Inhibitory Effect of Purpurogallin on Osteoclast Differentiation in Vitro through the Downregulation of c-Fos and NFATc1



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
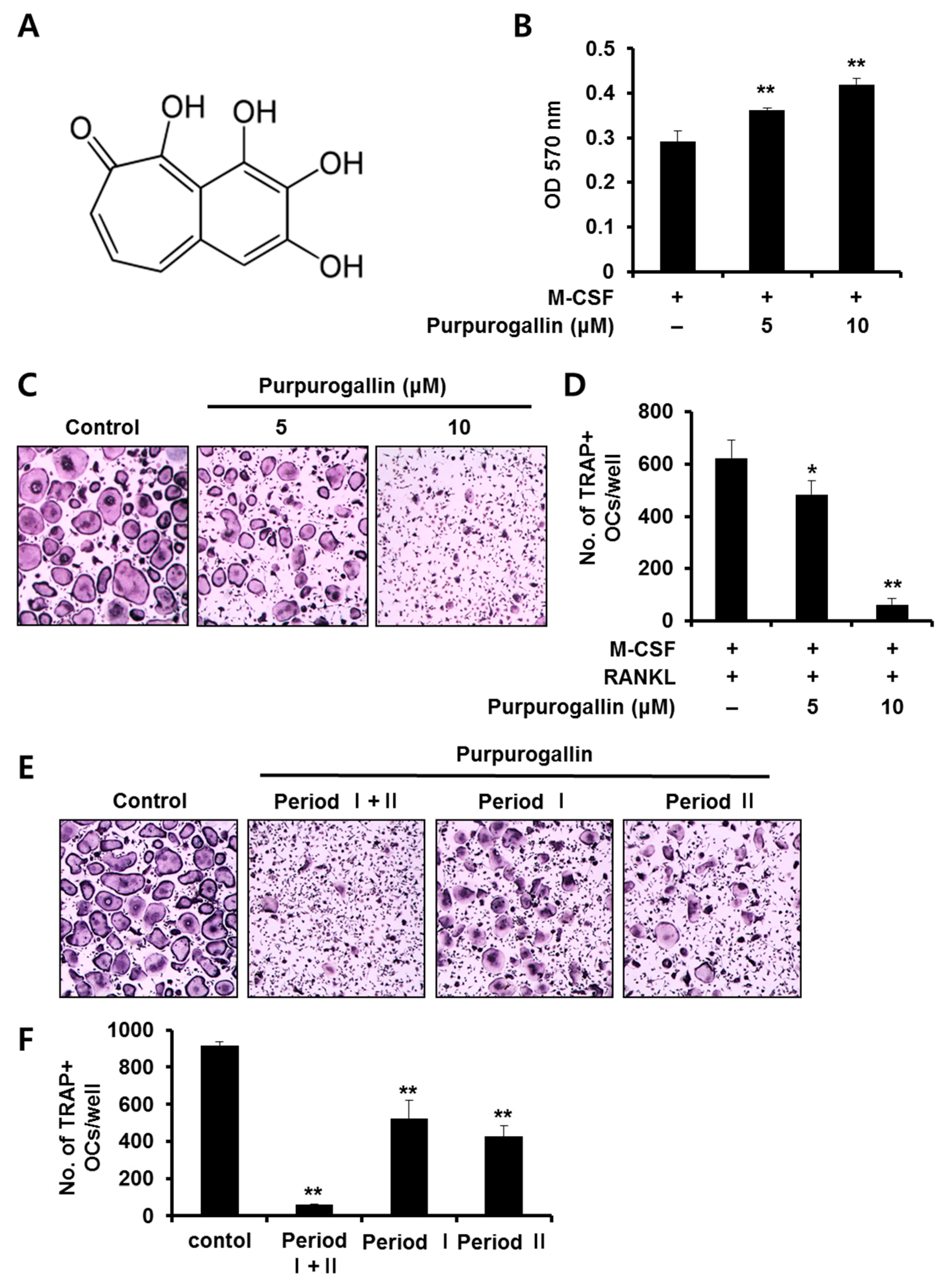
2.1. Purpurogallin Suppresses Receptor Activator of Nuclear Factor Kappa-B Ligand (RANKL)-Mediated Osteoclast Differentiation of Bone Marrow Macrophages (BMMs)
2.2. Purpurogallin Inhibits both Early and Late Stages of Osteoclast Differentiation
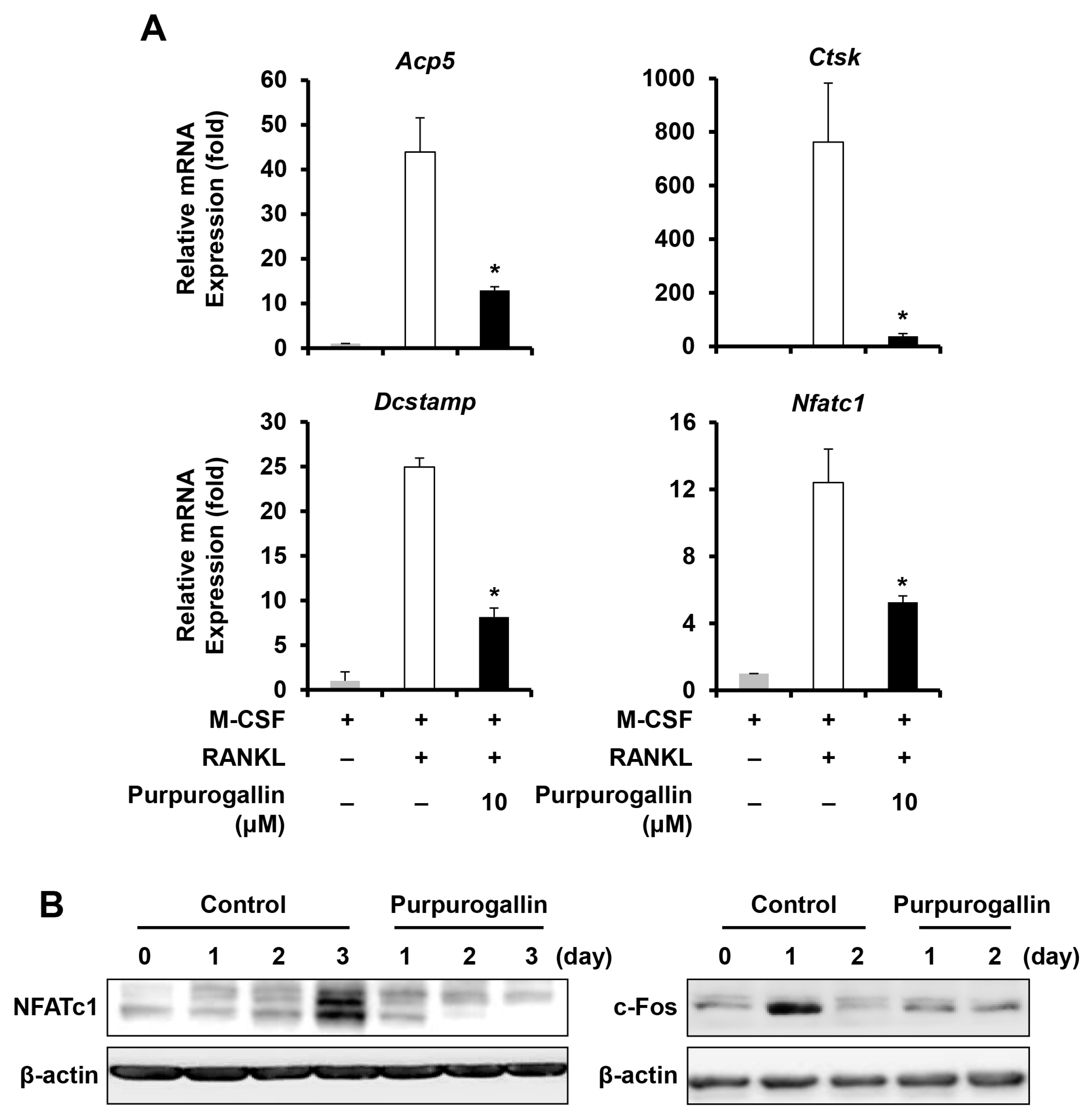
2.3. Purpurogallin Downregulates the Expression of Key Regulatory Factors and Osteoclast Marker Genes
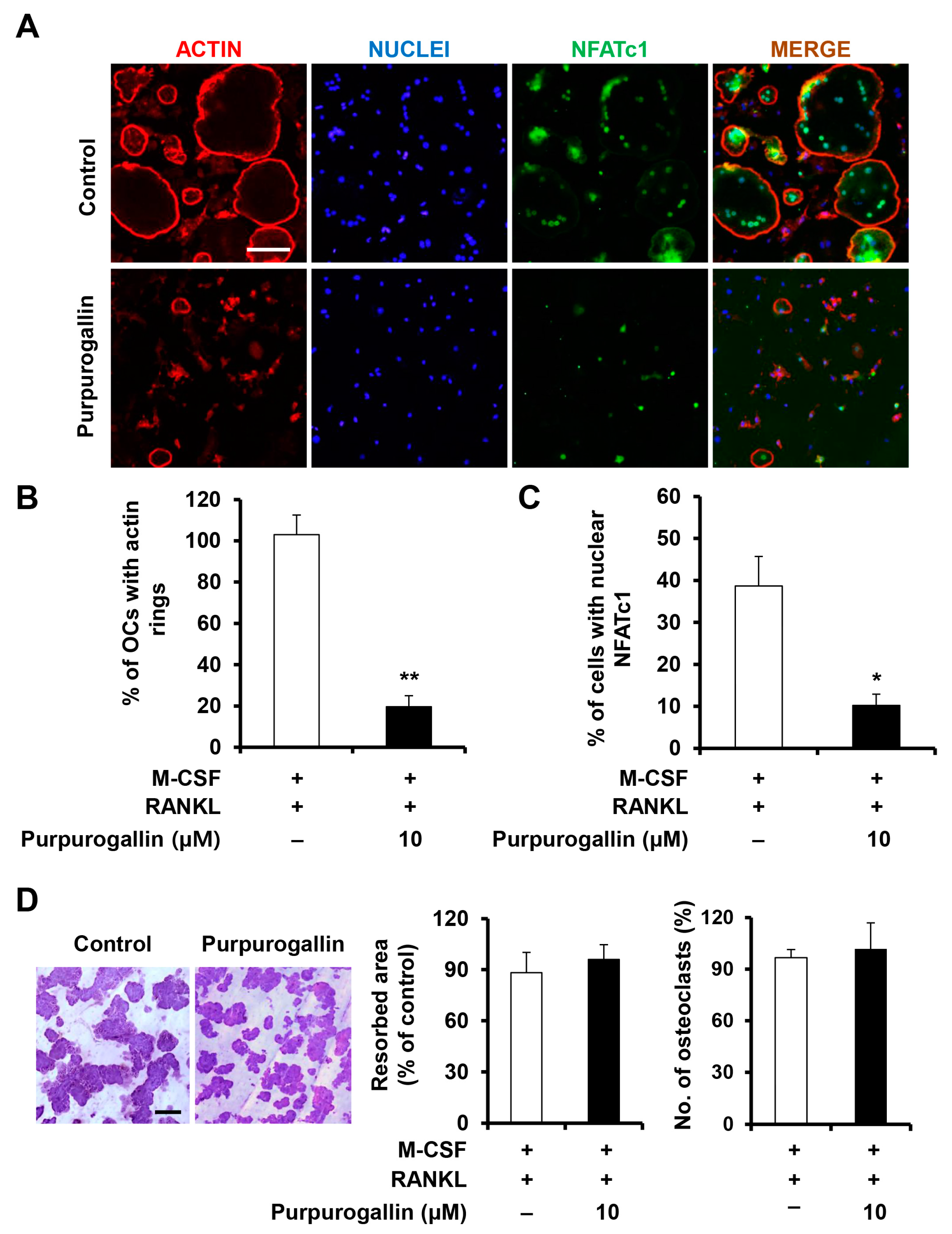
2.4. Purpurogallin Inhibits the Formation of Actin Rings
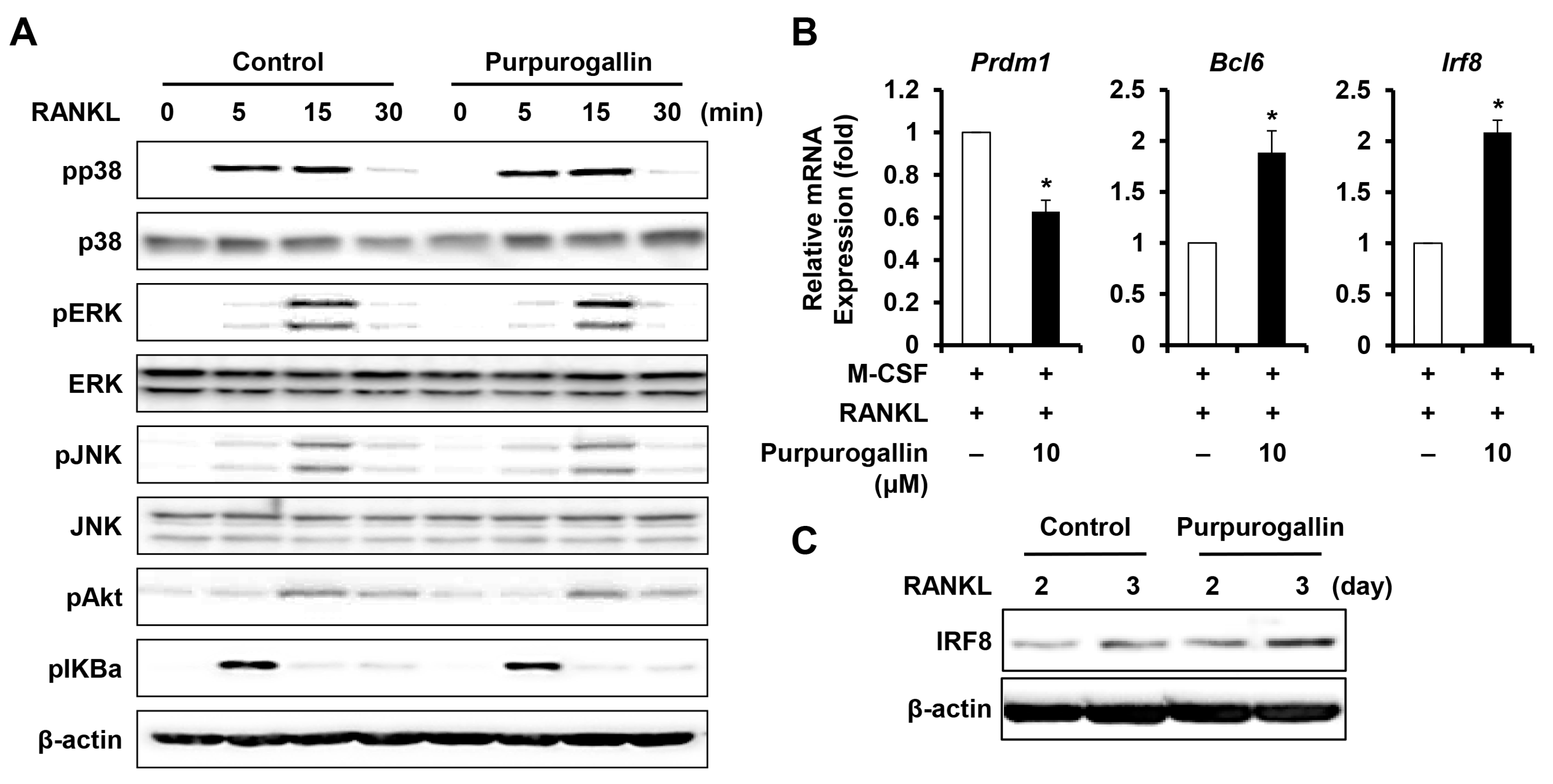
2.5. Purpurogallin Induces the Expression of Negative Regulating Factors of RANKL-Induced Osteoclast Differentiation
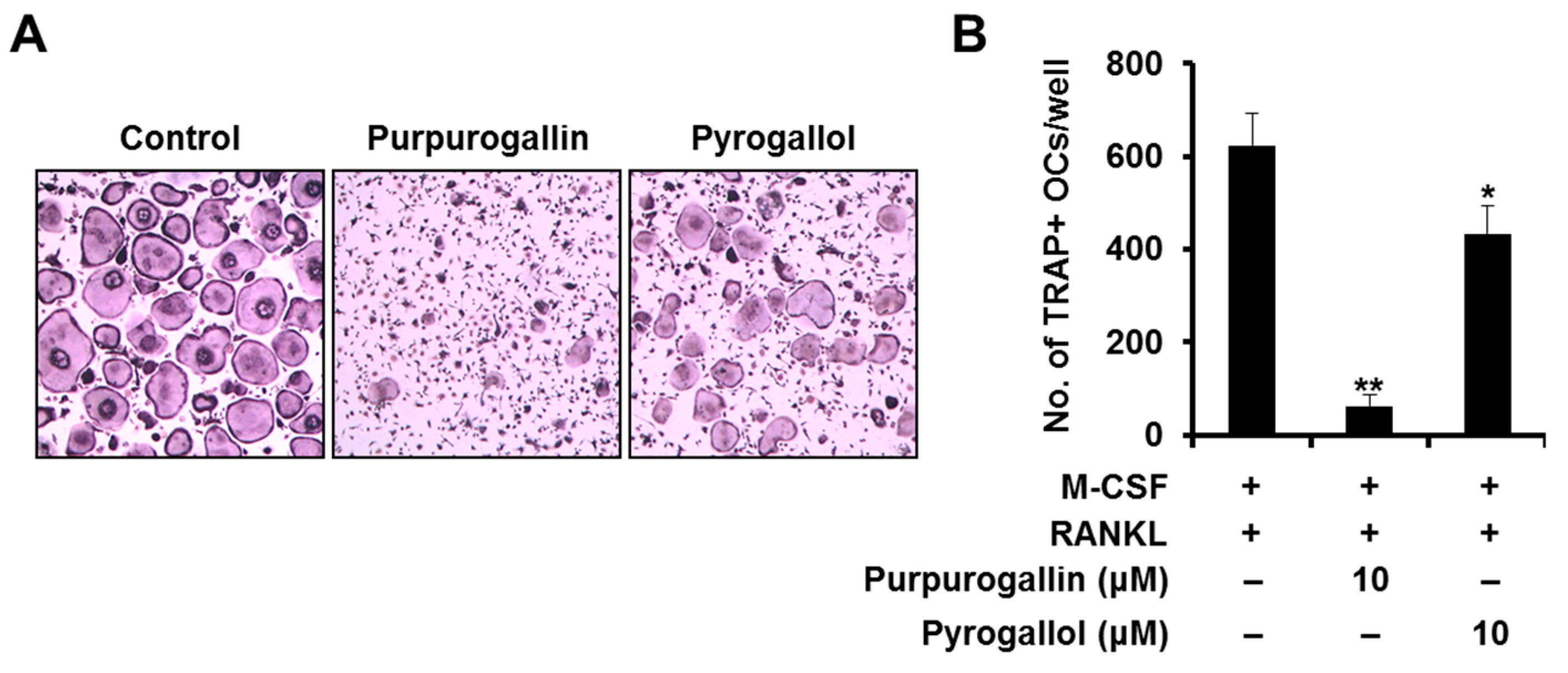
2.6. Purpurogallin Strongly Inhibits Osteoclast Differentiation more than Pyrogallol
3. Discussion
4. Materials and Methods
4.1. Osteoclast Differentiation and TRAP Staining
4.2. Cell Viability Assay
4.3. Real-Time PCR
4.4. Western Blot Analysis
4.5. Resorption Pit Assay
4.6. Immunofluorescence
4.7. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Phan, T.C.; Xu, J.; Zheng, M.H. Interaction between osteoblast and osteoclast: Impact in bone disease. Histol. Histopathol. 2004, 19, 1325–1344. [Google Scholar] [PubMed]
- Roodman, G.D. Pathogenesis of myeloma bone disease. Leukemia 2009, 23, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Kwak, H.B.; Lee, S.W.; Li, Y.J.; Kim, Y.A.; Han, S.Y.; Jhon, G.J.; Kim, H.H.; Lee, Z.H. Inhibition of osteoclast differentiation and bone resorption by a novel lysophosphatidylcholine derivative, scoh. Biochem. Pharmacol. 2004, 67, 1239–1248. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Wachi, M.; Woo, J.T.; Kato, M.; Kasai, S.; Takahashi, F.; Lee, I.S.; Nagai, K. Fenton reaction is primarily involved in a mechanism of (−)-epigallocatechin-3-gallate to induce osteoclastic cell death. Biochem. Biophys. Res. Commun. 2002, 292, 94–101. [Google Scholar] [CrossRef] [PubMed]
- McClung, M.; Harris, S.T.; Miller, P.D.; Bauer, D.C.; Davison, K.S.; Dian, L.; Hanley, D.A.; Kendler, D.L.; Yuen, C.K.; Lewiecki, E.M. Bisphosphonate therapy for osteoporosis: Benefits, risks, and drug holiday. Am. J. Med. 2013, 126, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.G.; Watts, N.B.; Ebetino, F.H.; Rogers, M.J. Mechanisms of action of bisphosphonates: Similarities and differences and their potential influence on clinical efficacy. Osteoporos. Int. 2008, 19, 733–759. [Google Scholar] [CrossRef] [PubMed]
- Khosla, S.; Burr, D.; Cauley, J.; Dempster, D.W.; Ebeling, P.R.; Felsenberg, D.; Gagel, R.F.; Gilsanz, V.; Guise, T.; Koka, S.; et al. Bisphosphonate-associated osteonecrosis of the jaw: Report of a task force of the american society for bone and mineral research. J. Bone Miner. Res. 2007, 22, 1479–1491. [Google Scholar] [CrossRef] [PubMed]
- Papapetrou, P.D. Bisphosphonate-associated adverse events. Hormones 2009, 8, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Solomon, D.H.; Rekedal, L.; Cadarette, S.M. Osteoporosis treatments and adverse events. Curr. Opin. Rheumatol. 2009, 21, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Abou-Karam, M.; Shier, W.T. Inhibition of oncogene product enzyme activity as an approach to cancer chemoprevention. Tyrosine-specific protein kinase inhibition by purpurogallin from quercus sp. Nutgall. Phytother. Res. 1999, 13, 337–340. [Google Scholar] [CrossRef]
- Wu, T.W.; Zeng, L.H.; Wu, J.; Fung, K.P.; Weisel, R.D.; Hempel, A.; Camerman, N. Molecular structure and antioxidant specificity of purpurogallin in three types of human cardiovascular cells. Biochem. Pharmacol. 1996, 52, 1073–1080. [Google Scholar] [CrossRef]
- Das, M.; Bickers, D.R.; Mukhtar, H. Plant phenols as in vitro inhibitors of glutathione s-transferase(s). Biochem. Biophys. Res. Commun. 1984, 120, 427–433. [Google Scholar] [CrossRef]
- Honda, S.; Fukuyama, Y.; Nishiwaki, H.; Masuda, A.; Masuda, T. Conversion to purpurogallin, a key step in the mechanism of the potent xanthine oxidase inhibitory activity of pyrogallol. Free Radic. Biol. Med. 2017, 106, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Veser, J. Kinetics and inhibition studies of catechol o-methyltransferase from the yeast candida tropicalis. J. Bacteriol. 1987, 169, 3696–3700. [Google Scholar] [CrossRef] [PubMed]
- Prasad, K.; Laxdal, V.A. Evaluation of hydroxyl radical-scavenging property of purpurogallin using high pressure liquid chromatography. Mol. Cell. Biochem. 1994, 135, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.W.; Wu, J.; Carey, D.; Zeng, L.H. Purpurogallin protects both ventricular myocytes and aortic endothelial cells of rats against oxyradical damage. Biochem. Cell Biol. 1992, 70, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.W.; Zeng, L.H.; Wu, J.; Carey, D. Purpurogallin—A natural and effective hepatoprotector in vitro and in vivo. Biochem. Cell Biol. 1991, 69, 747–750. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, H.; Fung, K.P.; Wu, T.W. Purpurogallin as an antioxidant protector of human erythrocytes against lysis by peroxyl radicals. Life Sci. 1993, 53, PL39–PL43. [Google Scholar] [CrossRef]
- Park, H.Y.; Kim, T.H.; Kim, C.G.; Kim, G.Y.; Kim, C.M.; Kim, N.D.; Kim, B.W.; Hwang, H.J.; Choi, Y.H. Purpurogallin exerts antiinflammatory effects in lipopolysaccharidestimulated bv2 microglial cells through the inactivation of the nfkappab and mapk signaling pathways. Int. J. Mol. Med. 2013, 32, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, F.; Matsubara, T.; Tsurukai, T.; Hata, K.; Nishimura, R.; Yoneda, T. Jnk/c-jun signaling mediates an anti-apoptotic effect of rankl in osteoclasts. J. Bone Miner. Res. 2008, 23, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.J.; Jeker, H.; Schaueblin, M. Wortmannin, a potent inhibitor of phosphatidylinositol 3-kinase, inhibits osteoclastic bone resorption in vitro. Calcif. Tissue Int. 1995, 56, 336–338. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Woo, K.M.; Kim, S.Y.; Kim, H.M.; Kwack, K.; Lee, Z.H.; Kim, H.H. The phosphatidylinositol 3-kinase, p38, and extracellular signal-regulated kinase pathways are involved in osteoclast differentiation. Bone 2002, 30, 71–77. [Google Scholar] [CrossRef]
- Asagiri, M.; Takayanagi, H. The molecular understanding of osteoclast differentiation. Bone 2007, 40, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Saltel, F.; Destaing, O.; Bard, F.; Eichert, D.; Jurdic, P. Apatite-mediated actin dynamics in resorbing osteoclasts. Mol. Biol. Cell 2004, 15, 5231–5241. [Google Scholar] [CrossRef] [PubMed]
- Miyauchi, Y.; Ninomiya, K.; Miyamoto, H.; Sakamoto, A.; Iwasaki, R.; Hoshi, H.; Miyamoto, K.; Hao, W.; Yoshida, S.; Morioka, H.; et al. The blimp1-bcl6 axis is critical to regulate osteoclast differentiation and bone homeostasis. J. Exp. Med. 2010, 207, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Tauber, H. Oxidation of pyrogallol to purpurogallin by crystallin catalase. J. Biol. Chem. 1953, 205, 395–400. [Google Scholar] [PubMed]
- Clarke, B. Normal bone anatomy and physiology. Clin. J. Am. Soc. Nephrol. 2008, 3 (Suppl. S3), S131–S139. [Google Scholar] [CrossRef] [PubMed]
- Dickson, K.M.; Bhakar, A.L.; Barker, P.A. Traf6-dependent nf-kb transcriptional activity during mouse development. Dev. Dyn. 2004, 231, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Lee, Z.H.; Kim, H.H. Signal transduction by receptor activator of nuclear factor kappa b in osteoclasts. Biochem. Biophys. Res. Commun. 2003, 305, 211–214. [Google Scholar] [CrossRef]
- Wada, T.; Nakashima, T.; Hiroshi, N.; Penninger, J.M. Rankl-rank signaling in osteoclastogenesis and bone disease. Trends Mol. Med. 2006, 12, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Asagiri, M.; Sato, K.; Usami, T.; Ochi, S.; Nishina, H.; Yoshida, H.; Morita, I.; Wagner, E.F.; Mak, T.W.; Serfling, E.; et al. Autoamplification of nfatc1 expression determines its essential role in bone homeostasis. J. Exp. Med. 2005, 202, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, K.; Nakashima, T.; Hayashi, M.; Fukunaga, T.; Kato, S.; Kodama, T.; Takahashi, S.; Calame, K.; Takayanagi, H. Blimp1-mediated repression of negative regulators is required for osteoclast differentiation. Proc. Natl. Acad. Sci. USA 2010, 107, 3117–3122. [Google Scholar] [CrossRef] [PubMed]
- Park-Min, K.H.; Lee, E.Y.; Moskowitz, N.K.; Lim, E.; Lee, S.K.; Lorenzo, J.A.; Huang, C.; Melnick, A.M.; Purdue, P.E.; Goldring, S.R.; et al. Negative regulation of osteoclast precursor differentiation by cd11b and beta2 integrin-b-cell lymphoma 6 signaling. J. Bone Miner. Res. 2013, 28, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Ihn, H.J.; Kim, J.A.; Cho, H.S.; Shin, H.I.; Kim, G.Y.; Choi, Y.H.; Jeon, Y.J.; Park, E.K. Diphlorethohydroxycarmalol from ishige okamurae suppresses osteoclast differentiation by downregulating the nf-kappab signaling pathway. Int. J. Mol. Sci. 2017, 18, 2635. [Google Scholar] [CrossRef] [PubMed]
- Ihn, H.J.; Lee, T.; Kim, J.A.; Lee, D.; Kim, N.D.; Shin, H.I.; Bae, Y.C.; Park, E.K. Ocli-023, a novel pyrimidine compound, suppresses osteoclastogenesis in vitro and alveolar bone resorption in vivo. PLoS ONE 2017, 12, e0170159. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, K.; Kim, T.H.; Ihn, H.J.; Kim, J.E.; Choi, J.-Y.; Shin, H.-I.; Park, E.K. Inhibitory Effect of Purpurogallin on Osteoclast Differentiation in Vitro through the Downregulation of c-Fos and NFATc1. Int. J. Mol. Sci. 2018, 19, 601. https://doi.org/10.3390/ijms19020601
Kim K, Kim TH, Ihn HJ, Kim JE, Choi J-Y, Shin H-I, Park EK. Inhibitory Effect of Purpurogallin on Osteoclast Differentiation in Vitro through the Downregulation of c-Fos and NFATc1. International Journal of Molecular Sciences. 2018; 19(2):601. https://doi.org/10.3390/ijms19020601
Chicago/Turabian StyleKim, Kiryeong, Tae Hoon Kim, Hye Jung Ihn, Jung Eun Kim, Je-Yong Choi, Hong-In Shin, and Eui Kyun Park. 2018. "Inhibitory Effect of Purpurogallin on Osteoclast Differentiation in Vitro through the Downregulation of c-Fos and NFATc1" International Journal of Molecular Sciences 19, no. 2: 601. https://doi.org/10.3390/ijms19020601