Critical Role of the Human ATP-Binding Cassette G1 Transporter in Cardiometabolic Diseases


Sorbonne Universités, UPMC Univ Paris 06, Inserm, Institute of Cardiometabolism and Nutrition (ICAN), UMR_S1166, Hôpital de la Pitié, F-75013 Paris, France
*
Author to whom correspondence should be addressed.
†
Current address: INSERM UMR_S1166, Faculté de Médecine UPMC, 91 boulevard de l’hôpital, 75013 Paris, France.
Int. J. Mol. Sci. 2017, 18(9), 1892; https://doi.org/10.3390/ijms18091892
Submission received: 1 August 2017
/
Revised: 30 August 2017
/
Accepted: 30 August 2017
/
Published: 2 September 2017
(This article belongs to the Special Issue Physiological and Pathological Roles of ABC Transporters)
Abstract
:ATP-binding cassette G1 (ABCG1) is a member of the large family of ABC transporters which are involved in the active transport of many amphiphilic and lipophilic molecules including lipids, drugs or endogenous metabolites. It is now well established that ABCG1 promotes the export of lipids, including cholesterol, phospholipids, sphingomyelin and oxysterols, and plays a key role in the maintenance of tissue lipid homeostasis. Although ABCG1 was initially proposed to mediate cholesterol efflux from macrophages and then to protect against atherosclerosis and cardiovascular diseases (CVD), it becomes now clear that ABCG1 exerts a larger spectrum of actions which are of major importance in cardiometabolic diseases (CMD). Beyond a role in cellular lipid homeostasis, ABCG1 equally participates to glucose and lipid metabolism by controlling the secretion and activity of insulin and lipoprotein lipase. Moreover, there is now a growing body of evidence suggesting that modulation of ABCG1 expression might contribute to the development of diabetes and obesity, which are major risk factors of CVD. In order to provide the current understanding of the action of ABCG1 in CMD, we here reviewed major findings obtained from studies in mice together with data from the genetic and epigenetic analysis of ABCG1 in the context of CMD.
1. Introduction to ABC Transporters and Lipid Metabolism
Adenosine triphosphate (ATP)-Binding Cassette (ABC) transporters belong to a large protein family composed of 48 members in humans, divided into 7 families (A–G) [1]. The ABC transporters mediate the active transport of molecules through membranes by hydrolyzing ATP. Thus, ABC transporters are able to drive transport of different types of molecules, from small inorganic or organic molecules to larger organic compounds, such as hormones, amino acids, ions, vitamins and lipids. Exchange of nutrients and metabolites, which are essential for the maintenance of cell or tissue homeostasis, often occurs against concentration gradients across cellular membranes. Therefore, it is not surprisingly that mutations in genes encoding ABC transporters lead to impaired homeostasis and genetic disorders associated with diseases. More especially, mutations in genes coding for ABC transporters involved in lipid transport are responsible for alterations in cellular lipid homeostasis and cause lipid-related disorders which may be associated with an increased risk of cardiovascular diseases (CVD) [2,3].
Among them, patients carrying mutations in ABCG5 or ABCG8 genes exhibit Sitosterolemia [4,5], a pathology characterized by elevated plasma levels of plant sterols due to an exacerbated absorbance of dietary phytosterols from the intestine. Indeed, the ABCG5/ABCG8 dimer promotes the efflux of cholesterol and plant sterols from both hepatocytes to bile and enterocytes to lumen. Patients harboring mutations in either ABCG5 or ABCG8 have premature atherosclerosis and an increased risk to develop CVD [6]. In addition, mutations in ABCA1 cause Tangier disease, a pathology characterized by a quasi-absence of high-density lipoprotein (HDL) and the appearance of lipid depots in tonsils, liver, spleen and lymph nodes [7,8,9]. Indeed, ABCA1 promotes cholesterol and phospholipid efflux to lipid-poor apolipoprotein A-I (ApoA-I) [10] and through this mechanism expression of ABCA1 in liver and intestine is crucial for HDL biogenesis [11]. Cholesterol efflux to HDL from macrophages in the arterial wall is proposed to protect against macrophage lipid accumulation and atherosclerosis development. Thus, mutations in ABCA1 are associated with a premature atherosclerosis and an increased CVD risk [12,13].
Although no genetic disorder caused by mutations in the ABCG1 gene is reported so far, the ABCG1 transporter draws a special attention since it is described to promote cellular cholesterol efflux to HDL [14,15]. The membrane ABCG1 transporter was initially thought to protect from atherosclerosis and CVD. However, investigation of the role of ABCG1 in mouse models as well as in human studies seems to indicate that the control of cellular lipid homeostasis operated by ABCG1 might play an important role in the wide spectrum of metabolic disorders, including obesity, diabetes and insulin resistance, which are risk factors for atherosclerosis and CVD.
The goal of the present review is therefore to provide a comprehensive view of the action of ABCG1 in lipid metabolism and lipid-related metabolic disorders in the context of cardiometabolic diseases (CMD). In addition to major findings obtained from studies in mice, the present review mainly focusses on the analysis of genetic and epigenetic modulation of ABCG1 in relation to CMD.
2. Structure and Regulation
The human ABCG1 gene was initially identified as the human homolog of the Drosophila melanogaster white gene which encodes a member of the ABC transporter family involved in guanine and tryptophan import, two precursors of Drosophila eye color pigments [16]. In humans, ABCG1 gene has been mapped to chromosome 21q22.3 [17] and multiple human ABCG1 transcripts have been detected resulting from different transcription initiation and alternative mRNA splicing [18,19,20]. Similar to others members of the ABC family (ABCA1, ABCG4, ABCG5 and ABCG8), the expression of human ABCG1 is highly stimulated by cholesterol loading and oxysterols [14,21] through the liver X receptors (LXR) pathway. Indeed LXR acts as a cholesterol sensor which is activated by oxysterols (cholesterol oxidative derivatives) when the cholesterol concentration increases in the cell. Induction of ABCG1 expression by LXR agonists likely involves the presence of multiple LXR responsive elements (LXRE) through the ABCG1 gene [20,22] and appears to only require the LXRα isoform in human macrophages [23]. Analysis of the molecular mechanism indicates that LXR recruitment at the human ABCG1 locus is facilitated by the G protein pathway suppressor 2 (GPS2) [24]. Although ABCG1 shares common gene expression regulatory pathways with others ABC family members, ABCG1 clearly exhibits a tissue specific expression pattern with high expression levels of ABCG1 found in lung, brain, spleen, adrenal glands, heart and liver [14,17].
Although discrepancies were initially found in the literature regarding the structure of the ABCG1 gene [18,19,20], it is now well established that the ABCG1 gene is composed of 23 exons encoding a protein forming a half transporter with 6 transmembrane spanning domains and a single intracellular nucleotide binding domain (NBD) (Figure 1). This NBD domain contains highly conserved Walker A and Walker B motifs and is required for the binding and the hydrolysis of ATP [25,26] which might provide required energy to transport substrates across the membrane [27]. An alternative splicing at the end of exon 17 led to the generation of a spliced variant of ABCG1 which lacks an internal segment of 12 amino acids in the region between the cytoplasmic domain and the ATP cassette (ABCG1(−12)) [28,29]. Interestingly, the two ABCG1 isoforms, ABCG1(+12) and ABCG1(−12), are expressed in humans whereas mice only express the shorter isoform putting forward the limitations of extrapolating results from mouse models to the human context [30]. Although both isoforms exhibit similar activity, expression of the ABCG1(−12) isoform is roughly two fold more abundant than the ABCG1(+12) isoform in human macrophages and vascular cells [28,29]. Indeed, Gelissen et al. demonstrated that the ABCG1(+12) isoform is more rapidly degraded as the result of the phosphorylation of a serine residue located in the 11 amino acid segment by protein kinase A (PKA) [31]. Others mechanisms contribute to the stability and activity of human ABCG1. Thus, ABCG1 degradation is proposed to be regulated via the ubiquitin proteasome system by the E3 ubiquitin ligases HUWE1 (HECT, UBA, and WWE domain containing 1, E3 ubiquitin protein ligase) and NEDD4-1 (Neural precursor cell-expressed developmentally down regulated gene 4) [32]. Interestingly, ubiquitination and proteasomal degradation of human ABCG1 was suppressed by cholesterol through interaction with a cholesterol recognition/interaction amino acid consensus (CRAC) motif located in the final transmembrane domain of ABCG1 [33,34].
A debate still occurs regarding the cellular localization of ABCG1. Indeed, the use of expression vectors containing the human ABCG1 cDNA sequence fused or not to a tag protein for ABCG1 visualization led to conflicting results with ABCG1 being predominantly detected at the plasma membrane [27,28,35,36,37] or/and in the intracellular compartment [14,38,39,40]. Indeed, localization of human ABCG1 was reported at the cell surface and in the endocytic pathway (early, recycling, late endosomes and lysosomes) as well as in the Golgi apparatus [14,28,35,38,39,40] and ABCG1 was proposed to traffic along the secretory pathway from the endoplasmic reticulum to the cell surface as well as along the endocytic pathway to the late endosomes/lysosomes [41]. The presence of Leu 562 [37] or both Asn 316 and Phe 320 in the highly conserved sequence located between the NBD and the transmembrane domain [42] has been reported to be important for ABCG1 localization at the plasma membrane.
3. Functions and Mechanism of Action
The ABC transporters are membrane-bound proteins that mediate the ATP-dependent translocation of many amphiphilic and lipophilic molecules including lipids, drugs or endogenous metabolites. Among them, twenty transporters have been shown to be implicated in lipids, lipid-like or steroids transport [43]. The ABCG1 transporter is a half ABC protein that must dimerize to form an active transporter. It has been reported that human ABCG1 can form homodimers [38] or heterodimers, especially with ABCG4 [26,44]. Although others potential partners for ABCG1 may exist, it is very likely that ABCG1 mainly acts as a homodimer since ABCG1 and ABCG4 are generally spatially separated at the exception of the brain [45]. Human ABCG1 was identified as a lipid transporter able to promote the export of numerous lipid species including cholesterol, phospholipids (phosphatidylcholine), sphingomyelin (SM) and oxysterols (7-ketocholesterol, 7β-hydroxycholesterol and 25-hydroxycholesterol) [36,38,46,47,48,49,50]. However, it appears that ABCG1 exerts a broader array of substrates which is not restricted to lipids as ABCG1 can equally promote export of liposoluble molecules such as vitamin E (α- and γ-tocopherol) [51]. In contrast to ABCA1, ABCG1 promotes cellular cholesterol efflux to high density lipoproteins (HDL2 and HDL3) but not to lipid-free ApoA-I; the phospholipid content of the acceptor being the major parameter driving ABCG1-mediated cholesterol efflux [38,46]. Importantly, lipidation of ApoA-I by ABCA1 generates nascent or preβ-HDL which may serve as a substrate for ABCG1 indicating that ABCA1 and ABCG1 act sequentially to export cellular cholesterol efflux [38,46]. However, even if HDL is the most efficient lipid acceptor for promoting lipid efflux through ABCG1, the latter may also occur in the presence of non-specific acceptors such as serum albumin although to a lesser degree considering the efflux of cholesterol [36,50].
The mechanism of action of ABCG1 is not yet fully understood and is controversial. A prevailing view is that ABCG1 acts within the endocytic pathway and at the plasma membrane by redistributing membrane cholesterol through the vesicular pathway between endoplasmic reticulum and plasma membrane. The reorganization of membrane cholesterol and SM operated by ABCG1 might increase their accessibility to extracellular acceptors for removal allowing a reduction of lipid rafts formation at the plasma membrane [41,52,53]. However, this process might occur independently of the presence of such acceptors and cholesterol might desorb in a nonspecific manner when ABCG1 is expressed [54].
4. Critical Role of ATP-Binding Cassette G1 (ABCG1) in Cardiometabolic Diseases
Identification of a role of ABCG1 in promoting cellular lipid efflux to extracellular acceptors led to propose that ABCG1 may be essential to maintain cell or tissue lipid homeostasis in metabolic contexts associated with major perturbation of lipid homeostasis and metabolism as classically observed in CMD. However, no genetic disease caused by ABCG1 mutations has been documented and generation of mouse models with targeted disruption of Abcg1 or expression of human ABCG1 helped to appreciate the contribution of ABCG1 in physiological and physiopathological situations. Although major finding from studies in mice will be reported here, we will mainly focus on information available when human ABCG1 is explored at both genetic and epigenetic levels which are of major importance for uncovering association of ABCG1 with metabolic traits and providing insights in CMD.
5. ABCG1 and Lipid Metabolism and Homeostasis
5.1. Major Findings from Studies in Mice
In agreement with a role of ABCG1 in promoting cellular cholesterol efflux, the use of Abcg1−/− and human ABCG1 transgenic mice confirmed that ABCG1 has a critical role in maintaining tissue lipid homeostasis when mice were challenged with a high-fat and high-cholesterol diet [55]. Indeed although neutral lipid accumulation was observed in lung from deficient-Abcg1 mice on a chow diet [55,56], a massive elevation of the amount of cholesterol ester, triglycerides (TG) and phospholipids was detected in hepatocytes and macrophages from liver and lung when deficient-Abcg1 mice were fed a high-fat and high-cholesterol diet. By contrast, animals expressing the human ABCG1 transgene were protected from lipid accumulation in those tissues [55]. Although not the topic of the present review, it is however important to mention here that ABCG1 plays a key role in maintaining lung lipid homeostasis and that impaired ABCG1 expression was reported in pulmonary alveolar proteinosis (PAP) [57]. Indeed, PAP is characterized by deficient surfactant clearance and lipid accumulation in alveolar macrophages which were proposed to result from a defective ABCG1 expression [57,58,59].
The apparent preponderant role of macrophage Abcg1 to export cholesterol was confirmed in peritoneal macrophages from Abcg1-deficient mice which exhibited a reduced cellular cholesterol efflux to HDL but not to lipid free ApoA-I in comparison to peritoneal macrophages from wild-type mice when stimulated with LXR ligands. Despite a role of ABCG1 in promoting cellular cholesterol export to HDL, lack of Abcg1 in mice has no consequence on plasma lipid levels, especially HDL-C. Indeed, both Abcg1-deficient mice and human ABCG1 transgenic mice fed either a chow diet or a high-fat and high-cholesterol diet exhibited no differences in plasma lipid levels as compared to wild-type mice [55,60,61]. However, Wiersma et al. observed a less pronounced increase in plasma HDL-C in Abcg1 knockout (KO) mice following a 2-week period of high-cholesterol diet or when fed a chow diet after LXR agonist (T0901317) administration, suggesting that LXR activation is required to elicit an effect of Abcg1 on plasma HDL-C levels [62].
5.2. Genetic Modulation of Human ABCG1
Development of genome- and epigenome-wide association studies (GWAS and EWAS) provides very useful tools to study the association between genetic and epigenetic variations of the human ABCG1 gene and metabolic phenotypes. Numerous polymorphisms have been identified in the ABCG1 locus [63] and several functional single nucleotide polymorphisms (SNP) located in the promoter region as well as in the coding region of the ABCG1 gene served as genetic markers to explore the association between ABCG1 and plasma lipid levels and to decipher the role of ABCG1 in CMD. Genotyping of a functional ABCG1 SNP (rs1378577, T > G) located in the promoter region of the ABCG1 gene in 109 Japanese men with a coronary artery disease (CAD) did not allow to detect any statistical differences in serum TG, total cholesterol (TC), low-density lipoproteins (LDL)-C and HDL-C according to the ABCG1 genotype [64]. Similar observations were reported in 609 dyslipidemic white men with a minimum 50% obstruction of a major coronary artery in regression growth evaluation satin study (REGRESS) when two different SNPs located in the ABCG1 promoter (rs1378577, T > G and rs1893590, A > C) were studied [65]. Indeed, these two ABCG1 SNPs were associated with neither plasma lipid concentrations (TC, LDL-C, HDL-C, TG and Lipoprotein(a)) nor circulating cholesterol ester transfer protein (CETP) concentrations. Nevertheless in this study, the authors brought to light an unexpected association between both ABCG1 SNPs and plasma lipoprotein lipase (LPL) activity independently of any effect on circulating LPL mass [65]. However the absence of association between the two SNPs and TG levels observed in REGRESS (plasma TG levels < 3.5 g/L) suggests that modulation of LPL activity by ABCG1 might only alter circulating TG concentrations in subjects exhibiting fasting or postprandial hypertriglyceridemia. The absence of any association between ABCG1 promoter SNPs (rs2234714, −768G > A; and rs57137919, −367G > A) and plasma lipids levels (HDL-C, LDL-C, TG and TC) was equally reported in 1021 patients with CAD and 1013 unaffected control subjects in a Chinese Han population [66]. Genotyping of the rs57137919 in 200 Chinese healthy volunteers led to similar conclusions [67]. Association studies in larger cohorts enabled detection of an association between ABCG1 genotype and plasma HDL-C levels. Indeed, genotyping of 10,237 individuals from the Copenhagen city heart study (CCHS) for 16 ABCG1 SNPs identified two rare variants (rs140837853, C > T and rs56140811, C > T) located in the promoter region and exon 9 of the ABCG1 gene that were associated with modest reductions in HDL-C (7% and 3%, respectively); the carriers of the rare alleles exhibited lower HDL-C levels than the non-carriers [68]. However except for HDL-C, none of those ABCG1 variants were associated with plasma lipids, lipoproteins and apolipoproteins. Moreover, a high-density genotyping array containing SNPs from HDL-C candidates provided suggestive evidence for an association of an intronic ABCG1 SNP (rs914189, C > G) with HDL-C concentrations in a meta-analysis comprising 7857 individuals [69]. Interestingly, Abellán et al. reported that the ABCG1 SNP rs1893590 (A > C) was associated with plasma HDL-C levels in the postprandial state while no association was detected with postprandial TC or LDL-C [70]. Indeed, in 1473 Spanish subjects from the population-based Hortega study, the carriers of the rare C allele (CC + AC) were significantly associated with higher postprandial HDL-C concentrations 3 h after lunch as compared to individuals homozygous for the frequent A allele (genotype AA) [70]. Strikingly, this association was only observed in women but not in men suggesting a gender-specific effect of the ABCG1 genotype on postprandial HDL-C levels. Since people in Western countries spend most of the day in the postprandial state, this study provides new insights regarding the potential contribution of ABCG1 in determining circulating levels of HDL-C, which are firmly reported to be inversely associated to CAD [71]. However, the diet intake appears to be essential in the effect of ABCG1 on plasma HDL-C levels. Indeed, the association between rs1893590 and postprandial HDL-C disappeared after adjustment for caloric intake or lipid consumption [70] whereas an association with the ABCG1 SNP rs1044317 (G > A, 3’UTR region) with variations in fasting HDL-C concentrations was only observed in subjects with a high polyunsaturated fatty acid (PUFA) intake (>13.6 g/day) [72]. A similar diet-interaction was reported regarding the association of the ABCG1 SNP rs4148102 (G > A, intronic region) with TC and LDL-C in this latter study [72]. To note that the body mass index (BMI) and age equally interact in the relation between ABCG1 polymorphisms and plasma HDL-C. Thus, the association between the ABCG1 SNP rs1893590 (A > C) with HDL-C in an asymptomatic Brazilian population (654 normolipidemic volunteers) was only detectable for individuals under 60 years or BMI < 25 kg/m2 with homozygous AA subjects displaying higher plasma HDL-C levels than carriers of the C allele (AC + CC) [73]. Finally, rare ABCG1 SNPs were reported to be associated with plasma TC (rs170444, A > G) and TG levels (rs170444, A > G and rs8126601, G > A) in a large-scale meta-analysis across 32 studies (66,240 individuals of European ancestry) [74]. However no association between ABCG1 SNPs (rs1378577, −134 T > G and rs1893590, −204 A > C) with circulating lipids levels was observed in a population of morbidly obese patients (BMI > 40 kg/m2, n = 1320) [75].
5.3. Epigenetic Modulation of Human ABCG1
Epigenetic mechanisms such as DNA methylation might contribute to the determination of circulating lipid concentrations in humans [76]. In this context, recent evidence indicates that epigenetic marks within the ABCG1 gene including cytosine guanine dinucleotide (CpG) site methylation are associated with blood lipid levels. Genome-wide DNA methylation analysis in whole blood samples of 1776 subjects from the general population of the cooperative health research in the region of Augsburg (KORA) identified CpG sites located in ABCG1 associated with HDL-C and TG levels [77]. Indeed, cg27243685 and cg07397296 were strongly associated with TG levels whereas cg06500161 was associated in opposite directions with plasma HDL-C and TG levels. Interestingly, association of ABCG1 DNA methylation with TG levels was also found in adipose tissue, a tissue in which ABCG1 gene was expressed. Methylation of ABCG1 at cg06500161 was negatively associated with ABCG1 mRNA levels indicating that this CpG site was functional in repressing ABCG1 expression [77,78,79]. Moreover, functional analysis of cg06500161 by electrophoretic mobility shift assay revealed a higher binding of a protein complex for the unmethylated status of the CpG site as compared to the methylated status, suggesting that cg06500161 modulates transcriptional activity of the ABCG1 promoter. In agreement with those observations, ABCG1 mRNA levels showed a strong positive association with HDL-C and a negative association with TG levels [77].
The association of DNA methylation at ABCG1 locus in whole blood DNA with plasma TG and HDL-C levels was replicated in several studies recapitulated in Table 1 which were conducted in the Botnia prospective study (cg06500161, TG, n = 258) [80], the Rotterdam study (cg06500161, TG and HDL-C, n = 1485) [78], the Framingham heart study (FHS) and prospective investigation of the vasculature in uppsala seniors study (PIVUS) (cg06500161, TG and HDL-C, cg27243685 and cg01176028, TG, n = 2036) [79] and in Canadian familial hypercholesterolemia patients (CpGC3, TG, HDL-C and TC, n = 98) [81]. In addition, Lai et al. reported that methylation of the ABCG1 CpG site (cg06500161) in blood CD4+ T cells from 979 subjects of the genetics of lipid lowering drugs and diet network (GOLDN) study was significantly associated with postprandial hypertriglyceridemia (PPHT) after consumption of a high-fat test meal [82]. Indeed, an increased methylation at ABCG1 was correlated with an increased PPHT response. However cg06500161 was equally associated with fasting TG, then the effect on PPHT disappeared after adjustment for baseline fasting TG levels. Finally, beyond TG and HDL-C, EWAS between DNA methylation and lipidomic traits in human blood revealed an association between methylation at cg06500161 and SM, phosphatidylcholine, lysophosphatidylcholine and monoacylglycerol [79,83].
Taken together, the use of polymorphisms in the ABCG1 gene as genetic markers in association studies appears poorly informative in order to reveal a potential impact of ABGC1 on lipid metabolism. However, the results from DNA methylation studies univocally revealed a strong association of ABCG1 with circulating TG levels and in a lesser extend those of HDL-C. The inverse association between methylation at cg06500161 and HDL-C is coherent with the largely described role of ABCG1 in promoting cellular cholesterol efflux to HDL and suggests that DNA methylation of ABCG1 in key tissues controlling HDL metabolism (liver or intestine) might contribute to the determination of circulating HDL-C levels. In addition, the strong positive relationship existing between DNA methylation at the ABCG1 locus and circulating TG levels is in agreement with the role of ABCG1 in controlling the bioavailability and activity of LPL, a key enzyme in TG metabolism [65]. However, it must be kept in mind that although DNA methylation may modulate circulating lipid concentrations, an inverse relationship may equally occur. Indeed, Dekkers et al. demonstrated that blood lipids influenced DNA methylation with methylation of ABCG1 CpG sites (cg06500161 and cg27243685) being modulated by TG or HDL-C [84].
6. ABCG1: Role in Atherosclerosis and Cardiovascular Diseases
Cardiovascular diseases are one of the major causes of mortality and morbidity worldwide and are frequently associated to atherosclerosis, a physiopathological process leading to lipid accumulation, mainly in macrophages in the arterial wall. Epidemiological studies have largely reported that plasma LDL-C levels are positively associated to CAD whereas an inverse relationship exists with those of HDL-C [71].
6.1. Major Findings from Studies in Mice
Because of the important role of Abcg1 in maintaining tissue lipid homeostasis, especially in macrophages [55], Abcg1 was proposed to protect from atherosclerosis development by promoting cholesterol efflux to HDL from arterial macrophages. In addition, ABCG1 was reported to protect macrophage from apoptosis induced by oxidized LDL [85] as well as against endothelial dysfunction [48] and activation [86] in mice, which are key features of atherosclerosis. However investigation of the role of Abcg1 in atherosclerosis in animal models led to conflicting results. Indeed, the expression of human ABCG1 in a rabbit fed a high-cholesterol diet [87] or in Ldlr−/− [88] or Apoe−/− [61] mice fed a high-fat and high-cholesterol diet showed either attenuation or increased or no effect on atherosclerosis, respectively. In addition, Abcg1 deficiency in mice under high-fat and high-cholesterol feeding was either protective or deleterious against atherosclerosis in wild-type mice [60] or in Apoe−/− mice [89], respectively. In order to evaluate the specific contribution of macrophage Abcg1 in atherosclerosis development, bone marrow transfer (BMT) from donor Abcg1−/− mice into recipient Ldlr−/− or Apoe−/− mice fed a high-fat and/or a high-cholesterol diet was simultaneously conducted in three different laboratories [89,90,91,92,93,94]. However, consistent with what was observed for modulation of whole body Abcg1 expression, those studies led to contrasting results which did not resolve the role of macrophage Abcg1 in atherosclerosis. In an attempt to reconcile the results obtained in Ldlr−/− mice, Meurs et al. proposed that the effect of Abcg1 deficiency on atherosclerosis depends on the stage of the disease [95]. Thus, Abcg1 would be protective in early lesions likely by promoting macrophage cholesterol efflux and reducing foam cell formation whereas Abcg1 would be deleterious in advanced lesions by protecting macrophage from apoptosis through the export of cellular oxysterols. However further investigations are needed to validate this model and to elucidate the whole spectrum of action of Abcg1 in the process of atherogenesis in which Abcg1 appears to exert a more complex role that initially thought.
Finally, since Abcg1 was demonstrated to cooperate with Abca1 in promoting cholesterol efflux from macrophages [38,46], Abca1/Abcg1 double KO mice have been generated in order to determine if the impact of macrophage Abcg1 deficiency in atherogenesis could be unmasked when Abca1 is absent. A much more pronounced reduction of cholesterol efflux to HDL was observed in double Abca1/Abcg1-deficient macrophages in comparison to single KO mice whereas cholesterol efflux to ApoA-I was not affected [92,96]. However, similar to what was observed in single Abcg1-deficiency, BMT from donor Abca1−/−Abcg1−/− mice into recipient Ldlr−/− mice led to conflicting results and did not help to elucidate the role of macrophage Abcg1 in atherosclerosis [92,93]. Indeed, an exacerbated size of atherosclerotic lesions was detected in Ldlr−/− mice transplanted with Abca1−/−Abcg1−/− BM fed a high-cholesterol diet in comparison to single KO animals [92] whereas an independent study showed no difference in lesion size between Ldlr−/− mice transplanted with either Abca1−/−Abcg1−/− or Abcg1−/− BM when animals were fed a high-fat and high-cholesterol diet [93].
6.2. Genetic Modulation of Human ABCG1
Whole genome expression arrays performed in circulating monocytes (CD14+ cells) from young male with premature familial CAD and control individuals (n = 22 in each group) revealed that ABCG1 expression was down-regulated in CAD patients suggesting that ABCG1 might be associated with early atherosclerosis [97]. However, association studies between ABCG1 SNPs and CAD or atherosclerosis led to conflicting results. Indeed, analysis of the effect of the functional ABCG1 SNP (rs1378577, T > G) on CAD severity in 109 Japanese men with CAD indicated that individuals homozygous for the T allele (TT) exhibited an increased risk of multi-vessel disease compared with single-vessel disease which was independent of any effect on plasma lipid levels [64]. A protective role of ABCG1 regarding CAD was also reported by Schou et al. who identified two ABCG1 variants associated with an increased risk for myocardial infarction (MI) and ischemic heart disease (IHD) (rs72542412, −376 C > T and S630L) and one ABCG1 variant only associated with MI (rs138515663, −311 T > A) in 10,237 individuals from theCCHS [68]. It is worthy to note that those very rare variants were not associated with plasma HDL-C levels. Conversely, ABCG1 SNPs associated with circulating HDL-C concentrations were not found associated to either MI or IHD indicating that the effect of ABGC1 variation on the risk of ischemic vascular disease was not the result of any modulation of HDL-C. However, genotyping of two ABCG1 SNPs located in the ABCG1 promoter (rs1378577, −134 T > G and rs1893590, −204 A > C) in REGRESS indicated that those SNPs were not associated with angiographic parameters (minimum segment diameter and mean obstruction diameter) in individuals (n = 609) developing atherosclerosis [65]. On the contrary, analysis of the association of four ABCG1 polymorphisms, three located in the promoter (rs2234714, −768 G > A; rs2234715, A > G and rs57137919, −367 G > A) and one in the 3’UTR region (rs1044317, G>A) in patients with CAD (n = 1021) and unaffected control subjects (n = 1013) in a Chinese Han population concluded to a deleterious role of ABCG1 regarding CAD [66]. Indeed, a significant difference in allele frequency and genotype distribution of both rs2234714 and rs57137919 was observed between CAD patients and control subjects with the frequency of the less minor allele being lower in the CAD group (with MI or not) than in controls. Thus, those polymorphisms showed a decreased risk for CAD (rs2234714 and rs57137919) and MI (rs57137919) and led to identification of a GAGA haplotype (rs2234714, rs2234715, rs1044317 and rs57137919) including the more frequent allele of each SNP that was associated with an increased risk of CAD or MI [66]. In addition, rs57137919 was shown to be associated with angiographic severity of CAD (multi-vessel versus single vessel). Interestingly, those SNPs were not in apparent linkage disequilibrium and were not associated with blood lipid concentrations. In order to elucidate the mechanism underlying the deleterious effect of the ABCG1 SNP rs57137919 in CAD, the same group extended their investigation by carrying out functional experiments in 200 Chinese healthy volunteers [67]. In this study, human monocyte-derived macrophages from individuals carrying the rare AA genotype for the rs57137919 displayed reduced ABCG1 expression, impaired oxidized NBD-cholesterol efflux to HDL and increased apoptosis as compared to macrophages from individuals carrying the frequent GG genotype. In addition, mRNA levels of proapoptotic Bcl-2 related ovarian killer (BOK) and BH3 interacting domain death agonist (BID) genes from macrophages with the AA genotype were more elevated than those from macrophages with the GG genotype following exposure with oxidized LDL. Those results suggest that the reduced risk of developing atherosclerosis observed in individuals carrying the less frequent rs57137919 A allele could result from an increased macrophage apoptosis, thereby supporting the model proposed by Meurs et al. from studies in mice about the role of Abcg1 in atherosclerosis [95]. Finally, the association of two functional ABCG1 SNPs (rs1378577, T > G and rs571137919, G > A) with the risk of ischemic stroke in a case-control study including 389 ischemic stroke patients and 380 healthy subjects in the Chinese Han population provided additional mechanistic insights about the deleterious effect of ABCG1 [98]. Indeed, Li et al. demonstrated that the frequency of homozygosity for the rare alleles (GG and AA) was lower in patients as compared to control individuals in a subgroup of hypertriglyceridemia whereas genotypic distribution was similar in patients and controls when the whole population was studied. In addition, both rs1378577 and rs571137919 were associated with a reduced risk of developing ischemic stroke in hypertriglyceridemic individuals. Although the sample size was here relatively low, this study supports the mechanism proposed by Olivier et al. through which ABCG1 could exert a deleterious role in metabolic situations associated with high levels of circulating TG-rich lipoproteins [65]. Indeed, Olivier et al. demonstrated that ABCG1 controls LPL activity and promotes lipid accumulation in macrophages in a presence of TG-rich lipoproteins and, therefore, might contribute to foam cell formation and atherosclerosis development in hypertriglyceridemic individuals.
6.3. Epigenetic Modulation of Human ABCG1
Analysis of the relationship between ABCG1 promoter methylation and coronary heart disease (CHD) was investigated in a small group of 85 CHD patients and 54 participants without CHD in the Han Chinese population [99]. In this study, ABCG1 promoter hypermethylation was frequently observed in patients with CHD (90.5%) but not in individuals without CHD (29.6%). Further analysis indicated that hypermethylation of the ABCG1 promoter gene was associated with an increased risk of CHD after adjustment for age, gender, smoking, lipid levels, hypertension and diabetes. In support of this preliminary study, analysis of genome-wide DNA methylation in whole blood in subjects (n = 1776) from the general population of the KORA study indicated that the ABCG1 CpG site cg06500161 was associated with MI [77]. This effect was independent of the association of cg06500161 with HDL-C and TG in this cohort. However, this result must be interpreted with caution since only 60 individuals with previous hospitalized MI were included in this analysis. A very recent study reported that whole blood methylation at the ABCG1 CpG site cg27243685 (5’UTR region of ABCG1) was significantly associated with CHD in a meta-analysis of theFHS and PIVUS (2306 participants, number of CHD events = 193). Altogether, this study highlighted a pathway linking hypermethylation of ABCG1 at cg27243685 with reduced ABCG1 expression, higher TG, lower HDL-C and increased risk for CHD [79]. Measurement of blood DNA methylation in Canadian Familial Hypercholesterolemia patients did not allow to establish any association between methylation levels at a single ABCG1 CpG site (CpGC3) and a prior history of CAD in a small subset of individuals (Non-CAD (n = 22) vs CAD (n = 22)) [81].
To conclude, association studies in humans using methylation marks at the ABCG1 locus support a protective role for ABCG1 in CAD in the general population with methylation levels at ABCG1 CpG sites being associated with an increased risk of CHD and MI. However, this effect is independent of any effect on blood lipid concentrations. Thus, modulation of DNA methylation by environmental factors such as smoking and nutrition might play an important role in the contribution of ABCG1 in CAD. Nevertheless, genome-wide DNA methylation analysis in large cohorts of CAD patients or in case-control studies for CAD will be required to conclude on the impact of methylation at the ABCG1 locus on CAD. By contrast, results from the genotyping of ABCG1 SNPs in CAD patients or in case-control studies for CAD support the contention that ABCG1 might exert a pro-atherogenic effect locally in the arterial wall independently of any modulation of circulating HDL-C or TG levels [100]. Such a deleterious role could result from the critical contribution of human ABCG1 in reducing macrophage apoptosis through efflux of oxysterol [50,101] and in promoting macrophage lipid accumulation through modulation of LPL activity in a TG-rich metabolic context [65]. In contrast to what is proposed in mice, human ABCG1 does not promote macrophage cholesterol efflux to HDL [52] and therefore is not expected to attenuate foam cell formation in early atherosclerosis lesions. However, ABCG1 could protect from atherosclerosis by preserving vascular endothelium from dietary cholesterol-induced dysfunction [48,102].
7. ABCG1: Role in Diabetes and Insulin Resistance
The presence of diabetes and insulin resistance (IR) is frequently observed in CMD. In addition, diabetes is growing worldwide because of the emergence of obesity and is a risk factor for accelerated atherosclerosis and CAD. The loss of insulin signaling or secretion as well as the loss of glycemic control are critical features that characterize type 2 diabetes (T2D).
7.1. Major Findings from Studies in Mice
Macrophages from diabetic mice (db/db mice or treated with streptozotocin) showed a decrease of Abcg1 expression and an impaired cellular cholesterol efflux to HDL [103,104]. As a consequence, an increased accumulation of esterified cholesterol was observed in diabetic db/db macrophages [103]. Interestingly, Mauldin et al. also demonstrated that elevated glucose led to a repression of Abcg1 expression in mouse macrophages, suggesting that hyperglycemia in diabetes could contribute to foam cell formation and accelerated atherosclerosis through inhibition of macrophage Abcg1-mediated cholesterol efflux. Advanced glycation end products (AGE), which are generated by non-enzymatic glycation and oxidation of proteins and lipids, accumulate in diabetes. Treatment of macrophages with AGEs led to a decreased ABCG1 expression through a peroxisome proliferator-activated receptor (PPAR)γ-dependent mechanism and a reduction of cholesterol efflux to HDL. Generation of mice deficient for the receptor of AGEs (RAGE/AGER) revealed that macrophage reverse cholesterol transport to feces was reduced in diabetic mice through RAGE [104]. Moreover, Nagelin et al. reported that macrophage Abcg1 expression was significantly suppressed by PUFAs such as linoleic or arachidonic acids which frequently accumulate in diabetes and IR [105]. Interestingly, those PUFAs are ligands for 12/15-lipoxygenase (12/15LO) for producing hydroperoxy FA 12S- and 15S-hydroxyeicosatetranoic acids (12SHETE/15SHETE) and 13S-hydroxyoctadecadienoic acid (13SHODE) and 12/15LO was reported to play a role in diabetes and atherosclerosis. Genetic manipulation of 12/15LO in mice indicated that 12/15 LO regulates the degradation of Abcg1 through p38- and c-Jun N-terminal kinase 2 (JNK2)-dependent pathways and macrophage cholesterol efflux to HDL [106].
Beyond macrophages, Abcg1 was demonstrated to play a major role in regulation of subcellular cholesterol distribution in mouse pancreatic β cells [107]. Indeed, loss of Abcg1 in β cells reduced granule cholesterol content and altered granule morphology leading to an impaired insulin secretion. Thus, Abcg1−/− mice fed a chow diet exhibited an impaired glucose tolerance and insulin secretion as compared to Abcg1+/+ animals whereas insulin sensitivity was not altered. The relevance of those findings was strengthened by the observation that islet Abcg1 expression was decreased in hyperglycemic diabetic db/db mice and restored by the antidiabetic thiazolidinedione (PPARγ agonist). However, although insulin secretion was equally reduced in Abcg1−/− mice fed a chow diet in the work from Kruit et al., glucose tolerance was not affected in Abcg1−/− mice when compared to Abcg1+/+ animals [108]. On the contrary, Abcg1-deficient mice fed a high fat diet were protected from glucose intolerance in comparison to their wild-type littermates [109]. Taken together, a large body of evidence suggests that Abcg1 could play a role in diabetes as well as in accelerated atherosclerosis associated to diabetes through modulation of insulin secretion and foam cell formation, respectively. However, it is still difficult to evaluate the impact of the impaired insulin secretion consecutive to Abcg1 deficiency in diabetes since glucose intolerance was not a common feature in Abcg1−/− mice. In addition, although macrophage Abcg1 expression was consistently found reduced in diabetic mice or in response to stimuli relevant to diabetes (PUFA, glucose, AGEs), the impact of such a reduction on macrophage cholesterol efflux and potentially atherogenesis was not evaluated directly in experimental models in which Abcg1 expression may be manipulated (Abcg1 KO or Abcg1 KD). Additional investigations in appropriate mouse models are therefore required in order to evaluate the overall physiopathological consequences of the reduced Abcg1 expression in diabetes.
7.2. Genetic Modulation of Human ABCG1
Similar to what was observed in diabetic mice, expression of macrophage ABCG1 was reduced in patients with T2D as compared to control subjects [110]. Indeed, blood monocyte-derived macrophages from T2D patients displayed a decreased cholesterol efflux to HDL, but not to ApoA-I, and an increased cholesterol accumulation relative to control individuals. Interestingly, treatment with a LXR agonist induced ABCG1 expression and attenuated cholesterol accumulation in macrophages from T2D patients [110]. A reduction of ABCG1 expression was also reported in peripheral blood mononuclear cells from Iranian patients with Metabolic Syndrome exhibiting high fasting glycemia in comparison to the control group [111]. Consistent with studies in murine cells, high glucose and PUFAs repressed ABCG1 expression in human macrophages [112,113] whereas a conjugated linoleic acid isomer (trans-9, trans-11-CLA) was reported to activate ABCG1 expression through sterol regulatory element-binding protein (SREBP)-1c [114]. In order to determine whether genetic variation in ABCG1 may predict T2D in the general population, Schou et al. genotyped several ABCG1 SNPs in 40,600 individuals from the CCHS [115]. However, none of the 14 ABCG1 SNPs tested was associated with an increased risk of T2D. Finally, analysis of two functional ABCG1 SNPs (rs1378577, −134 T > G and rs1893590, −204 A > C in the promoter region) in 1320 French morbidly obese patients (BMI > 40 kg/m2) failed to detect any association between the 2 SNPs and homeostatic model assessment of insulin resistance (HOMA-IR) with or without adjustment for BMI [75].
7.3. Epigenetic Modulation of Human ABCG1
Although analysis of genetic variations in ABCG1 failed to report any relationship between ABCG1 and diabetes, numerous studies highlighted a link between epigenetic modulation of ABCG1 and IR and diabetes. Hidalgo et al. first reported that DNA methylation at the ABCG1 locus was significantly associated with insulin and HOMA-IR [116]. Indeed, EWAS conducted in blood CD4+ T cells from 837 non-diabetic individuals in the GOLDN study identified that methylation at two ABCG1 CpG site (cg06500161 and cg01881899) was associated with fasting insulin and HOMA-IR. This observation was replicated and enriched in a recent EWAS performed in whole blood samples from 1440 non-diabetic individuals of the KORA F4 study [117]. In this study, the ABCG1 CpG site cg06500161 was associated with reduced ABCG1 expression and positively with fasting glucose, 2-h glucose, fasting insulin, 2-h insulin, glycated haemoglobin (HbA1c) and HOMA-IR, effects persisting after adjustment with BMI. Analysis of DNA methylation in a nested case-control study in Indian Asians and Europeans with incident T2D from the 8-year follow-up of 25,372 participants in the London life sciences prospective population (LOLIPOP) revealed that methylation at the ABCG1 locus (cg06500161) was associated with future T2D incidence [118]. Indeed, the relative risk for incident T2D per 1% increase in methylation was 1.09 (95% CI: 1.07–1.11) at the ABCG1 methylation marker. A similar risk for future T2D (OR = 1.09, 95% CI: 1.02–1.16) was reported by Dayeh et al. when DNA methylation at the ABCG1 locus cg06500161 was conducted in blood DNA from 258 non-diabetic individuals from the Botnia prospective study [80]. In those two studies, cg06500161 was positively associated with diabetes-related traits including fasting glucose, HbA1c, fasting insulin and HOMA-IR. Interestingly, DNA methylation at cg06500161 was increased in blood and adipose tissue from subjects with T2D versus non-diabetic subjects [80]. Additional investigations in 850 pedigreed Mexican-American individuals from 39 families in the San Antonio family heart study (SAFHS) confirmed that methylation in ABCG1 is a determinant of T2D in this population at high risk of developing T2D [119]. Thus, even after adjustment with BMI, cg06500161 was found significantly associated with fasting blood glucose, HOMA-IR and T2D. Further exploration was conducted in this population in order to evaluate genetic and epigenetic associations with the hypertriglyceridemic waist (HTGW) phenotype (high waist circumference (≥95 cm in men and ≥80 cm in women) combined with high serum TG concentration (≥2 mmol/L in men and ≥1.5 mmol/L in women)), as a marker of T2D and CVD [120]. Whereas GWAS detected no SNP in association with HTGW, EWAS identified that the ABCG1 CpG site (cg06500161) was associated with HTGW and T2D, the association with T2D being significantly explained in part through HTGW.
As a whole, genetic and epigenetic studies of ABCG1 in T2D and IR univocally demonstrated that genetic modulation of ABCG1 was without effect on T2D and associated traits whereas methylation marks at the ABCG1 locus were consistently associated with T2D as well as the risk to develop T2D. Indeed, methylation at ABCG1 CpG sites was positively associated with all the diabetes-related traits including fasting glucose, fasting insulin, HbA1c and HOMA-IR suggesting that epigenetic regulation of ABCG1 expression by environmental exposure is a critical event in the prevalence and incidence of diabetes.
8. ABCG1: Role in Obesity and Weight Gain
8.1. Major Findings from Studies in Mice
No difference in body weight was reported in Abcg1-deficent mice or mice expressing human ABCG1 when fed a chow or a high-cholesterol diet supplemented with or without fat [55,60,61,88]. However, genome-wide screens in drosophila and mice led to the identification of Abcg1 as a candidate for TG storage and obesity [109]. Indeed, Buchman et al. first reported that targeted disruption of Abcg1 protected mice against diet-induced obesity (DIO) by reducing body weight gain and adipose tissue mass. In addition, Abcg1−/− exhibited a markedly decrease of the adipocyte size, an elevation of total energy expenditure and a reduction of food intake as compared to wild-type littermates. The possible underlying mechanism was afterward elucidated by Frisdal et al. who provided evidence that adipocyte Abcg1 exerts a major role in adiposity and fat mass growth [75]. Indeed, Abcg1 ensured the optimal LPL-mediated TG hydrolysis in mouse adipocytes through promoting SM export which contributed to a concomitant induction of adipogenesis under the control of PPARγ and an increase of de novo TG synthesis and storage. Validation of the role of Abcg1 in adipocyte maturation was achieved by the local RNAi-silencing of Abcg1 expression in adipose tissue from mice fed a high-fat diet which led to a rapid decrease of adiposity and fat mass growth. Whereas an increased expression of adipose perilipin (Plin), Pparγ, and hormone-sensitive lipase (hsl) was observed in adipose tissue from male Abcg1−/− mice as compared to wild-type littermates upon a high-fat diet [109], the expression of those genes was markedly reduced in Abcg1-deficient adipocytes or in Abcg1 knock down adipose tissue from mice fed a high-fat diet [75]. Those contrasting results clearly highlight that the expression of Abcg1 in cells others than adipocytes contributes, likely to an opposite way, to adipogenesis and adipocyte maturation. However, expression of Abcg1 appears equally critical in controlling inflammation in adipose tissue (AT). Indeed, Abcg1-mediated cholesterol efflux to HDL in mouse adipocyte generated an anti-inflammatory response that reduced expression of pro-inflammatory cytokines and chemotactic factors [121]. Thus, inhibition of Abcg1 expression in mouse adipocytes led to a reduction of plasma cholesterol content, a disruption of lipid rafts and an inhibition of the translocation of NADPH oxidase 4 into lipid rafts. As a consequence, reactive oxygen species (ROS) generation and inflammatory response induced by palmitate was abrogated following silencing of Abcg1. Moreover, BMT from donor Abcg1−/− or Abcg1+/+ mice into recipient C57BL/6 mice fed a high-fat diet revealed that Abcg1−/− adipose tissue macrophages (ATM) from obese mice were enriched in cholesterol as compared to Abcg1+/+ ATM [122]. Analysis of ATM activation indicated that alternatively activated (M2) macrophages were more abundant in Abcg1−/− ATM and that this effect could likely result from an impaired M2 macrophage chemotaxis due to Abcg1 deficiency. However, deletion of Abcg1 in myeloid cells in this study did not improve obesity or glucose intolerance [122]. Intriguingly, whereas Abcg1 expression was consistently increased in AT from obese mice as compared to lean mice [109,123], a further elevation of Abcg1 in epididymal AT was observed in db/db mice following caloric restriction [123]. The increase of Abcg1 expression in AT following caloric restriction was thought to result from an induction of Abcg1 in ATM and more especially in F4/80 expressing cells. Thus, the effect of myeloid Abcg1 on the regulation of ATM cholesterol content and M2 abundance in AT from obese mice was exacerbated by caloric restriction [122]. Then, although Abcg1 deficiency protects mice from DIO, the exact role of Abcg1 in AT appears complex with a potential antagonist role of Abcg1 in AT, in promoting adipocyte maturation and fat mass growth on the one hand and in attenuating inflammation on the other hand. In this context, it is interesting to note that glucagon-like peptide 1 (GLP-1)-based therapy (vildagliptin and exendin-4) was able to induce the expression of Abcg1 and cholesterol efflux in mouse adipocytes [124].
8.2. Genetic Modulation of Human ABCG1
Several studies reported that human ABCG1 was expressed in adipose tissue [77,80] and similar to what observed in mice, ABCG1 expression was highly induced during adipocyte maturation [75]. No association between ABCG1 SNPs and BMI or waist-to-hip ratio was observed in individuals with a BMI < 30 kg/m2 [65,67,73]. However, genotyping of two functional ABCG1 SNPs (rs1378577, −134 T > G and rs1893590, −204 A > C in the promoter region) in 1320 French morbidly obese patients (BMI > 40 kg/m2) revealed that those two SNPs were associated with BMI after adjustment for diabetes and HOMA-IR [75]. Indeed, obese individuals homozygous for the frequent allele of each SNP (rs1378577, TT and rs1893590, AA) displayed the higher BMI. Also BMI increased in parallel with the amount of the frequent AT haplotype. In coherence with this result, obese individuals carrying the −134TT or −204AA genotype exhibited the higher fat mass index. Analysis of ABCG1 expression in adipose tissue biopsies indicated that ABCG1 mRNA levels were more elevated in adipose tissue from obese patients carrying the AT haplotype than those carrying the CG haplotype and ABCG1 mRNA levels in adipose tissue were positively correlated to adipocyte diameter. Association of ABCG1 genotype with obesity was replicated in two independent populations composed of either 595 severely obese (35 < BMI < 40 kg/m2) or 216 diabetic obese (30 < BMI < 35 kg/m2) from the diabetes atorvastatin lipid intervention (DALI) subjects [75]. This study reported for the first time that genetic modulation of ABCG1 was intimately linked to fat mass growth and obesity in humans suggesting that ABCG1 might represent a potential therapeutic target in obesity [125]. Although this study suggests that an increase of ABCG1 expression in adipose tissue was positively associated with weight gain and BMI, it however appears that weight loss in obese individuals was not accompanied by a decrease of ABCG1 expression in adipose tissue. Indeed, quantification of ABCG1 mRNA levels in subcutaneous adipose tissue from obese individuals following caloric restriction [126,127,128,129] or surgery [130] indicated that ABCG1 expression was not altered, the latter could even be increased [131].
8.3. Epigenetic Modulation of Human ABCG1
Although most of the metabolic disturbances in fat cells are normalized following gastric bypass surgery, patients in a post-obese (PO) state display adipose hyperplasia characterized by a smaller size but an increased number of fat cells as compared with never-obese (NO) women. In order to identify differentially methylated DNA sites linked to adipocyte hyperplasia in PO, genome-wide DNA methylation was analyzed in abdominal subcutaneous fat cells from women two years after gastric bypass surgery (PO, n = 16) and from never-obese women (NO, n = 14) [132]. Reduced DNA methylation at the ABCG1 CpG site cg10192877 was detected in fat cells from PO women in comparison to NO women, which was associated with increased ABCG1 mRNA levels in subcutaneous adipose tissue. Those results suggest that hyperplasia observed in PO women is associated with elevated ABCG1 expression in fat cells as compared NO women.
In order to decipher molecular mechanisms associated to obesity that contribute to obesity-related diseases, such as T2D and CVD, transcriptome and epigenome of circulating monocytes were conducted in 1264 participants from the multi-ethnic study of atherosclerosis (MESA) [133]. This study demonstrated that alterations of a cellular cholesterol metabolism network of 11 BMI-associated genes (including ABCG1) were associated with T2D and coronary artery calcium. More precisely, ABCG1 expression in the coexpressed network module was downregulated with increasing BMI whereas methylation at the ABCG1 CpG site cg06500161 was positively associated with BMI. Thus, the relationship between BMI and the expression of ABCG1 in the module was partially explained through methylation [133]. Analysis of whole-genome DNA methylation in peripheral blood leucocytes from 2097 African American adults in the atherosclerosis risk in communities (ARIC) study reported that the increased methylation at two CpG sites within the ABCG1 gene (cg06500161 and cg27243685) was positively associated with BMI [134]. Moreover, this study highlighted that methylation at cg06500161 was positively associated with waist circumference (WC). Finally, a very recent analysis carried out in a cohort of 1058 US women who had a sister with breast cancer but had not been diagnosed for the disease themselves (Sister Study) confirmed that blood DNA methylation of ABCG1 (cg06500161) was associated with BMI [135]. Although the primary endpoint of those studies was T2D, a positive association between the ABCG1 locus cg06500161 was reported with BMI in the Botnia prospective study [80], the LOLIPOP study [118] and with WC in the SAFHS [120]. Interestingly, the association of cg06500161 with obesity-related traits, such a BMI, waist-to-hip ratio and fat mass (total, android, ginoid and trunk) was only significant in the main group of individuals with BMI < 25 kg/m2 [118].
Taken together, results from studies in mice and in humans led to conflicting conclusions. Findings from Abcg1-deficient mice suggest that Abcg1 promotes diet-induced obesity through the contribution of adipocyte Abcg1 in adipogenesis and fat mass growth. The deleterious role of Abcg1 in obesity is supported by genetic modulation of ABCG1 in populations of obese individuals, although analysis of adipose ABCG1 expression following weight loss in both mice and humans appears in contradiction with the proposed model for ABCG1. On the contrary, epigenetic modulation of ABCG1 was undoubtedly positively associated with an increased BMI and BMI-related traits which would support a protective role of ABCG1 in obesity. Those apparent contrasting observations clearly indicate that the role of adipose ABCG1 is still not yet fully elucidated and that further investigations are required in order to decipher the puzzling role of ABCG1 in obesity.
9. ABCG1: Role in Non-Alcoholic Fatty Liver Diseases (NAFLD)
9.1. Major Findings from Studies in Mice
Although no study aimed to investigate the contribution of Abcg1 in non-alcoholic steatohepatitis (NASH) and more widely in fatty liver diseases (NAFLD) has been conducted, several lines of evidence indicate that Abcg1 plays an important role in lipid homeostasis in the liver. In rat liver, most of the Abcg1 expression resided in Kupffer cells (KC) (51% of total) whereas liver endothelial and parenchymal cells only accounted for 24% and 26% of total expression at the basal level. However a high-cholesterol diet highly induced Abcg1 expression in parenchymal cells but not in KC and endothelial cells which brought the Abcg1 expression in parenchymal cells up to 60% of total liver [136]. The use of Abcg1 deficient mice harboring a lacZ cassette insertion in exon 3 of the Abcg1 locus confirmed that Abcg1 was expressed at very low levels in hepatocytes under a chow diet although stimulation with a LXR agonist (TO901317) led to significant increase of Abcg1 expression [55]. Nevertheless, the feeding of Abcg1-deficient mice with a high-cholesterol and high-fat diet led to a massive accumulation of lipids (TG, cholesterol ester and phospholipids) in the liver, in both parenchymal cells and KC, indicating that the very low levels of Abcg1 mRNA in hepatocytes are critical in liver lipid homeostasis. By contrast, expression of the human ABCG1 transgene led to a reduction of both cholesterol ester and phospholipids in the liver from mice fed a high-cholesterol and high-fat diet as compared to wild-type mice [55]. In an independent study, a discrete increase in sterol biosynthetic intermediate levels (lathosterol, lanosterol and desmosterol) was observed in liver from mice expressing the human ABCG1 transgene fed a high-fat and high-cholesterol diet in comparison to wild-type animals [61]. Moreover, an increase of the hepatic expression of genes controlling cholesterol and fatty acids synthesis was observed in Abcg1−/− mice fed a chow diet with an increase of 3-hydroxy-3-methyl-glutaryl-coenzyme A reductase (Hmgcr), farnesyl pyrophosphate (Fpp), acetylCoA carboxylase (Acc), sterol-CoA desaturase (Sdc1) and Srebp1c mRNA levels in comparison to control mice. However, an opposite decrease of Srebp2, Ldlr and Hmgcr mRNA levels was equally reported in Abcg1−/− mice fed a chow or a high-cholesterol diet relative to control mice, suggesting a decreased hepatic cholesterol synthesis and uptake in Abcg1-deficient mice [62]. In the latter study, feeding with a high-cholesterol diet led to a significant more elevated biliary cholesterol secretion in Abcg1−/− mice compared with controls. Although Abcg1 was initially proposed to protect from lipid accumulation in the liver under a high-fat and high-cholesterol diet [55], Buchmann et al. unexpectedly reported that ablation of Abcg1 prevented the diet-induced lipid accumulation in liver from mice fed a high-fat diet devoid of cholesterol [109]. This opposite phenotype might result from the activation of the LXR pathway in cholesterol-enriched diets which in turn might stimulate lipogenesis and TG synthesis through SREBP-1c activation. However, the observation that liver TG were found decreased in Abcg1−/− mice fed a chow diet supplemented with a LXR agonist (TO901317) as compared to control mice [62] does not support this hypothesis and suggests that a different mechanism occurs in order to explain the diet-specific control of liver lipid homeostasis by Abcg1.
9.2. Sparse Observations in Humans
Little information is available about the role of ABCG1 in lipid homeostasis in human liver. Analysis of expression of genes involved in cholesterol homeostasis in liver biopsies from obese patients that underwent bariatric surgery (14 with NASH and 17 with hepatosteatosis) in comparison to control liver biopsies from cadaveric liver donors or resection of liver metastasis (n = 7) indicated that liver ABCG1 mRNA levels were modestly increased in patients with steatosis with or without NASH [137]. Interestingly, and in coherence with observation in Abcg1-defiencient mice fed a high-cholesterol diet [62], an increased expression of 3-hydroxy-3-methylglutaryl-CoA reductase (HMGCR) and sterol regulatory element-binding protein 2 (SREBP2) was also observed in liver from steatosis with or without NASH as compared to control biopsies [137]. However a more recent study performed in liver biopsies from 84 morbidly obese Mexican mestizo subjects who underwent bariatric surgery made the opposite observations [138]. Indeed, in this study, a reduction of liver ABCG1 protein levels was detected in patients with NASH as compared to control or steatosic livers while corresponding mRNA levels tended to be increased. Interestingly, ABCG1 protein levels were further decreased according to the different grades of fibrosis.
Thus, although ABCG1 appears to play an important role in the control of liver lipid homeostasis in mice in a diet-specific manner, such a role in humans remains to be demonstrated. Therefore, studies analyzing the genetic and epigenetic modulation of ABCG1 in populations of patients with hepatosteatosis or NASH will greatly help to evaluate the contribution of ABCG1 expression in NAFLD.
10. Conclusions
A comprehensive review of the literature brought to light the critical role of the human ABCG1 transporter in CMD. Indeed, by redistributing cholesterol between cell membranes, ABCG1 not only maintains cellular cholesterol homeostasis but is also involved in the secretion and bioavailability of key molecules (LPL and insulin) controlling glucose and lipid metabolism. Studies in both mice and in humans, demonstrate modulation of ABCG1 expression in association with a large spectrum of metabolic disorders including diabetes, insulin resistance, obesity and CAD. However the precise role of ABCG1 in those disorders is still under debate since results obtained from studies in mice do not corroborate observations in humans and vice-versa. Nevertheless, it becomes clear that the epigenetic modulation of ABCG1 is consistently associated with several metabolic disorders, including TG metabolism, diabetes-related traits, T2D risk, obesity and in a lesser degree to CAD (Table 2). Those studies highlight that methylation at the ABCG1 in response to environmental exposure might be critical in CMD. Indeed, CMD are complex diseases that result from close interactions between genetic and epigenetic mechanisms which are greatly influenced by environmental stimuli. However, further studies are needed to elucidate mechanisms underlying such regulations and to determine whether ABCG1 might constitute a therapeutic target in CMD.
Acknowledgments
INSERM and UPMC provided generous support for this study. This work was supported by the Fondation de France (WLG). LMH has received a research fellowship from the Île-de-France region, France (CORDDIM).
Author contributions
Lise M. Hardy, Eric Frisdal and Wilfried Le Goff conducted the bibliographic search and wrote the manuscript.
Conflicts of interest
The authors declare no conflict of interest.
Abbreviations
| ABC | ATP-Binding Cassette |
| ApoA-I | Apolipoprotein A-I |
| BMI | Body Mass Index |
| CAD | Coronary Artery Disease |
| CHD | Coronary Heart Disease |
| CMD | CardioMetabolic Disease |
| CpG | Cytosine Guanine dinucleotide |
| CVD | CardioVascular Diseases |
| EWAS | Epigenome-Wide Association Study |
| GWAS | Genome-Wide Association Study |
| HDL HOMA-IR | High-Density Lipoprotein HOmeostatic Model Assessment of Insulin Resistance |
| LDL | Low-Density Lipoprotein |
| LPL | Lipoprotein Lipase |
| LXR | Liver X Receptor |
| MI | Myocardial Infarction |
| SM | Sphingomyelin |
| SNP SREBP | Single Nucelotide Polymorphism Sterol Regulatory Element-Binding Protein |
| T2D | Type 2 Diabetes |
| TC | Total Cholesterol |
| TG | Triglycerides |
| WC | Waist Circumference |
References
- Neumann, J.; Rose-Sperling, D.; Hellmich, U.A. Diverse relations between ABC transporters and lipids: An overview. Biochim. Biophys. Acta-Biomembr. 2016. [Google Scholar] [CrossRef] [PubMed]
- Tarling, E.J.; de Aguiar Vallim, T.Q.; Edwards, P.A. Role of ABC transporters in lipid transport and human disease. Trends Endocrinol. Metab. 2013, 24, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, T.; Benndorf, R.A. ABC transport proteins in cardiovascular disease—A brief summary. Molecules 2017, 22, 589. [Google Scholar] [CrossRef] [PubMed]
- Berge, K.E.; Tian, H.; Graf, G.A.; Yu, L.; Grishin, N.V.; Schultz, J.; Kwiterovich, P.; Shan, B.; Barnes, R.; Hobbs, H.H. Accumulation of dietary cholesterol in sitosterolemia caused by mutations in adjacent ABC transporters. Science 2000, 290, 1771–1775. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Lu, K.; Hazard, S.; Yu, H.; Shulenin, S.; Hidaka, H.; Kojima, H.; Allikmets, R.; Sakuma, N.; Pegoraro, R.; et al. Identification of a gene, ABCG5, important in the regulation of dietary cholesterol absorption. Nat. Genet. 2001, 27, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Tzavella, E.; Hatzimichael, E.; Kostara, C.; Bairaktari, E.; Elisaf, M.; Tsimihodimos, V. Sitosterolemia: A multifaceted metabolic disorder with important clinical consequences. J. Clin. Lipidol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Brooks-Wilson, A.; Marcil, M.; Clee, S.M.; Zhang, L.H.; Roomp, K.; van Dam, M.; Yu, L.; Brewer, C.; Collins, J.A.; Molhuizen, H.O.; et al. Mutations in ABC1 in Tangier disease and familial high-density lipoprotein deficiency. Nat. Genet. 1999, 22, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Bodzioch, M.; Orsó, E.; Klucken, J.; Langmann, T.; Böttcher, A.; Diederich, W.; Drobnik, W.; Barlage, S.; Büchler, C.; Porsch-Ozcürümez, M.; et al. The gene encoding ATP-binding cassette transporter 1 is mutated in Tangier disease. Nat. Genet. 1999, 22, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Rust, S.; Rosier, M.; Funke, H.; Real, J.; Amoura, Z.; Piette, J.C.; Deleuze, J.F.; Brewer, H.B.; Duverger, N.; Denèfle, P.; et al. Tangier disease is caused by mutations in the gene encoding ATP-binding cassette transporter 1. Nat. Genet. 1999, 22, 352–355. [Google Scholar] [CrossRef] [PubMed]
- Lawn, R.M.; Wade, D.P.; Garvin, M.R.; Wang, X.; Schwartz, K.; Porter, J.G.; Seilhamer, J.J.; Vaughan, A.M.; Oram, J.F. The Tangier disease gene product ABC1 controls the cellular apolipoprotein-mediated lipid removal pathway. J. Clin. Investig. 1999, 104, R25–R31. [Google Scholar] [CrossRef] [PubMed]
- Brunham, L.R.; Singaraja, R.R.; Duong, M.; Timmins, J.M.; Fievet, C.; Bissada, N.; Kang, M.H.; Samra, A.; Fruchart, J.-C.; McManus, B.; et al. Tissue-specific roles of ABCA1 influence susceptibility to atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Frikke-Schmidt, R.; Nordestgaard, B.G.; Jensen, G.B.; Steffensen, R.; Tybjaerg-Hansen, A. Genetic variation in ABCA1 predicts ischemic heart disease in the general population. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Bochem, A.E.; van Wijk, D.F.; Holleboom, A.G.; Duivenvoorden, R.; Motazacker, M.M.; Dallinga-Thie, G.M.; de Groot, E.; Kastelein, J.J.P.; Nederveen, A.J.; Hovingh, G.K.; et al. ABCA1 mutation carriers with low high-density lipoprotein cholesterol are characterized by a larger atherosclerotic burden. Eur. Heart J. 2013, 34, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Klucken, J.; Büchler, C.; Orsó, E.; Kaminski, W.E.; Porsch-Özcürümez, M.; Liebisch, G.; Kapinsky, M.; Diederich, W.; Drobnik, W.; Dean, M.; et al. ABCG1 (ABC8), the human homolog of the Drosophila white gene, is a regulator of macrophage cholesterol and phospholipid transport. Proc. Natl. Acad. Sci. USA 2000, 97, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Lan, D.; Chen, W.; Matsuura, F.; Tall, A.R. ATP-binding cassette transporters G1 and G4 mediate cellular cholesterol efflux to high-density lipoproteins. Proc. Natl. Acad. Sci. USA 2004, 101, 9774–9779. [Google Scholar] [CrossRef] [PubMed]
- Savary, S.; Denizot, F.; Luciani, M.; Mattei, M.; Chimini, G. Molecular cloning of a mammalian ABC transporter homologous to Drosophila white gene. Mamm. Genome 1996, 7, 673–676. [Google Scholar] [CrossRef] [PubMed]
- Croop, J.M.; Tiller, G.E.; Fletcher, J.A.; Lux, M.L.; Raab, E.; Goldenson, D.; Son, D.; Arciniegas, S.; Wu, R.L. Isolation and characterization of a mammalian homolog of the Drosophila white gene. Gene 1997, 185, 77–85. [Google Scholar] [CrossRef]
- Langmann, T.; Porsch-Ozcürümez, M.; Unkelbach, U.; Klucken, J.; Schmitz, G. Genomic organization and characterization of the promoter of the human ATP-binding cassette transporter-G1 (ABCG1) gene. Biochim. Biophys. Acta 2000, 1494, 175–180. [Google Scholar] [CrossRef]
- Lorkowski, S.; Kratz, M.; Wenner, C.; Schmidt, R.; Weitkamp, B.; Fobker, M.; Reinhardt, J.; Rauterberg, J.; Galinski, E.A.; Cullen, P. Expression of the ATP-binding cassette transporter gene ABCG1 (ABC8) in Tangier disease. Biochem. Biophys. Res. Commun. 2001, 283, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, M.A.; Venkateswaran, A.; Tarr, P.T.; Xenarios, I.; Kudoh, J.; Shimizu, N.; Edwards, P.A. Characterization of the human ABCG1 gene liver X receptor activates an internal promoter that produces a novel transcript encoding an alternative form of the protein. J. Biol. Chem. 2001, 276, 39438–39447. [Google Scholar] [CrossRef] [PubMed]
- Venkateswaran, A.; Repa, J.J.; Lobaccaro, J.M.; Bronson, A.; Mangelsdorf, D.J.; Edwards, P.A. Human white/murine ABC8 mRNA levels are highly induced in lipid-loaded macrophages. A transcriptional role for specific oxysterols. J. Biol. Chem. 2000, 275, 14700–14707. [Google Scholar] [CrossRef] [PubMed]
- Sabol, S.L.; Brewer, H.B.; Santamarina-Fojo, S. The human ABCG1 gene: Identification of LXR response elements that modulate expression in macrophages and liver. J. Lipid Res. 2005, 46, 2151–2167. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, M.; Filomenko, R.; Rébé, C.; Chevriaux, A.; Varin, A.; Derangère, V.; Bessède, G.; Gambert, P.; Lagrost, L.; Masson, D. Knock-down of the oxysterol receptor LXRα impairs cholesterol efflux in human primary macrophages: Lack of compensation by LXRβ activation. Biochem. Pharmacol. 2013, 86, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Jakobsson, T.; Venteclef, N.; Toresson, G.; Damdimopoulos, A.E.; Ehrlund, A.; Lou, X.; Sanyal, S.; Steffensen, K.R.; Gustafsson, J.-A.; Treuter, E. GPS2 is required for cholesterol efflux by triggering histone demethylation, LXR recruitment, and coregulator assembly at the ABCG1 locus. Mol. Cell 2009, 34, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, H.; Kimura, Y.; Kioka, N.; Matsuo, M.; Ueda, K. ATPase activity of human ABCG1 is stimulated by cholesterol and sphingomyelin. J. Lipid Res. 2013, 54, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Cserepes, J.; Szentpétery, Z.; Seres, L.; Ozvegy-Laczka, C.; Langmann, T.; Schmitz, G.; Glavinas, H.; Klein, I.; Homolya, L.; Váradi, A.; et al. Functional expression and characterization of the human ABCG1 and ABCG4 proteins: Indications for heterodimerization. Biochem. Biophys. Res. Commun. 2004, 320, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, A.M.; Oram, J.F. ABCG1 redistributes cell cholesterol to domains removable by high density lipoprotein but not by lipid-depleted apolipoproteins. J. Biol. Chem. 2005, 280, 30150–30157. [Google Scholar] [CrossRef] [PubMed]
- Engel, T.; Bode, G.; Lueken, A.; Knop, M.; Kannenberg, F.; Nofer, J.-R.; Assmann, G.; Seedorf, U. Expression and functional characterization of ABCG1 splice variant ABCG1(666). FEBS Lett. 2006, 580, 4551–4559. [Google Scholar] [CrossRef] [PubMed]
- Gelissen, I.C.; Cartland, S.; Brown, A.J.; Sandoval, C.; Kim, M.; Dinnes, D.L.; Lee, Y.; Hsieh, V.; Gaus, K.; Kritharides, L.; et al. Expression and stability of two isoforms of ABCG1 in human vascular cells. Atherosclerosis 2010, 208, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Burns, V.; Sharpe, L.J.; Gelissen, I.C.; Brown, A.J. Species variation in ABCG1 isoform expression: Implications for the use of animal models in elucidating ABCG1 function. Atherosclerosis 2013, 226, 408–411. [Google Scholar] [CrossRef] [PubMed]
- Gelissen, I.C.; Sharpe, L.J.; Sandoval, C.; Rao, G.; Kockx, M.; Kritharides, L.; Jessup, W.; Brown, A.J. Protein kinase A modulates the activity of a major human isoform of ABCG1. J. Lipid Res. 2012, 53, 2133–2140. [Google Scholar] [CrossRef] [PubMed]
- Aleidi, S.M.; Howe, V.; Sharpe, L.J.; Yang, A.; Rao, G.; Brown, A.J.; Gelissen, I.C. The E3 ubiquitin ligases, HUWE1 and NEDD4-1, are involved in the post-translational regulation of the ABCG1 and ABCG4 lipid transporters. J. Biol. Chem. 2015, 290, 24604–24613. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, V.; Kim, M.-J.; Gelissen, I.C.; Brown, A.J.; Sandoval, C.; Hallab, J.C.; Kockx, M.; Traini, M.; Jessup, W.; Kritharides, L. Cellular cholesterol regulates ubiquitination and degradation of the cholesterol export proteins ABCA1 and ABCG1. J. Biol. Chem. 2014, 289, 7524–7536. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, L.J.; Rao, G.; Jones, P.M.; Glancey, E.; Aleidi, S.M.; George, A.M.; Brown, A.J.; Gelissen, I.C. Cholesterol sensing by the ABCG1 lipid transporter: Requirement of a CRAC motif in the final transmembrane domain. Biochim. Biophys. Acta 2015, 1851, 956–964. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Engel, T.; Schnoor, M.; Niehaus, J.; Hofnagel, O.; Buers, I.; Cullen, P.; Seedorf, U.; Assmann, G.; Lorkowski, S. Cell surface localization of ABCG1 does not require LXR activation. Arterioscler. Thromb. Vasc. Biol. 2006, 26, e143–e144. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.; Takanezawa, Y.; Hirata, T.; Shimizu, Y.; Misasa, K.; Kioka, N.; Arai, H.; Ueda, K.; Matsuo, M. Efflux of sphingomyelin, cholesterol, and phosphatidylcholine by ABCG1. J. Lipid Res. 2006, 47, 1791–1802. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Wang, F.; Alabi, A.; Deng, S.; Qin, S.; Zhang, D. Identification of an amino acid residue critical for plasma membrane localization of ATP-binding cassette transporter G1—Brief report. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 253–255. [Google Scholar] [CrossRef] [PubMed]
- Gelissen, I.C.; Harris, M.; Rye, K.-A.; Quinn, C.; Brown, A.J.; Kockx, M.; Cartland, S.; Packianathan, M.; Kritharides, L.; Jessup, W. ABCA1 and ABCG1 synergize to mediate cholesterol export to apoA-I. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Ranalletta, M.; Matsuura, F.; Peng, F.; Tall, A.R. LXR-induced redistribution of ABCG1 to plasma membrane in macrophages enhances cholesterol mass efflux to HDL. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1310–1316. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Li, G.; Gao, X.; Berthiaume, L.G.; Zhang, D. Characterization of palmitoylation of ATP binding cassette transporter G1: Effect on protein trafficking and function. Biochim. Biophys. Acta 2013, 1831, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Neufeld, E.B.; O’Brien, K.; Walts, A.D.; Stonik, J.A.; Malide, D.; Combs, C.A.; Remaley, A.T. The human ABCG1 transporter mobilizes plasma membrane and late endosomal non-sphingomyelin-associated-cholesterol for efflux and esterification. Biology 2014, 3, 866–891. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, G.; Gu, H.; Zhang, D. Characterization of the role of a highly conserved sequence in ATP binding cassette transporter G (ABCG) family in ABCG1 stability, oligomerization, and trafficking. Biochemistry 2013, 52, 9497–9509. [Google Scholar] [CrossRef] [PubMed]
- Borst, P.; Elferink, R.O. Mammalian ABC transporters in health and disease. Annu. Rev. Biochem. 2002, 71, 537–592. [Google Scholar] [CrossRef] [PubMed]
- Hegyi, Z.; Homolya, L. Functional cooperativity between ABCG4 and ABCG1 isoforms. PLoS ONE 2016, 11, e0156516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarr, P.T.; Edwards, P.A. ABCG1 and ABCG4 are coexpressed in neurons and astrocytes of the CNS and regulate cholesterol homeostasis through SREBP-2. J. Lipid Res. 2008, 49, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, A.M.; Oram, J.F. ABCA1 and ABCG1 or ABCG4 act sequentially to remove cellular cholesterol and generate cholesterol-rich HDL. J. Lipid Res. 2006, 47, 2433–2443. [Google Scholar] [CrossRef] [PubMed]
- Engel, T.; Kannenberg, F.; Fobker, M.; Nofer, J.-R.; Bode, G.; Lueken, A.; Assmann, G.; Seedorf, U. Expression of ATP binding cassette-transporter ABCG1 prevents cell death by transporting cytotoxic 7beta-hydroxycholesterol. FEBS Lett. 2007, 581, 1673–1680. [Google Scholar] [CrossRef] [PubMed]
- Terasaka, N.; Yu, S.; Yvan-Charvet, L.; Wang, N.; Mzhavia, N.; Langlois, R.; Pagler, T.; Li, R.; Welch, C.L.; Goldberg, I.J.; et al. ABCG1 and HDL protect against endothelial dysfunction in mice fed a high-cholesterol diet. J. Clin. Investig. 2008, 118, 3701–3713. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, Y.; Ma, J.; Ling, W.; Xia, M. Adenosine monophosphate activated protein kinase regulates ABCG1-mediated oxysterol efflux from endothelial cells and protects against hypercholesterolemia-induced endothelial dysfunction. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1354–1362. [Google Scholar] [CrossRef] [PubMed]
- Engel, T.; Fobker, M.; Buchmann, J.; Kannenberg, F.; Rust, S.; Nofer, J.-R.; Schürmann, A.; Seedorf, U. 3β,5α,6β-Cholestanetriol and 25-hydroxycholesterol accumulate in ATP-binding cassette transporter G1 (ABCG1)-deficiency. Atherosclerosis 2014, 235, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Olivier, M.; Bottg, R.; Frisdal, E.; Nowick, M.; Plengpanich, W.; Desmarchelier, C.; Roi, S.; Quinn, C.M.; Gelissen, I.; Jessup, W.; et al. ABCG1 is involved in vitamin E efflux. Biochim. Biophys. Acta 2014, 1841, 1741–1751. [Google Scholar] [CrossRef] [PubMed]
- Larrede, S.; Quinn, C.M.; Jessup, W.; Frisdal, E.; Olivier, M.; Hsieh, V.; Kim, M.-J.; van Eck, M.; Couvert, P.; Carrie, A.; et al. Stimulation of cholesterol efflux by LXR agonists in cholesterol-loaded human macrophages is ABCA1-dependent but ABCG1-independent. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1930–1936. [Google Scholar] [CrossRef] [PubMed]
- Tarling, E.J.; Edwards, P.A. ATP binding cassette transporter G1 (ABCG1) is an intracellular sterol transporter. Proc. Natl. Acad. Sci. USA 2011, 108, 19719–19724. [Google Scholar] [CrossRef] [PubMed]
- Sankaranarayanan, S.; Oram, J.F.; Asztalos, B.F.; Vaughan, A.M.; Lund-Katz, S.; Adorni, M.P.; Phillips, M.C.; Rothblat, G.H. Effects of acceptor composition and mechanism of ABCG1-mediated cellular free cholesterol efflux. J. Lipid Res. 2009, 50, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, M.A.; Barrera, G.C.; Nakamura, K.; Baldán, A.; Tarr, P.; Fishbein, M.C.; Frank, J.; Francone, O.L.; Edwards, P.A. ABCG1 has a critical role in mediating cholesterol efflux to HDL and preventing cellular lipid accumulation. Cell Metab. 2005, 1, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Baldán, Á.; Tarr, P.; Vales, C.S.; Frank, J.; Shimotake, T.K.; Hawgood, S.; Edwards, P.A. Deletion of the transmembrane transporter ABCG1 results in progressive pulmonary lipidosis. J. Biol. Chem. 2006, 281, 29401–29410. [Google Scholar] [CrossRef] [PubMed]
- Thomassen, M.J.; Barna, B.P.; Malur, A.G.; Bonfield, T.L.; Farver, C.F.; Malur, A.; Dalrymple, H.; Kavuru, M.S.; Febbraio, M. ABCG1 is deficient in alveolar macrophages of GM-CSF knockout mice and patients with pulmonary alveolar proteinosis. J. Lipid Res. 2007, 48, 2762–2768. [Google Scholar] [CrossRef] [PubMed]
- Malur, A.; Huizar, I.; Wells, G.; Barna, B.P.; Malur, A.G.; Thomassen, M.J. Lentivirus-ABCG1 instillation reduces lipid accumulation and improves lung compliance in GM-CSF knock-out mice. Biochem. Biophys. Res. Commun. 2011, 415, 288–293. [Google Scholar] [CrossRef] [PubMed]
- De Aguiar Vallim, T.Q.; Lee, E.; Merriott, D.J.; Goulbourne, C.N.; Cheng, J.; Cheng, A.; Gonen, A.; Allen, R.M.; Palladino, E.N. D.; Ford, D.A.; et al. ABCG1 regulates pulmonary surfactant metabolism in mice and men. J. Lipid Res. 2017, 58, 941–954. [Google Scholar] [CrossRef] [PubMed]
- Out, R.; Hoekstra, M.; Meurs, I.; de Vos, P.; Kuiper, J.; van Eck, M.; van Berkel, T.J. C. Total body ABCG1 expression protects against early atherosclerotic lesion development in mice. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Burgess, B.; Naus, K.; Chan, J.; Hirsch-Reinshagen, V.; Tansley, G.; Matzke, L.; Chan, B.; Wilkinson, A.; Fan, J.; Donkin, J.; et al. Overexpression of human ABCG1 does not affect atherosclerosis in fat-fed ApoE-deficient mice. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1731–1737. [Google Scholar] [CrossRef] [PubMed]
- Wiersma, H.; Nijstad, N.; de Boer, J.F.; Out, R.; Hogewerf, W.; van Berkel, T.J.; Kuipers, F.; Tietge, U.J. F. Lack of Abcg1 results in decreased plasma HDL cholesterol levels and increased biliary cholesterol secretion in mice fed a high cholesterol diet. Atherosclerosis 2009, 206, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Iida, A.; Saito, S.; Sekine, A.; Mishima, C.; Kitamura, Y.; Kondo, K.; Harigae, S.; Osawa, S.; Nakamura, Y. Catalog of 605 single-nucleotide polymorphisms (SNPs) among 13 genes encoding human ATP-binding cassette transporters: ABCA4, ABCA7, ABCA8, ABCD1, ABCD3, ABCD4, ABCE1, ABCF1, ABCG1, ABCG2, ABCG4, ABCG5, and ABCG8. J. Hum. Genet. 2002, 47, 285–310. [Google Scholar] [CrossRef] [PubMed]
- Furuyama, S.; Uehara, Y.; Zhang, B.; Baba, Y.; Abe, S.; Iwamoto, T.; Miura, S.-I.; Saku, K. Genotypic effect of ABCG1 gene promoter -257T>G polymorphism on coronary artery disease severity in Japanese men. J. Atheroscler. Thromb. 2009, 16, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Olivier, M.; Tanck, M.W.; Out, R.; Villard, E.F.; Lammers, B.; Bouchareychas, L.; Frisdal, E.; Superville, A.; van Berkel, T.; Kastelein, J.J.; et al. Human ATP-binding cassette G1 controls macrophage lipoprotein lipase bioavailability and promotes foam cell formation. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2223–2231. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, W.; Zhang, L.; Qi, L.-P.; Li, L.-Y.; Chen, L.-F.; Fang, Q.; Dang, A.-M.; Yan, X.-W. A polymorphism in the ABCG1 promoter is functionally associated with coronary artery disease in a Chinese Han population. Atherosclerosis 2011, 219, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wang, W.; Xu, Y.; Wang, Y.; Chen, L.-F.; Fang, Q.; Yan, X.-W. ABCG1 rs57137919G>a polymorphism is functionally associated with varying gene expression and apoptosis of macrophages. PLoS ONE 2014, 9, e97044. [Google Scholar] [CrossRef] [PubMed]
- Schou, J.; Frikke-Schmidt, R.; Kardassis, D.; Thymiakou, E.; Nordestgaard, B.G.; Jensen, G.; Grande, P.; Tybjærg-Hansen, A. Genetic variation in ABCG1 and risk of myocardial infarction and ischemic heart disease. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Edmondson, A.C.; Braund, P.S.; Stylianou, I.M.; Khera, A.V.; Nelson, C.P.; Wolfe, M.L.; Derohannessian, S.L.; Keating, B.J.; Qu, L.; He, J.; et al. Dense genotyping of candidate gene loci identifies variants associated with high-density lipoprotein cholesterol. Circ. Cardiovasc. Genet. 2011, 4, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Abellán, R.; Mansego, M.L.; Martínez-Hervás, S.; Martín-Escudero, J.C.; Carmena, R.; Real, J.T.; Redon, J.; Castrodeza-Sanz, J.J.; Chaves, F.J. Association of selected ABC gene family single nucleotide polymorphisms with postprandial lipoproteins: Results from the population-based Hortega study. Atherosclerosis 2010, 211, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Ko, D.T.; Alter, D.A.; Guo, H.; Koh, M.; Lau, G.; Austin, P.C.; Booth, G.L.; Hogg, W.; Jackevicius, C.A.; Lee, D.S.; et al. High-density lipoprotein cholesterol and cause-specific mortality in individuals without previous cardiovascular conditions: The CANHEART study. J. Am. Coll. Cardiol. 2016, 68, 2073–2083. [Google Scholar] [CrossRef] [PubMed]
- Abellán, R.; Mansego, M.L.; Martínez-Hervás, S.; Morcillo, S.; Pineda-Alonso, M.; Carmena, R.; Real, J.T.; Redon, J.; Rojo-Martínez, G.; Martín-Escudero, J.C.; et al. Dietary polyunsaturated fatty acids may increase plasma LDL-cholesterol and plasma cholesterol concentrations in carriers of an ABCG1 gene single nucleotide polymorphism: Study in two Spanish populations. Atherosclerosis 2011, 219, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Zago, V.H. S.; Scherrer, D.Z.; Parra, E.S.; Panzoldo, N.B.; Alexandre, F.; Nakandakare, E.R.; Quintão, E.C.R.; de Faria, E.C. Association between ABCG1 polymorphism rs1893590 and high-density lipoprotein (HDL) in an asymptomatic Brazilian population. Mol. Biol. Rep. 2015, 42, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Asselbergs, F.W.; Guo, Y.; van Iperen, E.P. A.; Sivapalaratnam, S.; Tragante, V.; Lanktree, M.B.; Lange, L.A.; Almoguera, B.; Appelman, Y.E.; Barnard, J.; et al. Large-scale gene-centric meta-analysis across 32 studies identifies multiple lipid loci. Am. J. Hum. Genet. 2012, 91, 823–838. [Google Scholar] [CrossRef] [PubMed]
- Frisdal, E.; Le Lay, S.; Hooton, H.; Poupel, L.; Olivier, M.; Alili, R.; Plengpanich, W.; Villard, E.F.; Gilibert, S.; Lhomme, M.; et al. Adipocyte ATP-binding cassette G1 promotes triglyceride storage, fat mass growth, and human obesity. Diabetes 2015, 64, 840–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, K.V.E.; Voortman, T.; Dhana, K.; Troup, J.; Bramer, W.M.; Troup, J.; Chowdhury, R.; Dehghan, A.; Muka, T.; Franco, O.H. The role of DNA methylation in dyslipidaemia: A systematic review. Prog. Lipid Res. 2016, 64, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, L.; Wahl, S.; Pilling, L.C.; Reischl, E.; Sandling, J.K.; Kunze, S.; Holdt, L.M.; Kretschmer, A.; Schramm, K.; Adamski, J.; et al. DNA methylation of lipid-related genes affects blood lipid levels. Circ. Cardiovasc. Genet. 2015, 8, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Braun, K.V.E.; Dhana, K.; de Vries, P.S.; Voortman, T.; van Meurs, J.B.J.; Uitterlinden, A.G.; BIOS consortium; Hofman, A.; Hu, F.B.; Franco, O.H.; et al. Epigenome-wide association study (EWAS) on lipids: The Rotterdam study. Clin. Epigenet. 2017, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Hedman, Å.K.; Mendelson, M.M.; Marioni, R.E.; Gustafsson, S.; Joehanes, R.; Irvin, M.R.; Zhi, D.; Sandling, J.K.; Yao, C.; Liu, C.; et al. Epigenetic patterns in blood associated with lipid traits predict incident coronary heart disease events and are enriched for results from genome-wide association studies. Circ. Cardiovasc. Genet. 2017, 10, e001487. [Google Scholar] [CrossRef] [PubMed]
- Dayeh, T.; Tuomi, T.; Almgren, P.; Perfilyev, A.; Jansson, P.-A.; de Mello, V.D.; Pihlajamäki, J.; Vaag, A.; Groop, L.; Nilsson, E.; et al. DNA methylation of loci within ABCG1 and PHOSPHO1 in blood DNA is associated with future type 2 diabetes risk. Epigenetics 2016, 11, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Guay, S.-P.; Brisson, D.; Lamarche, B.; Gaudet, D.; Bouchard, L. Epipolymorphisms within lipoprotein genes contribute independently to plasma lipid levels in familial hypercholesterolemia. Epigenetics 2014, 9, 718–729. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.-Q.; Wojczynski, M.K.; Parnell, L.D.; Hidalgo, B.A.; Irvin, M.R.; Aslibekyan, S.; Province, M.A.; Absher, D.M.; Arnett, D.K.; Ordovás, J.M. Epigenome-wide association study of triglyceride postprandial responses to a high-fat dietary challenge. J. Lipid Res. 2016, 57, 2200–2207. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.-K.; Zeilinger, S.; Kastenmüller, G.; Römisch-Margl, W.; Brugger, M.; Peters, A.; Meisinger, C.; Strauch, K.; Hengstenberg, C.; Pagel, P.; et al. Epigenetics meets metabolomics: An epigenome-wide association study with blood serum metabolic traits. Hum. Mol. Genet. 2014, 23, 534–545. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, K.F.; van Iterson, M.; Slieker, R.C.; Moed, M.H.; Bonder, M.J.; van Galen, M.; Mei, H.; Zhernakova, D.V.; van den Berg, L.H.; Deelen, J.; et al. Blood lipids influence DNA methylation in circulating cells. Genome Biol. 2016, 17, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terasaka, N.; Wang, N.; Yvan-Charvet, L.; Tall, A.R. High-density lipoprotein protects macrophages from oxidized low-density lipoprotein-induced apoptosis by promoting efflux of 7-ketocholesterol via ABCG1. Proc. Natl. Acad. Sci. USA 2007, 104, 15093–15098. [Google Scholar] [CrossRef] [PubMed]
- Whetzel, A.M.; Sturek, J.M.; Nagelin, M.H.; Bolick, D.T.; Gebre, A.K.; Parks, J.S.; Bruce, A.C.; Skaflen, M.D.; Hedrick, C.C. ABCG1 deficiency in mice promotes endothelial activation and monocyte-endothelial interactions. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Münch, G.; Bültmann, A.; Li, Z.; Holthoff, H.-P.; Ullrich, J.; Wagner, S.; Ungerer, M. Overexpression of ABCG1 protein attenuates arteriosclerosis and endothelial dysfunction in atherosclerotic rabbits. Heart Int. 2012, 7, e12. [Google Scholar] [CrossRef] [PubMed]
- Basso, F.; Amar, M.J.; Wagner, E.M.; Vaisman, B.; Paigen, B.; Santamarina-Fojo, S.; Remaley, A.T. Enhanced ABCG1 expression increases atherosclerosis in LDLr-KO mice on a western diet. Biochem. Biophys. Res. Commun. 2006, 351, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Baldán, A.; Pei, L.; Lee, R.; Tarr, P.; Tangirala, R.K.; Weinstein, M.M.; Frank, J.; Li, A.C.; Tontonoz, P.; Edwards, P.A. Impaired development of atherosclerosis in hyperlipidemic Ldlr−/− and ApoE−/− mice transplanted with Abcg1−/− bone marrow. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 2301–2307. [Google Scholar] [CrossRef] [PubMed]
- Ranalletta, M.; Wang, N.; Han, S.; Yvan-Charvet, L.; Welch, C.; Tall, A.R. Decreased atherosclerosis in low-density lipoprotein receptor knockout mice transplanted with Abcg1−/− bone marrow. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 2308–2315. [Google Scholar] [CrossRef] [PubMed]
- Out, R.; Hoekstra, M.; Hildebrand, R.B.; Kruit, J.K.; Meurs, I.; Li, Z.; Kuipers, F.; van Berkel, T.J.C.; van Eck, M. Macrophage ABCG1 deletion disrupts lipid homeostasis in alveolar macrophages and moderately influences atherosclerotic lesion development in LDL receptor-deficient mice. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 2295–2300. [Google Scholar] [CrossRef] [PubMed]
- Yvan-Charvet, L.; Ranalletta, M.; Wang, N.; Han, S.; Terasaka, N.; Li, R.; Welch, C.; Tall, A.R. Combined deficiency of ABCA1 and ABCG1 promotes foam cell accumulation and accelerates atherosclerosis in mice. J. Clin. Investig. 2007, 117, 3900–3908. [Google Scholar] [CrossRef] [PubMed]
- Out, R.; Hoekstra, M.; Habets, K.; Meurs, I.; de Waard, V.; Hildebrand, R.B.; Wang, Y.; Chimini, G.; Kuiper, J.; van Berkel, T.J.C.; et al. Combined deletion of macrophage ABCA1 and ABCG1 leads to massive lipid accumulation in tissue macrophages and distinct atherosclerosis at relatively low plasma cholesterol levels. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Tarling, E.J.; Bojanic, D.D.; Tangirala, R.K.; Wang, X.; Lovgren-Sandblom, A.; Lusis, A.J.; Bjorkhem, I.; Edwards, P.A. Impaired development of atherosclerosis in Abcg1−/− Apoe−/− mice: Identification of specific oxysterols that both accumulate in Abcg1−/− Apoe−/− tissues and induce apoptosis. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1174–1180. [Google Scholar] [CrossRef] [PubMed]
- Meurs, I.; Lammers, B.; Zhao, Y.; Out, R.; Hildebrand, R.B.; Hoekstra, M.; van Berkel, T.J.C.; van Eck, M. The effect of ABCG1 deficiency on atherosclerotic lesion development in LDL receptor knockout mice depends on the stage of atherogenesis. Atherosclerosis 2012, 221, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Out, R.; Jessup, W.; Le Goff, W.; Hoekstra, M.; Gelissen, I.C.; Zhao, Y.; Kritharides, L.; Chimini, G.; Kuiper, J.; Chapman, M.J.; et al. Coexistence of foam cells and hypocholesterolemia in mice lacking the ABC transporters A1 and G1. Circ. Res. 2008, 102, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Sivapalaratnam, S.; Basart, H.; Watkins, N.A.; Maiwald, S.; Rendon, A.; Krishnan, U.; Sondermeijer, B.M.; Creemers, E.E.; Pinto-Sietsma, S.J.; Hovingh, K.; et al. Monocyte gene expression signature of patients with early onset coronary artery disease. PLoS ONE 2012, 7, e32166. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Liu, X.; Zhu, R.; He, Z. Association of ATP-binding cassette transporter G1 polymorphisms with risk of ischemic stroke in the Chinese Han population. J. Stroke Cerebrovasc. Dis. 2015, 24, 1397–1404. [Google Scholar] [CrossRef] [PubMed]
- Peng, P.; Wang, L.; Yang, X.; Huang, X.; Ba, Y.; Chen, X.; Guo, J.; Lian, J.; Zhou, J. A preliminary study of the relationship between promoter methylation of the ABCG1, GALNT2 and HMGCR genes and coronary heart disease. PLoS ONE 2014, 9, e102265. [Google Scholar] [CrossRef] [PubMed]
- Le Goff, W.; Dallinga-Thie, G.M. ABCG1: Not as good as expected? Atherosclerosis 2011, 219, 393–394. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Zhou, H.; Tan, K.C.B.; Guo, R.; Shiu, S.W.M.; Wong, Y. ABCG1 mediated oxidized LDL-derived oxysterol efflux from macrophages. Biochem. Biophys. Res. Commun. 2009, 390, 1349–1354. [Google Scholar] [CrossRef] [PubMed]
- Terasaka, N.; Westerterp, M.; Koetsveld, J.; Fernández-Hernando, C.; Yvan-Charvet, L.; Wang, N.; Sessa, W.C.; Tall, A.R. ATP-binding cassette transporter G1 and high-density lipoprotein promote endothelial NO synthesis through a decrease in the interaction of caveolin-1 and endothelial NO synthase. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2219–2225. [Google Scholar] [CrossRef] [PubMed]
- Mauldin, J.P.; Srinivasan, S.; Mulya, A.; Gebre, A.; Parks, J.S.; Daugherty, A.; Hedrick, C.C. Reduction in ABCG1 in Type 2 diabetic mice increases macrophage foam cell formation. J. Biol. Chem. 2006, 281, 21216–21224. [Google Scholar] [CrossRef] [PubMed]
- Daffu, G.; Shen, X.; Senatus, L.; Thiagarajan, D.; Abedini, A.; Hurtado Del Pozo, C.; Rosario, R.; Song, F.; Friedman, R.A.; Ramasamy, R.; et al. RAGE suppresses ABCG1-mediated macrophage cholesterol efflux in diabetes. Diabetes 2015, 64, 4046–4060. [Google Scholar] [CrossRef] [PubMed]
- Nagelin, M.H.; Srinivasan, S.; Lee, J.; Nadler, J.L.; Hedrick, C.C. 12/15-Lipoxygenase activity increases the degradation of macrophage ATP-binding cassette transporter G1. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1811–1819. [Google Scholar] [CrossRef] [PubMed]
- Nagelin, M.H.; Srinivasan, S.; Nadler, J.L.; Hedrick, C.C. Murine 12/15-lipoxygenase regulates ATP-binding cassette transporter G1 protein degradation through p38- and JNK2-dependent pathways. J. Biol. Chem. 2009, 284, 31303–31314. [Google Scholar] [CrossRef] [PubMed]
- Sturek, J.M.; Castle, J.D.; Trace, A.P.; Page, L.C.; Castle, A.M.; Evans-Molina, C.; Parks, J.S.; Mirmira, R.G.; Hedrick, C.C. An intracellular role for ABCG1-mediated cholesterol transport in the regulated secretory pathway of mouse pancreatic beta cells. J. Clin. Investig. 2010, 120, 2575–2589. [Google Scholar] [CrossRef] [PubMed]
- Kruit, J.K.; Wijesekara, N.; Westwell-Roper, C.; Vanmierlo, T.; de Haan, W.; Bhattacharjee, A.; Tang, R.; Wellington, C.L.; LütJohann, D.; Johnson, J.D.; et al. Loss of both ABCA1 and ABCG1 results in increased disturbances in islet sterol homeostasis, inflammation, and impaired β-cell function. Diabetes 2012, 61, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Buchmann, J.; Meyer, C.; Neschen, S.; Augustin, R.; Schmolz, K.; Kluge, R.; Al-Hasani, H.; Jürgens, H.; Eulenberg, K.; Wehr, R.; et al. Ablation of the cholesterol transporter adenosine triphosphate-binding cassette transporter G1 reduces adipose cell size and protects against diet-induced obesity. Endocrinology 2007, 148, 1561–1573. [Google Scholar] [CrossRef] [PubMed]
- Mauldin, J.P.; Nagelin, M.H.; Wojcik, A.J.; Srinivasan, S.; Skaflen, M.D.; Ayers, C.R.; McNamara, C.A.; Hedrick, C.C. Reduced expression of ATP-binding cassette transporter G1 increases cholesterol accumulation in macrophages of patients with type 2 diabetes mellitus. Circulation 2008, 117, 2785–2792. [Google Scholar] [CrossRef] [PubMed]
- Tavoosi, Z.; Moradi-Sardareh, H.; Saidijam, M.; Yadegarazari, R.; Borzuei, S.; Soltanian, A.; Goodarzi, M.T. Cholesterol transporters ABCA1 and ABCG1 gene expression in peripheral blood mononuclear cells in patients with metabolic syndrome. Cholesterol 2015, 2015, 682904. [Google Scholar] [CrossRef] [PubMed]
- Uehara, Y.; Miura, S.; von Eckardstein, A.; Abe, S.; Fujii, A.; Matsuo, Y.; Rust, S.; Lorkowski, S.; Assmann, G.; Yamada, T.; et al. Unsaturated fatty acids suppress the expression of the ATP-binding cassette transporter G1 (ABCG1) and ABCA1 genes via an LXR/RXR responsive element. Atherosclerosis 2007, 191, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Mauerer, R.; Ebert, S.; Langmann, T. High glucose, unsaturated and saturated fatty acids differentially regulate expression of ATP-binding cassette transporters ABCA1 and ABCG1 in human macrophages. Exp. Mol. Med. 2009, 41, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Ecker, J.; Langmann, T.; Moehle, C.; Schmitz, G. Isomer specific effects of conjugated linoleic acid on macrophage ABCG1 transcription by a SREBP-1c dependent mechanism. Biochem. Biophys. Res. Commun. 2007, 352, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Schou, J.; Tybjærg-Hansen, A.; Møller, H.J.; Nordestgaard, B.G.; Frikke-Schmidt, R. ABC transporter genes and risk of type 2 diabetes: A study of 40,000 individuals from the general population. Diabetes Care 2012, 35, 2600–2606. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, B.; Irvin, M.R.; Sha, J.; Zhi, D.; Aslibekyan, S.; Absher, D.; Tiwari, H.K.; Kabagambe, E.K.; Ordovas, J.M.; Arnett, D.K. Epigenome-wide association study of fasting measures of glucose, insulin, and HOMA-IR in the genetics of lipid lowering drugs and diet network study. Diabetes 2014, 63, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Kriebel, J.; Herder, C.; Rathmann, W.; Wahl, S.; Kunze, S.; Molnos, S.; Volkova, N.; Schramm, K.; Carstensen-Kirberg, M.; Waldenberger, M.; et al. Association between DNA methylation in whole blood and measures of glucose metabolism: KORA F4 study. PLoS ONE 2016, 11, e0152314. [Google Scholar] [CrossRef] [PubMed]
- Chambers, J.C.; Loh, M.; Lehne, B.; Drong, A.; Kriebel, J.; Motta, V.; Wahl, S.; Elliott, H.R.; Rota, F.; Scott, W.R.; et al. Epigenome-wide association of DNA methylation markers in peripheral blood from Indian Asians and Europeans with incident type 2 diabetes: A nested case-control study. Lancet Diabetes Endocrinol. 2015, 3, 526–534. [Google Scholar] [CrossRef]
- Kulkarni, H.; Kos, M.Z.; Neary, J.; Dyer, T.D.; Kent, J.W.; Göring, H.H.H.; Cole, S.A.; Comuzzie, A.G.; Almasy, L.; Mahaney, M.C.; et al. Novel epigenetic determinants of type 2 diabetes in Mexican-American families. Hum. Mol. Genet. 2015, 24, 5330–5344. [Google Scholar] [CrossRef] [PubMed]
- Mamtani, M.; Kulkarni, H.; Dyer, T.D.; Göring, H.H.H.; Neary, J.L.; Cole, S.A.; Kent, J.W.; Kumar, S.; Glahn, D.C.; Mahaney, M.C.; et al. Genome- and epigenome-wide association study of hypertriglyceridemic waist in Mexican American families. Clin. Epigenet. 2016, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Umemoto, T.; Han, C.Y.; Mitra, P.; Averill, M.M.; Tang, C.; Goodspeed, L.; Omer, M.; Subramanian, S.; Wang, S.; Den Hartigh, L.J.; et al. Apolipoprotein AI and high-density lipoprotein have anti-inflammatory effects on adipocytes via cholesterol transporters: ATP-binding cassette A-1, ATP-binding cassette G-1, and scavenger receptor B-1. Circ. Res. 2013, 112, 1345–1354. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Tarling, E.J.; McMillen, T.S.; Tang, C.; LeBoeuf, R.C. ABCG1 regulates mouse adipose tissue macrophage cholesterol levels and ratio of M1 to M2 cells in obesity and caloric restriction. J. Lipid Res. 2015, 56, 2337–2347. [Google Scholar] [CrossRef] [PubMed]
- Edgel, K.A.; McMillen, T.S.; Wei, H.; Pamir, N.; Houston, B.A.; Caldwell, M.T.; Mai, P.-O.T.; Oram, J.F.; Tang, C.; Leboeuf, R.C. Obesity and weight loss result in increased adipose tissue ABCG1 expression in db/db mice. Biochim. Biophys. Acta 2012, 1821, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, A.M.; Hamdy, N.M.; El-Mesallamy, H.O.; Abdel-Rahman, S.Z. Glucagon-like peptide 1 (GLP-1)-based therapy upregulates LXR-ABCA1/ABCG1 cascade in adipocytes. Biochem. Biophys. Res. Commun. 2015, 468, 900–905. [Google Scholar] [CrossRef] [PubMed]
- Frisdal, E.; Le Goff, W. Adipose ABCG1: A potential therapeutic target in obesity? Adipocyte 2015, 4, 315–318. [Google Scholar] [CrossRef] [PubMed]
- Campbell, K.L.; Foster-Schubert, K.E.; Makar, K.W.; Kratz, M.; Hagman, D.; Schur, E.A.; Habermann, N.; Horton, M.; Abbenhardt, C.; Kuan, L.-Y.; et al. Gene expression changes in adipose tissue with diet- and/or exercise-induced weight loss. Cancer Prev. Res. 2013, 6, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Nookaew, I.; Svensson, P.-A.; Jacobson, P.; Jernås, M.; Taube, M.; Larsson, I.; Andersson-Assarsson, J.C.; Sjöström, L.; Froguel, P.; Walley, A.; et al. Adipose tissue resting energy expenditure and expression of genes involved in mitochondrial function are higher in women than in men. J. Clin. Endocrinol. Metab. 2013, 98, E370–E378. [Google Scholar] [CrossRef] [PubMed]
- Rizkalla, S.W.; Prifti, E.; Cotillard, A.; Pelloux, V.; Rouault, C.; Allouche, R.; Laromiguière, M.; Kong, L.; Darakhshan, F.; Massiera, F.; et al. Differential effects of macronutrient content in 2 energy-restricted diets on cardiovascular risk factors and adipose tissue cell size in moderately obese individuals: A randomized controlled trial. Am. J. Clin. Nutr. 2012, 95, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Mutch, D.M.; Pers, T.H.; Temanni, M.R.; Pelloux, V.; Marquez-Quiñones, A.; Holst, C.; Martinez, J.A.; Babalis, D.; van Baak, M.A.; Handjieva-Darlenska, T.; et al. DiOGenes Project A distinct adipose tissue gene expression response to caloric restriction predicts 6-mo weight maintenance in obese subjects. Am. J. Clin. Nutr. 2011, 94, 1399–1409. [Google Scholar] [CrossRef] [PubMed]
- Hoggard, N.; Cruickshank, M.; Moar, K.-M.; Bashir, S.; Mayer, C.-D. Using gene expression to predict differences in the secretome of human omental vs. subcutaneous adipose tissue. Obesity 2012, 20, 1158–1167. [Google Scholar] [CrossRef] [PubMed]
- Johansson, L.E.; Danielsson, A.P.H.; Parikh, H.; Klintenberg, M.; Norström, F.; Groop, L.; Ridderstråle, M. Differential gene expression in adipose tissue from obese human subjects during weight loss and weight maintenance. Am. J. Clin. Nutr. 2012, 96, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Dahlman, I.; Sinha, I.; Gao, H.; Brodin, D.; Thorell, A.; Rydén, M.; Andersson, D.P.; Henriksson, J.; Perfilyev, A.; Ling, C.; et al. The fat cell epigenetic signature in post-obese women is characterized by global hypomethylation and differential DNA methylation of adipogenesis genes. Int. J. Obes. 2015, 39, 910–919. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Reynolds, L.M.; Zeller, T.; Müller, C.; Lohman, K.; Nicklas, B.J.; Kritchevsky, S.B.; Huang, Z.; de la Fuente, A.; Soranzo, N.; et al. Alterations of a cellular cholesterol metabolism network are a molecular feature of obesity-related type 2 diabetes and cardiovascular disease. Diabetes 2015, 64, 3464–3474. [Google Scholar] [CrossRef] [PubMed]
- Demerath, E.W.; Guan, W.; Grove, M.L.; Aslibekyan, S.; Mendelson, M.; Zhou, Y.-H.; Hedman, Å.K.; Sandling, J.K.; Li, L.-A.; Irvin, M.R.; et al. Epigenome-wide association study (EWAS) of BMI, BMI change and waist circumference in African American adults identifies multiple replicated loci. Hum. Mol. Genet. 2015, 24, 4464–4479. [Google Scholar] [CrossRef] [PubMed]
- Wilson, L.E.; Harlid, S.; Xu, Z.; Sandler, D.P.; Taylor, J.A. An epigenome-wide study of body mass index and DNA methylation in blood using participants from the Sister Study cohort. Int. J. Obes. 2017, 41, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Hoekstra, M.; Kruijt, J.K.; van Eck, M.; van Berkel, T.J.C. Specific gene expression of ATP-binding cassette transporters and nuclear hormone receptors in rat liver parenchymal, endothelial, and Kupffer cells. J. Biol. Chem. 2003, 278, 25448–25453. [Google Scholar] [CrossRef] [PubMed]
- Caballero, F.; Fernández, A.; de Lacy, A.M.; Fernández-Checa, J.C.; Caballería, J.; García-Ruiz, C. Enhanced free cholesterol, SREBP-2 and StAR expression in human NASH. J. Hepatol. 2009, 50, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Vega-Badillo, J.; Gutiérrez-Vidal, R.; Hernández-Pérez, H.A.; Villamil-Ramírez, H.; León-Mimila, P.; Sánchez-Muñoz, F.; Morán-Ramos, S.; Larrieta-Carrasco, E.; Fernández-Silva, I.; Méndez-Sánchez, N.; et al. Hepatic miR-33a/miR-144 and their target gene ABCA1 are associated with steatohepatitis in morbidly obese subjects. Liver Int. 2016, 36, 1383–1391. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structure of the membrane ATP-binding cassette G1 (ABCG1) transporter. ABCG1 is a member of the ABC superfamily of transporters and contains one ABC domain containing three highly conserved domains (Walker A (A), Walker B (B) and Signature motif (S)) and one transmembrane domain (TMD) consisting of six membrane-spanning α-helices. The 12 amino acids present in the long ABCG1 isoform (+12AA) are indicated. Functional ABC transporters require two ABCs and two TMDs, therefore, the ABCG1 half-transporter must homodimerize or heterodimerize with others ABC partners in order to be functional. Nt: N-terminus; Ct: C-terminus.
Figure 1.
Structure of the membrane ATP-binding cassette G1 (ABCG1) transporter. ABCG1 is a member of the ABC superfamily of transporters and contains one ABC domain containing three highly conserved domains (Walker A (A), Walker B (B) and Signature motif (S)) and one transmembrane domain (TMD) consisting of six membrane-spanning α-helices. The 12 amino acids present in the long ABCG1 isoform (+12AA) are indicated. Functional ABC transporters require two ABCs and two TMDs, therefore, the ABCG1 half-transporter must homodimerize or heterodimerize with others ABC partners in order to be functional. Nt: N-terminus; Ct: C-terminus.


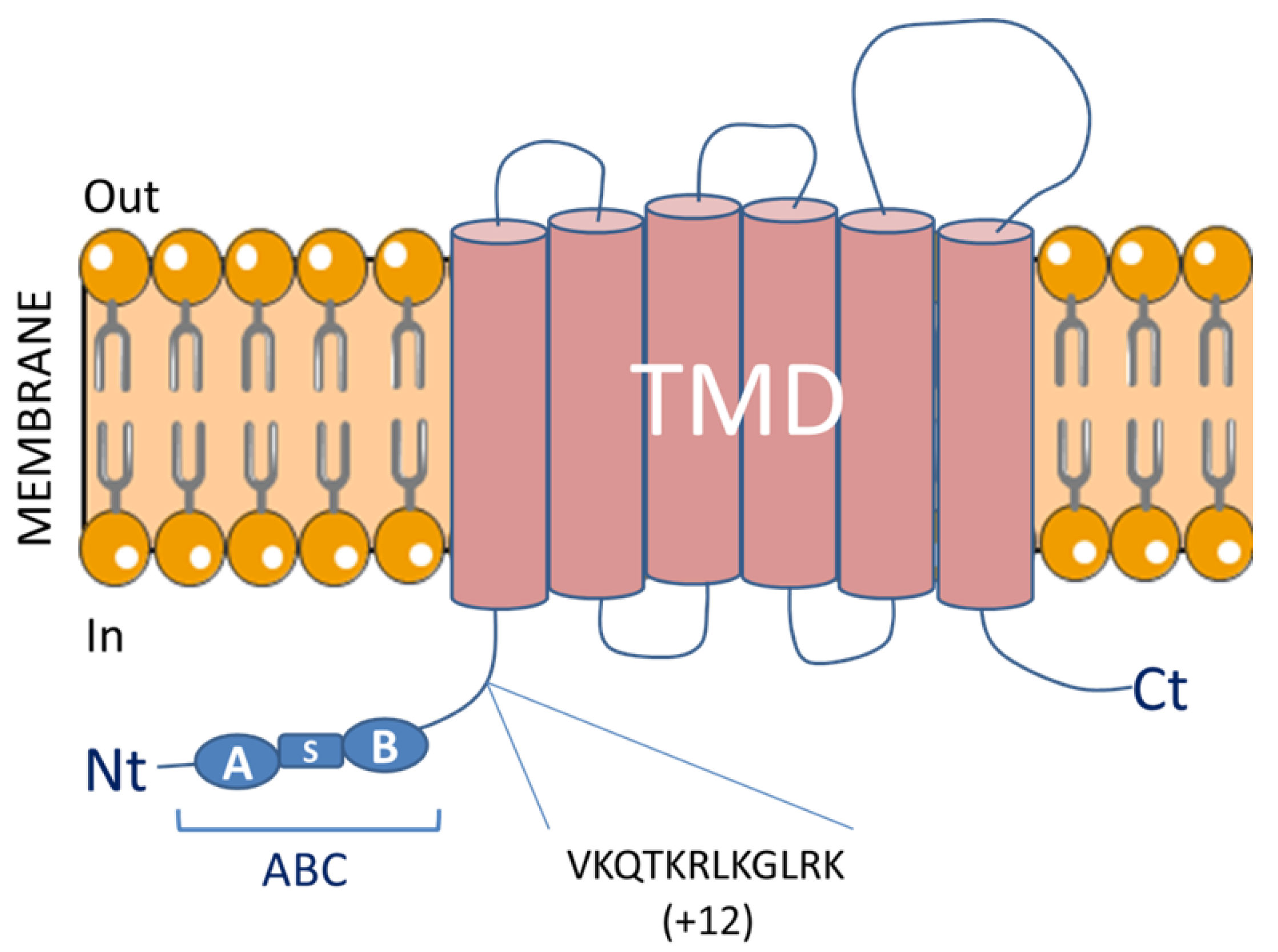{kind=link}
Table 1.
Association between methylation at different ATP-binding cassette G1 (ABCG1) CpG loci and plasma lipid levels. HDL, high-density lipoprotein; TG, triglycerides; KORA, cooperative health research in the region of Augsburg; InCHIANTI, aging in the chianti area; FHS, Framingham heart study; PIVUS, prospective investigation of the vasculature in Uppsala seniors; PPHT; postprandial hypertriglyceridemia; GOLDN, genetics of lipid lowering drugs and diet network.
Table 1.
Association between methylation at different ATP-binding cassette G1 (ABCG1) CpG loci and plasma lipid levels. HDL, high-density lipoprotein; TG, triglycerides; KORA, cooperative health research in the region of Augsburg; InCHIANTI, aging in the chianti area; FHS, Framingham heart study; PIVUS, prospective investigation of the vasculature in Uppsala seniors; PPHT; postprandial hypertriglyceridemia; GOLDN, genetics of lipid lowering drugs and diet network.
| Lipid Trait | CpG Site | Position | Direction | Cohort | n | p Value | Reference |
|---|---|---|---|---|---|---|---|
| HDL | cg06500161 | 43,656,588 | − | KORA F3, KORA F4 and InCHIANTI (meta-analysis) | 2747 | 9.00 × 10−11 | [77] |
| − | Rotterdam discovery/replication | 725/760 | 9.5 × 10−23 | [78] | |||
| − | FHS and PIVUS | 2306 | 1.2 × 10−34 | [79] | |||
| TG | cg06500161 | 43,656,588 | + | KORA F3, KORA F4 and InCHIANTI (meta-analysis) | 2747 | 5.56 × 10−10 | [77] |
| + | Botnia prospective study | 258 | 0.001 | [80] | |||
| + | Rotterdam discovery/replication | 725/760 | 1.4 × 10−24 | [78] | |||
| + | FHS and PIVUS | 2306 | 2.29 × 10−48 | [79] | |||
| +(PPHT) | GOLDN | 979 | 4.25 × 10−9 | [82] | |||
| cg27243685 | 43,642,366 | + | KORA F3, KORA F4 and InCHIANTI (meta-analysis) | 2747 | 2.49 × 10−5 | [77] | |
| + | FHS and PIVUS | 2306 | 8.12 × 10−26 | [79] | |||
| cg07397296 | 42,235,165 | + | KORA F3, KORA F4 and InCHIANTI (meta-analysis) | 2747 | 3.78 × 10−3 | [77] | |
| cg01176028 | 43,653,234 | + | FHS and PIVUS | 2306 | 5 × 10−9 | [79] | |
| CpGC3 | Not specified | + | Canadian familial Hypercholesterolemia | 98 | 0.02 | [81] |
Table 2.
Relation between methylation at different ABCG1 CpG loci and cardiovascular diseases, prevalence and incidence of type 2 diabete and obesity. CHD, coronary heart diseases; T2D, type 2 diabetes; MI, myocardial infarction; KORA, cooperative health research in the region of Augsburg; InCHIANTI, aging in the Chianti area; HOMA-IR, homeostatic model assessment of insulin resistance; GOLDN, genetic of lipid lowering drugs and diet network; HbA1c, glycated haemoglobin; BMI, body mass index; LOLIPOP, London life science prospective population; HTGW, hypertriglyceridemic waist; WC, waist circumference; SAFHS, San Antonio family heart study; MESA, multi-ethnic study of atherosclerosis; ARIC, atherosclerosis risk in communities; FHS, Framingham heart study; PIVUS, prospective investigation of the vasculature in Uppsala seniors.
Table 2.
Relation between methylation at different ABCG1 CpG loci and cardiovascular diseases, prevalence and incidence of type 2 diabete and obesity. CHD, coronary heart diseases; T2D, type 2 diabetes; MI, myocardial infarction; KORA, cooperative health research in the region of Augsburg; InCHIANTI, aging in the Chianti area; HOMA-IR, homeostatic model assessment of insulin resistance; GOLDN, genetic of lipid lowering drugs and diet network; HbA1c, glycated haemoglobin; BMI, body mass index; LOLIPOP, London life science prospective population; HTGW, hypertriglyceridemic waist; WC, waist circumference; SAFHS, San Antonio family heart study; MESA, multi-ethnic study of atherosclerosis; ARIC, atherosclerosis risk in communities; FHS, Framingham heart study; PIVUS, prospective investigation of the vasculature in Uppsala seniors.
| CpG Site | Position | CVD | Type 2 Diabete | Obesity | Direction | Cohort | n | Reference |
|---|---|---|---|---|---|---|---|---|
| CpGC3 | Not specified | CAD | ↔ | Canadian Familial Hypercholesterolemia | 22/22 | [81] | ||
| ABCG1 promoter | Not specified | CHD | + | Chinese Han population | 139 | [99] | ||
| cg06500161 | 43,656,588 | MI | + | KORA F3, KORA F4 and InCHIANTI (meta-analysis) | 2747 | [77] | ||
| Fasting insulin and HOMA-IR | + | GOLDN | 837 | [116] | ||||
| Fasting glucose, fasting insulin, HbA1c and HOMA-IR | + | KORA F4 | 1440 | [117] | ||||
| Risk of future T2D (fasting glucose, HbA1c, fasting insulin and HOMA-IR) | BMI and WHR, Fat mass | + | LOLIPOP | 25,372 | [118] | |||
| Risk of future T2D (fasting glucose, HbA1c, fasting insulin and HOMA-IR) | BMI | + | Non-diabetic from Botnia prospective study | 258 | [80] | |||
| Risk of future T2D (fasting glucose and HOMA-IR) | + | SAFHS | 850 | [119] | ||||
| HTGW and risk of T2D | WC | + | [120] | |||||
| BMI | + | MESA | 1264 | [133] | ||||
| BMI and WC | + | ARIC | 2097 | [134] | ||||
| BMI | + | Sister study | 1058 | [135] | ||||
| cg27243685 | 43,642,366 | CHD | + | FHS and PIVUS | 2306 | [79] | ||
| BMI | + | ARIC | 2097 | [134] | ||||
| cg10192877 | 43,641,690 | Post obese women after by-pass surgery | − | Women after bypass surgery/Controls | 16/14 | [132] | ||
| cg01881899 | 43,652,704 | Fasting insulin and HOMA-IR | + | GOLDN | 837 | [116] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hardy, L.M.; Frisdal, E.; Le Goff, W. Critical Role of the Human ATP-Binding Cassette G1 Transporter in Cardiometabolic Diseases. Int. J. Mol. Sci. 2017, 18, 1892. https://doi.org/10.3390/ijms18091892
AMA Style
Hardy LM, Frisdal E, Le Goff W. Critical Role of the Human ATP-Binding Cassette G1 Transporter in Cardiometabolic Diseases. International Journal of Molecular Sciences. 2017; 18(9):1892. https://doi.org/10.3390/ijms18091892
Chicago/Turabian StyleHardy, Lise M., Eric Frisdal, and Wilfried Le Goff. 2017. "Critical Role of the Human ATP-Binding Cassette G1 Transporter in Cardiometabolic Diseases" International Journal of Molecular Sciences 18, no. 9: 1892. https://doi.org/10.3390/ijms18091892
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.