Arthroprotective Effects of Cf-02 Sharing Structural Similarity with Quercetin

 ,
,  , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
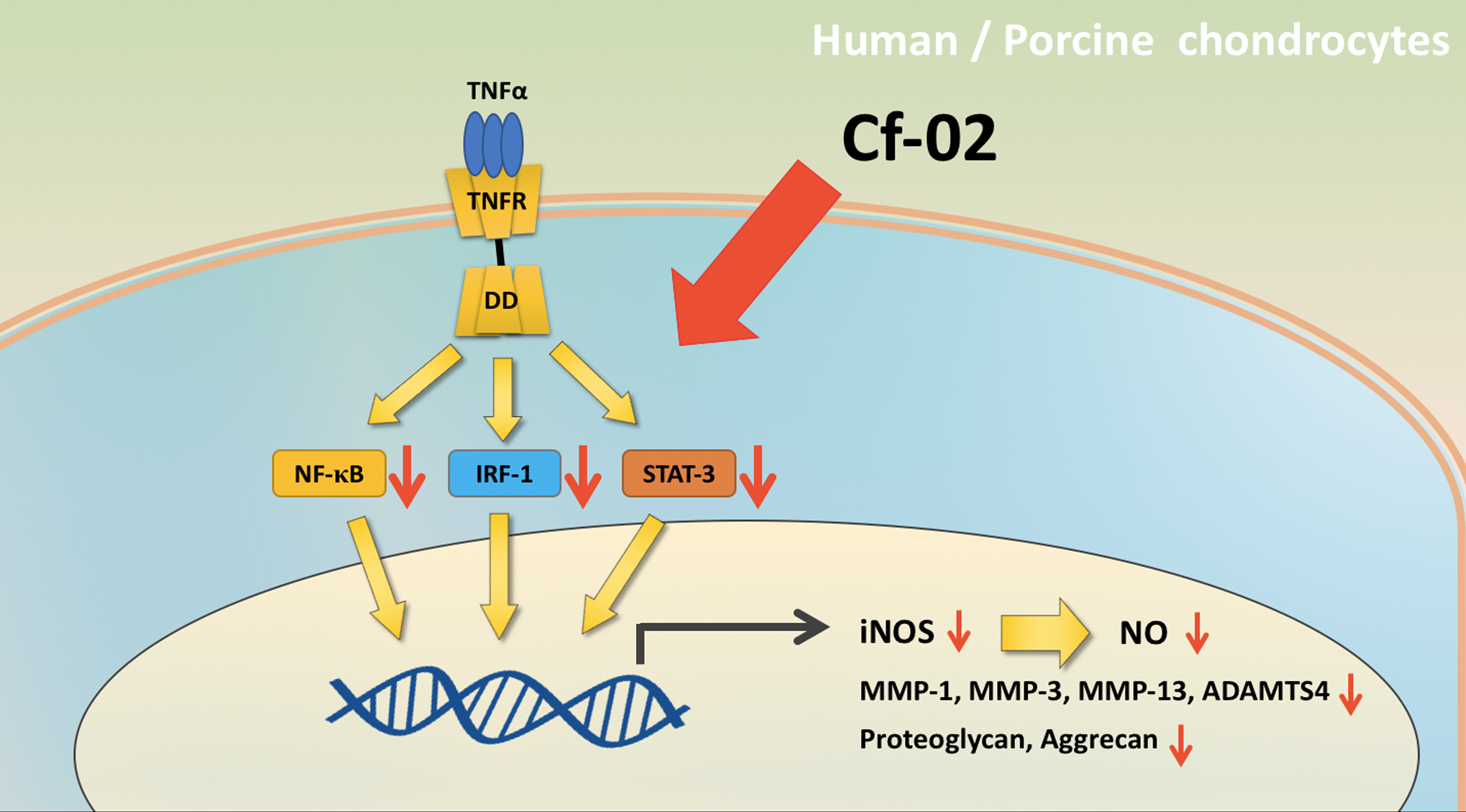
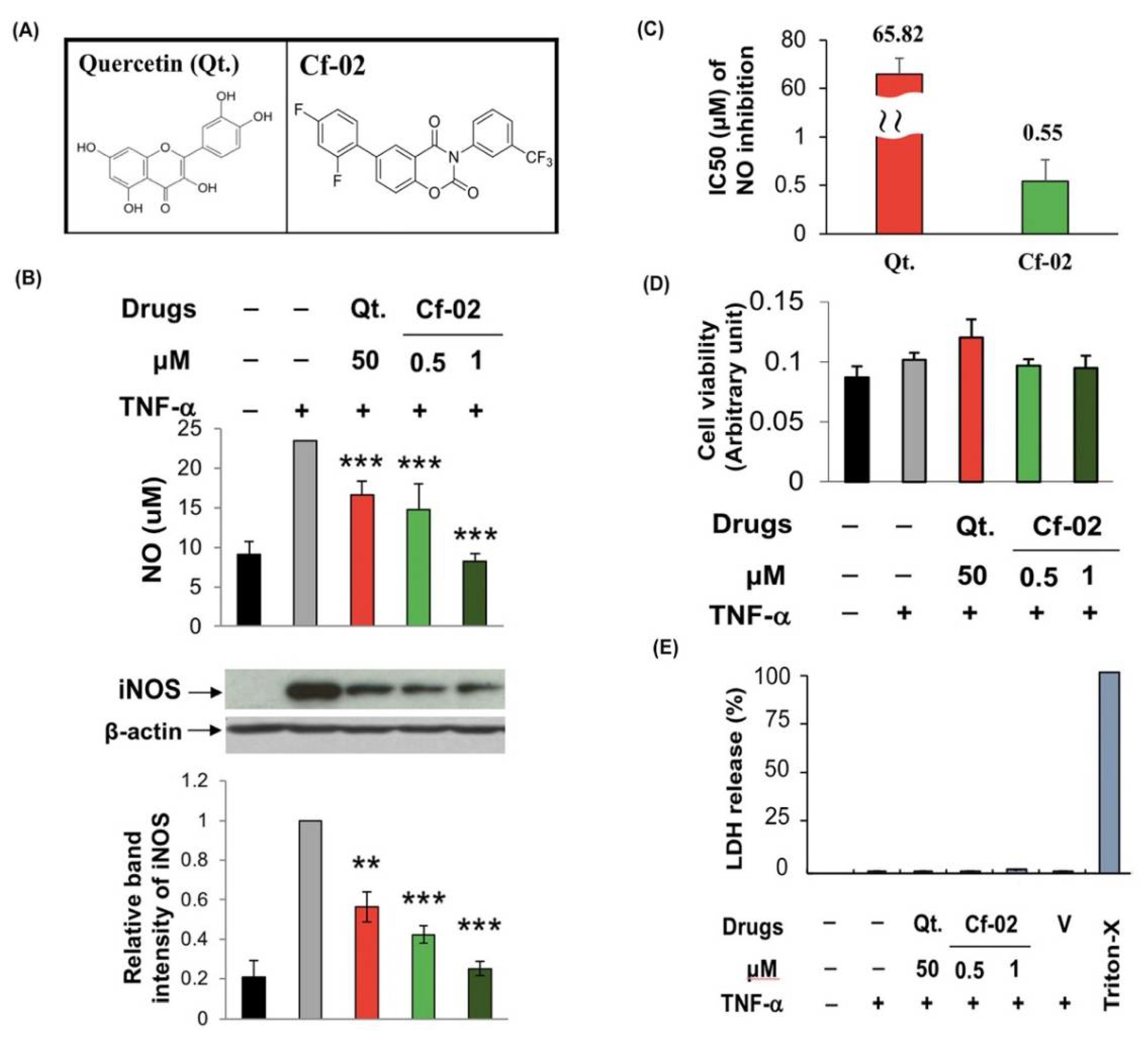
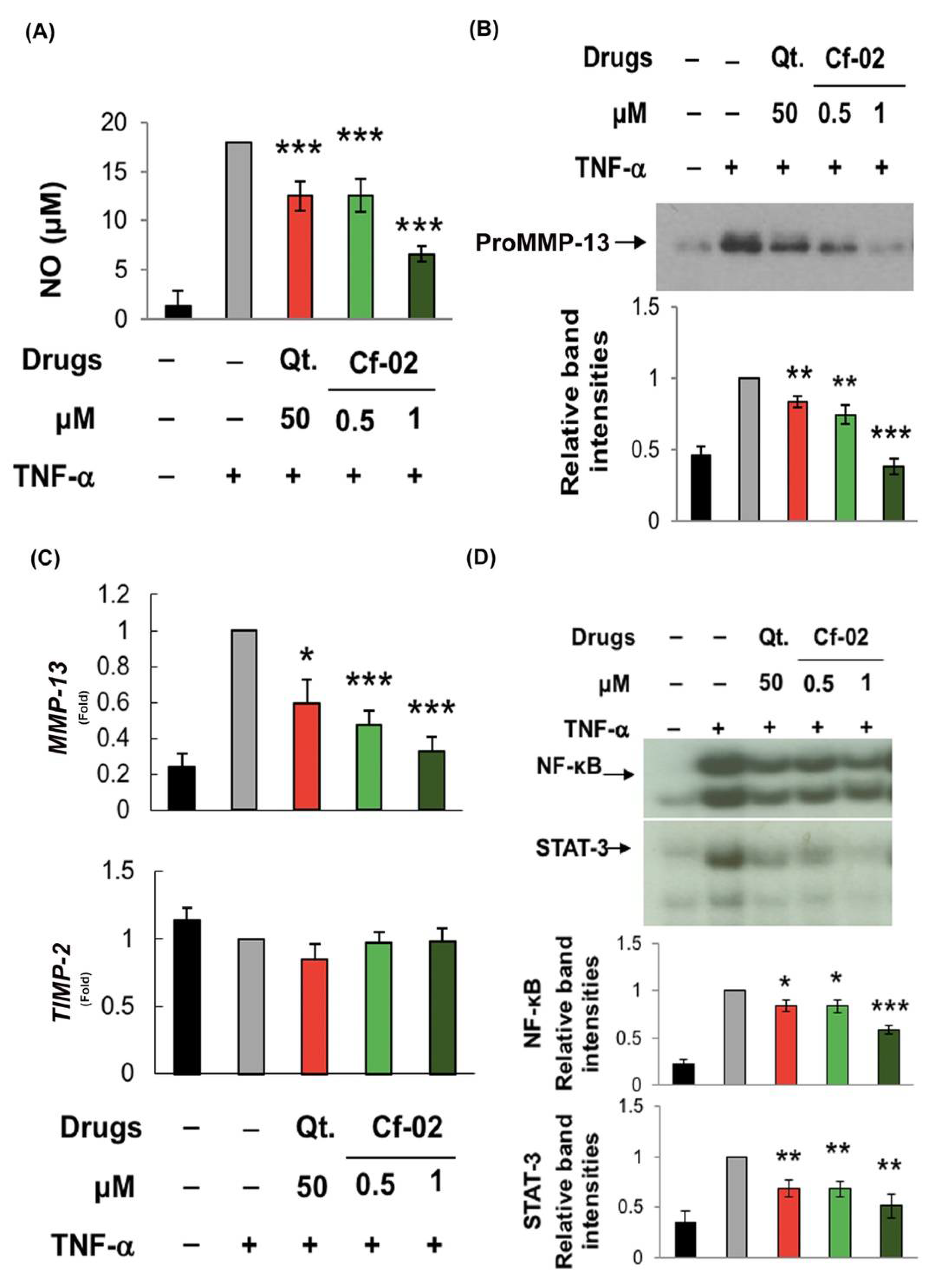
2.1. Inhibiting iNOS–NO Production in TNF-α-Stimulated Porcine Chondrocytes via Cf-02
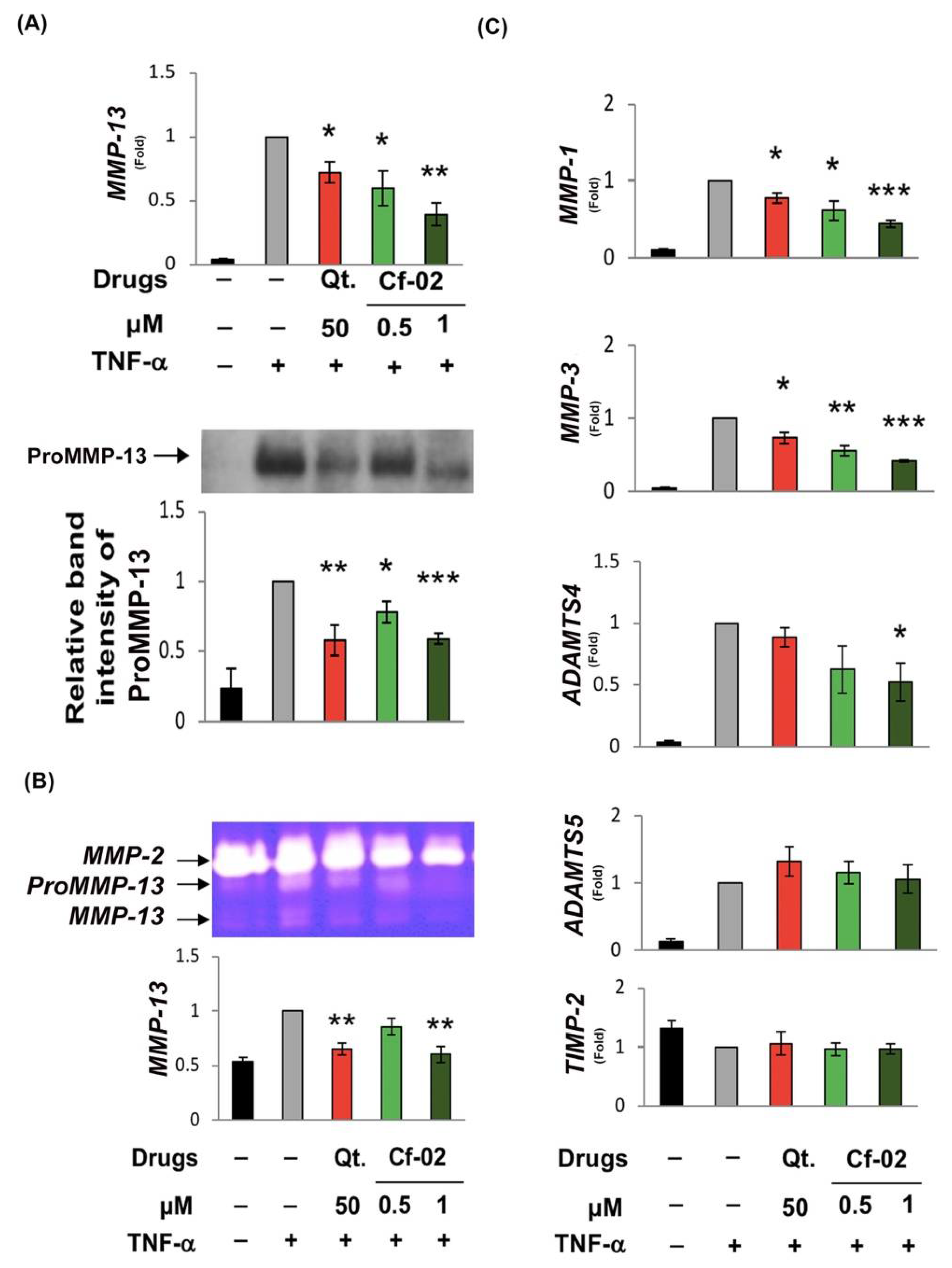
2.2. Inhibiting the Production of Chondro-Destructive Enzymes via Cf-02
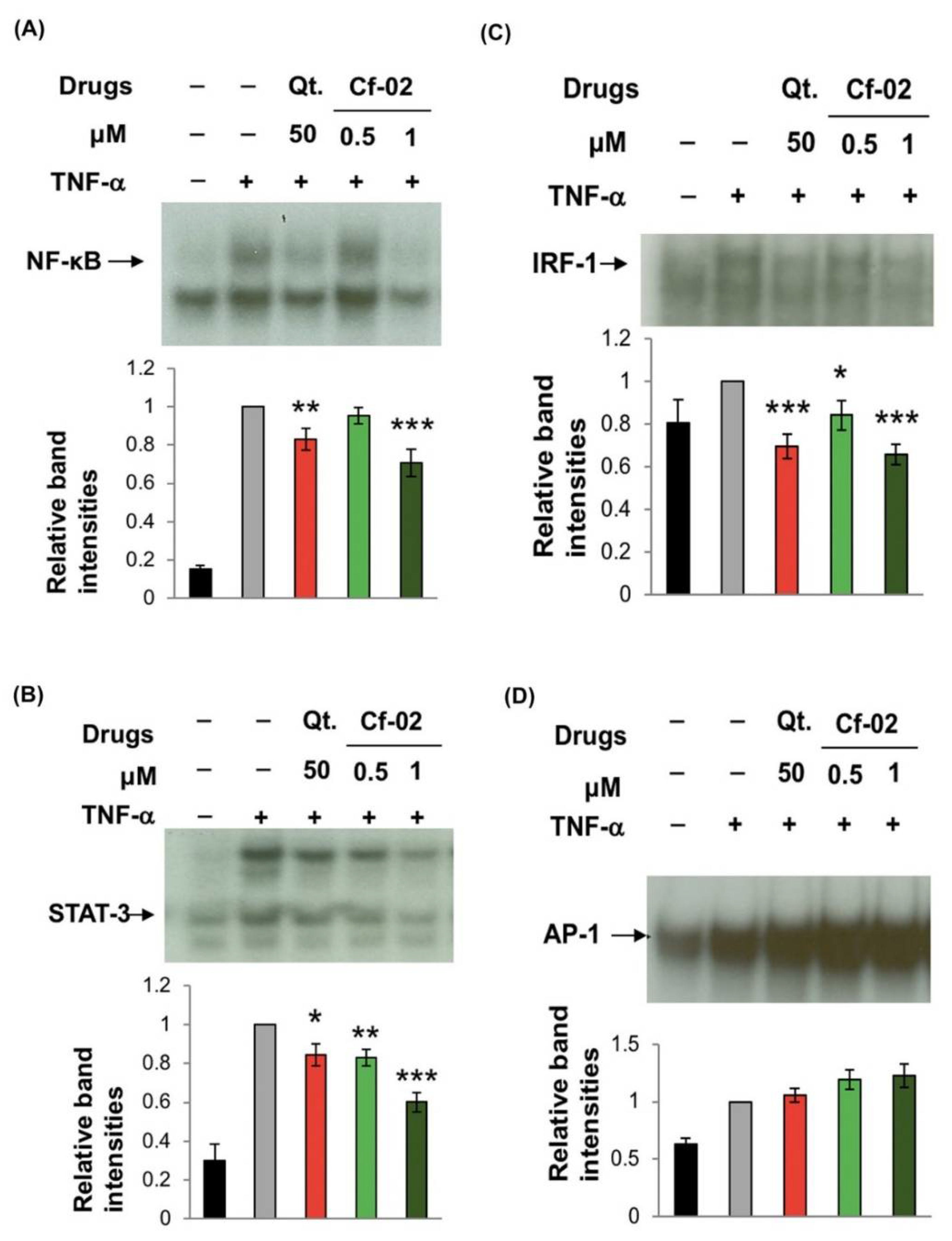
2.3. Regulating the Activity of Transcriptional Factors via Cf-02
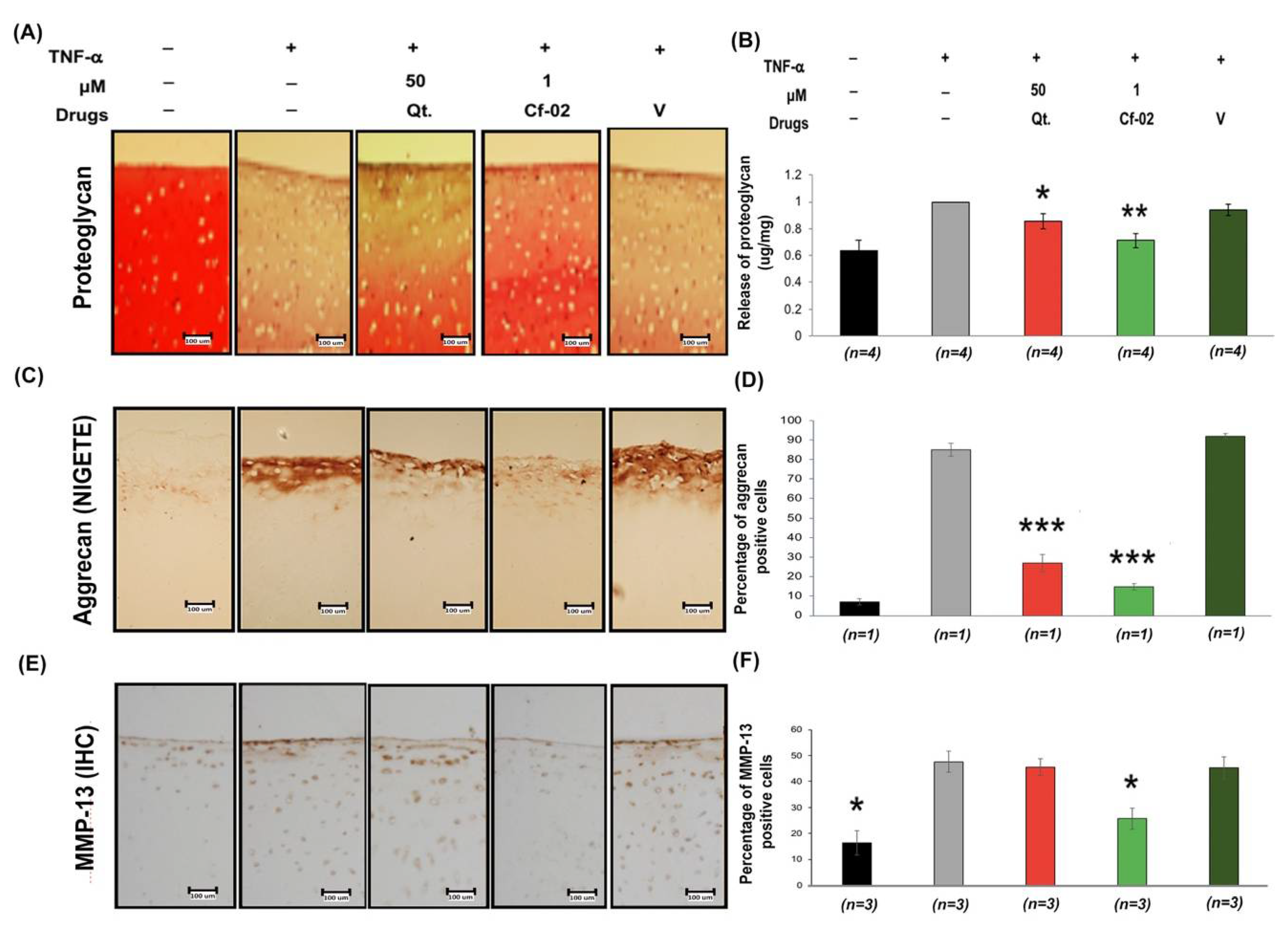
2.4. Effects of Cf-02 on TNF-α-Induced Proteoglycan/Aggrecan Degradation in Cartilage Explants
2.5. Immunomodulatory Effects of Cf-02 on Human Chondrocytes
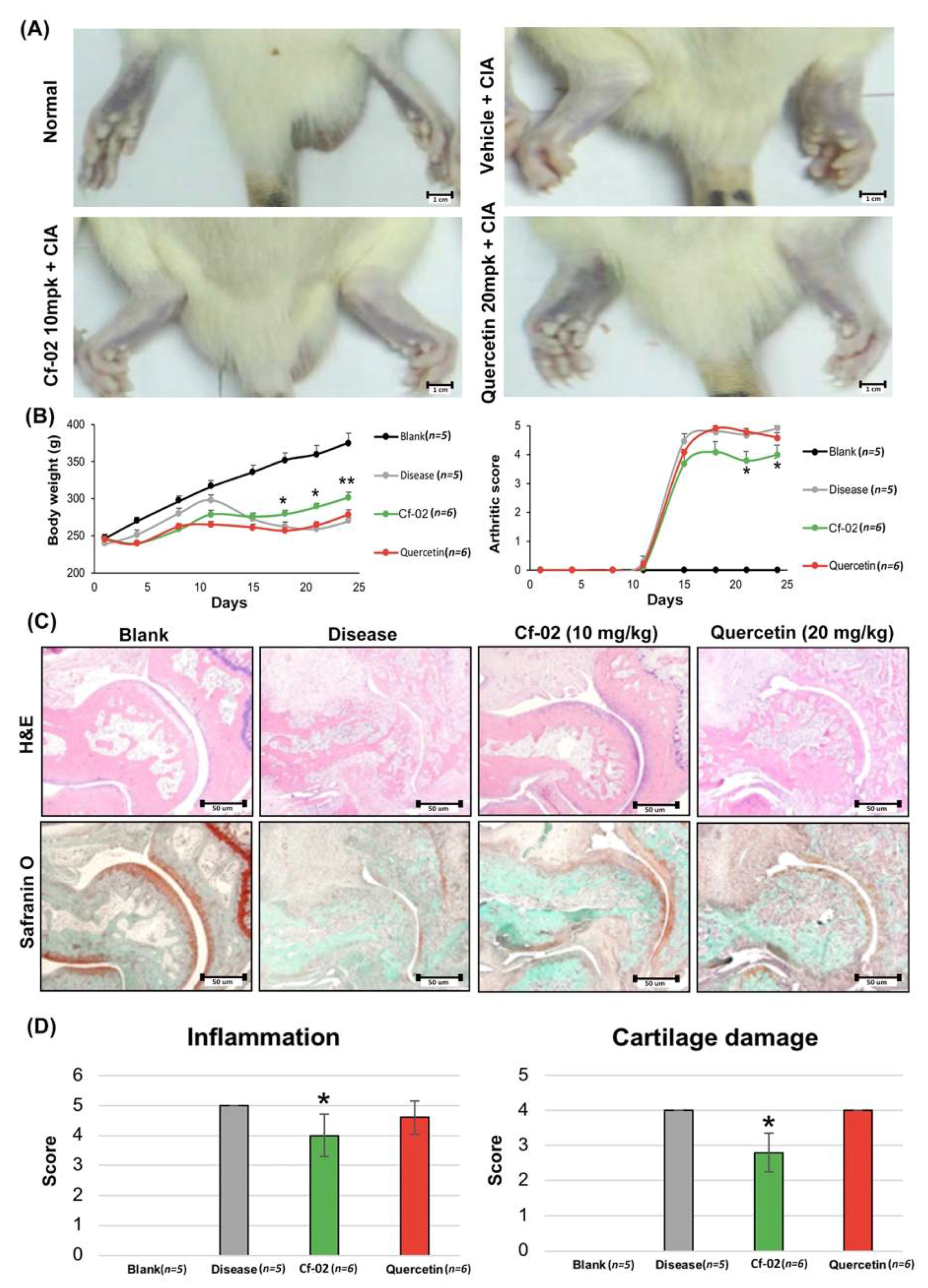
2.6. Prevention of Collagen Loss by Cf-02 in an Arthritis Animal Model
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Isolation and Culture of Porcine and Human Chondrocytes
4.3. Cytotoxicity Analysis and Measurement of NO Concentrations
4.4. Nuclear Extract Preparation and EMSA
4.5. Real-Time RT-PCR and Western Blotting
4.6. Gelatin Zymography
4.7. Preparation of Cartilage Explants and Analysis of Cartilage Degradation
4.8. Safranin-O Staining, IHC Staining, and Measurement of Aggrecan NITEGE Neoepitopes
4.9. Collagen-Induced Arthritis Model
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| TNF-α | tumor necrosis factor-alpha |
| HS-Cf | 2-hydroxy-N-[3-(trifluoromethyl)phenyl]benzamide |
| Cf-02 | 6-(2,4-difluorophenyl)-3-(3-(trifluoromethyl)phenyl)-2H-benzo[e][1,3]oxazine-2,4(3H)-dione |
| iNOS | nitric oxide synthase |
| NO | nitric oxide |
| MMPs | matrix metalloproteinases |
| ADAMTS | aggrecanases like a disintegrin and metalloproteinase with thrombospondin motifs |
| IRF-1 | interferon regulatory factor-1 |
| NF-κB | nuclear factor-kappaB |
| AP-1 | activator protein-1 |
| STAT-3 | signal transducer and activator of transcription-3 |
References
- Pap, T.; Korb-Pap, A. Cartilage damage in osteoarthritis and rheumatoid arthritis—Two unequal siblings. Nat. Rev. Rheumatol. 2015, 11, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Robinson, W.H.; Lepus, C.M.; Wang, Q.; Raghu, H.; Mao, R.; Lindstrom, T.M.; Sokolove, J. Low-grade inflammation as a key mediator of the pathogenesis of osteoarthritis. Nat. Rev. Rheumatol. 2016, 12, 580–592. [Google Scholar] [CrossRef] [PubMed]
- Herrero-Beaumont, G.; Roman-Blas, J.A.; Castaneda, S.; Jimenez, S.A. Primary osteoarthritis no longer primary: Three subsets with distinct etiological, clinical, and therapeutic characteristics. Semin. Arthritis Rheum. 2009, 39, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Abramson, S.B.; Attur, M. Developments in the scientific understanding of osteoarthritis. Arthritis Res. Ther. 2009, 11, 227. [Google Scholar] [CrossRef] [PubMed]
- Toivanen, A.T.; Heliovaara, M.; Impivaara, O.; Arokoski, J.P.; Knekt, P.; Lauren, H.; Kroger, H. Obesity, physically demanding work and traumatic knee injury are major risk factors for knee osteoarthritis—A population-based study with a follow-up of 22 years. Rheumatology 2010, 49, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Peffers, M.J.; Balaskas, P.; Smagul, A. Osteoarthritis year in review 2017: Genetics and epigenetics. Osteoarthr. Cartil. 2018, 26, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Billinghurst, R.C.; Dahlberg, L.; Ionescu, M.; Reiner, A.; Bourne, R.; Rorabeck, C.; Mitchell, P.; Hambor, J.; Diekmann, O.; Tschesche, H.; et al. Enhanced cleavage of type II collagen by collagenases in osteoarthritic articular cartilage. J. Clin. Investig. 1997, 99, 1534–1545. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, J.P.; Martel-Pelletier, J. New trends in the treatment of osteoarthritis. Semin. Arthritis Rheum. 2005, 34, 13–14. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.L. ADAMTS: A novel family of extracellular matrix proteases. Int. J. Biochem. Cell Biol. 2001, 33, 33–44. [Google Scholar] [CrossRef]
- Pelletier, J.P.; Jovanovic, D.; Fernandes, J.C.; Manning, P.; Connor, J.R.; Currie, M.G.; Di Battista, J.A.; Martel-Pelletier, J. Reduced progression of experimental osteoarthritis in vivo by selective inhibition of inducible nitric oxide synthase. Arthritis Rheum. 1998, 41, 1275–1286. [Google Scholar] [CrossRef]
- Pelletier, J.P.; Jovanovic, D.V.; Lascau-Coman, V.; Fernandes, J.C.; Manning, P.T.; Connor, J.R.; Currie, M.G.; Martel-Pelletier, J. Selective inhibition of inducible nitric oxide synthase reduces progression of experimental osteoarthritis in vivo: Possible link with the reduction in chondrocyte apoptosis and caspase 3 level. Arthritis Rheum. 2000, 43, 1290–1299. [Google Scholar] [CrossRef]
- Pelletier, J.P.; Martel-Pelletier, J.; Abramson, S.B. Osteoarthritis, an inflammatory disease: Potential implication for the selection of new therapeutic targets. Arthritis Rheum. 2001, 44, 1237–1247. [Google Scholar] [CrossRef]
- Reynard, L.N.; Loughlin, J. Insights from human genetic studies into the pathways involved in osteoarthritis. Nat. Rev. Rheumatol. 2013, 9, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A. Osteoarthritis year 2013 in review: Genetics and genomics. Osteoarthr. Cartil. 2013, 21, 1443–1451. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.C.; Huang, H.S.; Huang, C.Y.; Yang, R.; Chang, D.M.; Lai, J.H.; Ho, L.J. A benzamide-linked small molecule HS-Cf inhibits TNF-alpha-induced interferon regulatory factor-1 in porcine chondrocytes: A potential disease-modifying drug for osteoarthritis therapeutics. J. Clin. Immunol. 2011, 31, 1131–1142. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.J.; Hung, L.F.; Liu, F.C.; Hou, T.Y.; Lin, L.C.; Huang, C.Y.; Lai, J.H. Ginkgo biloba extract individually inhibits JNK activation and induces c-Jun degradation in human chondrocytes: Potential therapeutics for osteoarthritis. PLoS ONE 2013, 8, e82033. [Google Scholar] [CrossRef] [PubMed]
- Rigoglou, S.; Papavassiliou, A.G. The NF-κB signalling pathway in osteoarthritis. Int. J. Biochem. Cell Biol. 2013, 45, 2580–2584. [Google Scholar] [CrossRef] [PubMed]
- Goldring, M.B.; Otero, M. Inflammation in osteoarthritis. Curr. Opin. Rheumatol. 2011, 23, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Raman, S.; FitzGerald, U.; Murphy, J.M. Interplay of Inflammatory Mediators with Epigenetics and Cartilage Modifications in Osteoarthritis. Front. Bioeng. Biotechnol. 2018, 6, 22. [Google Scholar] [CrossRef] [PubMed]
- Felson, D.T. Osteoarthritis in 2010: New takes on treatment and prevention. Nat. Rev. Rheumatol. 2011, 7, 75–76. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.J. Pharmacologic therapy for osteoarthritis—The era of disease modification. Nat. Rev. Rheumatol. 2011, 7, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.C.; Hung, L.F.; Wu, W.L.; Chang, D.M.; Huang, C.Y.; Lai, J.H.; Ho, L.J. Chondroprotective effects and mechanisms of resveratrol in advanced glycation end products-stimulated chondrocytes. Arthritis Res. Ther. 2010, 12, R167. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Ding, S. Small molecules that modulate embryonic stem cell fate and somatic cell reprogramming. Trends Pharmacol. Sci. 2010, 31, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.J.; Lai, J.H. Small-molecule inhibitors for autoimmune arthritis: Success, failure and the future. Eur. J. Pharmacol. 2014, 747, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Liou, J.T.; Huang, H.S.; Chiang, M.L.; Lin, C.S.; Yang, S.P.; Ho, L.J.; Lai, J.H. A salicylate-based small molecule HS-Cm exhibits immunomodulatory effects and inhibits dipeptidyl peptidase-IV activity in human T cells. Eur. J. Pharmacol. 2014, 726, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.; Spagnuolo, C.; Tedesco, I.; Bilotto, S.; Russo, G.L. The flavonoid quercetin in disease prevention and therapy: Facts and fancies. Biochem. Pharmacol. 2012, 83, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Ji, Q.; Xu, X.; Zhang, Q.; Kang, L.; Xu, Y.; Zhang, K.; Li, L.; Liang, Y.; Hong, T.; Ye, Q.; et al. The IL-1beta/AP-1/miR-30a/ADAMTS-5 axis regulates cartilage matrix degradation in human osteoarthritis. J. Mol. Med. 2016, 94, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Hung, L.F.; Liang, C.C.; Ho, L.J. COX-2 and iNOS are critical in advanced glycation end product-activated chondrocytes in vitro. Eur. J. Clin. Investig. 2009, 39, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.J.; Lin, L.C.; Hung, L.F.; Wang, S.J.; Lee, C.H.; Chang, D.M.; Lai, J.H.; Tai, T.Y. Retinoic acid blocks pro-inflammatory cytokine-induced matrix metalloproteinase production by down-regulating JNK-AP-1 signaling in human chondrocytes. Biochem. Pharmacol. 2005, 70, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.W.; Liu, F.C.; Hung, L.F.; Huang, C.Y.; Lien, S.B.; Lin, L.C.; Lai, J.H.; Ho, L.J. Chondroprotective Effects and Mechanisms of Dextromethorphan: Repurposing Antitussive Medication for Osteoarthritis Treatment. Int. J. Mol. Sci. 2018, 19, 825. [Google Scholar] [CrossRef] [PubMed]
- Upton, M.L.; Chen, J.; Setton, L.A. Region-specific constitutive gene expression in the adult porcine meniscus. J. Orthop. Res. 2006, 24, 1562–1570. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.H.; Cheng, W.T.; Kuo, T.F.; Sun, J.S.; Lin, F.H.; Tsai, J.C. Fibrin glue mixed with gelatin/hyaluronic acid/chondroitin-6-sulfate tri-copolymer for articular cartilage tissue engineering: The results of real-time polymerase chain reaction. J. Biomed. Mater. Res. A 2007, 82, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Hansen, B.; Ornas, D.; Grigorian, M.; Klingelhofer, J.; Tulchinsky, E.; Lukanidin, E.; Ambartsumian, N. Extracellular S100A4(mts1) stimulates invasive growth of mouse endothelial cells and modulates MMP-13 matrix metalloproteinase activity. Oncogene 2004, 23, 5487–5495. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Hu, H.; Zhang, Y.; Yue, P.; Ning, L.; Zhou, Y.; Shi, P.; Yuan, R. Amelioration of collagen-induced arthritis using antigen-loaded dendritic cells modified with NF-kappaB decoy oligodeoxynucleotides. Drug Des. Dev. Ther. 2017, 11, 2997–3007. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, F.-C.; Lu, J.-W.; Chien, C.-Y.; Huang, H.-S.; Lee, C.-C.; Lien, S.-B.; Lin, L.-C.; Chen, L.W.; Ho, Y.-J.; Shen, M.-C.; et al. Arthroprotective Effects of Cf-02 Sharing Structural Similarity with Quercetin. Int. J. Mol. Sci. 2018, 19, 1453. https://doi.org/10.3390/ijms19051453
Liu F-C, Lu J-W, Chien C-Y, Huang H-S, Lee C-C, Lien S-B, Lin L-C, Chen LW, Ho Y-J, Shen M-C, et al. Arthroprotective Effects of Cf-02 Sharing Structural Similarity with Quercetin. International Journal of Molecular Sciences. 2018; 19(5):1453. https://doi.org/10.3390/ijms19051453
Chicago/Turabian StyleLiu, Feng-Cheng, Jeng-Wei Lu, Chiao-Yun Chien, Hsu-Shan Huang, Chia-Chung Lee, Shiu-Bii Lien, Leou-Chyr Lin, Liv Weichien Chen, Yi-Jung Ho, Min-Chung Shen, and et al. 2018. "Arthroprotective Effects of Cf-02 Sharing Structural Similarity with Quercetin" International Journal of Molecular Sciences 19, no. 5: 1453. https://doi.org/10.3390/ijms19051453