Mangiferin Accelerates Glycolysis and Enhances Mitochondrial Bioenergetics

Abstract
:
1. Introduction
2. Results
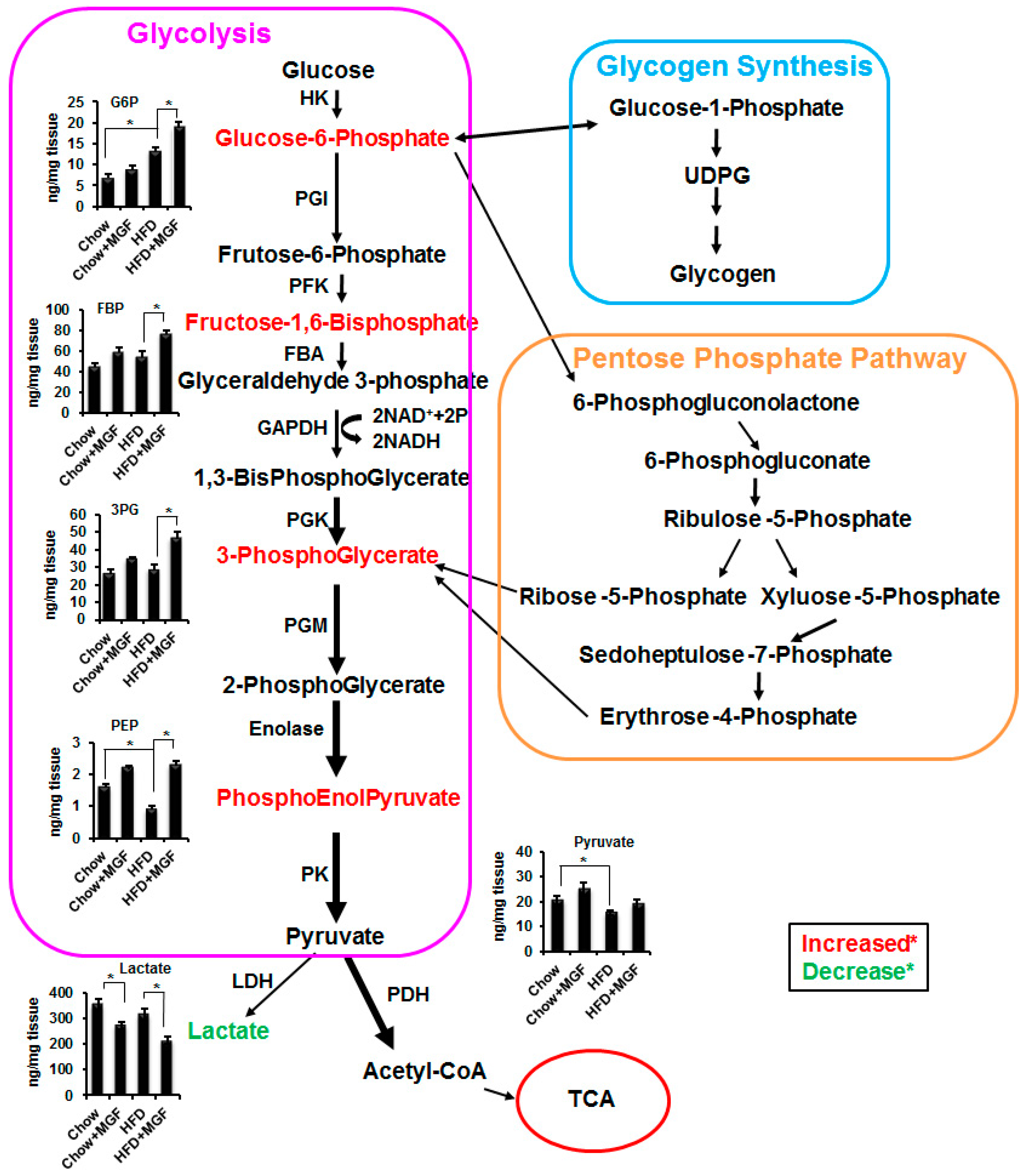
2.1. Profiling of Metabolites in Glucose Metabolism
2.2. MGF Accelerates Glycolysis
2.2.1. MGF Increases Glucose Metabolism via Glycolysis
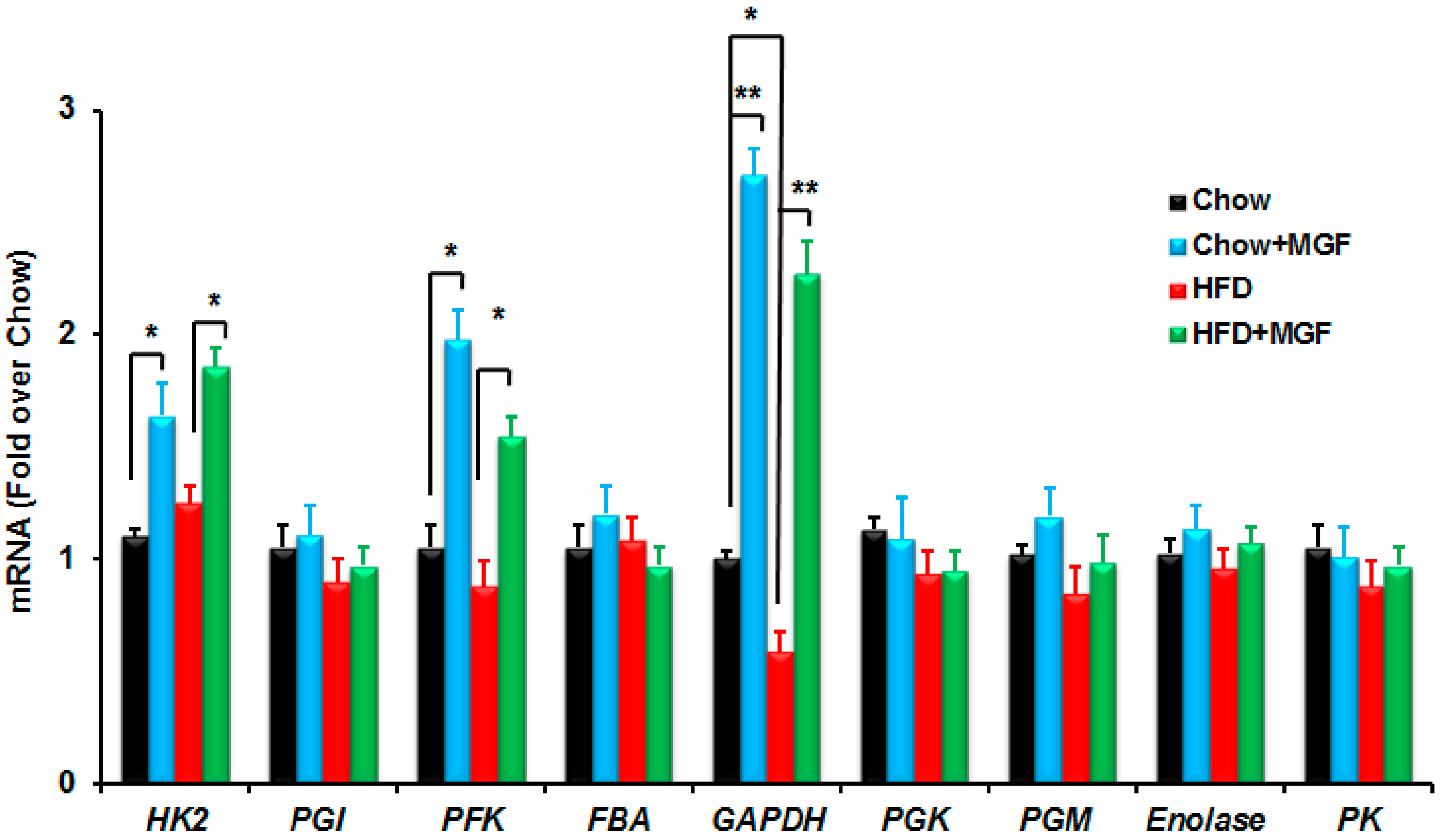
2.2.2. MGF Induces Some Enzymes in Glycolysis
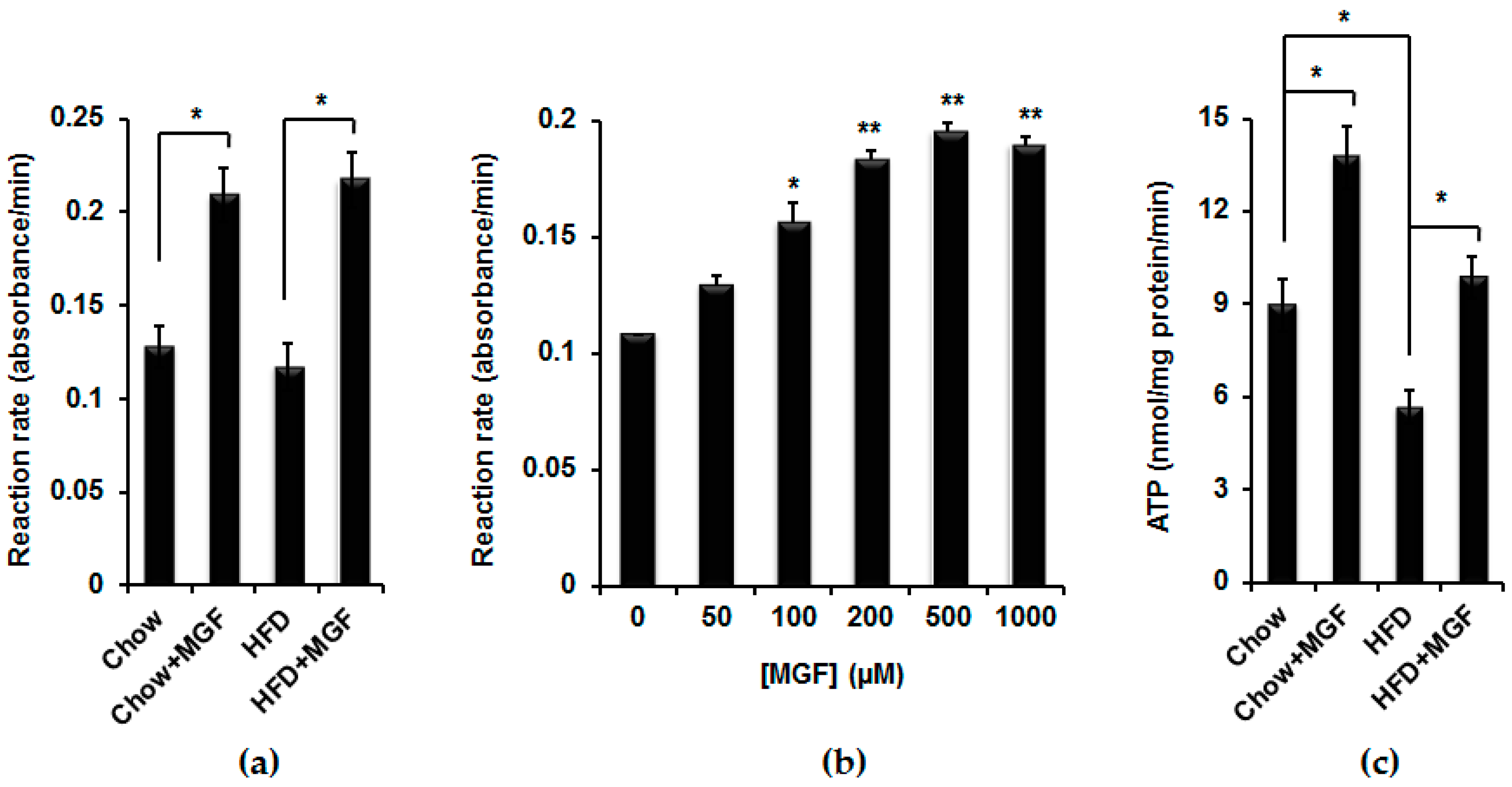
2.3. MGF Enhances Mitochondrial Bioenergetics
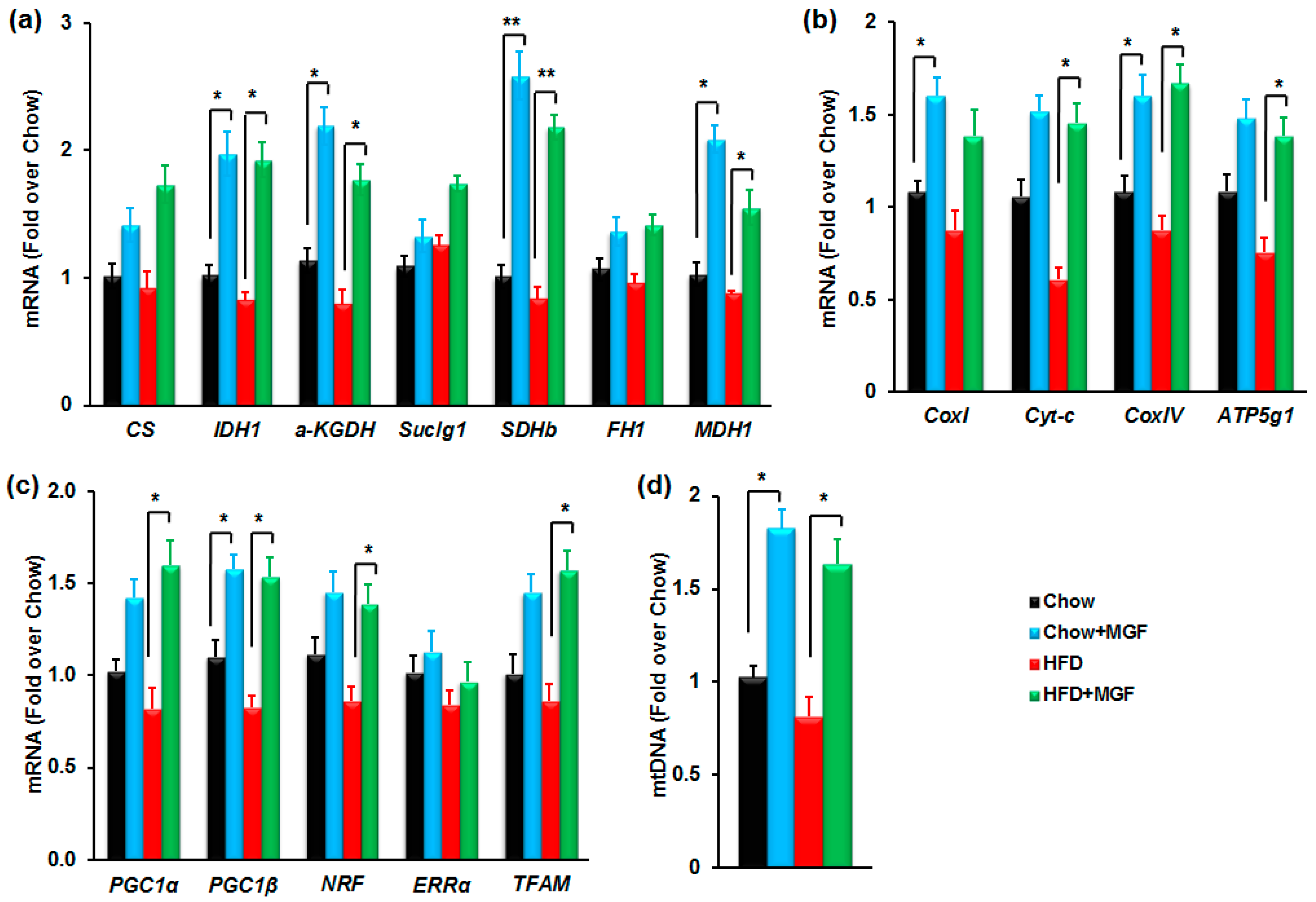
2.3.1. MGF Enhances Metabolism in the TCA Cycle
2.3.2. MGF Enhances SDH Activity and Overall Energy Production in Mitochondria
2.3.3. MGF Induces Mitochondrial Biogenesis
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Animal Experiments
4.3. Metabolite Profiling
4.4. mRNA Measurement by Real Time qRT-PCR
4.5. Mitochondria Isolation
4.6. Measurement of Mitochondrial Complex II Activity
4.7. Measurement of ATP Production
4.8. Mitochondrial DNA (mtDNA)
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| TCA | Tricarboxylic acid |
| ETC | Electron transport chain |
| PDH | Pyruvate dehydrogenase |
| CS | Citrate synthase |
| α-KG | α-Ketoglutarate |
| α-KGDH | α-Ketoglutarate dehydrogenase |
| PFK | Phosphofructose kinase |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| HFD | High fat diet |
| MGF | Mangiferin |
| UDPG | Urindine diphosphate glucose |
| PEP | Phosphoenolpyruvate |
| HK | Hexose kinase |
| PGI | Phosphoglucose isomerase |
| PFK | Phosphofructokinase |
| FBA | Fructose-1,6-bisphosphate aldolase |
| PGK | Phosphoglycerate Kinase |
| PGM | Phosphoglycerate mutase |
| PK | Pyruvate kinase |
| LDH | Lactate dehydrogenase |
| PC | Pyruvate carboxylase |
| CS | Citrate synthase |
| AC | Aconitase |
| IDH | Isocitrate dehydrogenase |
| Sucl | Succinyl coenzyme A synthetase |
| SDH | Succinate dehydrogenase |
| FH | Fumarase |
| MDH | Malate dehydrogenase |
| Atp5g1 | ATP synthase subunit 5g1 |
| TFAM | Transcriptional factor A |
| NRF1 | Nuclear respiratory factor 1 |
| ERRα | Estrogen-related receptor α |
| PGC-1α | Peroxisome proliferator-activated receptor-γ coactivator-1α |
| PGC-1β | Peroxisome proliferator-activated receptor-γ coactivator-1β |
| mtDNA | Mitochondrial DNA |
| PBS | Phosphate-buffered saline |
References
- Owen, O.E.; Kalhan, S.C.; Hanson, R.W. The key role of anaplerosis and cataplerosis for citric acid cycle function. J. Biol. Chem. 2002, 277, 30409–30412. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, G.M.; de Souza, D.P.; Burch, M.L.; Hamley, S.; Kloehn, J.; Selathurai, A.; Tull, D.; O’Callaghan, S.; McConville, M.J.; Bruce, C.R. Application of dynamic metabolomics to examine in vivo skeletal muscle glucose metabolism in the chronically high-fat fed mouse. Biochem. Biophys. Res. Commun. 2015, 462, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Chinopoulos, C. Which way does the citric acid cycle turn during hypoxia? The critical role of α-ketoglutarate dehydrogenase complex. J. Neurosci. Res. 2013, 91, 1030–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul-Ghani, M.A.; DeFronzo, R.A. Mitochondrial dysfunction, insulin resistance, and type 2 diabetes mellitus. Curr. Diabetes Rep. 2008, 8, 173–178. [Google Scholar] [CrossRef]
- Ristow, M.; Carlqvist, H.; Hebinck, J.; Vorgerd, M.; Krone, W.; Pfeiffer, A.; Muller-Wieland, D.; Ostenson, C.G. Deficiency of phosphofructo-1-kinase/muscle subtype in humans is associated with impairment of insulin secretory oscillations. Diabetes 1999, 48, 1557–1561. [Google Scholar] [CrossRef] [PubMed]
- Ristow, M.; Vorgerd, M.; Mohlig, M.; Schatz, H.; Pfeiffer, A. Deficiency of phosphofructo-1-kinase/muscle subtype in humans impairs insulin secretion and causes insulin resistance. J. Clin. Investig. 1997, 100, 2833–2841. [Google Scholar] [CrossRef] [PubMed]
- Ristow, M.; Vorgerd, M.; Mohlig, M.; Schatz, H.; Pfeiffer, A. Insulin resistance and impaired insulin secretion due to phosphofructo-1-kinase-deficiency in humans. J. Mol. Med. 1999, 77, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, D.; Zancan, P.; Coelho, W.S.; Gomez, L.S.; Sola-Penna, M. Metformin reverses hexokinase and 6-phosphofructo-1-kinase inhibition in skeletal muscle, liver and adipose tissues from streptozotocin-induced diabetic mouse. Arch. Biochem. Biophys. 2010, 496, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Matsumura, T.; Edelstein, D.; Rossetti, L.; Zsengeller, Z.; Szabo, C.; Brownlee, M. Inhibition of GAPDH activity by poly(ADP-ribose) polymerase activates three major pathways of hyperglycemic damage in endothelial cells. J. Clin. Investig. 2003, 112, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, M. The pathobiology of diabetic complications: A unifying mechanism. Diabetes 2005, 54, 1615–1625. [Google Scholar] [CrossRef] [PubMed]
- Wentzel, P.; Ejdesjo, A.; Eriksson, U.J. Maternal diabetes in vivo and high glucose in vitro diminish GAPDH activity in rat embryos. Diabetes 2003, 52, 1222–1228. [Google Scholar] [CrossRef] [PubMed]
- Senhaji, N.; Elkhalfi, B.; Soukri, A. Contribution to the study of glyceraldehyde-3-phosphate dehydrogenase in patients with type 2 diabetes. Pathol.-Biol. 2015, 63, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Befroy, D.E.; Petersen, K.F.; Dufour, S.; Mason, G.F.; de Graaf, R.A.; Rothman, D.L.; Shulman, G.I. Impaired mitochondrial substrate oxidation in muscle of insulin-resistant offspring of type 2 diabetic patients. Diabetes 2007, 56, 1376–1381. [Google Scholar] [CrossRef] [PubMed]
- Petersen, K.F.; Dufour, S.; Befroy, D.; Garcia, R.; Shulman, G.I. Impaired mitochondrial activity in the insulin-resistant offspring of patients with type 2 diabetes. N. Engl. J. Med. 2004, 350, 664–671. [Google Scholar] [CrossRef] [PubMed]
- Szendroedi, J.; Schmid, A.I.; Chmelik, M.; Toth, C.; Brehm, A.; Krssak, M.; Nowotny, P.; Wolzt, M.; Waldhausl, W.; Roden, M. Muscle mitochondrial ATP synthesis and glucose transport/phosphorylation in type 2 diabetes. PLoS Med. 2007, 4, e154. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.P.; Krausz, K.W.; Xie, C.; Beyoglu, D.; Gonzalez, F.J.; Idle, J.R. Metabolic profiling by gas chromatography-mass spectrometry of energy metabolism in high-fat diet-fed obese mice. PLoS ONE 2017, 12, e0177953. [Google Scholar] [CrossRef] [PubMed]
- Patti, M.E.; Butte, A.J.; Crunkhorn, S.; Cusi, K.; Berria, R.; Kashyap, S.; Miyazaki, Y.; Kohane, I.; Costello, M.; Saccone, R.; et al. Coordinated reduction of genes of oxidative metabolism in humans with insulin resistance and diabetes: Potential role of PGC1 and NRF1. Proc. Natl. Acad. Sci. USA 2003, 100, 8466–8471. [Google Scholar] [CrossRef] [PubMed]
- Ritov, V.B.; Menshikova, E.V.; He, J.; Ferrell, R.E.; Goodpaster, B.H.; Kelley, D.E. Deficiency of subsarcolemmal mitochondria in obesity and type 2 diabetes. Diabetes 2005, 54, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Apontes, P.; Liu, Z.; Su, K.; Benard, O.; Youn, D.Y.; Li, X.; Li, W.; Mirza, R.H.; Bastie, C.C.; Jelicks, L.A.; et al. Mangiferin Stimulates Carbohydrate Oxidation and Protects against High Fat Diet Induced Metabolic Disorders. Diabetes 2014, 63, 3626–3636. [Google Scholar] [CrossRef] [PubMed]
- Kirby, D.M.; Thorburn, D.R.; Turnbull, D.M.; Taylor, R.W. Biochemical assays of respiratory chain complex activity. Methods Cell Biol. 2007, 80, 93–119. [Google Scholar] [PubMed]
- Puigserver, P. Tissue-specific regulation of metabolic pathways through the transcriptional coactivator PGC1-α. Int. J. Obes. 2005, 29, S5–S9. [Google Scholar] [CrossRef] [PubMed]
- Vega, R.B.; Huss, J.M.; Kelly, D.P. The coactivator PGC-1 cooperates with peroxisome proliferator-activated receptor α in transcriptional control of nuclear genes encoding mitochondrial fatty acid oxidation enzymes. Mol. Cell Biol. 2000, 20, 1868–1876. [Google Scholar] [CrossRef] [PubMed]
- Huss, J.M.; Kopp, R.P.; Kelly, D.P. Peroxisome proliferator-activated receptor coactivator-1α (PGC-1α) coactivates the cardiac-enriched nuclear receptors estrogen-related receptor-alpha and -gamma. Identification of novel leucine-rich interaction motif within PGC-1α. J. Biol. Chem. 2002, 277, 40265–40274. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Delerive, P.; Chin, W.W.; Burris, T.P. Requirement of helix 1 and the AF-2 domain of the thyroid hormone receptor for coactivation by PGC-1. J. Biol. Chem. 2002, 277, 8898–8905. [Google Scholar] [CrossRef] [PubMed]
- Scarpulla, R.C. Nuclear control of respiratory chain expression by nuclear respiratory factors and PGC-1-related coactivator. Ann. N. Y. Acad. Sci. 2008, 1147, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, J.T.; Lerin, C.; Haas, W.; Gygi, S.P.; Spiegelman, B.M.; Puigserver, P. Nutrient control of glucose homeostasis through a complex of PGC-1α and SIRT1. Nature 2005, 434, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Sellamuthu, P.S.; Muniappan, B.P.; Perumal, S.M.; Kandasamy, M. Antihyperglycemic Effect of Mangiferin in Streptozotocin Induced Diabetic Rats. J. Health Sci. 2009, 55, 206–214. [Google Scholar] [CrossRef]
- Simi, B.; Sempore, B.; Mayet, M.H.; Favier, R.J. Additive effects of training and high-fat diet on energy metabolism during exercise. J. Appl. Physiol. (1985) 1991, 71, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Leibowitz, S.F.; Dourmashkin, J.T.; Chang, G.Q.; Hill, J.O.; Gayles, E.C.; Fried, S.K.; Wang, J. Acute high-fat diet paradigms link galanin to triglycerides and their transport and metabolism in muscle. Brain Res. 2004, 1008, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Roy, C.; Paglialunga, S.; Fisette, A.; Schrauwen, P.; Moonen-Kornips, E.; St-Onge, J.; Hesselink, M.K.; Richard, D.; Joanisse, D.R.; Cianflone, K. Shift in metabolic fuel in acylation-stimulating protein-deficient mice following a high-fat diet. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E1051–E1059. [Google Scholar] [CrossRef] [PubMed]
- Chen-Zion, M.; Livnat, T.; Beitner, R. Effects of long-term streptozotocin diabetes on cytoskeletal and cytosolic phosphofructokinase and the levels of glucose 1,6-bisphosphate and fructose 2,6-bisphosphate in different rat muscles. Biochem. Med. Metab. Biol. 1994, 53, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Vestergaard, H. Studies of gene expression and activity of hexokinase, phosphofructokinase and glycogen synthase in human skeletal muscle in states of altered insulin-stimulated glucose metabolism. Dan. Med. Bull. 1999, 46, 13–34. [Google Scholar] [PubMed]
- Aithal, H.N.; Walsh-Reitz, M.M.; Toback, F.G. Regulation of glyceraldehyde-3-phosphate dehydrogenase by a cytosolic protein. Am. J. Physiol. 1985, 249, C111–C116. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, C.; Li, H.; Liu, J.P. GAPDH: A common enzyme with uncommon functions. Clin. Exp. Pharmacol. Physiol. 2012, 39, 674–679. [Google Scholar] [CrossRef] [PubMed]
- Modrego, J.; de las Heras, N.; Zamorano-Leon, J.J.; Mateos-Caceres, P.J.; Martin-Fernandez, B.; Valero-Munoz, M.; Lahera, V.; Lopez-Farre, A.J. Changes in cardiac energy metabolic pathways in overweighed rats fed a high-fat diet. Eur. J. Nutr. 2013, 52, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Sellayah, D.; Sek, K.; Anthony, F.W.; Hanson, M.A.; Cagampang, F.R. Sensitivity of housekeeping genes in the hypothalamus to mismatch in diets between pre- and postnatal periods in mice. Neurosci. Lett. 2008, 447, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Ma, X.; Cui, B.; Li, X.; Ning, G.; Wang, S. Selection of reference genes for qRT-PCR in high fat diet-induced hepatic steatosis mice model. Mol. Biotechnol. 2011, 48, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Van Vranken, J.G.; Na, U.; Winge, D.R.; Rutter, J. Protein-mediated assembly of succinate dehydrogenase and its cofactors. Crit. Rev. Biochem. Mol. Biol. 2015, 50, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Bourgeron, T.; Rustin, P.; Chretien, D.; Birch-Machin, M.; Bourgeois, M.; Viegas-Pequignot, E.; Munnich, A.; Rotig, A. Mutation of a nuclear succinate dehydrogenase gene results in mitochondrial respiratory chain deficiency. Nat. Genet. 1995, 11, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, L.M.; Raya, A.I.; Martinez-Moreno, J.M.; Aguilera-Tejero, E.; Rivero, J.L. Mangiferin protects against adverse skeletal muscle changes and enhances muscle oxidative capacity in obese rats. PLoS ONE 2017, 12, e0173028. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, S.; Jainu, M.; Sabitha, K.E.; Shyamala Devi, C.S. Effect of mangiferin on mitochondrial energy production in experimentally induced myocardial infarcted rats. Vasc. Pharmacol. 2006, 44, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Liu, Z.; Apontes, P.; Feng, D.; Pessin, J.E.; Sauve, A.A.; Angeletti, R.H.; Chi, Y. Dual mode action of mangiferin in mouse liver under high fat diet. PLoS ONE 2014, 9, e90137. [Google Scholar] [CrossRef] [PubMed]
- Muruganandan, S.; Srinivasan, K.; Gupta, S.; Gupta, P.K.; Lal, J. Effect of mangiferin on hyperglycemia and atherogenicity in streptozotocin diabetic rats. J. Ethnopharmacol. 2005, 97, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Huang, C.; Liao, X.; Wang, Y.; He, Y.; Feng, R.; Li, Y.; Sun, C. Beneficial effects of mangiferin on hyperlipidemia in high-fat-fed hamsters. Mol. Nutr. Food Res. 2011, 55, 1809–1818. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Li, S.; Na, L.; Feng, R.; Liu, L.; Li, Y.; Sun, C. Mangiferin decreases plasma free fatty acids through promoting its catabolism in liver by activation of AMPK. PLoS ONE 2012, 7, e30782. [Google Scholar] [CrossRef] [PubMed]
- Xing, X.; Li, D.; Chen, D.; Zhou, L.; Chonan, R.; Yamahara, J.; Wang, J.; Li, Y. Mangiferin treatment inhibits hepatic expression of acyl-coenzyme A:diacylglycerol acyltransferase-2 in fructose-fed spontaneously hypertensive rats: A link to amelioration of fatty liver. Toxicol. Appl. Pharmacol. 2014, 280, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Pan, Y.; Chonan, R.; Batey, R.; Rong, X.; Yamahara, J.; Wang, J.; Li, Y. Mitigation of Insulin Resistance by Mangiferin in a Rat Model of Fructose-Induced Metabolic Syndrome Is Associated with Modulation of CD36 Redistribution in the Skeletal Muscle. J. Pharmacol. Exp. Ther. 2016, 356, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Na, L.X.; Zhang, Q.; Jiang, S.; Du, S.S.; Zhang, W.; Li, Y.; Sun, C.H.; Niu, Y.C. Mangiferin supplementation improves serum lipid profiles in overweight patients with hyperlipidemia: A double-blind randomized controlled trial. Sci. Rep.-UK 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Chen, C.; Zhang, C.; Zhang, Y.; Tang, X. Determination of mangiferin in rat plasma by liquid-liquid extraction with UPLC-MS/MS. J. Pharm. Biomed. Anal. 2010, 51, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.Y.; Wang, F.; Li, Y.M.; Li, Y.; Wang, M.Q.; Sun, D.J.; Sun, C.H. Pharmacokinetic study of mangiferin in human plasma after oral administration. Food Chem. 2012, 132, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Chen, H.; Sun, L.; Tong, L.; Zhang, T. Improving permeability and oral absorption of mangiferin by phospholipid complexation. Fitoterapia 2014, 93, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Serasinghe, M.N.; Wieder, S.Y.; Renault, T.T.; Elkholi, R.; Asciolla, J.J.; Yao, J.L.; Jabado, O.; Hoehn, K.; Kageyama, Y.; Sesaki, H.; et al. Mitochondrial Division Is Requisite to RAS-Induced Transformation and Targeted by Oncogenic MAPK Pathway Inhibitors. Mol. Cell 2015, 57, 521–536. [Google Scholar] [CrossRef] [PubMed]
- Jeoung, N.H.; Sanghani, P.C.; Zhai, L.; Harris, R.A. Assay of the pyruvate dehydrogenase complex by coupling with recombinant chicken liver arylamine N-acetyltransferase. Anal. Biochem. 2006, 356, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Constantin-Teodosiu, D.; Cederblad, G.; Hultman, E. A sensitive radioisotopic assay of pyruvate dehydrogenase complex in human muscle tissue. Anal. Biochem. 1991, 198, 347–351. [Google Scholar] [CrossRef]
- Drew, B.; Phaneuf, S.; Dirks, A.; Selman, C.; Gredilla, R.; Lezza, A.; Barja, G.; Leeuwenburgh, C. Effects of aging and caloric restriction on mitochondrial energy production in gastrocnemius muscle and heart. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 284, R474–R480. [Google Scholar] [CrossRef] [PubMed]
- Ajaz, S.; Czajka, A.; Malik, A. Accurate measurement of circulating mitochondrial DNA content from human blood samples using real-time quantitative PCR. Methods Mol. Biol. 2015, 1264, 117–131. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Chow | HFD | HFD/Chow (Ctl) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ctl | MGF | Ratio (MGF/Ctl) | * p (MGF Versus Ctl) | Ctl | MGF | Ratio (MGF/Ctl) | * p (MGF Versus Ctl) | Ratio | * p | |
| UDPG | 7.95 | 7.63 | 0.96 | 0.115 | 6.40 | 5.89 | 0.92 | 0.316 | 0.80 | 0.098 |
| Glucose-6-phosphate | 6.93 | 8.90 | 1.28 | 0.272 | 13.19 | 18.84 | 1.43 | 0.018 | 1.90 | 0.034 |
| Fructose-1,6-bisphosphate | 44.17 | 58.90 | 1.33 | 0.099 | 54.28 | 76.40 | 1.41 | 0.043 | 1.23 | 0.451 |
| 3-Phosphoglycerate | 26.03 | 34.09 | 1.31 | 0.089 | 28.39 | 46.18 | 1.63 | 0.030 | 1.09 | 0.445 |
| Phosphoeolpyruate | 1.58 | 2.18 | 1.38 | 0.054 | 0.89 | 2.27 | 2.54 | 0.024 | 0.57 | 0.025 |
| Pyruvate | 20.48 | 25.11 | 1.23 | 0.175 | 15.41 | 18.94 | 1.23 | 0.083 | 0.75 | 0.038 |
| Lactate | 350.59 | 272.19 | 0.78 | 0.046 | 313.57 | 213.15 | 0.68 | 0.029 | 0.89 | 0.144 |
| Glycerol-3-phosphate | 120.48 | 131.00 | 1.09 | 0.329 | 103.34 | 133.68 | 1.29 | 0.152 | 0.86 | 0.274 |
| Ribose-5-phosphate | 0.39 | 0.43 | 1.09 | 0.216 | 0.44 | 0.47 | 1.05 | 0.688 | 1.13 | 0.406 |
| 6-Phosphogluconate | 1.40 | 1.58 | 1.13 | 0.193 | 1.19 | 1.07 | 0.89 | 0.217 | 0.85 | 0.130 |
| α-Ketoglutarate | 0.45 | 0.67 | 1.48 | 0.043 | 0.28 | 0.62 | 2.20 | 0.041 | 0.63 | 0.035 |
| Succinate | 50.81 | 31.23 | 0.61 | 0.038 | 36.96 | 21.85 | 0.59 | 0.032 | 0.73 | 0.050 |
| Fumarate | 8.17 | 11.24 | 1.38 | 0.049 | 6.01 | 9.09 | 1.51 | 0.041 | 0.74 | 0.049 |
| Malate | 43.89 | 48.31 | 1.10 | 0.121 | 35.64 | 40.45 | 1.13 | 0.322 | 0.81 | 0.084 |
| Aspartate | 26.48 | 29.53 | 1.12 | 0.090 | 19.45 | 20.82 | 1.07 | 0.210 | 0.73 | 0.042 |
| Gene | Forward Sequence from 5′ to 3′ | Reverse Sequencefrom 5′ to 3′ |
|---|---|---|
| HK2 | TGATCGCCTGCTTATTCACGG | AACCGCCTAGAAATCTCCAGA |
| PGI | TGGCCCAGACTGAGGCCCTG | CTCTAGGTGTCTTTATTCTA |
| PFK | CAGATCAGTGCCAACATAACCAA | CGGGATGCAGAGCTCATCA |
| FBA | AGCCTTCTGAGAAGGATGCTC | GTCCAGCATGAAGCAGTTGAC |
| GAPDH | ACTGGCATGGCCTTCCG | CAGGCGGCACGTCAGATC |
| PGK1 | GGTCTTGCCAAAATGTCGCT | TCCAATCGAATCCTCGTGTC |
| PGMb | TGCCCGTCGCCATGGCTGCC- | CGGTGGGACATCATAAGATC- |
| Enolase1 | GCCGGCTTTACGTTCACCTC | GTTGAAGCACCACTGGGCAC |
| CS | AAGGACGAGGCAGGATGAG | TGCAGCTGTAGCTCTCTCCC |
| Idh1 | ATTGGTGGCATCACGATTCT | TGGAGATGCAAGGAGATGAA |
| AKGDH | AGTGGTGGTGGGTAAGTGGA | GCAAAACTTGATCCTCTCGG |
| Suclg1 | TGTTTCCGAGAGGCTGTGTA | CAACCATGGTCTCCAGCAG |
| SDHb | GTCTGTGCCCCTCGACAG | TGACGTCAGGAGCCAAAAT |
| FH1 | ACACGGAAGGAATTTTGGCT | ACCATGTACCGCGCACTC |
| MDH1 | TGCTCCAGTCACAAGGACTC | GACTGCTGGAGACTGCCTTT |
| CoxI (Ndufa9) | TAAGGGATGAAGGTCCGATG | GATCCAGATGCCGTAGGAAA |
| Cyc (cytochrome c) | GGAGGCAAGCATAAGACTGG | TCCATCAGGGTATCCTCTCC |
| CoxIV | AGAAGGAAATGGCTGCAGAA | GCTCGGCTTCCAGTATTGAG |
| Atp5g1 | GCTGCTTGAGAGATGGGTTC | AGTTGGTGTGGCTGGATCA |
| PGC-1α | AAGTGTGGAACTCTCTGGAACTG | GGGTTATCTTGGTTGGCTTTATG |
| PGC-1β | TGCTGCTGTCCTCAAATACG | TGGAGACTGCTCTGGAAGGT |
| NRF | CGGAGTGACCCAAACTGAAC | GATGACCACCTCGACCGTTT |
| ERRα | ACTGCCACTGCAGGATGAG | CACAGCCTCAGCATCTTCAA |
| TFAM | CCAAAAAGACCTCGTTCAGC | ATGTCTCCGGATCGTTTCAC |
| Β-actin | CATGGAGTCCTGTGGCATC | AGCACTGTGTTGGCGTAC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Apontes, P.; Fomenko, E.V.; Chi, N.; Schuster, V.L.; Kurland, I.J.; Pessin, J.E.; Chi, Y. Mangiferin Accelerates Glycolysis and Enhances Mitochondrial Bioenergetics. Int. J. Mol. Sci. 2018, 19, 201. https://doi.org/10.3390/ijms19010201
Liu Z, Apontes P, Fomenko EV, Chi N, Schuster VL, Kurland IJ, Pessin JE, Chi Y. Mangiferin Accelerates Glycolysis and Enhances Mitochondrial Bioenergetics. International Journal of Molecular Sciences. 2018; 19(1):201. https://doi.org/10.3390/ijms19010201
Chicago/Turabian StyleLiu, Zhongbo, Pasha Apontes, Ekaterina V. Fomenko, Nan Chi, Victor L. Schuster, Irwin J. Kurland, Jeffrey E. Pessin, and Yuling Chi. 2018. "Mangiferin Accelerates Glycolysis and Enhances Mitochondrial Bioenergetics" International Journal of Molecular Sciences 19, no. 1: 201. https://doi.org/10.3390/ijms19010201