Enhancement of Radiation Effectiveness in Cervical Cancer Cells by Combining Ionizing Radiation with Hyperthermia and Molecular Targeting Agents

 , ,
, ,  and
and
Abstract
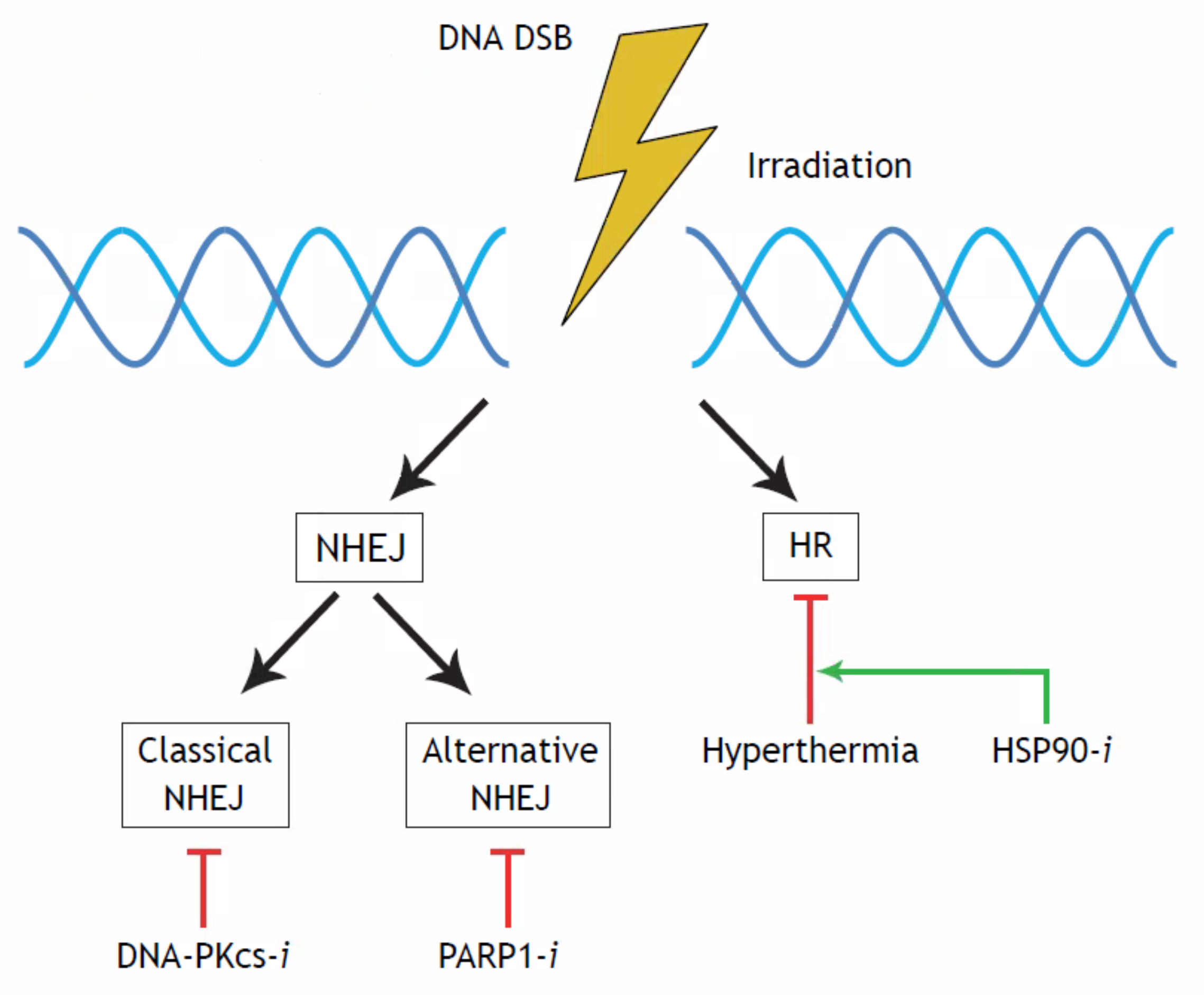
:1. Introduction
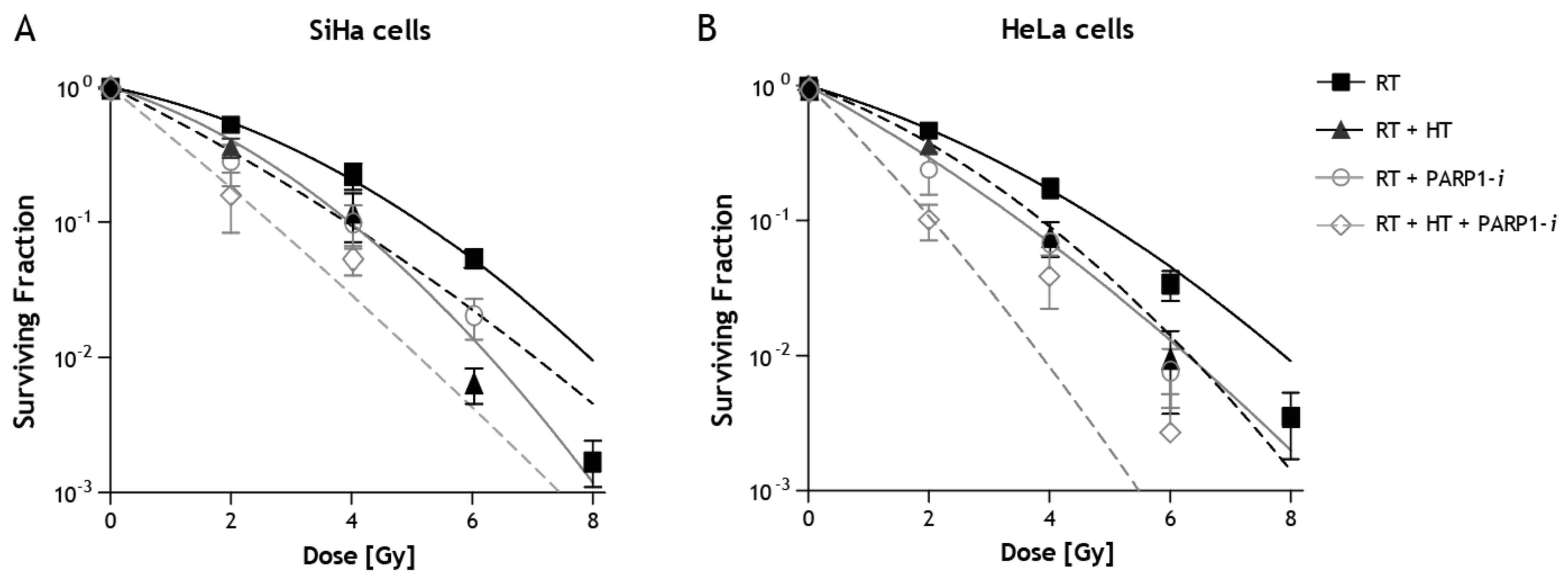
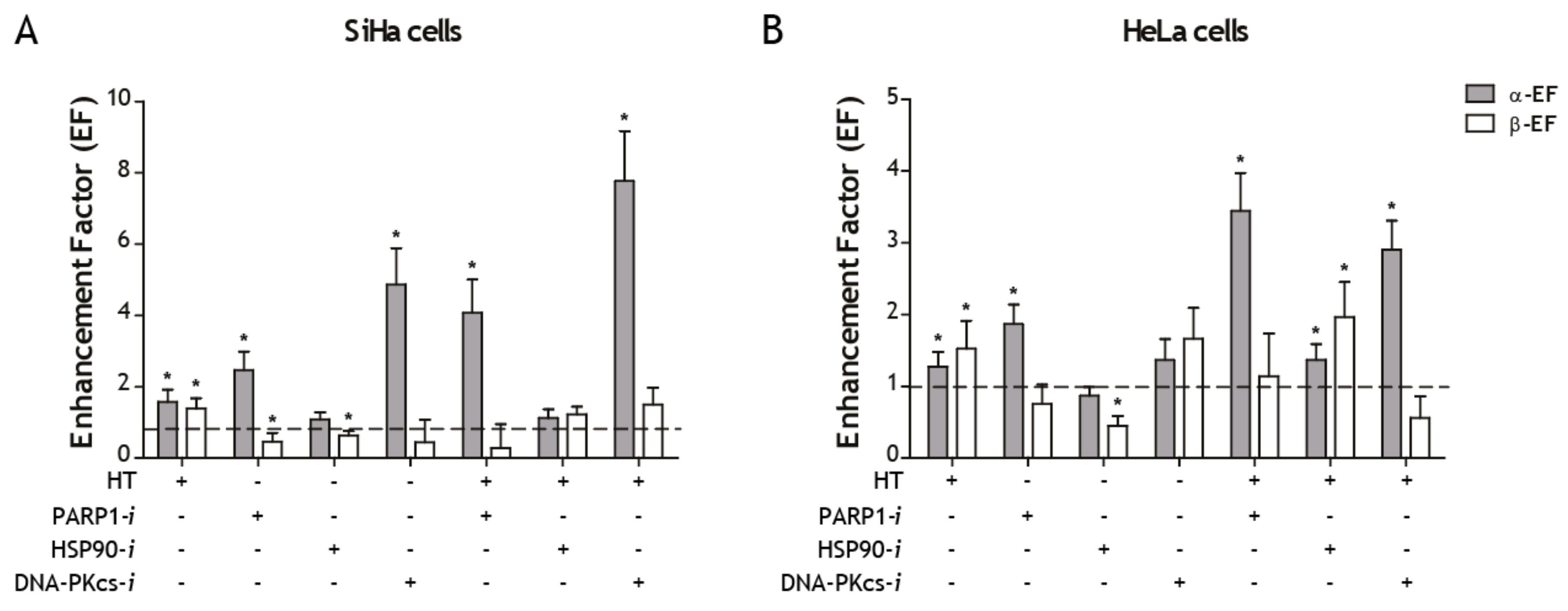
2. Results
3. Discussion
4. Methods
4.1. Linear-Quadratic (LQ) Model
4.2. Cell Cultures
4.3. Irradiation
4.4. Hyperthermia
4.5. Molecular Targeting Agents
4.6. Clonogenic Assay
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Cervical Cancer—Early Diagnosis and Screening; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Rose, P.G.; Bundy, B.N.; Watkins, E.B.; Thigpen, J.T.; Deppe, G.; Maiman, M.A.; Clarke-Pearson, D.L.; Insalaco, S. Concurrent cisplatin-based radiotherapy and chemotherapy for locally advanced cervical cancer. N. Engl. J. Med. 1999, 340, 1144–1153. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.; Eifel, P.J.; Lu, J.; Grigsby, P.W.; Levenback, C.; Stevens, R.E.; Rotman, M.; Gershenson, D.M.; Mutch, D.G. Pelvic radiation with concurrent chemotherapy compared with pelvic and para-aortic radiation for high-risk cervical cancer. N. Engl. J. Med. 1999, 340, 1137–1143. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wu, X.; Cheng, X. Advances in diagnosis and treatment of metastatic cervical cancer. J. Gynecol. Oncol. 2016, 27, e43. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.Y.; Yang, L.; Yang, G.; Tu, X.Y.; Wu, X.; Cheng, X.; Wei, Q. DNA polymerase zeta as a potential biomarker of chemoradiation resistance and poor prognosis for cervical cancer. Med. Oncol. 2013, 30, 500. [Google Scholar] [CrossRef] [PubMed]
- Van der Zee, J.; van Rhoon, G.C. Cervical cancer: Radiotherapy and hyperthermia. Int. J. Hyperth. 2006, 22, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Linthorst, M.; van Geel, A.N.; Baaijens, M.; Ameziane, A.; Ghidey, W.; van Rhoon, G.C.; van der Zee, J. Re-irradiation and hyperthermia after surgery for recurrent breast cancer. Radiother. Oncol. 2013, 109, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Bergs, J.W.; Franken, N.A.; Haveman, J.; Geijsen, E.D.; Crezee, J.; van Bree, C. Hyperthermia, cisplatin and radiation trimodality treatment: A promising cancer treatment? A review from preclinical studies to clinical application. Int. J. Hyperth. 2007, 23, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Lutgens, L.C.; Koper, P.C.; Jobsen, J.J.; van der Steen-Banasik, E.M.; Creutzberg, C.L.; van den Berg, H.A.; Ottevanger, P.B.; van Rhoon, G.C.; van Doorn, H.C.; Houben, R.; et al. Radiation therapy combined with hyperthermia versus cisplatin for locally advanced cervical cancer: Results of the randomized radchoc trial. Radiother. Oncol. 2016, 120, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Dietlein, F.; Thelen, L.; Reinhardt, H.C. Cancer-specific defects in DNA repair pathways as targets for personalized therapeutic approaches. Trends Genet. 2014, 30, 326–339. [Google Scholar] [CrossRef] [PubMed]
- Krajewska, M.; Fehrmann, R.S.; de Vries, E.G.; van Vugt, M.A. Regulators of homologous recombination repair as novel targets for cancer treatment. Front. Genet. 2015, 6, 96. [Google Scholar] [CrossRef] [PubMed]
- Iliakis, G.; Murmann, T.; Soni, A. Alternative end-joining repair pathways are the ultimate backup for abrogated classical non-homologous end-joining and homologous recombination repair: Implications for the formation of chromosome translocations. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2015, 793, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Kotter, A.; Cornils, K.; Borgmann, K.; Dahm-Daphi, J.; Petersen, C.; Dikomey, E.; Mansour, W.Y. Inhibition of PARP1-dependent end-joining contributes to Olaparib-mediated radiosensitization in tumor cells. Mol. Oncol. 2014, 8, 1616–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Oorschot, B.; Granata, G.; Di Franco, S.; Ten Cate, R.; Rodermond, H.M.; Todaro, M.; Medema, J.P.; Franken, N.A. Targeting DNA double strand break repair with hyperthermia and DNA-PKcs inhibition to enhance the effect of radiation treatment. Oncotarget 2016, 7, 65504–65513. [Google Scholar] [CrossRef] [PubMed]
- Curtin, N.J. Inhibiting the DNA damage response as a therapeutic manoeuvre in cancer. Br. J. Pharmacol. 2013, 169, 1745–1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oei, A.L.; Ahire, V.R.; van Leeuwen, C.M.; Ten Cate, R.; Stalpers, L.J.A.; Crezee, J.; Kok, H.P.; Franken, N.A.P. Enhancing radiosensitisation of BRCA2-proficient and BRCA2-deficient cell lines with hyperthermia and PARP1-i. Int. J. Hyperth. 2018, 34, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, P.M.; Eppink, B.; Essers, J.; Stap, J.; Rodermond, H.; Odijk, H.; Zelensky, A.; van Bree, C.; Stalpers, L.J.; Buist, M.R.; et al. Mild hyperthermia inhibits homologous recombination, induces BRCA2 degradation, and sensitizes cancer cells to poly (ADP-ribose) polymerase-1 inhibition. Proc. Natl. Acad. Sci. USA 2011, 108, 9851–9856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oei, A.L.; Vriend, L.E.; Crezee, J.; Franken, N.A.; Krawczyk, P.M. Effects of hyperthermia on DNA repair pathways: One treatment to inhibit them all. Radiat. Oncol. 2015, 10, 165. [Google Scholar] [CrossRef] [PubMed]
- Ihara, M.; Takeshita, S.; Okaichi, K.; Okumura, Y.; Ohnishi, T. Heat exposure enhances radiosensitivity by depressing DNA-PK kinase activity during double strand break repair. Int. J. Hyperth. 2014, 30, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Uzui, K.; Nishi, R.; Shigemi, H.; Ueda, T. Gemtuzumab ozogamicin and olaparib exert synergistic cytotoxicity in CD33-positive HL-60 myeloid leukemia cells. Anticancer Res. 2014, 34, 5487–5494. [Google Scholar] [PubMed]
- Deriano, L.; Roth, D.B. Modernizing the nonhomologous end-joining repertoire: Alternative and classical NHEJ share the stage. Annu. Rev. Genet. 2013, 47, 433–455. [Google Scholar] [CrossRef] [PubMed]
- Newman, E.A.; Lu, F.; Bashllari, D.; Wang, L.; Opipari, A.W.; Castle, V.P. Alternative NHEJ pathway components are therapeutic targets in high-risk neuroblastoma. Mol. Cancer Res. 2015, 13, 470–482. [Google Scholar] [CrossRef] [PubMed]
- Simsek, D.; Brunet, E.; Wong, S.Y.; Katyal, S.; Gao, Y.; McKinnon, P.J.; Lou, J.; Zhang, L.; Li, J.; Rebar, E.J.; et al. DNA ligase III promotes alternative nonhomologous end-joining during chromosomal translocation formation. PLoS Genet. 2011, 7, e1002080. [Google Scholar] [CrossRef] [PubMed]
- Jasin, M.; Haber, J.E. The democratization of gene editing: Insights from site-specific cleavage and double-strand break repair. DNA Repair 2016, 44, 6–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderwood, S.K.; Ciocca, D.R. Heat shock proteins: Stress proteins with Janus-like properties in cancer. Int. J. Hyperth. 2008, 24, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, R.; Ascenzi, P.; di Masi, A. Hsp90: A new player in DNA repair? Biomolecules 2015, 5, 2589–2618. [Google Scholar] [CrossRef] [PubMed]
- Vriend, L.E.M.; van den Tempel, N.; Oei, A.L.; L’Acosta, M.; Pieterson, F.J.; Franken, N.A.P.; Kanaar, R.; Krawczyk, P.M. Boosting the effects of hyperthermia-based anticancer treatments by HSP90 inhibition. Oncotarget 2017, 8, 97490–97503. [Google Scholar] [CrossRef] [PubMed]
- Barendsen, G.W.; Van Bree, C.; Franken, N.A. Importance of cell proliferative state and potentially lethal damage repair on radiation effectiveness: Implications for combined tumor treatments (review). Int. J. Oncol. 2001, 19, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Franken, N.A.; Oei, A.L.; Kok, H.P.; Rodermond, H.M.; Sminia, P.; Crezee, J.; Stalpers, L.J.; Barendsen, G.W. Cell survival and radiosensitisation: Modulation of the linear and quadratic parameters of the lq model (review). Int. J. Oncol. 2013, 42, 1501–1515. [Google Scholar] [CrossRef] [PubMed]
- Barendsen, G.W. Dose fractionation, dose rate and iso-effect relationships for normal tissue responses. Int. J. Radiat. Oncol. Biol. Phys. 1982, 8, 1981–1997. [Google Scholar] [CrossRef]
- Franken, N.A.; Kok, H.P.; Crezee, J.; Barendsen, G.W. Analysis of enhancement at small and large radiation doses for effectiveness of inactivation in cultured cells by combining two agents with radiation. Int. J. Radiat. Biol. 2016, 92, 521–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franken, N.A.; Barendsen, G.W. Enhancement of radiation effectiveness by hyperthermia and incorporation of halogenated pyrimidines at low radiation doses as compared with high doses: Implications for mechanisms. Int. J. Radiat. Biol. 2014, 90, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, C.M.; Oei, A.L.; Ten Cate, R.; Franken, N.A.P.; Bel, A.; Stalpers, L.J.A.; Crezee, J.; Kok, H.P. Measurement and analysis of the impact of time-interval, temperature and radiation dose on tumour cell survival and its application in thermoradiotherapy plan evaluation. Int. J. Hyperth. 2018, 34, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Mansour, W.Y.; Borgmann, K.; Petersen, C.; Dikomey, E.; Dahm-Daphi, J. The absence of ku but not defects in classical non-homologous end-joining is required to trigger PARP1-dependent end-joining. DNA Repair 2013, 12, 1134–1142. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.G.; Sarkaria, J.N.; Kaufmann, S.H. Nonhomologous end joining drives poly(ADP-ribose) polymerase (PARP) inhibitor lethality in homologous recombination-deficient cells. Proc. Natl. Acad. Sci. USA 2011, 108, 3406–3411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Zee, J.; Gonzalez Gonzalez, D.; van Rhoon, G.C.; van Dijk, J.D.; van Putten, W.L.; Hart, A.A. Comparison of radiotherapy alone with radiotherapy plus hyperthermia in locally advanced pelvic tumours: A prospective, randomised, multicentre trial. Dutch deep hyperthermia group. Lancet 2000, 355, 1119–1125. [Google Scholar] [CrossRef]
- Westermann, A.; Mella, O.; Van Der Zee, J.; Jones, E.L.; Van Der Steen-Banasik, E.; Koper, P.; Uitterhoeve, A.L.; De Wit, R.; Van Der Velden, J.; Burger, C.; et al. Long-term survival data of triple modality treatment of stage IIB-III-IVA cervical cancer with the combination of radiotherapy, chemotherapy and hyperthermia—An update. Int. J. Hyperth. 2012, 28, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Ohguri, T.; Harima, Y.; Imada, H.; Sakurai, H.; Ohno, T.; Hiraki, Y.; Tuji, K.; Tanaka, M.; Terashima, H. Relationships between thermal dose parameters and the efficacy of definitive chemoradiotherapy plus regional hyperthermia in the treatment of locally advanced cervical cancer: Data from a multicentre randomised clinical trial. Int. J. Hyperth. 2018, 34, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Feng, F.Y.; de Bono, J.S.; Rubin, M.A.; Knudsen, K.E. Chromatin to clinic: The molecular rationale for parp1 inhibitor function. Mol. Cell 2015, 58, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Thijssen, R.; Ter Burg, J.; Garrick, B.; van Bochove, G.G.; Brown, J.R.; Fernandes, S.M.; Rodriguez, M.S.; Michot, J.M.; Hallek, M.; Eichhorst, B.; et al. Dual TORK/DNA-PK inhibition blocks critical signaling pathways in chronic lymphocytic leukemia. Blood 2016, 128, 574–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanai, M.; Makino, H.; Ping, B.; Takeda, K.; Tanaka, N.; Sakamoto, T.; Yamaguchi, K.; Kodani, M.; Yamasaki, A.; Igishi, T.; et al. DNA-PK inhibition by nu7441 enhances chemosensitivity to topoisomerase inhibitor in non-small cell lung carcinoma cells by blocking DNA damage repair. Yonago Acta Med. 2017, 60, 9–15. [Google Scholar] [PubMed]
- Van Bree, C.; Franken, N.A.; Bakker, P.J.; Klomp-Tukker, L.J.; Barendsen, G.W.; Kipp, J.B. Hyperthermia and incorporation of halogenated pyrimidines: Radiosensitization in cultured rodent and human tumor cells. Int. J. Radiat. Oncol. Biol. Phys. 1997, 39, 489–496. [Google Scholar] [CrossRef] [Green Version]
- Franken, N.A.; Rodermond, H.M.; Stap, J.; Haveman, J.; van Bree, C. Clonogenic assay of cells in vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cell Line | Sensitizing Agent | Surviving Fraction |
|---|---|---|
| SiHa | RT | 0.53 ± 0.08 |
| SiHa | RT + HT42 | 0.36 ± 0.10 |
| SiHa | RT + PARP1-i | 0.28 ± 0.09 |
| SiHa | RT + HT42 + PARP1-i | 0.16 ± 0.07 |
| SiHa | RT + HSP90-i | 0.58 ± 0.04 |
| SiHa | RT + HT42 + HSP90-i | 0.51 ± 0.05 |
| SiHa | RT + DNA-PKcs-i | 0.12 ± 0.03 |
| SiHa | RT + HT42 + DNA-PKcs-i | 0.03 ± 0.00 |
| HeLa | RT | 0.46 ± 0.03 |
| HeLa | RT + HT42 | 0.36 ± 0.09 |
| HeLa | RT + PARP1-i | 0.24 ± 0.08 |
| HeLa | RT + HT42 + PARP1-i | 0.10 ± 0.03 |
| HeLa | RT + HSP90-i | 0.56 ± 0.03 |
| HeLa | RT + HT42 + HSP90-i | 0.33 ± 0.04 |
| HeLa | RT + DNA-PKcs-i | 0.35 ± 0.06 |
| HeLa | RT + HT42 + DNA-PKcs-i | 0.16 ± 0.02 |
| Cell Line | Sensitizing Agent | α (Gy−1) | β (Gy−2) | α-EF * | β-EF * |
|---|---|---|---|---|---|
| SiHa | RT | 0.21 ± 0.04 | 0.05 ± 0.01 | 1.00 ± 0.00 | 1.00 ± 0.21 |
| SiHa | RT + HT42 | 0.32 ± 0.07 | 0.07 ± 0.01 | 1.56 ± 0.45 | 1.38 ± 0.28 |
| SiHa | RT + PARP1-i | 0.51 ± 0.05 | 0.02 ± 0.01 | 2.46 ± 0.51 | 0.45 ± 0.24 |
| SiHa | RT + HSP-i | 0.22 ± 0.02 | 0.03 ± 0.00 | 1.07 ± 0.21 | 0.62 ± 0.13 |
| SiHa | RT + DNA-PKcs-i | 1.00 ± 0.11 | 0.02 ± 0.03 | 4.85 ± 1.02 | 0.43 ± 0.64 |
| SiHa | RT + HT + PARP1-i | 0.84 ± 0.12 ** | 0.01 ± 0.03 | 4.06 ± 0.94 ** | 0.25 ± 0.68 |
| SiHa | RT + HT + HSP-i | 0.23 ± 0.03 | 0.06 ± 0.01 *** | 1.12 ± 0.24 | 1.21 ± 0.22 *** |
| SiHa | RT + HT + DNA-PKcs-i | 1.60 ± 0.02 ** | 0.07 ± 0.02 | 7.77 ± 1.40 ** | 1.49 ± 0.48 |
| HeLa | RT | 0.30 ± 0.04 | 0.04 ± 0.01 | 1.00 ± 0.18 | 1.00 ± 0.00 |
| HeLa | RT + HT42 | 0.38 ± 0.04 | 0.06 ± 0.01 | 1.27 ± 0.21 | 1.53 ± 0.39 |
| HeLa | RT + PARP1-i | 0.56 ± 0.04 | 0.03 ± 0.01 | 1.88 ± 0.27 | 0.75 ± 0.27 |
| HeLa | RT + HSP-i | 0.26 ± 0.02 | 0.02 ± 0.00 | 0.87 ± 0.12 | 0.44 ± 0.14 |
| HeLa | RT + DNA-PKcs-i | 0.41 ± 0.07 | 0.06 ± 0.01 | 1.37 ± 0.29 | 1.67 ± 0.43 |
| HeLa | RT + HT + PARP1-i | 1.03 ± 0.09 ** | 0.04 ± 0.02 | 3.46 ± 0.53 ** | 1.14 ± 0.60 |
| HeLa | RT + HT + HSP-i | 0.41 ± 0.04 *** | 0.07 ± 0.01 *** | 1.37 ± 0.22 *** | 1.97 ± 0.49 *** |
| HeLa | RT + HT + DNA-PKcs-i | 0.87 ± 0.06 ** | 0.02 ± 0.01 | 2.91 ± 0.41 ** | 0.56 ± 0.30 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
IJff, M.; Van Oorschot, B.; Oei, A.L.; Krawczyk, P.M.; Rodermond, H.M.; Stalpers, L.J.A.; Kok, H.P.; Crezee, J.; Franken, N.A.P. Enhancement of Radiation Effectiveness in Cervical Cancer Cells by Combining Ionizing Radiation with Hyperthermia and Molecular Targeting Agents. Int. J. Mol. Sci. 2018, 19, 2420. https://doi.org/10.3390/ijms19082420
IJff M, Van Oorschot B, Oei AL, Krawczyk PM, Rodermond HM, Stalpers LJA, Kok HP, Crezee J, Franken NAP. Enhancement of Radiation Effectiveness in Cervical Cancer Cells by Combining Ionizing Radiation with Hyperthermia and Molecular Targeting Agents. International Journal of Molecular Sciences. 2018; 19(8):2420. https://doi.org/10.3390/ijms19082420
Chicago/Turabian StyleIJff, Marloes, Bregje Van Oorschot, Arlene L. Oei, Przemek M. Krawczyk, Hans M. Rodermond, Lukas J. A. Stalpers, H. Petra Kok, Johannes Crezee, and Nicolaas A. P. Franken. 2018. "Enhancement of Radiation Effectiveness in Cervical Cancer Cells by Combining Ionizing Radiation with Hyperthermia and Molecular Targeting Agents" International Journal of Molecular Sciences 19, no. 8: 2420. https://doi.org/10.3390/ijms19082420