of tolerance and immunity to intestinal antigens. Nat. Rev. Immunol. 2003, 3, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, N.; Pathak, M.; Lal, G. Role of chemokine receptors and intestinal epithelial cells in the mucosal inflammation and tolerance. J. Leukoc. Biol. 2017, 101, 377–394. [Google Scholar] [CrossRef] [PubMed]
- Koelink, P.J.; Overbeek, S.A.; Braber, S.; de Kruijf, P.; Folkerts, G.; Smit, M.J.; Kraneveld, A.D. Targeting chemokine receptors in chronic inflammatory diseases: An extensive review. Pharmacol. Ther. 2012, 133, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys. Acta 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [PubMed]
- Baba, M.; Imai, T.; Nishimura, M.; Kakizaki, M.; Takagi, S.; Hieshima, K.; Nomiyama, H.; Yoshie, O. Identification of CCR6, the specific receptor for a novel lymphocyte-directed CC chemokine LARC. J. Biochem. Physiol. 1997, 272, 14893–14898. [Google Scholar] [CrossRef]
- Greaves, D.R.; Wang, W.; Dairaghi, D.J.; Dieu, M.C.; Saint-Vis, B.; Franz-Bacon, K.; Rossi, D.; Caux, C.; McClanahan, T.; Gordon, S.; et al. CCR6, a CC chemokine receptor that interacts with macrophage inflammatory protein 3alpha and is highly expressed in human dendritic cells. J. Exp. Med. 1997, 186, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.N.; Prosser, D.M.; Forster, R.; Zhang, J.; Kuklin, N.A.; Abbondanzo, S.J.; Niu, X.-D.; Chen, S.-C.; Manfra, D.J.; Wiekowski, M.T.; et al. CCR6 mediates dendritic cell localization, lymphocyte homeostasis, and immune responses in mucosal tissue. Immunity 2000, 12, 495–503. [Google Scholar] [CrossRef]
- Dieu-Nosjean, M.C.; Massacrier, C.; Homey, B.; Vanbervliet, B.; Pin, J.J.; Vicari, A.; Lebecque, S.; Dezutter-Dambuyant, C.; Schmitt, D.; Zlotnik, A.; et al. Macrophage inflammatory protein 3α is expressed at inflamed epithelial surfaces and is the most potent chemokine known in attracting Langerhans cell precursors. J. Exp. Med. 2000, 192, 705–718. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Kelsall, B.L. Localization of distinct Peyer’s patch dendritic cell subsets and their recruitment by chemokines macrophage inflammatory protein (MIP)-3α, MIP-3β, and secondary lymphoid organ chemokine. J. Exp. Med. 2000, 191, 1381–1394. [Google Scholar] [CrossRef] [PubMed]
- Dieu, M.C.; Vanbervliet, B.; Vicari, A.; Bridon, J.M.; Oldham, E.; Ait-Yahia, S.; Brière, F.; Zlotnik, A.; Lebecque, S.; Caux, C. Selective recruitment of immature and mature dendritic cells by distinct chemokines expressed in different anatomic sites. J. Exp. Med. 1998, 188, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Laumonnier, Y.; Syrovets, T.; Simmet, T. Recruitment of CCR6-expressing Th17 cells by CCL20 secreted from plasmin-stimulated macrophages. Acta Biochim. Biophys. Sin. (Shanghai) 2013, 45, 593–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comerford, I.; Bunting, M.; Fenix, K.; Haylock-Jacobs, S.; Litchfield, W.; Harata-Lee, Y.; Turvey, M.; Brazzatti, J.; Gregor, C.; Nguyen, P.; et al. An immune paradox: How can the same chemokine axis regulate both immune tolerance and activation? CCR6/CCL20: A chemokine axis balancing immunological tolerance and inflammation in autoimmune disease. Bioessays 2010, 32, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, T.; Yang, X.O.; Chung, Y.; Fukunaga, A.; Nurieva, R.; Pappu, B.; Martin-Orozco, N.; Kang, H.S.; Ma, L.; Panopoulos, A.D.; et al. CCR6 regulates the migration of inflammatory and regulatory T cells. J. Immunol. 2008, 181, 8391–8401. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.H.; Keates, S.; Bassani, L.; Mayer, L.F.; Keates, A.C. Colonic epithelial cells are a major site of macrophage inflammatory protein 3α (MIP-3α) production in normal colon and inflammatory bowel disease. Gut 2002, 51, 818–826. [Google Scholar] [CrossRef] [PubMed]
- Kaser, A.; Ludwiczek, O.; Holzmann, S.; Moschen, A.R.; Weiss, G.; Enrich, B.; Graziadei, I.; Dunzendorfer, S.; Wiedermann, C.J.; Mürzl, E.; et al. Increased expression of CCL20 in human inflammatory bowel disease. J. Clin. Immunol. 2004, 24, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Skovdahl, H.K.; Granlund, A.; Ostvik, A.E.; Bruland, T.; Bakke, I.; Torp, S.H.; Damås, J.K.; Sandvik, A.K. Expression of CCL20 and Its Corresponding Receptor CCR6 Is Enhanced in Active Inflammatory Bowel Disease, and TLR3 Mediates CCL20 Expression in Colonic Epithelial Cells. PLoS ONE 2015, 10, e0141710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.Z.; van Sommeren, S.; Huang, H.; Ng, S.C.; Alberts, R.; Takahashi, A.; Ripke, S.; Lee, J.C.; Jostins, L.; Shah, T.; et al. Association analyses identify 38 susceptibility loci for inflammatory bowel disease and highlight shared genetic risk across populations. Nat. Genet. 2015, 47, 979–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackay, C.R. Moving targets: Cell migration inhibitors as new anti-inflammatory therapies. Nat. Immunol. 2008, 9, 988–998. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Choi, S.C.; Lee, M.H.; Oh, H.M.; Choi, E.Y.; Choi, E.J.; Yun, K.-J.; Seo, G.-S.; Kim, S.-W.; Lee, J.-G.; et al. Increased expression of MIP-3α/CCL20 in peripheral blood mononuclear cells from patients with ulcerative colitis and its down-regulation by sulfasalazine and glucocorticoid treatment. Inflamm. Bowel Dis. 2005, 11, 1070–1079. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Zhang, S.L.; Hu, C.J.; Tong, D.W.; Li, Y.Z. Higher levels of CCL20 expression on peripheral blood mononuclear cells of chinese patients with inflammatory bowel disease. Immunol. Investig. 2010, 39, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Spagnulolo, R.; Dattilo, V.; D’Antona, L.; Cosco, C.; Tallerico, R.; Ventura, V.; Conforti, F.; Camastra, C.; Mancina, R.M.; Catalogna, G.; et al. Deregulation of SGK1 in Ulcerative Colitis: A Paradoxical Relationship between Immune Cells and Colonic Epithelial Cells. Inflamm. Bowel Dis. 2018, 24, 1967–1977. [Google Scholar] [CrossRef] [PubMed]
- Cayatte, C.; Joyce-Shaikh, B.; Vega, F.; Boniface, K.; Grein, J.; Murphy, E.; Blumenschein, W.M.; Chen, S.; Malinao, M.-C.; Basham, B.; et al. Biomarkers of Therapeutic Response in the IL-23 Pathway in Inflammatory Bowel Disease. Clin. Transl. Gastroenterol. 2012, 3, e10. [Google Scholar] [CrossRef] [PubMed]
- Levine, J.S.; Burakoff, R. Extraintestinal manifestations of inflammatory bowel disease. Gastroenterol. Hepatol. (N. Y.) 2011, 7, 235–241. [Google Scholar]
- Hoffmann, R.M.; Kruis, W. Rare extraintestinal manifestations of inflammatory bowel disease. Inflamm. Bowel Dis. 2004, 10, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Mesko, B.; Poliska, S.; Szegedi, A.; Szekanecz, Z.; Palatka, K.; Papp, M.; Nagy, L. Peripheral blood gene expression patterns discriminate among chronic inflammatory diseases and healthy controls and identify novel targets. BMC Med. Genom. 2010, 3, 15. [Google Scholar] [CrossRef] [PubMed]
- Granlund, A.; Flatberg, A.; Ostvik, A.E.; Drozdov, I.; Gustafsson, B.I.; Kidd, M.; Beisvag, V.; Torp, S.H.; Waldum, H.L.; Martinsen, T.C.; et al. Whole genome gene expression meta-analysis of inflammatory bowel disease colon mucosa demonstrates lack of major differences between Crohn’s disease and ulcerative colitis. PLoS ONE 2013, 8, e56818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hausmann, M.; Zeitler, C.; Weber, A.; Krebs, M.; Kellermeier, S.; Rosenstiel, P.; de Vallière, C.; Kosovac, K.; Fried, M.; Holler, E.; et al. MIP-3alpha expression in macrophages is NOD dependent. Digestion 2012, 85, 192–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokol, C.L.; Luster, A.D. The chemokine system in innate immunity. Cold Spring Harb. Perspect. Biol. 2015, 7, a016303. [Google Scholar] [CrossRef] [PubMed]
- Krzysiek, R.; Lefevre, E.A.; Bernard, J.; Foussat, A.; Galanaud, P.; Louache, F.; Richard, Y. Regulation of CCR6 chemokine receptor expression and responsiveness to macrophage inflammatory protein-3alpha/CCL20 in human B cells. Blood 2000, 96, 2338–2345. [Google Scholar] [PubMed]
- Liao, F.; Rabin, R.L.; Smith, C.S.; Sharma, G.; Nutman, T.B.; Farber, J.M. CC-chemokine receptor 6 is expressed on diverse memory subsets of T cells and determines responsiveness to macrophage inflammatory protein 3 α. J. Immunol. 1999, 162, 186–194. [Google Scholar] [PubMed]
- Omenetti, S.; Pizarro, T.T. The Treg/Th17 Axis: A Dynamic Balance Regulated by the Gut Microbiome. Front. Immunol. 2015, 6, 639. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.J.; Ebsworth, K.; Ertl, L.S.; McMahon, J.P.; Newland, D.; Wang, Y.; Liu, S.; Miao, Z.; Dang, T.; Zhang, P.; et al. IL-17-Secreting gammadelta T Cells Are Completely Dependent upon CCR6 for Homing to Inflamed Skin. J. Immunol. 2017, 199, 3129–3136. [Google Scholar] [CrossRef] [PubMed]
- Alaaeddine, N.; Antoniou, J.; Moussa, M.; Hilal, G.; Kreichaty, G.; Ghanem, I.; Abouchedid, W.; Saghbini, E.; Di Battista, J.A. The chemokine CCL20 induces proinflammatory and matrix degradative responses in cartilage. Inflamm. Res. 2015, 64, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.C.; Lee, E.K.; Lee, S.; Chae, S.C.; Lee, M.S.; Seo, G.S.; Kim, S.W.; Yeom, J.J.; Jun, C.D. Ulcerative Colitis is Associated with Novel Polymorphisms in the Promoter Region of MIP-3α/CCL20 Gene. Immune Netw. 2005, 5, 205–214. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | N | UC | CD |
|---|---|---|---|
| Number | 8 | 16 | 16 |
| Female sex | 5 | 8 | 7 |
| Age mean (range) | 43 (22–68) | 45 (19–76) | 40 (24–57) |
| 5-ASA/S-ASA | 0 | 13 | 4 |
| Corticosteroids | 0 | 1 | 1 |
| Mayo/HBI < 1 | - | 10 | 9 |
| Mayo/HBI ≥ 1 | - | 6 | 7 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skovdahl, H.K.; Damås, J.K.; Granlund, A.V.B.; Østvik, A.E.; Doseth, B.; Bruland, T.; Mollnes, T.E.; Sandvik, A.K. C-C Motif Ligand 20 (CCL20) and C-C Motif Chemokine Receptor 6 (CCR6) in Human Peripheral Blood Mononuclear Cells: Dysregulated in Ulcerative Colitis and a Potential Role for CCL20 in IL-1β Release. Int. J. Mol. Sci. 2018, 19, 3257. https://doi.org/10.3390/ijms19103257
Skovdahl HK, Damås JK, Granlund AVB, Østvik AE, Doseth B, Bruland T, Mollnes TE, Sandvik AK. C-C Motif Ligand 20 (CCL20) and C-C Motif Chemokine Receptor 6 (CCR6) in Human Peripheral Blood Mononuclear Cells: Dysregulated in Ulcerative Colitis and a Potential Role for CCL20 in IL-1β Release. International Journal of Molecular Sciences. 2018; 19(10):3257. https://doi.org/10.3390/ijms19103257
Chicago/Turabian StyleSkovdahl, Helene Kolstad, Jan Kristian Damås, Atle Van Beelen Granlund, Ann Elisabet Østvik, Berit Doseth, Torunn Bruland, Tom Eirik Mollnes, and Arne Kristian Sandvik. 2018. "C-C Motif Ligand 20 (CCL20) and C-C Motif Chemokine Receptor 6 (CCR6) in Human Peripheral Blood Mononuclear Cells: Dysregulated in Ulcerative Colitis and a Potential Role for CCL20 in IL-1β Release" International Journal of Molecular Sciences 19, no. 10: 3257. https://doi.org/10.3390/ijms19103257