T Cell Calcium Signaling Regulation by the Co-Receptor CD5


Department of Microbiology and Molecular Biology, Brigham Young University, Provo, UT 84604, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(5), 1295; https://doi.org/10.3390/ijms19051295
Submission received: 27 February 2018
/
Revised: 19 April 2018
/
Accepted: 24 April 2018
/
Published: 26 April 2018
(This article belongs to the Special Issue Calcium Signaling in Human Health and Diseases)
Abstract
:Calcium influx is critical for T cell effector function and fate. T cells are activated when T cell receptors (TCRs) engage peptides presented by antigen-presenting cells (APC), causing an increase of intracellular calcium (Ca2+) concentration. Co-receptors stabilize interactions between the TCR and its ligand, the peptide-major histocompatibility complex (pMHC), and enhance Ca2+ signaling and T cell activation. Conversely, some co-receptors can dampen Ca2+ signaling and inhibit T cell activation. Immune checkpoint therapies block inhibitory co-receptors, such as cytotoxic T-lymphocyte associated antigen 4 (CTLA-4) and programmed death 1 (PD-1), to increase T cell Ca2+ signaling and promote T cell survival. Similar to CTLA-4 and PD-1, the co-receptor CD5 has been known to act as a negative regulator of T cell activation and to alter Ca2+ signaling and T cell function. Though much is known about the role of CD5 in B cells, recent research has expanded our understanding of CD5 function in T cells. Here we review these recent findings and discuss how our improved understanding of CD5 Ca2+ signaling regulation could be useful for basic and clinical research.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
T cells are a critical component of the adaptive immune system. T cell responses are influenced by signals that modulate the effects of the T cell receptor (TCR) and peptide-major histocompatibility complex (pMHC) interaction and initiate the transcription of genes involved in cytokine production, proliferation, and differentiation [1,2,3]. T cell activation requires multiple signals. First, the TCR engages the pMHC leading to tyrosine phosphorylation of CD3 and initiation of the Ca2+/Calcineurin/Nuclear factor of activated T cells (NFAT) or Protein kinase C-theta (PKCθ)/Nuclear factor-κ-light chain enhancer of activated B cells (NF-κB) or Mitogen-activated protein kinase (MAP kinase)/AP-1 pathways [4,5,6]. Second, cell surface costimulatory molecules, such as co-receptor CD28, amplify TCR-pMHC complex signals and promote stronger intracellular interactions to prevent T cell anergy [7,8]. Finally, cytokines such as interleukin-12 (IL-12), interferon α (INFα), and interleukin-1 (IL-1) promote T cell proliferation, differentiation, and effector functions [6].
Co-receptors such as CD4 and CD8 interact with MHC molecules and additional co-receptors interact with surface ligands present on antigen-presenting cells (APCs) to regulate T cell homeostasis, survival, and effector functions with stimulatory or inhibitory signals [9]. Altering co-receptor levels, balance, or function dramatically affects immune responses and their dysfunction is implicated in autoimmune diseases [10]. Stimulatory co-receptors such as CD28, inducible T cell co-stimulator (ICOS), Tumor necrosis factor receptor superfamily member 9 (TNFRSF9 or 4-1BB), member of the TNR-superfamily receptor (CD134 or OX40), glucocorticoid-induced tumor necrosis factor (TNF) receptor (GITR), CD137, and CD77 promote T cell activation and protective responses [11]. Co-receptor signaling is initiated by the phosphorylation of tyrosine residues located in immunoreceptor tyrosine-based activation motifs (ITAMs) or immunoreceptor tyrosine-based inhibitory motifs (ITIMs) [7,12]. The phosphorylated tyrosines serve as docking sites for spleen tyrosine kinase (Syk) family members such as zeta-chain-associated protein kinase 10 (ZAP-70) and Syk which activate the phospholipase C γ (PLCγ), RAS, and extracellular signal-regulated kinase (ERK) pathways in addition to mobilizing intracellular Ca2+ stores [13].
One of the best described T cell co-receptors, CD28, is a stimulatory T cell surface receptor from the Ig superfamily with a single Ig variable-like domain which binds to B7-1 (CD80) and B7-2 (CD86) [2]. Ligand binding phosphorylates CD28 cytoplasmic domain tyrosine motifs such as YMNM and PYAP and initiates binding and activation of phosphatidylinositide 3 kinase (PI3K) which interacts with protein kinase B (Akt) and promotes T cell proliferation and survival [1]. CD28 also activates the NFAT pathway and mobilizes intracellular Ca2+ stores through association with growth factor receptor-bound protein 2 (GRB2) and the production of phosphatidylinositol 4,5-bisphosphate (PIP2), the substrate of PLCγ1, respectively [2,14]. Blocking stimulatory co-receptors suppresses T cell effector function. For example, blocking stimulatory CD28 with anti-CD28 antibodies promotes regulatory T cell function and represses activation of auto- and allo-reactive T effector cells after organ transplantation [8,15].
T cells also have inhibitory co-receptors which regulate T cell responses [8]. The best characterized are immunoglobulin (Ig) superfamily members cytotoxic T-lymphocyte-associated protein 4 (CTLA-4) and programmed cell death protein 1 (PD-1) [8,16]. CTLA-4 binds CD80 and CD86 with greater avidity than CD28, and its inhibitory role refines early phase activation signals for proliferation and cytokine production [16,17,18,19]. PD-1, another CD28/B7 family member, regulates late phase effector and memory response [20]. Inhibitory co-receptors such as CTLA-4 and PD-1, known as “immune checkpoints”, block the interaction between CD28 and its ligands altering downstream secondary T cell activation signals [19]. Therefore, blocking CTLA-4 or PD-1 promotes effector T cell function in immunosuppressive environments [19,21].
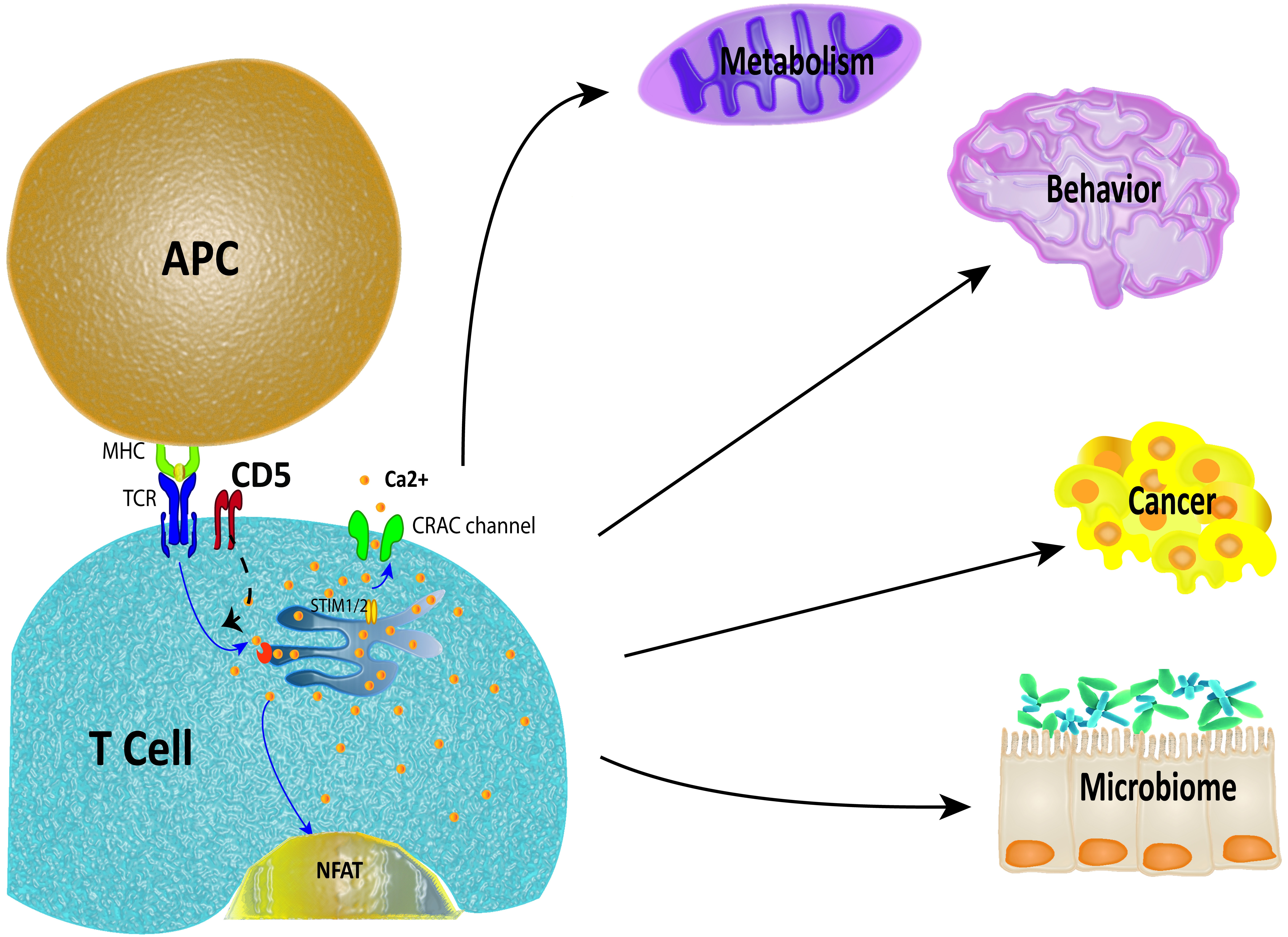
There are also a number of co-receptors that have differential modulatory properties. For example, CD5, a lymphocyte glycoprotein expressed on thymocytes and all mature T cells, has contradictory roles at different time points. CD5 expression is set during thymocyte development and decreases the perceived strength of TCR-pMHC signaling in naïve T cells by clustering at the TCR-pMHC complex and reducing TCR downstream signals such as the Ca2+ response when its cytoplasmic pseudo-ITAM domain is phosphorylated [22,23,24,25]. The CD5 cytoplasmic domain has four tyrosine residues (Y378, Y429, Y411, and Y463), and residues Y429 and Y441 are found in a YSQP-(x8)-YPAL pseudo ITAM motif while other tyrosine residues make up a pseudo-ITIM domain [23]. Phosphorylated tyrosines recruit several effector molecules and may sequester activation kinases away from the TCR complex, effectively reducing activation signaling strength [23]. Recruited proteins include Src homology-2 protein phosphatase-1 (SHP-1), Ras GTPase protein (rasGAP), CBL, casein kinase II (CK2), zeta-chain-associated protein kinase 70 (ZAP70), and PI3K which are involved in regulating both positive and negative TCR-induced responses [26,27,28]. For example, ZAP-70 phosphorylates other substrates and eventually recruits effector molecules such as PLC gamma and promotes Ca2+ signaling and Ras activation which stimulates the ERK pathway and leads to cellular activation [13,29]. Conversely, SHP1 inhibits Ca2+ signaling and PKC activation via decreased tyrosine phosphorylation of PLCγ [13,26,30,31]. Further, Y463 serves as a docking site for c-Cb1, a ubiquitin ligase, which is phosphorylated upon CD3–CD5 ligation and leads to increased ubiquitylation and lysosomal/proteasomal degradation of TCR downstream signaling effectors and CD5 itself [32]. Thus, CD5 has a mix of downstream effects that both promote and inhibit T cell activation. Curiously, recent work suggests that in contrast to its initial inhibitory nature, CD5 also co-stimulates resting and mature T cells by augmenting CD3-mediated signaling [25,33,34,35].
Ca2+ is an important second messenger in many cells types, including lymphocytes, and plays a key role in shaping immune responses. In naïve T cells, intracellular Ca2+ is maintained at low levels, but when TCR-pMHC complexes are formed, inositol triphosphate (IP3) initiates Ca2+ release from intracellular stores of the endoplasmic reticulum (ER) which opens the Ca2+ release-activated Ca2+ channels (CRAC) and initiates influx of extracellular Ca2+ through store-operated Ca2+ entry (SOCE) [36,37,38,39,40,41]. The resulting elevation of intracellular Ca2+ levels activates transcription factors involved in T cell proliferation, differentiation, and cytokine production (e.g., nuclear factor of activated cells (NFAT)) [36,37]. Thus, impaired Ca2+ mobilization affects T cell development, activation, differentiation, and function [42,43]. Examples of diseases with impaired Ca2+ signaling in T cells include systemic lupus erythematosus, type 1 diabetes mellitus, and others [44,45].
In this review, we will focus on CD5 co-receptor signaling and its functional effects on T cell activation. First, we will discuss how the inhibitory co-receptors CTLA-4 and PD-1 modulate T cell function. Then we will compare CTLA-4 and PD-1 function to CD5 function, examine recent findings that expand our understanding of the role of CD5, and assess how these findings apply to T cell Ca2+ signaling. Finally, we will consider CD5 Ca2+ signaling regulation in T cells and its potential physiological impact on immunometabolism, cell differentiation, homeostasis, and behavior.
2. Roles of Negative Regulatory T Cell Co-Receptors
2.1. Cytotoxic T-Lymphocyte Antigen-4 (CTLA-4)
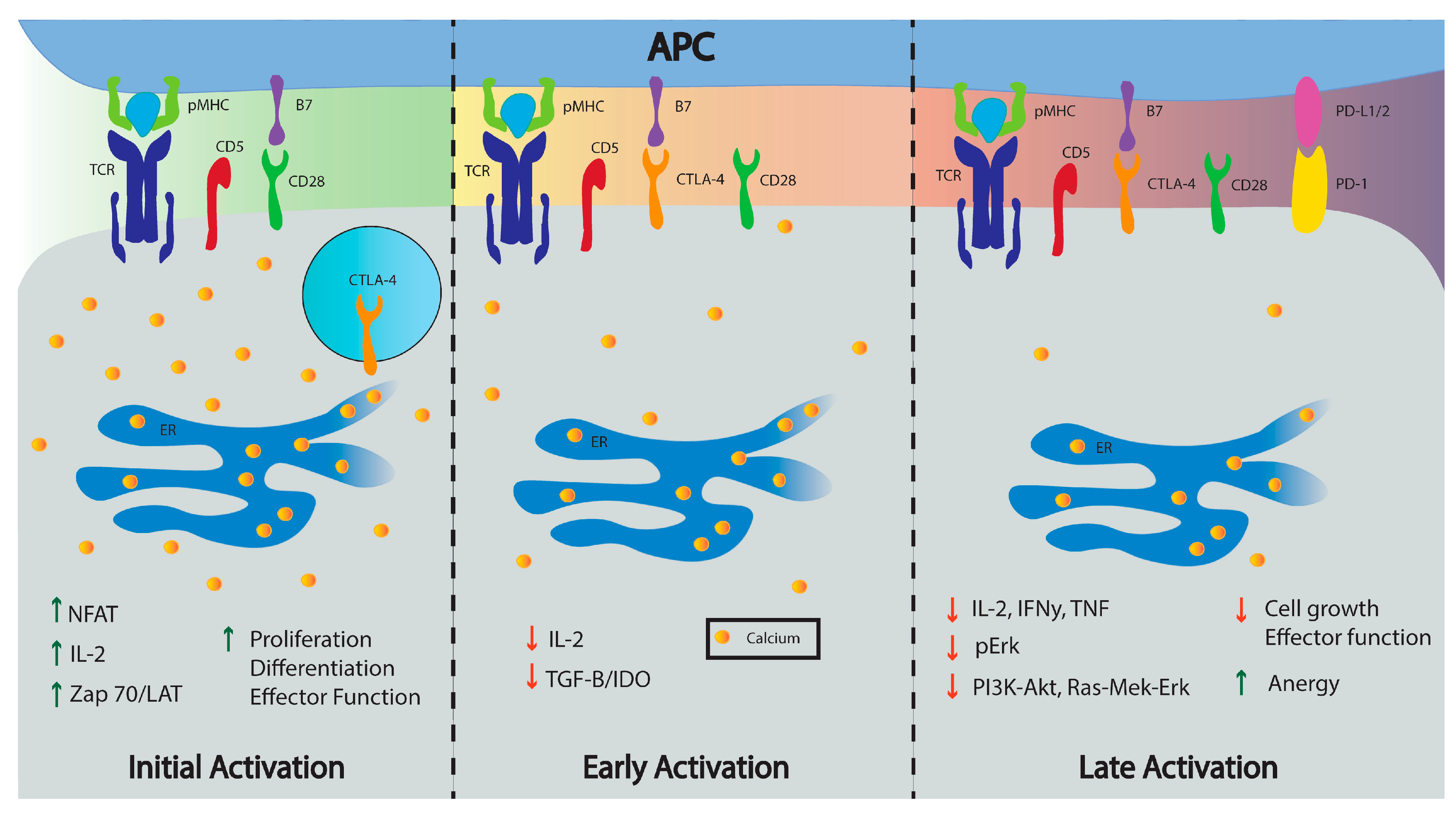
Cytotoxic T-lymphocyte antigen-4 (CTLA-4, CD152) inhibits early stages of T cell activation by recruiting inhibitory proteins such as SHP-2 and type II serine/threonine phosphatase PP2A that interfere with T cell synapse signaling [21,46,47,48]. CTLA-4 binds B7, a protein on activated APCs, with higher affinity than the stimulatory co-receptor CD28; the resulting balance between inhibitory and stimulatory signals controls T cell activation or anergy [19,49]. In naïve T cells, CTLA-4 is located in intracellular vesicles which localize at TCR binding sites following antigen recognition and intracellular Ca2+ mobilization [19,50]. Like CD28, CTLA-4 aggregates to the central supramolecular activation complex (cSMAC) where it then extrinsically controls activation by decreasing immunological synapse contact time [51,52,53]. This suppresses proactivation signals by activating ligands (B7-1 and B7-2) and induces the enzyme Inoleamine 2,3-dioxygenase (IDO) which impairs Ca2+ mobilization and suppresses T cell activation, ultimately altering IL-2 production and other effector functions in T cells [51,54,55]. CTLA-4 also stimulates production of regulatory cytokines, such as transforming growth factor beta (TGF-β), which inhibit APC presentation and T cell effector function [47,52,53]. Compared to effector T cells (Teff), CTLA-4 is highly expressed in regulatory T cells (Treg) and plays a role in maintaining Treg homeostasis, proliferation, and immune responses [16,56,57]. Total or partial CTLA-4 deficiency inhibits Treg’s ability to control cytokine production and can cause immune dysregulation [58,59,60,61]. Thus, CTLA-4 has an important role in the Treg suppressive response [60]. Additionally, CTLA-4 mutations are associated with autoimmune diseases as thoroughly reviewed by Kristiansen et al. [62].
The loss of CTLA-4 results in removal of CTLA-4 competition with CD28 for B7-1 and B7-2 and is implicated in autoimmunity and cancer [15,63]. Because CTLA-4 inhibits TCR signaling, CTLA-4 deficiency leads to T cell overactivation as measured by increased CD3ζ phosphorylation and Ca2+ mobilization [64]. Thus, modulating CTLA-4 signaling is an attractive target for immunotherapies that seek to boost or impair early TCR signaling for cancer and autoinflammatory diseases [65,66]. For example, Ipilimunab, an IgG1 antibody-based melanoma treatment, is a T cell potentiator that blocks CTLA-4 to stimulate T cell proliferation and stem malignant disease progression by delaying tumor progression and has been shown to significantly increase life expectancy [19,67,68]. Additionally, Tremelimumab, a noncomplement fixing IgG2 antibody, has been tested alone or in combination with other antibodies such as Durvalumab (a PD-1 inhibitor) and improves antitumor activity in patients with non-small cell lung cancer (NSCLC), melanoma, colon cancer, gastric cancer, and mesothelioma treatment [69,70,71,72,73,74].
2.2. Programmed Death 1 (PD-1)
Programmed cell death protein-1 (PD-1, CD279) is a 288-amino acid (50–55 KDa) type I transmembrane protein and a member of the B7/CD28 immunoglobulin superfamily expressed on activated T cells, B cells, and myeloid cells [19,75,76]. PD-1 has two known ligands, PD-L1 and PD-L2, which inhibit T cell activation signals [77]. Like CTLA-4, PD-1 also inhibits T cell proliferation and cytokine production (INF-γ, TNF and IL-2) but is expressed at a later phase of T cell activation [19]. PD-1 has an extracellular single immunoglobulin (Ig) superfamily domain and a cytoplasmic domain containing an ITIM and an immunoreceptor tyrosine-based switch motif (ITSM) subunit critical for PD-1 inhibitory function [78]. Upon T cell activation, PD-1 is upregulated and initiates ITIM and ITSM tyrosine interaction with SHP-2 which mediates TCR signaling inhibition by decreasing ERK phosphorylation and intracellular Ca2+ mobilization [79,80]. PD-1 can block the activation signaling pathways PI3K-Akt and Ras-Mek-ERK, which inhibit or regulate T cell activation [79,81]. Thus, engagement of PD-1 by its ligand affects intracellular Ca2+ mobilization, IL-2 and TNF-α production, supporting PD-1’s inhibitory role in TCR strength-mediated signals [82].
PD-1 signaling also affects regulatory T cell (Treg) homeostasis, expansion, and function [83]. Treg activation and proliferation are impacted by PD-1 expression which enhances their development and function while inhibiting T effector cells [75,84]. PD-1, PD-L, and Tregs help terminate immune responses [85]. Thus, PD-1 deficiency results not only in increased T cell activation, but in the breakdown of tolerance and the development of autoimmunity in diseases such as multiple sclerosis and systemic lupus erythematosus [85,86,87,88,89]. PD-1 and its ligands protect tissues from autoimmune attacks by regulating T cell activation and inducing and maintaining peripheral tolerance [90,91]. Studies done in PD-1-deficient mice observed the development of lupus-like glomerulonephritis and arthritis, cardiomyopathy, autoimmune hydronephrosis, and Type I diabetes, among other ailments [92,93,94]. PD-1 protects against autoimmunity and promotes Treg function. [85]. Enhancing Treg response with a PD-L1 agonist shows therapeutic potential for asthma and other autoimmune disorders [85,95]. Because PD-1 specifically modulates lymphocyte function, effective FDA-approved monoclonal antibodies targeting PD-1 are clinically available (i.e., Pembrolizumab and Nivolumab) to treat advanced malignancies [20]. Not only does blocking PD-1 decrease immunotolerance of tumor cells, it also increases cytotoxic T lymphocyte antitumor activity [20].
3. CD5: A Contradictory Co-Receptor
3.1. Overview of CD5 Signaling and Ca2+ Mobilization in T Cells
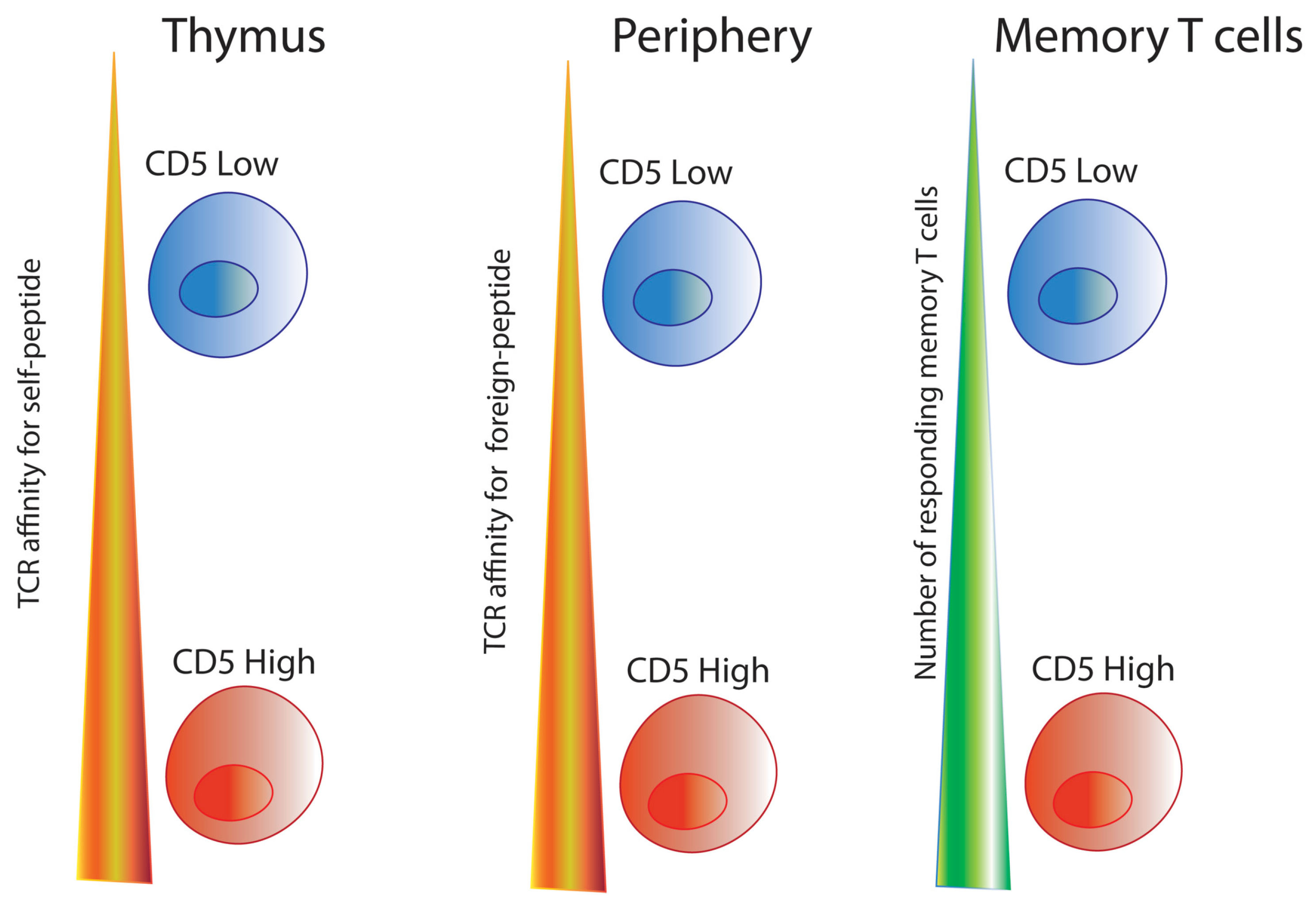
CD5, known as Ly-1 antigen in mice or as Leu-1 in humans, is a type I transmembrane glycoprotein (67 kDa) expressed on the surface of thymocytes, mature T cells, and a subset of B cells (B-1a) [96,97]. Although CD5 was discovered over 30 years ago, it was only in the last decade that CD5 gained attention as a key T cell activation regulator [98,99]. CD5 expression is set in the thymus during positive selection and correlates with how tightly the thymocyte TCR binds to self-peptide-MHC (self-pMHC); greater TCR affinity for self-peptide leads to increased CD5 expression in double positive (DP) thymocytes [100]. In other words, DP thymocytes that receive strong activation signals through their TCR express more CD5 than those DP thymocytes that receive weak TCR signals [100]. CD5 knockout mice (CD5−/−) have a defective negative and positive selection process, and therefore their thymocytes are hyper-responsive to TCR stimulation with increased Ca2+ mobilization, proliferation, and cytokine production [23,98]. On the other hand, because of the increased TCR avidity for self-pMHC, mature T cells with high CD5 expression (CD5hi) (peripheral or postpositive selection T cells) respond to foreign peptide with increased survival and activation compared to mature T cells with low CD5 expression (CD5lo) [34,101]. Therefore, CD5 is a negative regulator of TCR signaling in the thymus and modulates mature T cell response in the periphery [23,34,100,102].
While CTLA-4 and PD-1 belong to the immunoglobulin (Ig) family, CD5 belongs to group B of the scavenger receptor cysteine-rich (SRCR) superfamily and contains three extracellular SRCR domains [30,96,103]. The cytoplasmic tail of CD5 contains several tyrosine residues which mediate the negative regulatory activity independent of extracellular engagement [100,104,105]. As CD5 physically associates with TCRζ/CD3 complex upon TCR and pMHC interaction, the tyrosine residues in both TCRζ and CD5 are phosphorylated by tyrosine kinases associated with the complex [30,106,107,108,109,110]. This interaction is so intrinsic to T cell signaling that CD5 expression levels are proportional to the degree of TCRζ phosphorylation, IL-2 production capacity, and ERK phosphorylation which are critical for CD3-mediated signaling [33,111]. It is unknown whether posttranslational modifications, such as conserved domain 1 and domain 2 glycosylations, impact CD5 signaling [112,113]. CD5 is present in membrane lipids rafts of mature T cells where, upon activation, it helps augment TCR signaling, increases Ca2+ mobilization, and upregulates ZAP-70/LAT (linker for activation of T cells) activation [114,115,116]. This suggests that CD5 is not only a negative regulator in thymocytes, but also appears to positively influence T cell immune response to foreign antigens [117,118]. See Figure 1.
CD5 has three known ligands: CD72, a glycoprotein expressed by B cells, CD5 ligand or CD5L, an activation antigen expressed on splenocytes, and CD5 itself [120,121,122]. Crosslinking CD5L to CD5 increases intracellular Ca2+ concentrations [30,120,121,123,124]. Early studies with anti-CD5 monoclonal antibodies also demonstrated enhanced Ca2+ mobilization and proliferation, suggesting that CD5 co-stimulates and increases the T cell activation signal [125,126]. Following TCR:pMHC interaction, CD5 cytoplasmic ITAM and ITIM like-domains are phosphorylated by p56lck and bound by Src homology 2 (SH2) domain-containing protein tyrosine phosphatase (SHP-1) [108,127,128]. However, while SHP-1 affects Ca2+ mobilization and is a purported down-regulator of thymocyte activation, recent findings suggest that SHP-1 is not necessary for CD5 signaling as T cells deficient in SHP-1 have normal CD5 expression and continue to signal normally [26,129]. Thus, while CD5 is not a SHP-1 substrate and SHP-1 is likely unnecessary for CD5 signaling, CD5 signaling results in increased Ca2+ mobilization. It has yet to be resolved how CD5 can act as an inhibiting co-receptor in the thymus and as an activating co-receptor in the periphery.
3.2. CD5 as a Ca2+ Signaling Modulator
As previously mentioned, CD5 expression levels are set in the thymus during T cell development and are maintained on peripheral lymphocytes [117]. CD5 expression in T cells plays an important role during development and primes naïve T cells for responsiveness in the periphery [35,111,130]. CD5hi T cells have the highest affinity for self-peptides and respond with increased cytokine production and proliferation to infection [101,131,132].
Our laboratory works with two TCR transgenic mouse lines with different levels of CD5 expression: LLO56 (CD5hi) and LLO118 (CD5lo) [111,117,130]. While LLO56 (CD5hi) and LLO118 (CD5lo) have similar affinity for the same immunodominant epitope (listeriolysin O amino acids 190–205 or LLO190–205) from Listeria monocytogenes, on day 7 of primary response, LLO118 (CD5lo) has approximately three times the number of responding cells compared to LLO56 (CD5hi), and conversely, on day 4 during secondary infection, LLO56 (CD5hi) has approximately fifteen times more cells than LLO118 (CD5lo) [130]. This difference is not due to differential proliferative capacity, rather LLO56 (CD5hi) has higher levels of apoptosis during the primary response [130]. Thus, LLO56 CD5hi and LLO118 CD5lo’s capacity to respond to infection appears to be regulated by their CD5 expression levels [117]. LLO56 (CD5hi) thymocytes have greater affinity for self-peptide, which primes them to be highly apoptotic [130].
Recently we reported that in response to foreign peptide, LLO56 (CD5hi) naïve T cells have higher intracellular Ca2+ mobilization than LLO118 (CD5lo), which correlates with increased rate of apoptosis of LLO56 (CD5hi), as Ca2+ overloaded mitochondria release cytochrome c which activates caspase and nuclease enzymes, thus initiating the apoptotic pathways [35,133,134]. LLO56 (CD5hi) naïve T cell increased Ca2+ mobilization also provides additional support to the idea that CD5hi T cells have an enhanced response to foreign peptide [35,134]. This supports previous research that found that upon T cell activation, increased CD5 expression is correlated with greater basal TCRζ phosphorylation, increased ERK phosphorylation, and more IL-2 production [101,111].
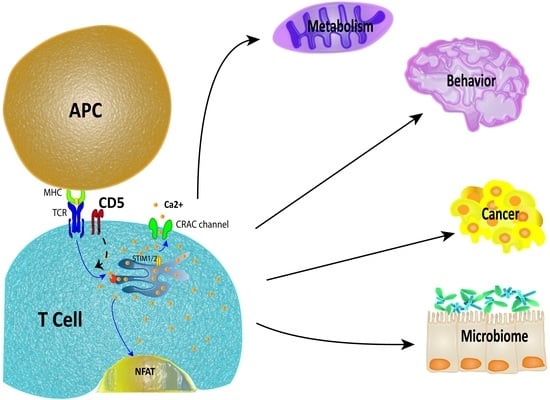
Thus, unlike CTLA-4 and PD-1 which are expressed only on activated T cells in the periphery during early and late phases of immune response, respectively, CD5 is set during T cell development, and influences T cells both during thymic development and during postthymic immune responses [19,101,111] (see Figure 2). CD5 not only has an important inhibitory role in the thymus, but also appears to positively influence the T cell population response; for example, more CD5hi T cells populate the memory T cell repertoire because CD5hi naïve T cells have a stronger primary response [34,135]. CD5 finetunes the sensitivity of TCR signaling to pMHC, altering intracellular Ca2+ mobilization and NFAT transcription, key players in T cell effector function [19,64,126]. As Ca2+ signaling plays a key role in T cell activation and function, controlling Ca2+ mobilization in T cells through CD5 expression could influence diverse areas of clinical research including metabolism, cancer treatments, and even cognitive behavior.
4. Physiological Impact of CD5 Expression in T Cells
4.1. Metabolism
Naive T cells are in a quiescent state and rely on oxidative phosphorylation (OXPHOS) to generate ATP for survival [136,137]. Upon TCR-pMHC interaction, T cells undergo metabolic reprograming to meet energetic demands by switching from OXPHOS to glycolysis [138]. Glycolysis is a rapid source of ATP and regulates posttranscriptional production of INF-γ, a critical effector cytokine [139]. Following the immune response, most effector T cells undergo apoptosis while a subset become quiescent memory T cells. Memory T cells have lower energetic requirements and rely on OXPHOS and Fatty Acid Oxidation (FAO) to enhance mitochondrial capacity for maintenance and survival [140].
Ca2+ signaling is a key second messenger in T cell activation and Ca2+ ions also modulate T cell metabolism through CRAC channel activity and NFAT activation [3,141]. During TCR-pMHC binding Ca2+ is released from the endoplasmic reticulum (ER) where it is absorbed by the mitochondria and initiates an influx of extracellular Ca2+ [3]. First, the rise of cytoplasmic Ca2+ activates stromal interaction molecule 1 (STIM1) located on the ER membrane to interact with the CRAC channel located on the cell membrane [142]. The release of the ER store and resulting extracellular Ca2+ influx increases the intracellular Ca2+ concentration and promotes AMPK (adenosine monophosphates activated protein kinase) expression and CaMKK (calmodulin-dependent protein kinase kinase) activity [3,142,143]. AMPK senses cellular energy levels through the ratio of AMP to ATP and generates ATP by inhibiting ATP-dependent pathways and stimulating catabolic pathways [144]. This indirectly controls T cell fate as AMPK indirectly inhibits mTOR (mammalian target of rapamycin complex) [145]. Because mTOR coordinates the metabolic cues that control T cell homeostasis, it plays a critical role in T cell fate [146]. T cells that are TSC1 (Tuberous sclerosis complex 1)-deficient show metabolic alterations through increased glucose uptake and glycolytic flux [147].
The rise of cytoplasmic Ca2+ also encourages mitochondria to uptake cytoplasmic Ca2+ through the mitochondrial Ca2+ uniporter (MCU) [148]. This MCU uptake increases Ca2+ influx by depleting Ca2+ near the ER which further activates the CRAC channels and promotes STIM1 oligomerization [3,149,150,151]. Ca2+ uptake in the mitochondria also enhances the function of the tricarboxylic acid cycle (TAC), which generates more ATP through OXPHOS [152,153]. OXPHOS is maintained by a glycolysis product, phosphoenolpyruvate (PEP), which sustains TCR-mediated Ca2+-NFAT signaling by inhibiting the sarcoendoplasmic reticulum (SR) calcium transport ATPase (SERCA) pump, thus promoting T cell effector function [154,155]. Downregulation of calmodulin kinase, CaMKK2, which controls NFAT signaling, decreases glycolytic flux, glucose uptake, and lactate and citrate metabolic processes [156]. Ca2+ may also orchestrate the metabolic reprogramming of naïve T cells by promoting glycolysis and OXPHOS through the SOCE/calcineurin pathway which controls the expression of glucose transporters GLUT1/GLUT3 and transcriptional co-regulator proteins important for the expression of electron transport chain complexes required for mitochondria respiration [141].
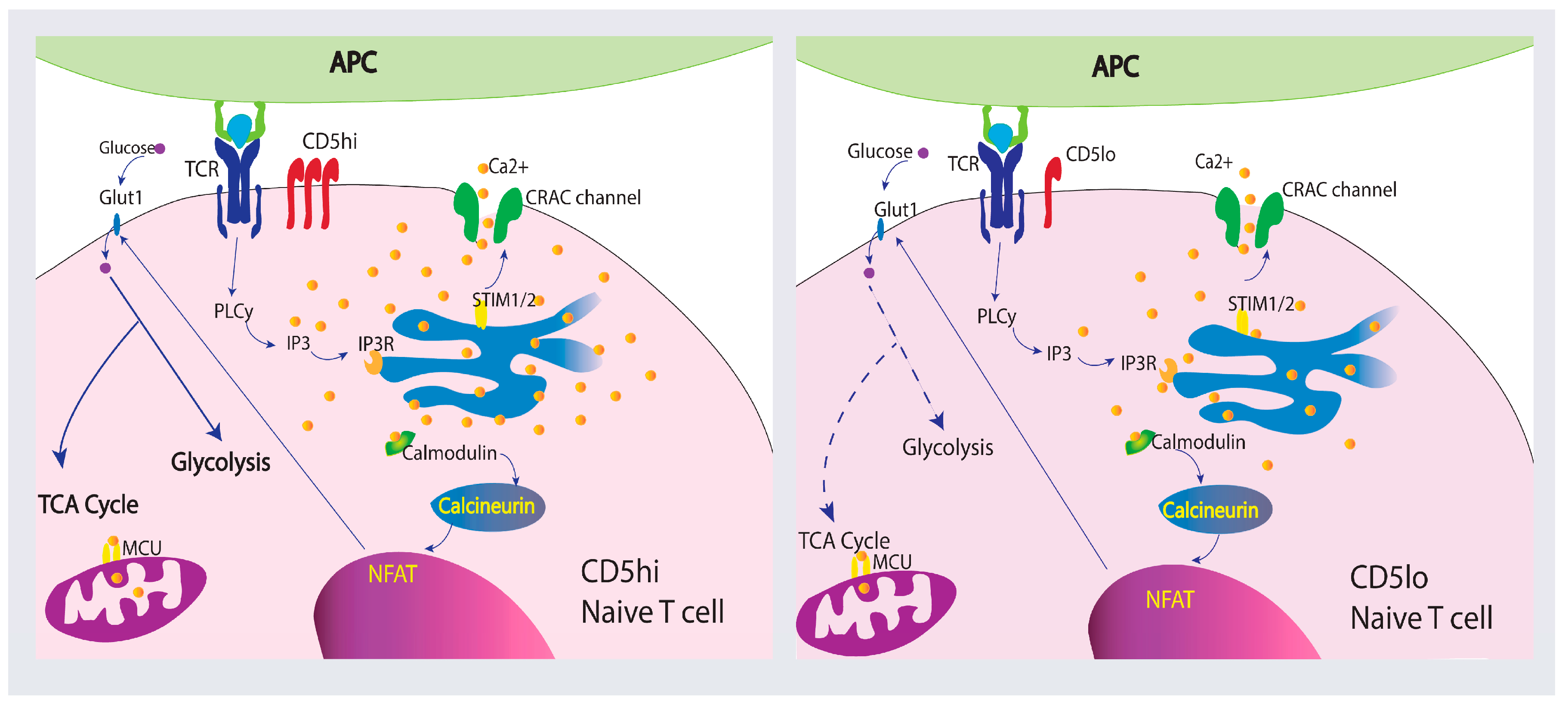
Co-receptor stimulation plays a pivotal role in T cell metabolism and function. A decrease in T cell Ca2+ signaling represses glycolysis and affects T cell effector function [152]. PD-1 and CTLA-4 depress Ca2+ signaling and glycolysis while promoting FAO and antibodies against CTLA-4 and PD-1 increase Ca2+ mobilization and glycolysis during T cell activation [157,158]. Like CTLA-4 and PD-1, CD5 modulatory function has the potential to influence T cell metabolism. Analysis of gene families modulated by CD5 in B cells found that CD5 upregulates metabolic-related genes including VEFG, Wnt signaling pathways genes, MAPK cascade genes, I-kB/NF-kB cascade genes, TGF β signaling genes, and adipogenesis process genes [159]. Therefore, proliferation differences correlated with CD5 expression in T cells may be caused by improved metabolic function as CD5lo T cells seem to be more quiescent than CD5hi T cells [160]. Although not much is known about how CD5 alters metabolic function in T cells, signaling strength differences of CD5hi and CD5lo T cell populations correlate with intracellular Ca2+ mobilization during activation and influence their immune response [35,111,130]. This implies that different metabolic processes may be initiated which would influence proliferation, memory cell generation, and cytokine production. Figure 3 summarizes how Ca2+ may be mobilized in CD5hi and CD5lo naïve T cells and the role Ca2+ may play on metabolism.
4.2. Neuroimmunology
The field of neuroimmunology examines the interplay between the immune system and the central nervous system (CNS) [161]. The adaptive immune system does influence the CNS as cognition is impaired by the absence of mature T cells [162]. In wild type mice, there is an increase in the number of T cells present in the meninges during the learning process, in stark contrast to mice with T helper 2 cytokine deficiencies (such as IL-4 and IL-13) who have decreased T cell recruitment and impaired learning [163]. Furthermore, regulation of T cell activation and cytokine production critically assists neuronal function and behavior, suggesting that manipulation of T cells could be a potential therapeutic target in treating neuroimmunological diseases [164,165].
T cells go through several microenvironments before reaching the CNS [166]. Many of the signal interactions present in these microenvironments affect T cell function and involve changes in intracellular Ca2+ levels [166,167]. In experimental autoimmune encephalitis (EAE), a model for human multiple sclerosis, autoreactive T cells have Ca2+ fluctuations throughout their journey to the CNS [166]. Prior to reaching the CNS, T cells interact with splenic stroma cells that do not display the cognate auto-antigen and this interaction produces short-lived low Ca2+ mobilization spikes [166]. Following entrance into the CNS, T cells encounter autoantigen-presenting cells and have sustained Ca2+ mobilization which results in NFAT translocation and T cell activation [166,168]. EAE mice display reduced social interaction and cognition demonstrating that autoimmune response impairs neuronal function and organismal behavior [169].
Inhibitory T cell co-receptors are implicated in CNS dysregulation and disease. Varicella zoster virus (VZV) infection is characterized by lifelong persistence in neurons. VZV increases the expression of CTLA-4 and PD-1 in infected T cells which reduces IL-2 production and increases T cell anergy [170,171]. PD-1-deficient mice (Pdcd1−/−) have increased T cell activation, leading to greater intracellular Ca2+ mobilization, and as previously discussed, increased glycolysis [86]. PD-1 deficiency causes elevated concentration of aromatic amino acids in the serum, specifically tryptophan and tyrosine, which decreases their availability in the brain where they are important for the synthesis of neurotransmitters such as dopamine and serotonin; consequently, there is an increase in anxiety-like behavior and fear in Pdcd1−/− mice [86]. Therefore, increased T cell activation caused by PD-1 deficiency can affect brain function and thus, affects cognitive behavior [86].
4.3. Cancer
T cells are critical components of the immune response to cancer. Helper T cells directly activate killer T cells to eradicate tumors and are essential in generating a strong antitumor response alone or in concert with killer T cells by promoting killer T cell activation, infiltration, persistence, and memory formation [172,173,174,175,176,177]. Tumor-specific T cells may not mount a robust response towards cancerous cells because the tumor microenvironment has numerous immunosuppressive factors; cancerous cells also downregulate cell surface co-stimulatory and MHC proteins which suppresses T cell activation [178,179,180,181,182]. Potent antitumor immune checkpoint blockade therapies using CTLA-4 and PD-1 monoclonal antibodies augment T cell response by suppressing the co-receptors’ inhibitory signals, thereby promoting increased Ca2+ mobilization, glycolysis, and activation [183,184]. CTLA-4 monoclonal antibodies such as ipilimumab (Yervoy) and tremelimumab block B7-interaction and have been used to treat melanoma [47,185,186]. The monoclonal antibody pembrolizumab is highly selective for PD-1 and prevents PD-1 from engaging PD-L1 and PD-L2, thus enhancing T cell immune response [19,187,188]. Further research will address whether combining anti-CTLA-4 and anti-PD-1 antibodies will improve cancer treatments [19].
As previously mentioned, Ca2+ is critical for T cell activation and immune response. Manipulating Ca2+ signaling to enhance T cell-directed immune response against cancer is an intriguing notion, yet the means to target the Ca2+ response of specific cells without tampering with the metabolic processes of other cells remains elusive [189]. Antitumor activity of tumor-infiltrating lymphocytes (TIL) is inversely related to CD5 expression [99]. CD5 levels in naïve T cells are constantly tuned in the periphery by interactions with self pMHC complexes to maintain homeostasis; therefore, CD5 expression on TILs can be downregulated in response to low affinity for cancer antigens [190,191,192]. Thus, the majority of TILs are CD5lo which increase their reactivity while CD5hi TILs do not elicit a Ca2+ response and become anergic and are unable to eliminate malignant cells [99,192]. While downregulation of CD5 on TILs enhances antitumor T cell activity, CD5lo T cells are also more likely to experience activation-induced cell death (AICD) as CD5 protects T cells from overstimulation [23]. To maximize TIL effectiveness, the inhibitory effects of CD5 could be blocked by neutralizing monoclonal antibodies or soluble CD5-Fc molecules combined with soluble FAS-Fc molecules to reduce the inherent AICD [23,193,194]. Soluble human CD5 (shCD5) may have a similar effect but avoids targeting issues by blocking CD5-mediated interaction via a “decoy receptor” effect. Mice constitutively expressing shCD5 had reduced melanoma and thyoma tumor cell growth and increased numbers of CD4+ and CD8+ T cells [195]. Wild type mice treated with an injection of recombinant shCD5 also had reduced tumor growth [195]. Finally, CD5-deficient mice engrafted with B16-F10 melanoma cells had slower tumor growth compared to wild type C57BL/6 mice [196]. This evidence suggests that CD5, along with PD-1 and CTLA-4, may be a potential target to specifically modulate T cell Ca2+ mobilization in an immunosuppressive tumor setting.
4.4. Microbiome
The gut microbiome, including the bacteria and their products, forms a dynamic beneficial symbiosis with the immune system influencing host genes and cellular response. The gut microbiome shapes and directs immune responses while the immune system dictates the bacterial composition of the gut microbiome [197]. As the gut is the major symbiotic system intersecting the immune system and microbiota, understanding their connection has implications for immune system development and function as the gut microbiome is involved in protecting against pathogens, influencing states of inflammation, and even affecting cancer patient outcomes [198,199].
The gut microbiome primes immune responses [200]. Alteration in the microbial composition can induce changes in T cell function in infectious disease, autoimmunity, and cancer [201]. For example, mice treated with antibiotics which restrict or reduce the microbial environment exhibit impaired immune response because their T cells have altered TCR signaling and compromised intracellular Ca2+ mobilization in infectious disease and cystic fibrosis models [202,203,204]. In contrast, administering oral antibiotics to mice with EAE increases the frequency of CD5+ B cell subpopulations in distal lymphoid sites and confers disease protection [205]. In cancer, the microbiome also influences patient response to immune checkpoint inhibitors such as CTLA-4 and PD-1 [206,207]. Mice and melanoma patients immunized or populated with Bacteriodes fragilis respond better to treatment with Ipilimumab, a monoclonal antibody against CTLA-4 [198]. Similarly, tumor-specific immunity improved when anti-PD-1/PD-L1 monoclonal antibodies where used in the presence of Bifidobacterium [208].
Though little is known about how CD5 influences T cell interaction with the microbiome, some tantalizing details are available. As specific bacterium promotes cancer regression during CTLA-4 and PD-1 checkpoint blockades, a CD5 blockade in conjunction with bacterial selection may also improve immune response. Such studies would lead to novel immunotherapeutic treatments for cancer and autoimmune diseases.
5. Conclusions
CD5, widely known as an inhibitory co-receptor in the thymus, appears to modulate the signaling intensity of peripheral T cells by increasing Ca2+ signaling activity and efficacy of CD5hi T cells. CD5 expression levels in the periphery correlates with intracellular Ca2+ mobilization, suggesting that CD5 promotes peripheral T cell activation and immune response. As such, CD5 may be a novel checkpoint therapy to regulate T cell activation and metabolism through altering Ca2+ mobilization, and could be used to affect neurological behavior, alter microbiome interactions, and treat cancer and autoinflammatory diseases. While this paper focuses on the role of co-receptor CD5 effects on calcium signaling and activation of T cells, CD5 itself may be regulated through posttranslational modifications, such as N-glycosylation, which may affect Ca2+ mobilization, T cell metabolism, activation, and function. In the future it would be interesting to determine the role of other posttranslational modifications (e.g., N-glycosylation, S-glutathionylation, lipidation) in CD5 signaling.
Author Contributions
C.M.T.F. is the first author and wrote the manuscript, D.K.J. contributed additional material and editing help, K.S.W. helped with the plan for the manuscript and editing and is the corresponding author.
Funding
This work was supported by a National Institute of Allergy and Infectious Diseases grant (R0102063) to K.S.W. The funder had no role in preparation of the manuscript.
Acknowledgments
We thank Kiara Vaden Whitley, Jeralyn Jones Fransen, Tyler Cox and Josie Tueller for their critical reviews of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| CTLA-4 | Cytotoxic T-lymphocyte antigen 4 |
| CD | Cluster of differenciation |
| PD-1 | Programmed cell death protein 1 |
| AMP | Adenosine monophosphate |
| ATP | Adenosine triphosphate |
| CaMKK | Calmodulin-dependent protein kinase kinase |
| AMPK | AMP-activated protein kinase |
| SOCE | Store-operated calcium channels |
| CRAC | Calcium+-release-activated channel |
| STIM | Stromal interaction molecule |
| SERCA | Sarcoendoplasmic reticulum calcium transport ATPase |
| ER | Endoplasmic reticulum |
| NFAT | Nuclear factor of activated T cells |
| INF-γ | Interferon gamma |
| TNF | Tumor necrosis factor |
| IL-2 | Interleukin 2 |
| GLUT1 | Glucose transporter 1 |
| GLUT3 | Glucose transporter 3 |
| TIL | Tumor infiltrating lymphocytes |
| ERK | Extracellular signal-regulated kinases |
References
- Chen, L.; Flies, D.B. Molecular mechanisms of T cell co-stimulation and co-inhibition. Nat. Rev. Immunol. 2013, 13, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Beyersdorf, N.; Kerkau, T.; Hünig, T. CD28 co-stimulation in T cell homeostasis: A recent perspective. Immunotargets Ther. 2015, 4, 111–122. [Google Scholar] [PubMed]
- Fracchia, K.M.; Pai, C.Y.; Walsh, C.M. Modulation of T cell metabolism and function through calcium signaling. Front. Immunol. 2013, 4, 324. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.J.; Lafferty, K.J. Letter: Cellular proliferation can be an unreliable index of immune competence. J. Immunol. 1974, 112, 436–437. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, T.; Yamashita, M. The TCR-mediated signaling pathways that control the direction of helper T cell differentiation. Semin. Immunol. 2010, 22, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Goral, S. The three-signal hypothesis of lymphocyte activation/targets for immunosuppression. Dial. Transplant. 2011, 40, 14–16. [Google Scholar] [CrossRef]
- Pennock, N.D.; White, J.T.; Cross, E.W.; Cheney, E.E.; Tamburini, B.A.; Kedl, R.M. T cell responses: Naïve to memory and everything in between. Adv. Physiol. Educ. 2013, 37, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, A.H.; Abbas, A.K. T cell costimulation—Biology, therapeutic potential, and challenges. N. Engl. J. Med. 2006, 355, 973–975. [Google Scholar] [CrossRef] [PubMed]
- Artyomov, M.N.; Lis, M.; Devadas, S.; Davis, M.M.; Chakraborty, A.K. CD4 and CD8 binding to MHC molecules primarily acts to enhance LCK delivery. Proc. Natl. Acad. Sci. USA 2010, 107, 16916–16921. [Google Scholar] [CrossRef] [PubMed]
- Ravetch, J.V.; Lanier, L.L. Immune inhibitory receptors. Science 2000, 290, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Mellman, I.; Coukos, G.; Dranoff, G. Cancer immunotherapy comes of age. Nature 2011, 480, 480. [Google Scholar] [CrossRef] [PubMed]
- Fuertes Marraco, S.A.; Neubert, N.J.; Verdeil, G.; Speiser, D.E. Inhibitory receptors beyond T cell exhaustion. Front. Immunol. 2015, 6, 310. [Google Scholar] [CrossRef] [PubMed]
- Barrow, A.D.; Trowsdale, J. You say ITAM and I say ITIM, let’s call the whole thing off: The ambiguity of immunoreceptor signalling. Eur. J. Immunol. 2006, 36, 1646–1653. [Google Scholar] [CrossRef] [PubMed]
- Esensten, J.H.; Helou, Y.A.; Chopra, G.; Weiss, A.; Bluestone, J.A. CD28 costimulation: From mechanism to therapy. Immunity 2016, 44, 973–988. [Google Scholar] [CrossRef] [PubMed]
- Dilek, N.; Poirier, N.; Hulin, P.; Coulon, F.; Mary, C.; Ville, S.; Vie, H.; Clémenceau, B.; Blancho, G.; Vanhove, B. Targeting CD28, CTLA-4 and PD-L1 costimulation differentially controls immune synapses and function of human regulatory and conventional t cells. PLoS ONE 2013, 8, e83139. [Google Scholar] [CrossRef] [PubMed]
- Chambers, C.A.; Sullivan, T.J.; Allison, J.P. Lymphoproliferation in CTLA-4-deficient mice is mediated by costimulation-dependent activation of CD4+ T cells. Immunity 1997, 7, 885–895. [Google Scholar] [CrossRef]
- Lindsten, T.; Lee, K.P.; Harris, E.S.; Petryniak, B.; Craighead, N.; Reynolds, P.J.; Lombard, D.B.; Freeman, G.J.; Nadler, L.M.; Gray, G.S.; et al. Characterization of CTLA-4 structure and expression on human T cells. J. Immunol. 1993, 151, 3489–3499. [Google Scholar] [PubMed]
- Boise, L.H.; Minn, A.J.; Noel, P.J.; June, C.H.; Accavitti, M.A.; Lindsten, T.; Thompson, C.B. CD28 costimulation can promote T cell survival by enhancing the expression of Bcl-XL. Immunity 1995, 3, 87–98. [Google Scholar] [CrossRef]
- Buchbinder, E.I.; Desai, A. CTLA-4 and PD-1 pathways: Similarities, differences, and implications of their inhibition. Am. J. Clin. Oncol. 2016, 39, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Iwai, Y.; Hamanishi, J.; Chamoto, K.; Honjo, T. Cancer immunotherapies targeting the PD-1 signaling pathway. J. Biomed. Sci. 2017, 24, 26. [Google Scholar] [CrossRef] [PubMed]
- Chambers, C.A.; Kuhns, M.S.; Egen, J.G.; Allison, J.P. CTLA-4-mediated inhibition in regulation of T cell responses: Mechanisms and manipulation in tumor immunotherapy. Annu. Rev. Immunol. 2001, 19, 565–594. [Google Scholar] [CrossRef] [PubMed]
- Brossard, C.; Semichon, M.; Trautmann, A.; Bismuth, G. CD5 inhibits signaling at the immunological synapse without impairing its formation. J. Immunol. 2003, 170, 4623–4629. [Google Scholar] [CrossRef] [PubMed]
- Tabbekh, M.; Mokrani-Hammani, M.B.; Bismuth, G.; Mami-Chouaib, F. T cell modulatory properties of CD5 and its role in antitumor immune responses. Oncoimmunology 2013, 2, e22841. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, K.M.; Freeman, G.J.; McDermott, D.F. The next immune-checkpoint inhibitors: PD-1/PD-L1 blockade in melanoma. Clin. Ther. 2015, 37, 764–782. [Google Scholar] [CrossRef] [PubMed]
- De Wit, J.; Souwer, Y.; van Beelen, A.J.; de Groot, R.; Muller, F.J.; Klaasse Bos, H.; Jorritsma, T.; Kapsenberg, M.L.; de Jong, E.C.; van Ham, S.M. CD5 costimulation induces stable Th17 development by promoting IL-23R expression and sustained STAT3 activation. Blood 2011, 118, 6107–6114. [Google Scholar] [CrossRef] [PubMed]
- Perez-Villar, J.J.; Whitney, G.S.; Bowen, M.A.; Hewgill, D.H.; Aruffo, A.A.; Kanner, S.B. CD5 negatively regulates the T cell antigen receptor signal transduction pathway: Involvement of SH2-containing phosphotyrosine phosphatase SHP-1. Mol. Cell. Biol. 1999, 19, 2903–2912. [Google Scholar] [CrossRef] [PubMed]
- Gary-Gouy, H.; Harriague, J.; Dalloul, A.; Donnadieu, E.; Bismuth, G. CD5-negative regulation of B cell receptor signaling pathways originates from tyrosine residue Y429 outside an immunoreceptor tyrosine-based inhibitory motif. J. Immunol. 2002, 168, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Dennehy, K.M.; Broszeit, R.; Garnett, D.; Durrheim, G.A.; Spruyt, L.L.; Beyers, A.D. Thymocyte activation induces the association of phosphatidylinositol 3-kinase and pp120 with CD5. Eur. J. Immunol. 1997, 27, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Samelson, L.E. Signal transduction mediated by the T cell antigen receptor: The role of adapter proteins. Annu. Rev. Immunol. 2002, 20, 371–394. [Google Scholar] [CrossRef] [PubMed]
- Burgess, K.E.; Yamamoto, M.; Prasad, K.V.S.; Rudd, C.E. CD5 acts as a tyrosine kinase substrate within a receptor complex comprising T cell receptor ζ-chain CD3 and protein-tyrosine kinases P56LCK and P59FYN. Proc. Natl. Acad. Sci. USA 1992, 89, 9311–9315. [Google Scholar] [CrossRef] [PubMed]
- Consuegra-Fernandez, M.; Aranda, F.; Simoes, I.; Orta, M.; Sarukhan, A.; Lozano, F. CD5 as a Target for Immune-Based Therapies. Crit. Rev. Immunol. 2015, 35, 85–115. [Google Scholar] [CrossRef] [PubMed]
- Roa, N.S.; Ordonez-Rueda, D.; Chavez-Rios, J.R.; Raman, C.; Garcia-Zepeda, E.A.; Lozano, F.; Soldevila, G. The carboxy-terminal region of CD5 is required for c-CBL mediated TCR signaling downmodulation in thymocytes. Biochem. Biophys. Res. Commun. 2013, 432, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Berney, S.M.; Schaan, T.; Wolf, R.E.; Kimpel, D.L.; van der Heyde, H.; Atkinson, T.P. CD5 (OKT1) augments CD3-mediated intracellular signaling events in human T lymphocytes. Inflammation 2001, 25, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Azzam, H.S.; DeJarnette, J.B.; Huang, K.; Emmons, R.; Park, C.-S.; Sommers, C.L.; El-Khoury, D.; Shores, E.W.; Love, P.E. Fine tuning of TCR signaling by CD5. J. Immunol. 2001, 166, 5464–5472. [Google Scholar] [CrossRef] [PubMed]
- Freitas, C.M.T.; Hamblin, G.J.; Raymond, C.M.; Weber, K.S. Naive helper T cells with high CD5 expression have increased calcium signaling. PLoS ONE 2017, 12, e0178799. [Google Scholar] [CrossRef] [PubMed]
- Feske, S. Calcium signalling in lymphocyte activation and disease. Nat. Rev. Immunol. 2007, 7, 690–702. [Google Scholar] [CrossRef] [PubMed]
- Joseph, N.; Reicher, B.; Barda-Saad, M. The calcium feedback loop and T cell activation: How cytoskeleton networks control intracellular calcium flux. Biochim. Biophys. Acta Biomembr. 2014, 1838, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Vig, M.; Kinet, J.-P. Calcium signaling in immune cells. Nat. Immunol. 2009, 10, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Wolf, I.M.A.; Guse, A.H. Ca2+ microdomains in T-lymphocytes. Front. Oncol. 2017, 7, 73. [Google Scholar] [CrossRef] [PubMed]
- Hogan, P.G.; Lewis, R.S.; Rao, A. Molecular basis of calcium signaling in lymphocytes: STIM and ORAI. Annu. Rev. Immunol. 2010, 28, 491–533. [Google Scholar] [CrossRef] [PubMed]
- Oh-hora, M.; Rao, A. Calcium signaling in lymphocytes. Curr. Opin. Immunol. 2008, 20, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A., Jr. The co-receptor function of CD4. Semin. Immunol. 1991, 3, 153–160. [Google Scholar] [PubMed]
- Moran, A.E.; Hogquist, K.A. T cell receptor affinity in thymic development. Immunology 2012, 135, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Kyttaris, V.C.; Zhang, Z.; Kampagianni, O.; Tsokos, G.C. Calcium signaling in systemic lupus erythematosus T cells: A treatment target. Arthritis Rheum. 2011, 63, 2058–2066. [Google Scholar] [CrossRef] [PubMed]
- Demkow, U.; Winklewski, P.; Ciepiela, O.; Popko, K.; Lipinska, A.; Kucharska, A.; Michalska, B.; Wasik, M. Modulatory effect of insulin on T cell receptor mediated calcium signaling is blunted in long lasting type 1 diabetes mellitus. Pharmacol. Rep. 2012, 64, 150–156. [Google Scholar] [CrossRef]
- Parry, R.V.; Chemnitz, J.M.; Frauwirth, K.A.; Lanfranco, A.R.; Braunstein, I.; Kobayashi, S.V.; Linsley, P.S.; Thompson, C.B.; Riley, J.L. CTLA-4 and PD-1 receptors inhibit T cell activation by distinct mechanisms. Mol. Cell. Biol. 2005, 25, 9543–9553. [Google Scholar] [CrossRef] [PubMed]
- Grosso, J.F.; Jure-Kunkel, M.N. CTLA-4 blockade in tumor models: An overview of preclinical and translational research. Cancer Immun. 2013, 13, 5. [Google Scholar] [PubMed]
- Rudd, C.E.; Taylor, A.; Schneider, H. CD28 and CTLA-4 coreceptor expression and signal transduction. Immunol. Rev. 2009, 229, 12–26. [Google Scholar] [CrossRef] [PubMed]
- Jago, C.B.; Yates, J.; Olsen Saraiva CÂMara, N.; Lechler, R.I.; Lombardi, G. Differential expression of CTLA-4 among T cell subsets. Clin. Exp. Immunol. 2004, 136, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Linsley, P.S.; Bradshaw, J.; Greene, J.; Peach, R.; Bennett, K.L.; Mittler, R.S. Intracellular trafficking of CTLA-4 and focal localization towards sites of TCR engagement. Immunity 1996, 4, 535–543. [Google Scholar] [CrossRef]
- Schneider, H.; Smith, X.; Liu, H.; Bismuth, G.; Rudd, C.E. CTLA-4 disrupts ZAP70 microcluster formation with reduced T cell/APC dwell times and calcium mobilization. Eur. J. Immunol. 2008, 38, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Grohmann, U.; Orabona, C.; Fallarino, F.; Vacca, C.; Calcinaro, F.; Falorni, A.; Candeloro, P.; Belladonna, M.L.; Bianchi, R.; Fioretti, M.C.; et al. CTLA-4-Ig regulates tryptophan catabolism in vivo. Nat. Immunol. 2002, 3, 1097–1101. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Jin, W.; Wahl, S.M. Engagement of cytotoxic T lymphocyte-associated antigen 4 (CTLA-4) induces transforming growth factor beta (TGF-β) production by murine CD4+ T cells. J. Exp. Med. 1998, 188, 1849–1857. [Google Scholar] [CrossRef] [PubMed]
- Hryniewicz, A.; Boasso, A.; Edghill-Smith, Y.; Vaccari, M.; Fuchs, D.; Venzon, D.; Nacsa, J.; Betts, M.R.; Tsai, W.-P.; Heraud, J.-M.; et al. CTLA-4 blockade decreases TGF-β, IDO, and viral RNA expression in tissues of SIVmac251-infected macaques. Blood 2006, 108, 3834–3842. [Google Scholar] [CrossRef] [PubMed]
- Iken, K.; Liu, K.; Liu, H.; Bizargity, P.; Wang, L.; Hancock, W.W.; Visner, G.A. Indoleamine 2,3-dioxygenase and metabolites protect murine lung allografts and impair the calcium mobilization of T cells. Am. J. Respir. Cell Mol. Biol. 2012, 47, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.S.K.; Sansom, D.M. Confusing signals: Recent progress in CTLA-4 biology. Trends Immunol. 2015, 36, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Cederbom, L.; Hall, H.; Ivars, F. CD4+CD25+ regulatory T cells down-regulate co-stimulatory molecules on antigen-presenting cells. Eur. J. Immunol. 2000, 30, 1538–1543. [Google Scholar] [CrossRef]
- Burnett, D.L.; Parish, I.A.; Masle-Farquhar, E.; Brink, R.; Goodnow, C.C. Murine LRBA deficiency causes CTLA-4 deficiency in Tregs without progression to immune dysregulation. Immunol. Cell Biol. 2017, 95, 775–778. [Google Scholar] [CrossRef] [PubMed]
- Verma, N.; Burns, S.O.; Walker, L.S.K.; Sansom, D.M. Immune deficiency and autoimmunity in patients with CTLA-4 (CD152) mutations. Clin. Exp. Immunol. 2017, 190, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wing, K.; Onishi, Y.; Prieto-Martin, P.; Yamaguchi, T.; Miyara, M.; Fehervari, Z.; Nomura, T.; Sakaguchi, S. CTLA-4 control over Foxp3+ regulatory T cell function. Science 2008, 322, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Sojka, D.K.; Hughson, A.; Fowell, D.J. CTLA-4 is Required by CD4+CD25+ treg to control CD4+ T cell lymphopenia-induced proliferation. Eur. J. Immunol. 2009, 39, 1544–1551. [Google Scholar] [CrossRef] [PubMed]
- Kristiansen, O.P.; Larsen, Z.M.; Pociot, F. CTLA-4 in autoimmune diseases--a general susceptibility gene to autoimmunity? Genes Immun. 2000, 1, 170–184. [Google Scholar] [CrossRef] [PubMed]
- Chikuma, S. CTLA-4, an essential immune-checkpoint for T cell activation. Curr. Top. Microbiol. Immunol. 2017, 410, 99–126. [Google Scholar] [PubMed]
- Tai, X.; Van Laethem, F.; Pobezinsky, L.; Guinter, T.; Sharrow, S.O.; Adams, A.; Granger, L.; Kruhlak, M.; Lindsten, T.; Thompson, C.B.; et al. Basis of CTLA-4 function in regulatory and conventional CD4+ T cells. Blood 2012, 119, 5155–5163. [Google Scholar] [CrossRef] [PubMed]
- Lo, B.; Abdel-Motal, U.M. Lessons from CTLA-4 deficiency and checkpoint inhibition. Curr. Opin. Immunol. 2017, 49, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Avogadri, F.; Yuan, J.; Yang, A.; Schaer, D.; Wolchok, J.D. Modulation of CTLA-4 and GITR for cancer immunotherapy. Curr. Top. Microbiol. Immunol. 2011, 344, 211–244. [Google Scholar] [PubMed]
- Royal, R.E.; Levy, C.; Turner, K.; Mathur, A.; Hughes, M.; Kammula, U.S.; Sherry, R.M.; Topalian, S.L.; Yang, J.C.; Lowy, I.; et al. Phase 2 trial of single agent Ipilimumab (anti-CTLA-4) for locally advanced or metastatic pancreatic adenocarcinoma. J. Immunother. 2010, 33, 828–833. [Google Scholar] [CrossRef] [PubMed]
- Le, D.T.; Lutz, E.; Uram, J.N.; Sugar, E.A.; Onners, B.; Solt, S.; Zheng, L.; Diaz, L.A., Jr.; Donehower, R.C.; Jaffee, E.M.; et al. Evaluation of ipilimumab in combination with allogeneic pancreatic tumor cells transfected with a GM-CSF gene in previously treated pancreatic cancer. J. Immunother. 2013, 36, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.Y.; Gore, I.; Fong, L.; Venook, A.; Beck, S.B.; Dorazio, P.; Criscitiello, P.J.; Healey, D.I.; Huang, B.; Gomez-Navarro, J.; et al. Phase II study of the anti-cytotoxic T-lymphocyte-associated antigen 4 monoclonal antibody, tremelimumab, in patients with refractory metastatic colorectal cancer. J. Clin. Oncol. 2010, 28, 3485–3490. [Google Scholar] [CrossRef] [PubMed]
- Ribas, A.; Camacho, L.H.; Lopez-Berestein, G.; Pavlov, D.; Bulanhagui, C.A.; Millham, R.; Comin-Anduix, B.; Reuben, J.M.; Seja, E.; Parker, C.A.; et al. Antitumor activity in melanoma and anti-self responses in a phase I trial with the anti-cytotoxic T lymphocyte-associated antigen 4 monoclonal antibody CP-675,206. J. Clin. Oncol. 2005, 23, 8968–8977. [Google Scholar] [CrossRef] [PubMed]
- Calabro, L.; Morra, A.; Fonsatti, E.; Cutaia, O.; Fazio, C.; Annesi, D.; Lenoci, M.; Amato, G.; Danielli, R.; Altomonte, M.; et al. Efficacy and safety of an intensified schedule of tremelimumab for chemotherapy-resistant malignant mesothelioma: An open-label, single-arm, phase 2 study. Lancet Respir. Med. 2015, 3, 301–309. [Google Scholar] [CrossRef]
- Comin-Anduix, B.; Escuin-Ordinas, H.; Ibarrondo, F.J. Tremelimumab: Research and clinical development. OncoTargets Ther. 2016, 9, 1767–1776. [Google Scholar]
- Ribas, A.; Comin-Anduix, B.; Chmielowski, B.; Jalil, J.; de la Rocha, P.; McCannel, T.A.; Ochoa, M.T.; Seja, E.; Villanueva, A.; Oseguera, D.K.; et al. Dendritic cell vaccination combined with CTLA4 blockade in patients with metastatic melanoma. Clin. Cancer Res. 2009, 15, 6267–6276. [Google Scholar] [CrossRef] [PubMed]
- Antonia, S.; Goldberg, S.B.; Balmanoukian, A.; Chaft, J.E.; Sanborn, R.E.; Gupta, A.; Narwal, R.; Steele, K.; Gu, Y.; Karakunnel, J.J.; et al. Safety and antitumour activity of durvalumab plus tremelimumab in non-small cell lung cancer: A multicentre, phase 1b study. Lancet Oncol. 2016, 17, 299–308. [Google Scholar] [CrossRef]
- Dong, Y.; Sun, Q.; Zhang, X. PD-1 and its ligands are important immune checkpoints in cancer. Oncotarget 2017, 8, 2171–2186. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Chen, S.; Yang, L.; Li, Y. The role of PD-1 and PD-L1 in T cell immune suppression in patients with hematological malignancies. J. Hematol. Oncol. 2013, 6, 74. [Google Scholar] [CrossRef] [PubMed]
- Keir, M.E.; Butte, M.J.; Freeman, G.J.; Sharpe, A.H. PD-1 and its ligands in tolerance and immunity. Annu. Rev. Immunol. 2008, 26, 677–704. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, T.; Wang, J. PD-1/PD-L pathway and autoimmunity. Autoimmunity 2005, 38, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Boussiotis, V.A. Molecular and biochemical aspects of the PD-1 checkpoint pathway. N. Engl. J. Med. 2016, 375, 1767–1778. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-F.; Fouquet, S.; Chapon, M.; Salmon, H.; Regnier, F.; Labroquère, K.; Badoual, C.; Damotte, D.; Validire, P.; Maubec, E.; et al. Early T cell signalling is reversibly altered in PD-1+ T lymphocytes infiltrating human tumors. PLoS ONE 2011, 6, e17621. [Google Scholar] [CrossRef] [PubMed]
- Gorentla, B.K.; Zhong, X.-P. T cell receptor signal transduction in T lymphocytes. J. Clin. Cell. Immunol. 2012, 2012, 005. [Google Scholar]
- Wei, F.; Zhong, S.; Ma, Z.; Kong, H.; Medvec, A.; Ahmed, R.; Freeman, G.J.; Krogsgaard, M.; Riley, J.L. Strength of PD-1 signaling differentially affects T cell effector functions. Proc. Natl. Acad. Sci. USA 2013, 110, E2480–E2489. [Google Scholar] [CrossRef] [PubMed]
- Cochain, C.; Chaudhari, S.M.; Koch, M.; Wiendl, H.; Eckstein, H.-H.; Zernecke, A. Programmed cell death-1 deficiency exacerbates T cell activation and atherogenesis despite expansion of regulatory T cells in atherosclerosis-prone mice. PLoS ONE 2014, 9, e93280. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Kishi, Y.; Meguri, Y.; Yoshioka, T.; Iwamoto, M.; Maeda, Y.; Yagita, H.; Tanimoto, M.; Koreth, J.; Ritz, J.; et al. PD-1 signaling has a critical role in maintaining regulatory T cell homeostasis; implication for treg depletion therapy by PD-1 blockade. Blood 2015, 126, 848. [Google Scholar]
- Francisco, L.M.; Sage, P.T.; Sharpe, A.H. The PD-1 pathway in tolerance and autoimmunity. Immunol. Rev. 2010, 236, 219–242. [Google Scholar] [CrossRef] [PubMed]
- Miyajima, M.; Zhang, B.; Sugiura, Y.; Sonomura, K.; Guerrini, M.M.; Tsutsui, Y.; Maruya, M.; Vogelzang, A.; Chamoto, K.; Honda, K.; et al. Metabolic shift induced by systemic activation of T cells in PD-1-deficient mice perturbs brain monoamines and emotional behavior. Nat. Immunol. 2017, 18, 1342–1352. [Google Scholar] [CrossRef] [PubMed]
- Riella, L.V.; Paterson, A.M.; Sharpe, A.H.; Chandraker, A. Role of the PD-1 pathway in the immune response. Am. J. Transplant. 2012, 12, 2575–2587. [Google Scholar] [CrossRef] [PubMed]
- Kroner, A.; Mehling, M.; Hemmer, B.; Rieckmann, P.; Toyka, K.V.; Maurer, M.; Wiendl, H. A PD-1 polymorphism is associated with disease progression in multiple sclerosis. Ann. Neurol 2005, 58, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Pawlak-Adamska, E.; Nowak, O.; Karabon, L.; Pokryszko-Dragan, A.; Partyka, A.; Tomkiewicz, A.; Ptaszkowski, J.; Frydecka, I.; Podemski, R.; Dybko, J.; et al. PD-1 gene polymorphic variation is linked with first symptom of disease and severity of relapsing-remitting form of MS. J. Neuroimmunol. 2017, 305, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Jia, R.; Zhang, X.; Fang, Q.; Huang, L. The PD-1/PD-Ls pathway and autoimmune diseases. Cell. Immunol. 2014, 290, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Gianchecchi, E.; Delfino, D.V.; Fierabracci, A. Recent insights into the role of the PD-1/PD-L1 pathway in immunological tolerance and autoimmunity. Autoimmun. Rev. 2013, 12, 1091–1100. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yoshida, T.; Nakaki, F.; Hiai, H.; Okazaki, T.; Honjo, T. Establishment of NOD-Pdcd1−/− mice as an efficient animal model of type I diabetes. Proc. Natl. Acad. Sci. USA 2005, 102, 11823–11828. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, T.; Otaka, Y.; Wang, J.; Hiai, H.; Takai, T.; Ravetch, J.V.; Honjo, T. Hydronephrosis associated with antiurothelial and antinuclear autoantibodies in BALB/c-Fcgr2b−/−Pdcd1−/− mice. J. Exp. Med. 2005, 202, 1643–1648. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, H.; Okazaki, T.; Tanaka, Y.; Nakatani, K.; Hara, M.; Matsumori, A.; Sasayama, S.; Mizoguchi, A.; Hiai, H.; Minato, N.; et al. Autoimmune dilated cardiomyopathy in PD-1 receptor-deficient mice. Science 2001, 291, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Yu, S.; Zhu, B.; Bedoret, D.; Bu, X.; Francisco, L.M.; Hua, P.; Duke-Cohan, J.S.; Umetsu, D.T.; Sharpe, A.H.; et al. RGMb is a novel binding partner for PD-L2 and its engagement with PD-L2 promotes respiratory tolerance. J. Exp. Med. 2014, 211, 943–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, K.; Kishimoto, T. CD5: A new partner for IL-6. Immunity 2016, 44, 720–722. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.J.; Jones, N.H.; Strominger, J.L.; Herzenberg, L.A. Molecular cloning of Ly-1, a membrane glycoprotein of mouse T lymphocytes and a subset of B cells: Molecular homology to its human counterpart Leu-1/T1 (CD5). Proc. Natl. Acad. Sci. USA 1987, 84, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Tarakhovsky, A.; Kanner, S.B.; Hombach, J.; Ledbetter, J.A.; Muller, W.; Killeen, N.; Rajewsky, K. A role for CD5 in TCR-mediated signal transduction and thymocyte selection. Science 1995, 269, 535–537. [Google Scholar] [CrossRef] [PubMed]
- Dalloul, A. CD5: A safeguard against autoimmunity and a shield for cancer cells. Autoimmun. Rev. 2009, 8, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Bhandoola, A.; Bosselut, R.; Yu, Q.; Cowan, M.L.; Feigenbaum, L.; Love, P.E.; Singer, A. CD5-mediated inhibition of TCR signaling during intrathymic selection and development does not require the CD5 extracellular domain. Eur. J. Immunol. 2002, 32, 1811–1817. [Google Scholar] [CrossRef]
- Mandl, J.N.; Monteiro, J.P.; Vrisekoop, N.; Germain, R.N. T cell positive selection uses self-ligand binding strength to optimize repertoire recognition of foreign antigens. Immunity 2013, 38, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Henderson, J.G.; Opejin, A.; Jones, A.; Gross, C.; Hawiger, D. CD5 instructs extrathymic regulatory T cell development in response to self and tolerizing antigens. Immunity 2015, 42, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Gringhuis, S.I.; de Leij, L.F.; Wayman, G.A.; Tokumitsu, H.; Vellenga, E. The Ca2+/calmodulin-dependent kinase type IV is involved in the CD5-mediated signaling pathway in human T lymphocytes. J. Biol. Chem. 1997, 272, 31809–31820. [Google Scholar] [CrossRef] [PubMed]
- Hassan, N.J.; Simmonds, S.J.; Clarkson, N.G.; Hanrahan, S.; Puklavec, M.J.; Bomb, M.; Barclay, A.N.; Brown, M.H. CD6 regulates T cell responses through activation-dependent recruitment of the positive regulator SLP-76. Mol. Cell. Biol. 2006, 26, 6727–6738. [Google Scholar] [CrossRef] [PubMed]
- Pena-Rossi, C.; Zuckerman, L.A.; Strong, J.; Kwan, J.; Ferris, W.; Chan, S.; Tarakhovsky, A.; Beyers, A.D.; Killeen, N. Negative regulation of CD4 lineage development and responses by CD5. J. Immunol. 1999, 163, 6494–6501. [Google Scholar] [PubMed]
- Davies, A.A.; Ley, S.C.; Crumpton, M.J. CD5 is phosphorylated on tyrosine after stimulation of the T cell antigen receptor complex. Proc. Natl. Acad. Sci. USA 1992, 89, 6368–6372. [Google Scholar] [CrossRef] [PubMed]
- Samelson, L.E.; Phillips, A.F.; Luong, E.T.; Klausner, R.D. Association of the fyn protein-tyrosine kinase with the T cell antigen receptor. Proc. Natl. Acad. Sci. USA 1990, 87, 4358–4362. [Google Scholar] [CrossRef] [PubMed]
- Raab, M.; Yamamoto, M.; Rudd, C.E. The T cell antigen CD5 acts as a receptor and substrate for the protein-tyrosine kinase p56lck. Mol.Cell. Biol. 1994, 14, 2862–2870. [Google Scholar] [CrossRef] [PubMed]
- Beyers, A.D.; Spruyt, L.L.; Williams, A.F. Molecular associations between the T-lymphocyte antigen receptor complex and the surface antigens CD2, CD4, or CD8 and CD5. Proc. Natl. Acad. Sci. USA 1992, 89, 2945–2949. [Google Scholar] [CrossRef] [PubMed]
- Spertini, F.; Stohl, W.; Ramesh, N.; Moody, C.; Geha, R.S. Induction of human T cell proliferation by a monoclonal antibody to CD5. J. Immunol. 1991, 146, 47–52. [Google Scholar] [PubMed]
- Persaud, S.P.; Parker, C.R.; Lo, W.-L.; Weber, K.S.; Allen, P.M. Intrinsic CD4+ T cell sensitivity and response to pathogen are set and sustained by avidity for thymic and peripheral self-pMHC. Nat. Immunol. 2014, 15, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Calvo, J.; Padilla, O.; Places, L.; Vigorito, E.; Vila, J.M.; Vilella, R.; Mila, J.; Vives, J.; Bowen, M.A.; Lozano, F. Relevance of individual CD5 extracellular domains on antibody recognition, glycosylation and co-mitogenic signalling. Tissue Antigen. 1999, 54, 16–26. [Google Scholar] [CrossRef]
- McAlister, M.S.; Brown, M.H.; Willis, A.C.; Rudd, P.M.; Harvey, D.J.; Aplin, R.; Shotton, D.M.; Dwek, R.A.; Barclay, A.N.; Driscoll, P.C. Structural analysis of the CD5 antigen—Expression, disulphide bond analysis and physical characterisation of CD5 scavenger receptor superfamily domain 1. Eur J. Biochem. 1998, 257, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.-H.; Kim, H.-O.; Surh, C.D.; Sprent, J. T cell receptor-dependent regulation of lipid rafts controls naive CD8+ T cell homeostasis. Immunity 2010, 32, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Yashiro-Ohtani, Y.; Zhou, X.-Y.; Toyo-oka, K.; Tai, X.-G.; Park, C.-S.; Hamaoka, T.; Abe, R.; Miyake, K.; Fujiwara, H. Non-CD28 costimulatory molecules present in T cell rafts induce T cell costimulation by enhancing the association of TCR with rafts. J. Immunol. 2000, 164, 1251–1259. [Google Scholar] [CrossRef] [PubMed]
- König, R. Chapter 315—Signal Transduction in T Lymphocytes A2—Bradshaw, Ralph A. In Handbook of Cell Signaling, 2nd ed.; Dennis, E.A., Ed.; Academic Press: San Diego, CA, USA, 2010; pp. 2679–2688. [Google Scholar]
- Milam, A.V.; Allen, P.M. Functional heterogeneity in CD4+ T cell responses against a bacterial pathogen. Front. Immunol 2015, 6, 621. [Google Scholar] [CrossRef] [PubMed]
- Lozano, F.; Simarro, M.; Calvo, J.; Vila, J.M.; Padilla, O.; Bowen, M.A.; Campbell, K.S. CD5 signal transduction: Positive or negative modulation of antigen receptor signaling. Crit. Rev. Immunol. 2000, 20, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Hogquist, K.A.; Jameson, S.C. The self-obsession of T cells: How TCR signaling thresholds affect fate decisions in the thymus and effector function in the periphery. Nat. Immunol. 2014, 15, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Van de Velde, H.; von Hoegen, I.; Luo, W.; Parnes, J.R.; Thielemans, K. The B-cell surface protein CD72/Lyb-2 is the ligand for CD5. Nature 1991, 351, 662–665. [Google Scholar] [CrossRef] [PubMed]
- Biancone, L.; Bowen, M.A.; Lim, A.; Aruffo, A.; Andres, G.; Stamenkovic, I. Identification of a novel inducible cell-surface ligand of CD5 on activated lymphocytes. J. Exp. Med. 1996, 184, 811–819. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.H.; Lacey, E. A ligand for CD5 is CD5. J. Immunol. 2010, 185, 6068–6074. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Van de Velde, H.; von Hoegen, I.; Parnes, J.R.; Thielemans, K. Ly-1 (CD5), a membrane glycoprotein of mouse T lymphocytes and a subset of B cells, is a natural ligand of the B cell surface protein Lyb-2 (CD72). J. Immunol. 1992, 148, 1630–1634. [Google Scholar] [PubMed]
- Vandenberghe, P.; Verwilghen, J.; Van Vaeck, F.; Ceuppens, J.L. Ligation of the CD5 or CD28 molecules on resting human T cells induces expression of the early activation antigen CD69 by a calcium- and tyrosine kinase-dependent mechanism. Immunology 1993, 78, 210–217. [Google Scholar] [PubMed]
- Ceuppens, J.L.; Baroja, M.L. Monoclonal antibodies to the CD5 antigen can provide the necessary second signal for activation of isolated resting T cells by solid-phase-bound OKT3. J. Immunol. 1986, 137, 1816–1821. [Google Scholar] [PubMed]
- June, C.H.; Rabinovitch, P.S.; Ledbetter, J.A. CD5 antibodies increase intracellular ionized calcium concentration in T cells. J. Immunol. 1987, 138, 2782–2792. [Google Scholar] [PubMed]
- Reth, M. Antigen receptor tail clue. Nature 1989, 338, 383–384. [Google Scholar] [CrossRef] [PubMed]
- Unkeless, J.C.; Jin, J. Inhibitory receptors, ITIM sequences and phosphatases. Curr. Opin. Immunol 1997, 9, 338–343. [Google Scholar] [CrossRef]
- Dong, B.; Somani, A.K.; Love, P.E.; Zheng, X.; Chen, X.; Zhang, J. CD5-mediated inhibition of TCR signaling proceeds normally in the absence of SHP-1. Int. J. Mol. Med. 2016, 38, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Weber, K.S.; Li, Q.J.; Persaud, S.P.; Campbell, J.D.; Davis, M.M.; Allen, P.M. Distinct CD4+ helper T cells involved in primary and secondary responses to infection. Proc. Natl. Acad. Sci. USA 2012, 109, 9511–9516. [Google Scholar] [CrossRef] [PubMed]
- Fulton, R.B.; Hamilton, S.E.; Xing, Y.; Best, J.A.; Goldrath, A.W.; Hogquist, K.A.; Jameson, S.C. The TCR’s sensitivity to self peptide–MHC dictates the ability of naive CD8+ T cells to respond to foreign antigens. Nat. Immunol. 2014, 16, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Palin, A.C.; Love, P.E. CD5 helps aspiring regulatory T cells ward off unwelcome cytokine advances. Immunity 2015, 42, 395–396. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P.; Chan, S.L. Calcium orchestrates apoptosis. Nat. Cell Biol. 2003, 5, 1041–1043. [Google Scholar] [CrossRef] [PubMed]
- Orrenius, S.; Nicotera, P. The calcium ion and cell death. J. Neural Transm. Suppl. 1994, 43, 1–11. [Google Scholar] [PubMed]
- Zhao, C.; Davies, J.D. A peripheral CD4+ T cell precursor for naive, memory, and regulatory T cells. J. Exp. Med. 2010, 207, 2883–2894. [Google Scholar] [CrossRef] [PubMed]
- Wahl, D.R.; Byersdorfer, C.A.; Ferrara, J.L.M.; Opipari, A.W.; Glick, G.D. Distinct metabolic programs in activated T cells: Opportunities for selective immunomodulation. Immunol. Rev. 2012, 249, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Pearce, E.L.; Pearce, E.J. Metabolic pathways in immune cell activation and quiescence. Immunity 2013, 38, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Van der Windt, G.J.; Pearce, E.L. Metabolic switching and fuel choice during T cell differentiation and memory development. Immunol. Rev. 2012, 249, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-H.; Curtis, J.D.; Maggi, L.B., Jr.; Faubert, B.; Villarino, A.V.; O’Sullivan, D.; Huang, S.C.-C.; van der Windt, G.J.W.; Blagih, J.; Qiu, J.; et al. Posttranscriptional control of T cell effector function by aerobic glycolysis. Cell 2013, 153, 1239–1251. [Google Scholar] [CrossRef] [PubMed]
- Almeida, L.; Lochner, M.; Berod, L.; Sparwasser, T. Metabolic pathways in T cell activation and lineage differentiation. Semin. Immunol. 2016, 28, 514–524. [Google Scholar] [CrossRef] [PubMed]
- Vaeth, M.; Maus, M.; Klein-Hessling, S.; Freinkman, E.; Yang, J.; Eckstein, M.; Cameron, S.; Turvey, S.E.; Serfling, E.; Berberich-Siebelt, F.; et al. Store-operated Ca2+ entry controls clonal expansion of T cells through metabolic reprogramming. Immunity 2017, 47, 664–679. [Google Scholar] [CrossRef] [PubMed]
- Feske, S.; Skolnik, E.Y.; Prakriya, M. Ion channels and transporters in lymphocyte function and immunity. Nat. Rev. Immunol. 2012, 12, 532–547. [Google Scholar] [CrossRef] [PubMed]
- Tamás, P.; Hawley, S.A.; Clarke, R.G.; Mustard, K.J.; Green, K.; Hardie, D.G.; Cantrell, D.A. Regulation of the energy sensor AMP-activated protein kinase by antigen receptor and Ca2+ in T lymphocytes. J. Exp. Med. 2006, 203, 1665–1670. [Google Scholar] [CrossRef] [PubMed]
- Ma, E.H.; Poffenberger, M.C.; Wong, A.H.; Jones, R.G. The role of AMPK in T cell metabolism and function. Curr. Opin. Immunol. 2017, 46, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Manning, B.D. The TSC1–TSC2 complex: A molecular switchboard controlling cell growth. Biochem. J. 2008, 412, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Chi, H. Regulation and function of mTOR signalling in T cell fate decision. Nat. Rev. Immunol. 2012, 12, 325–338. [Google Scholar] [CrossRef] [PubMed]
- MacIver, N.J.; Blagih, J.; Saucillo, D.C.; Tonelli, L.; Griss, T.; Rathmell, J.C.; Jones, R.G. The liver kinase B1 is a central regulator of T cell development, activation, and metabolism. J. Immunol. 2011, 187, 4187–4198. [Google Scholar] [CrossRef] [PubMed]
- Kirichok, Y.; Krapivinsky, G.; Clapham, D.E. The mitochondrial calcium uniporter is a highly selective ion channel. Nature 2004, 427, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Gilabert, J.A.; Bakowski, D.; Parekh, A.B. Energized mitochondria increase the dynamic range over which inositol 1,4,5-trisphosphate activates store-operated calcium influx. EMBO J. 2001, 20, 2672–2679. [Google Scholar] [CrossRef] [PubMed]
- Gilabert, J.A.; Parekh, A.B. Respiring mitochondria determine the pattern of activation and inactivation of the store-operated Ca2+ current I (CRAC). EMBO J. 2000, 19, 6401–6407. [Google Scholar] [CrossRef] [PubMed]
- Singaravelu, K.; Nelson, C.; Bakowski, D.; de Brito, O.M.; Ng, S.W.; di Capite, J.; Powell, T.; Scorrano, L.; Parekh, A.B. Mitofusin 2 regulates STIM1 migration from the Ca2+ store to the plasma membrane in cells with depolarized mitochondria. J. Biol. Chem. 2011, 286, 12189–12201. [Google Scholar] [CrossRef] [PubMed]
- Dimeloe, S.; Burgener, A.V.; Grahlert, J.; Hess, C. T cell metabolism governing activation, proliferation and differentiation; a modular view. Immunology 2017, 150, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Jouaville, L.S.; Pinton, P.; Bastianutto, C.; Rutter, G.A.; Rizzuto, R. Regulation of mitochondrial ATP synthesis by calcium: Evidence for a long-term metabolic priming. Proc. Natl. Acad. Sci. USA 1999, 96, 13807–13812. [Google Scholar] [CrossRef] [PubMed]
- Ho, P.C.; Bihuniak, J.D.; Macintyre, A.N.; Staron, M.; Liu, X.; Amezquita, R.; Tsui, Y.C.; Cui, G.; Micevic, G.; Perales, J.C.; et al. Phosphoenolpyruvate is a metabolic checkpoint of anti-tumor T cell responses. Cell 2015, 162, 1217–1228. [Google Scholar] [CrossRef] [PubMed]
- Rumi-Masante, J.; Rusinga, F.I.; Lester, T.E.; Dunlap, T.B.; Williams, T.D.; Dunker, A.K.; Weis, D.D.; Creamer, T.P. Structural basis for activation of calcineurin by calmodulin. J. Mol. Biol. 2012, 415, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Racioppi, L.; Means, A.R. Calcium/calmodulin-dependent protein kinase kinase 2: Roles in signaling and pathophysiology. J. Biol. Chem. 2012, 287, 31658–31665. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-H.; Qiu, J.; O’Sullivan, D.; Buck, M.D.; Noguchi, T.; Curtis, J.D.; Chen, Q.; Gindin, M.; Gubin, M.M.; van der Windt, G.J.W.; et al. Metabolic competition in the tumor microenvironment is a driver of cancer progression. Cell 2015, 162, 1229–1241. [Google Scholar] [CrossRef] [PubMed]
- Patsoukis, N.; Bardhan, K.; Chatterjee, P.; Sari, D.; Liu, B.; Bell, L.N.; Karoly, E.D.; Freeman, G.J.; Petkova, V.; Seth, P.; et al. PD-1 alters T cell metabolic reprogramming by inhibiting glycolysis and promoting lipolysis and fatty acid oxidation. Nat. Commun. 2015, 6, 6692. [Google Scholar] [CrossRef] [PubMed]
- Gary-Gouy, H.; Sainz-Perez, A.; Marteau, J.-B.; Marfaing-Koka, A.; Delic, J.; Merle-Beral, H.; Galanaud, P.; Dalloul, A. Natural phosphorylation of CD5 in chronic lymphocytic leukemia B cells and analysis of CD5-regulated genes in a B cell line suggest a role for CD5 in malignant phenotype. J. Immunol. 2007, 179, 4335–4344. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.J.; Mahajan, V.S.; Chen, J.; Irvine, D.J.; Lauffenburger, D.A. Signaling thresholds govern heterogeneity in IL-7-receptor-mediated responses of naive CD8+ T cells. Immunol Cell. Biol. 2011, 89, 581–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kipnis, J.; Gadani, S.; Derecki, N.C. Pro-cognitive properties of T cells. Nat. Rev. Immunol. 2012, 12, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Kipnis, J.; Cohen, H.; Cardon, M.; Ziv, Y.; Schwartz, M. T cell deficiency leads to cognitive dysfunction: Implications for therapeutic vaccination for schizophrenia and other psychiatric conditions. Proc. Natl. Acad. Sci. USA 2004, 101, 8180–8185. [Google Scholar] [CrossRef] [PubMed]
- Brombacher, T.M.; Nono, J.K.; De Gouveia, K.S.; Makena, N.; Darby, M.; Womersley, J.; Tamgue, O.; Brombacher, F. IL-13–mediated regulation of learning and memory. J. Immunol. 2017, 198, 2681–2688. [Google Scholar] [CrossRef] [PubMed]
- Oliveira-dos-Santos, A.J.; Matsumoto, G.; Snow, B.E.; Bai, D.; Houston, F.P.; Whishaw, I.Q.; Mariathasan, S.; Sasaki, T.; Wakeham, A.; Ohashi, P.S.; et al. Regulation of T cell activation, anxiety, and male aggression by RGS2. Proc. Natl. Acad. Sci. USA 2000, 97, 12272–12277. [Google Scholar] [CrossRef] [PubMed]
- Filiano, A.J.; Gadani, S.P.; Kipnis, J. How and why do T cells and their derived cytokines affect the injured and healthy brain? Nat. Rev. Neurosci. 2017, 18, 375. [Google Scholar] [CrossRef] [PubMed]
- Kyratsous, N.I.; Bauer, I.J.; Zhang, G.; Pesic, M.; Bartholomäus, I.; Mues, M.; Fang, P.; Wörner, M.; Everts, S.; Ellwart, J.W.; et al. Visualizing context-dependent calcium signaling in encephalitogenic T cells in vivo by two-photon microscopy. Proc. Natl. Acad. Sci. USA 2017, 114, E6381–E6389. [Google Scholar] [CrossRef] [PubMed]
- Smedler, E.; Uhlén, P. Frequency decoding of calcium oscillations. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 964–969. [Google Scholar] [CrossRef] [PubMed]
- Pesic, M.; Bartholomaus, I.; Kyratsous, N.I.; Heissmeyer, V.; Wekerle, H.; Kawakami, N. 2-photon imaging of phagocyte-mediated T cell activation in the CNS. J. Clin. Investig. 2013, 123, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- de Bruin, N.M.W.J.; Schmitz, K.; Schiffmann, S.; Tafferner, N.; Schmidt, M.; Jordan, H.; Häußler, A.; Tegeder, I.; Geisslinger, G.; Parnham, M.J. Multiple rodent models and behavioral measures reveal unexpected responses to FTY720 and DMF in experimental autoimmune encephalomyelitis. Behav. Brain Res. 2016, 300, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Schub, D.; Janssen, E.; Leyking, S.; Sester, U.; Assmann, G.; Hennes, P.; Smola, S.; Vogt, T.; Rohrer, T.; Sester, M.; et al. Altered phenotype and functionality of varicella zoster virus–specific cellular immunity in individuals with active infection. J. Infect. Dis. 2015, 211, 600–612. [Google Scholar] [CrossRef] [PubMed]
- Schub, D.; Fousse, M.; Faßbender, K.; Gärtner, B.C.; Sester, U.; Sester, M.; Schmidt, T. CTLA-4-expression on VZV-specific T cells in CSF and blood is specifically increased in patients with VZV related central nervous system infections. Eur. J. Immunol. 2018, 48, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Koebel, C.M.; Vermi, W.; Swann, J.B.; Zerafa, N.; Rodig, S.J.; Old, L.J.; Smyth, M.J.; Schreiber, R.D. Adaptive immunity maintains occult cancer in an equilibrium state. Nature 2007, 450, 903–907. [Google Scholar] [CrossRef] [PubMed]
- Mattes, J.; Hulett, M.; Xie, W.; Hogan, S.; Rothenberg, M.E.; Foster, P.; Parish, C. Immunotherapy of cytotoxic T cell-resistant tumors by T helper 2 cells: An eotaxin and STAT6-dependent process. J. Exp. Med. 2003, 197, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Hung, K.; Hayashi, R.; Lafond-Walker, A.; Lowenstein, C.; Pardoll, D.; Levitsky, H. The central role of CD4+ T cells in the antitumor immune response. J. Exp. Med. 1998, 188, 2357–2368. [Google Scholar] [CrossRef] [PubMed]
- Scholler, J.; Brady, T.L.; Binder-Scholl, G.; Hwang, W.T.; Plesa, G.; Hege, K.M.; Vogel, A.N.; Kalos, M.; Riley, J.L.; Deeks, S.G.; et al. Decade-long safety and function of retroviral-modified chimeric antigen receptor T cells. Sci. Transl. Med. 2012, 4, 132ra153. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.C.; Shan, L.; Hosmane, N.N.; Wang, J.; Laskey, S.B.; Rosenbloom, D.I.S.; Lai, J.; Blankson, J.N.; Siliciano, J.D.; Siliciano, R.F. Replication-competent noninduced proviruses in the latent reservoir increase barrier to HIV-1 cure. Cell 2013, 155, 540–551. [Google Scholar] [CrossRef] [PubMed]
- Huetter, G.; Nowak, D.; Mossner, M.; Ganepola, S.; Muessig, A.; Allers, K.; Schneider, T.; Hofmann, J.; Kuecherer, C.; Blau, O.; et al. Long-Term Control of HIV by CCR5 Δ32/Δ32 Stem-Cell Transplantaion. N. Engl. J. Med. 2009, 360, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Ahmadzadeh, M.; Johnson, L.A.; Heemskerk, B.; Wunderlich, J.R.; Dudley, M.E.; White, D.E.; Rosenberg, S.A. Tumor antigen-specific CD8 T cells infiltrating the tumor express high levels of PD-1 and are functionally impaired. Blood 2009, 114, 1537–1544. [Google Scholar] [CrossRef] [PubMed]
- Baitsch, L.; Baumgaertner, P.; Devevre, E.; Raghav, S.K.; Legat, A.; Barba, L.; Wieckowski, S.; Bouzourene, H.; Deplancke, B.; Romero, P.; et al. Exhaustion of tumor-specific CD8+ T cells in metastases from melanoma patients. J. Clin. Investig. 2011, 121, 2350–2360. [Google Scholar] [CrossRef] [PubMed]
- Staveley-O’Carroll, K.; Sotomayor, E.; Montgomery, J.; Borrello, I.; Hwang, L.; Fein, S.; Pardoll, D.; Levitsky, H. Induction of antigen-specific T cell anergy: An early event in the course of tumor progression. Proc. Natl. Acad. Sci. USA 1998, 95, 1178–1183. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, S.A.; Yang, J.C.; Sherry, R.M.; Kammula, U.S.; Hughes, M.S.; Phan, G.Q.; Citrin, D.E.; Restifo, N.P.; Robbins, P.F.; Wunderlich, J.R.; et al. Durable complete responses in heavily pretreated patients with metastatic melanoma using T cell transfer immunotherapy. Clin. Cancer Res. 2011, 17, 4550–4557. [Google Scholar] [CrossRef] [PubMed]
- Dudley, M.E.; Wunderlich, J.R.; Robbins, P.F.; Yang, J.C.; Hwu, P.; Schwartzentruber, D.J.; Topalian, S.L.; Sherry, R.; Restifo, N.P.; Hubicki, A.M.; et al. Cancer regression and autoimmunity in patients after clonal repopulation with antitumor lymphocytes. Science 2002, 298, 850–854. [Google Scholar] [CrossRef] [PubMed]
- Postow, M.A.; Callahan, M.K.; Wolchok, J.D. Immune checkpoint blockade in cancer therapy. J. Clin. Oncol. 2015, 33, 1974–1982. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.C.; Levine, J.H.; Cogdill, A.P.; Zhao, Y.; Anang, N.-A.A.S.; Andrews, M.C.; Sharma, P.; Wang, J.; Wargo, J.A.; Pe’er, D.; et al. Distinct cellular mechanisms underlie anti-CTLA-4 and anti-PD-1 checkpoint blockade. Cell 2017, 170, 1120–1133.e17. [Google Scholar] [CrossRef] [PubMed]
- Barbee, M.S.; Ogunniyi, A.; Horvat, T.Z.; Dang, T.O. Current status and future directions of the immune checkpoint inhibitors ipilimumab, pembrolizumab, and nivolumab in oncology. Ann. Pharmacother. 2015, 49, 907–937. [Google Scholar] [CrossRef] [PubMed]
- Sangro, B.; Gomez-Martin, C.; de la Mata, M.; Inarrairaegui, M.; Garralda, E.; Barrera, P.; Riezu-Boj, J.I.; Larrea, E.; Alfaro, C.; Sarobe, P.; et al. A clinical trial of CTLA-4 blockade with tremelimumab in patients with hepatocellular carcinoma and chronic hepatitis C. J. Hepatol. 2013, 59, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Reck, M.; Rodríguez-Abreu, D.; Robinson, A.G.; Hui, R.; Csőszi, T.; Fülöp, A.; Gottfried, M.; Peled, N.; Tafreshi, A.; Cuffe, S.; et al. Pembrolizumab versus chemotherapy for PD-L1–positive non–small-cell lung cancer. N. Engl. J. Med. 2016, 375, 1823–1833. [Google Scholar] [CrossRef] [PubMed]
- Hersey, P.; Gowrishankar, K. Pembrolizumab joins the anti-PD-1 armamentarium in the treatment of melanoma. Future Oncol. 2015, 11, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Rooke, R. Can calcium signaling be harnessed for cancer immunotherapy? Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 2334–2340. [Google Scholar] [CrossRef] [PubMed]
- Ernst, B.; Lee, D.S.; Chang, J.M.; Sprent, J.; Surh, C.D. The peptide ligands mediating positive selection in the thymus control T cell survival and homeostatic proliferation in the periphery. Immunity 1999, 11, 173–181. [Google Scholar] [CrossRef]
- Smith, K.; Seddon, B.; Purbhoo, M.A.; Zamoyska, R.; Fisher, A.G.; Merkenschlager, M. Sensory adaptation in naive peripheral CD4 T cells. J. Exp. Med. 2001, 194, 1253–1261. [Google Scholar] [CrossRef] [PubMed]
- Dorothée, G.; Vergnon, I.; El Hage, F.; Chansac, B.L.M.; Ferrand, V.; Lécluse, Y.; Opolon, P.; Chouaib, S.; Bismuth, G.; Mami-Chouaib, F. In situ sensory adaptation of tumor-infiltrating T lymphocytes to peptide-MHC levels elicits strong antitumor reactivity. J. Immunol. 2005, 174, 6888–6897. [Google Scholar] [CrossRef] [PubMed]
- Friedlein, G.; El Hage, F.; Vergnon, I.; Richon, C.; Saulnier, P.; Lecluse, Y.; Caignard, A.; Boumsell, L.; Bismuth, G.; Chouaib, S.; et al. Human CD5 protects circulating tumor antigen-specific CTL from tumor-mediated activation-induced cell death. J. Immunol. 2007, 178, 6821–6827. [Google Scholar] [CrossRef] [PubMed]
- Axtell, R.C.; Webb, M.S.; Barnum, S.R.; Raman, C. Cutting edge: Critical role for CD5 in experimental autoimmune encephalomyelitis: Inhibition of engagement reverses disease in mice. J. Immunol. 2004, 173, 2928–2932. [Google Scholar] [CrossRef] [PubMed]
- Simoes, I.T.; Aranda, F.; Carreras, E.; Andres, M.V.; Casado-Llombart, S.; Martinez, V.G.; Lozano, F. Immunomodulatory effects of soluble CD5 on experimental tumor models. Oncotarget 2017, 8, 108156–108169. [Google Scholar] [CrossRef] [PubMed]
- Tabbekh, M.; Franciszkiewicz, K.; Haouas, H.; Lecluse, Y.; Benihoud, K.; Raman, C.; Mami-Chouaib, F. Rescue of tumor-infiltrating lymphocytes from activation-induced cell death enhances the antitumor CTL response in CD5-deficient mice. J. Immunol. 2011, 187, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Mazmanian, S.K. The gut microbiome shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Vétizou, M.; Pitt, J.M.; Daillère, R.; Lepage, P.; Waldschmitt, N.; Flament, C.; Rusakiewicz, S.; Routy, B.; Roberti, M.P.; Duong, C.P.M.; et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science 2015, 350, 1079–1084. [Google Scholar] [CrossRef] [PubMed]
- Botticelli, A.; Zizzari, I.; Mazzuca, F.; Ascierto, P.A.; Putignani, L.; Marchetti, L.; Napoletano, C.; Nuti, M.; Marchetti, P. Cross-talk between microbiota and immune fitness to steer and control response to anti PD-1/PDL-1 treatment. Oncotarget 2017, 8, 8890–8899. [Google Scholar] [CrossRef] [PubMed]
- Kosiewicz, M.M.; Dryden, G.W.; Chhabra, A.; Alard, P. Relationship between gut microbiota and development of T cell associated disease. FEBS Lett. 2014, 588, 4195–4206. [Google Scholar] [CrossRef] [PubMed]
- Lathrop, S.K.; Bloom, S.M.; Rao, S.M.; Nutsch, K.; Lio, C.-W.; Santacruz, N.; Peterson, D.A.; Stappenbeck, T.S.; Hsieh, C.-S. Peripheral education of the immune system by colonic commensal microbiota. Nature 2011, 478, 250. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Perez, G.; Lamousé-Smith, E.S.N. Gastrointestinal microbiome dysbiosis in infant mice alters peripheral CD8+ T cell receptor signaling. Front. Immunol. 2017, 8, 265. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Wei, B.; Velazquez, P.; Borneman, J.; Braun, J. Commensal microbiota alter the abundance and TCR responsiveness of splenic naïve CD4+ T lymphocytes. Clin. Immunol. 2005, 117, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Bazett, M.; Bergeron, M.-E.; Haston, C.K. Streptomycin treatment alters the intestinal microbiome, pulmonary T cell profile and airway hyperresponsiveness in a cystic fibrosis mouse model. Sci. Rep. 2016, 6, 19189. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Repáraz, J.; Mielcarz, D.W.; Haque-Begum, S.; Kasper, L.H. Induction of a regulatory B cell population in experimental allergic encephalomyelitis by alteration of the gut commensal microflora. Gut Microbes 2010, 1, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Allison, J.P.; Krummel, M.F. The Yin and Yang of T cell costimulation. Science 1995, 270, 932–933. [Google Scholar] [CrossRef] [PubMed]
- Allison, J.P. Checkpoints. Cell 2015, 162, 1202–1205. [Google Scholar] [CrossRef] [PubMed]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.-L.; et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti–PD-L1 efficacy. Science 2015, 350, 1084–1089. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of CD5 on different stages of T cell development. CD5 expression on thymocytes is directly proportional to the signaling intensity of the TCR:self-pMHC interaction. In the periphery, T cells with higher CD5 levels (CD5hi) are better responders to foreign-peptide. Long-lived memory cells populations are enriched for CD5hi T cells [34,102,119].
Figure 1.
Effects of CD5 on different stages of T cell development. CD5 expression on thymocytes is directly proportional to the signaling intensity of the TCR:self-pMHC interaction. In the periphery, T cells with higher CD5 levels (CD5hi) are better responders to foreign-peptide. Long-lived memory cells populations are enriched for CD5hi T cells [34,102,119].

Figure 2.
Inhibiting co-receptors modulate T cell activation by increasing (green arrows) or decreasing activity (red arrows). CD5 is present in naïve T cells and localizes to the TCR:pMHC complex during activation. Initial activation cascades signal for the release of CTLA-4 from vesicles to the cell surface while the transcription factor NFAT transcribes PD-1. CTLA-4 provides inhibitory signals during early activation while PD-1 is expressed later and inhibits later stages of T cell activation. The initial Ca2+ mobilization is decreased by CTLA-4 and PD-1 downstream signals. A more detailed illustration of the calcium signaling pathway (i.e., IP3, STIM 1/2, CRAC channel, calmodulin, etc.) is outlined in Figure 3.
Figure 2.
Inhibiting co-receptors modulate T cell activation by increasing (green arrows) or decreasing activity (red arrows). CD5 is present in naïve T cells and localizes to the TCR:pMHC complex during activation. Initial activation cascades signal for the release of CTLA-4 from vesicles to the cell surface while the transcription factor NFAT transcribes PD-1. CTLA-4 provides inhibitory signals during early activation while PD-1 is expressed later and inhibits later stages of T cell activation. The initial Ca2+ mobilization is decreased by CTLA-4 and PD-1 downstream signals. A more detailed illustration of the calcium signaling pathway (i.e., IP3, STIM 1/2, CRAC channel, calmodulin, etc.) is outlined in Figure 3.

Figure 3.
CD5 expression levels in naïve T cells may influence T cell metabolism and function. Differential levels of CD5 result in differences in Ca2+ mobilization in naïve T cells. CD5hi naïve T cells have higher Ca2+ influx than CD5lo naïve T cells upon TCR:pMHC interaction [35]. Ca2+ signaling plays a significant role in T cell activation and influences metabolism and T cell function. Differential Ca2+ mobilization and expression of calcineurin and NFAT affect glycolysis and mitochondrial respiration (hypothetical levels of metabolic activation are shown with dashed (low) or solid (high) arrows), suggesting CD5 expression may affect metabolic reprograming during T cell activation [141].
Figure 3.
CD5 expression levels in naïve T cells may influence T cell metabolism and function. Differential levels of CD5 result in differences in Ca2+ mobilization in naïve T cells. CD5hi naïve T cells have higher Ca2+ influx than CD5lo naïve T cells upon TCR:pMHC interaction [35]. Ca2+ signaling plays a significant role in T cell activation and influences metabolism and T cell function. Differential Ca2+ mobilization and expression of calcineurin and NFAT affect glycolysis and mitochondrial respiration (hypothetical levels of metabolic activation are shown with dashed (low) or solid (high) arrows), suggesting CD5 expression may affect metabolic reprograming during T cell activation [141].

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Freitas, C.M.T.; Johnson, D.K.; Weber, K.S. T Cell Calcium Signaling Regulation by the Co-Receptor CD5. Int. J. Mol. Sci. 2018, 19, 1295. https://doi.org/10.3390/ijms19051295
AMA Style
Freitas CMT, Johnson DK, Weber KS. T Cell Calcium Signaling Regulation by the Co-Receptor CD5. International Journal of Molecular Sciences. 2018; 19(5):1295. https://doi.org/10.3390/ijms19051295
Chicago/Turabian StyleFreitas, Claudia M. Tellez, Deborah K. Johnson, and K. Scott Weber. 2018. "T Cell Calcium Signaling Regulation by the Co-Receptor CD5" International Journal of Molecular Sciences 19, no. 5: 1295. https://doi.org/10.3390/ijms19051295
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.