2.1. Preliminary Screening for the Best Carbon Source, Inducer and Supplementary Nitrogen and Carbon Sources for the Production of Laccases by P. sanguineus RP15
The best carbon source for laccase production by
P. sanguineus RP15 was wheat bran, reaching 29.8 ± 1.9 U·g
−1 (
Table 1). Negligible enzyme levels were produced in peanut hulls and rice husks, and the organism was not able to grow on corn husks. Laccase activity was undetectable in the crude extracts obtained after
P. sanguineus culture in all other carbon sources tested. Under SmF conditions, wheat bran was a poor carbon source for laccase production by
P. sanguineus CS2 [
21]. In contrast, the lignocellulosic agro-residues sago “hampas”, rubber wood sawdust and oil palm frond parenchyma were good carbon sources for the production of laccases by
P. sanguineus CY788 under SSF conditions [
28,
29,
30], while
Eucalyptus bark shavings were revealed to be a good solid substrate for enzymatic production by a Uruguayan strain of
P. sanguineus under semi-SSF conditions [
31]. Similarly, a variety of lignocellulosic waste materials or by-products derived from food and agricultural processing industries have been successfully used as solid substrates for the production of laccases by other white-rot basidiomycetes under SSF conditions [
32,
33,
34,
35,
36]. As observed for
P. sanguineus RP15, wheat bran has been successfully employed as the main or sole carbon source for the production of laccases by
Fomes sclerodermeus [
37],
Ganoderma sp. [
38] and
Fomes fomentarius [
39] under SSF conditions, possibly due to the high content of cellulose, hemicelluloses and lignin as well as growth factors, vitamins and proteins [
40,
41].
The addition of veratryl alcohol (0–10.0 mmol
−1) to wheat bran slightly inhibited the laccase production by
P. sanguineus RP15 with respect to the control (wheat bran and water only). The inhibition was concentration-dependent and a decrease of about 23% occurred at 10 mmol·L
−1 veratryl alcohol. In contrast, the addition of CuSO
4 (0–100 mmol·L
−1) resulted in enhanced laccase production, reaching 37.2 ± 2.1 U·g
−1 (17% above the control) at 50 mmol·L
−1. At higher concentrations, however, a steady decrease of the enzymatic production occurred and 66% lower levels, as compared to the control, were obtained at 100 mmol·L
−1 CuSO
4. The production of fungal laccases is often influenced by a series of inducers, which apparently regulate the enzyme synthesis at transcriptional level. Aromatic and phenolic compounds, especially those structurally related to lignin, and metallic ions are known inducers of laccase expression. However, the effect of each compound/ion and the optimum concentration for laccase production are highly variable among different fungal strains [
1,
4,
36,
42]. Veratryl alcohol and Cu
2+ are among the most common inducers of fungal laccases, and their effect on the enzymatic production by various strains under SmF conditions is well characterized [
4,
42]. Few studies, however, addressed this point under SSF conditions. Contrasting to our results, the production of laccases by
Pycnoporus cinnabarinus ss3 [
43] and
Trametes versicolor [
16] cultured in sugarcane bagasse and horticultural waste, respectively, was increased about nine- and three-fold, respectively, by supplementation with veratryl alcohol. Furthermore, laccase production levels by
Fomes fomentarius WRF-1 [
39] and
Ganoderma sp. [
38] cultured in wheat bran were highly enhanced in response to the addition of CuSO
4. Slight inhibition was observed, however, when cultures of
Fomes sclerodermeus in wheat bran were supplemented with micromolar concentrations of CuSO
4 [
37].
The supplementation of wheat bran with 1% (
w/
w) milled corncob resulted in an increase of about 18% of the laccase production by
P. sanguineus RP15 (
Table 1). With the exception of soybean meal and corn husks, which slightly reduced the production, all other supplementary carbon sources tested were practically without effect. Up to date, the use of corncob as the main or sole carbon source for laccase production by
Pycnoporus sanguineus has not been investigated. Furthermore, few authors have studied the use of this cheap agro-residue for laccase production by other basidiomycetes, although it should provide conditions for enzyme induction due to its lignocellulosic nature [
44]. Low production levels in media containing corncob as the carbon source have been described for
Pleurotus florida EM 1303 [
44],
Lentinula edodes CCB-42 [
45] and
Trametes versicolor IBL-04 [
46].
The best supplementary nitrogen sources for the production of laccases by
P. sanguineus RP15 decreased in the following order: NH
4Cl > peptone > malt extract > casein > urea > yeast extract > asparagine > KNO
3 > NH
4NO
3 > (NH
4)
2SO
4 > NaNO
3. The production was increased about 37% by 1.0% (
w/
w) peptone and 68% by 0.8% (
w/
w) NH
4Cl, while it was inhibited about 50% by 0.8% (
w/
w) NaNO
3 (
Table 1). Thus, the quality of the nitrogen source has strongly influenced the laccase production in wheat bran, a complex substrate that presents high nitrogen levels [
47]. Furthermore, when NH
4Cl was added to wheat bran at variable concentrations (0.0%–5%
w/
w), the highest laccase levels were produced by
P. sanguineus RP15 at 0.8%–1.0% (
w/
w). Contrasting to our results, the highest laccase production by
Phlebia floridensis under SSF in wheat straw was obtained with a mixture of 9% (
w/
w) NH
4Cl and 5% (
w/
w) malt extract as nitrogen sources [
48].
2.2. Optimization of Laccase Production by P. sanguineus RP 15 Using RSM
After preliminary studies to determine the experimental ranges for each independent variable (culture time, initial moisture of the medium, temperature and milled corncob concentration) the culture conditions for the production of laccase were optimized using a 2
4 full-factorial Central Composite Rotational Design (CCRD) (
Table 2). The maximum and minimum laccase levels were, respectively, 137.2 ± 14.2 U·g
−1 (run 26) and 29.4 ± 2.4 U·g
−1 (run 21).
From the results of the CCRD, the variables that exerted significant effects on laccase production at 95% confidence level (
p < 0.05) were identified. The linear terms of temperature and milled corncob concentration, and the quadratic terms of culture time, temperature and initial moisture of the medium, exerted statistically significant effects on the enzymatic production. The interaction between the initial moisture of the medium and the temperature also had a significant effect, but not the other interactions between the independent variables tested. An ANOVA analysis was performed considering only the significant effects and the regression coefficients obtained are presented in
Table 3. The quadratic effects of the initial moisture of the medium and the temperature were the most important factors that affected the production of laccases.
The second-order model that describes the production of laccases by
P. sanguineus as a function of the independent variables tested was expressed by the equation:
Based on the F test, the model was predictive of the production of laccases as a function of culture time, culture temperature, medium initial moisture and milled corncob concentration, since the calculated F value was higher than the listed one. The R2 coefficient of 0.80 confirmed the goodness of the model and indicated that it could explain 80% of the response variability.
The statistical analysis showed that in a medium composed of wheat bran as the carbon source supplemented with 0.8% (w/w) NH4Cl and 50 mmol·L−1 CuSO4, the maximum laccase production occurred after 8.19 culture days at 26.2 °C, with initial moisture of 4.10 mL·g−1 and addition of 17.98% (w/w) milled corncob as the supplementary carbon source. The maximum predicted laccase production was 140.78 U·g−1, about 4.7-fold higher than that obtained in wheat bran and water (29.8 ± 1.9 U·g−1).
For the experimental validation of the optimized culture conditions, the fungus was grown for eight days at 26 °C in wheat bran supplemented with 0.8% (
w/
w) NH
4Cl, 50 mmol·L
−1 CuSO
4 and 18% (
w/
w) milled corncob, with initial moisture of 4.1 mL·g
−1, and the mean value determined for the laccase activity was 138.6 ± 13.2 U·g
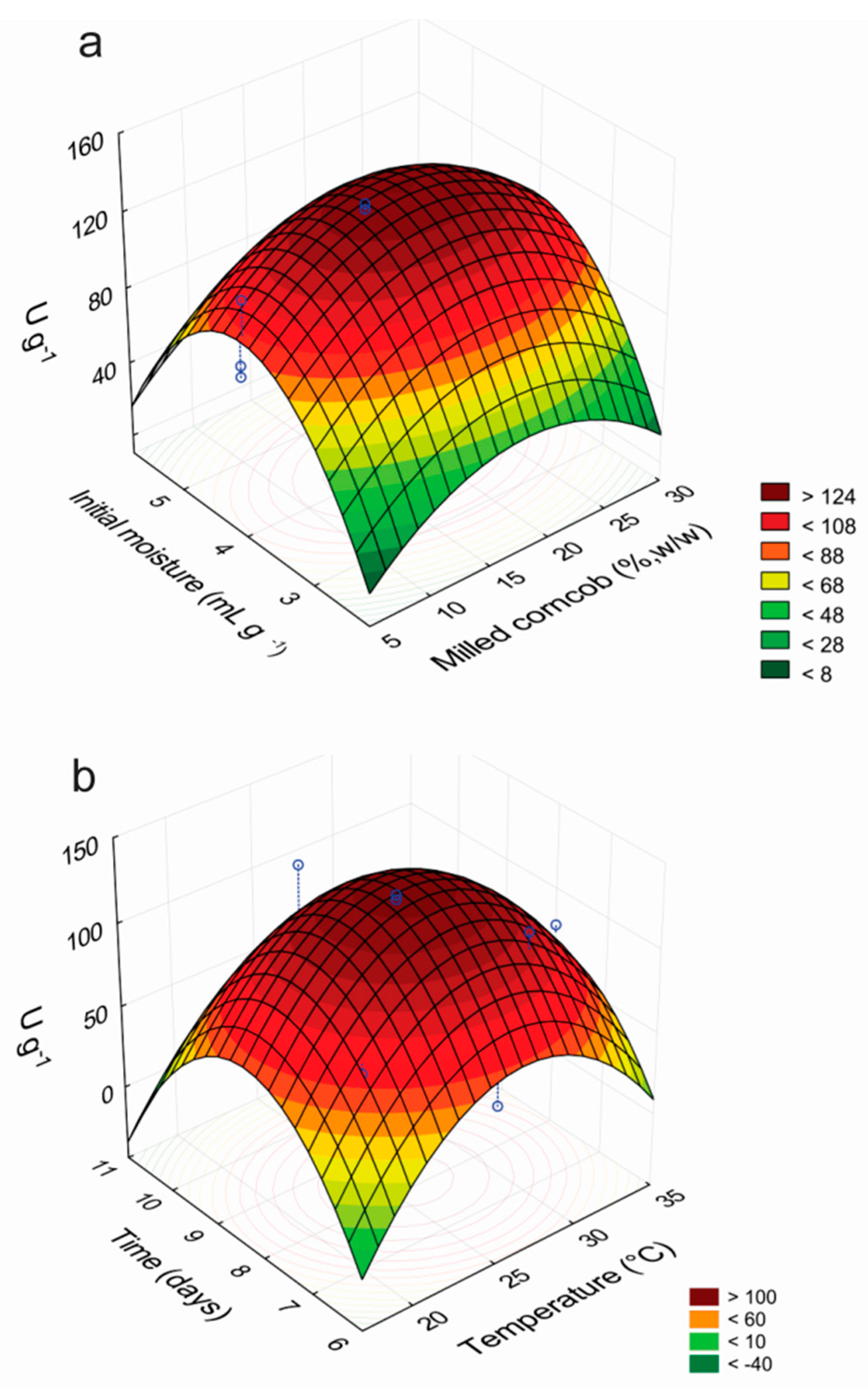
−1, in very good correlation with the value predicted by the model, confirming its validity. The response surfaces for the factors that affected the production of laccases by
P. sanguineus RP15 are presented in
Figure 1. The analysis of the curves revealed that the best conditions for the enzymatic production are in the ranges of 7.5–9.0 culture days, temperature of 25.5–27.5 °C, medium initial moisture of 3.5–4.5 mL·g
−1 and milled corncob concentration of 15%–25% (
w/
w).
The relatively short culture period, the low-cost salts employed as the supplementary nitrogen source (NH
4Cl) and inducer (CuSO
4), and the use of about 20% milled corncob as the supplementary carbon source constitute definite advantages of the culture medium employed for laccase production by
P. sanguineus RP15. In particular, corncobs constitute one of the most abundant and cheap residues of the agricultural industry around the world [
49] and the carbon source employed in a certain enzyme production process constitutes about 40%–60% of the total costs [
15].
Few authors have investigated the production of laccases by
P. sanguineus strains under conditions of SSF [
28,
29,
30] and semi-SSF [
31]. The main carbon sources used in these studies were lignocellulosic wastes and/or agro-industrial by-products, but only Annuar
et al. [
30] and Gioia
et al. [
31] performed the statistical optimization of the enzymatic production. Gioia
et al. [
31] employed ABTS to estimate the laccase activity, and under optimal conditions their
P sanguineus strain produced 106 U·g
−1 solid substrate, about 25% less than
P. sanguineus RP15. Furthermore, in spite of the low cost of the culture medium employed (bark shavings with the addition of salts, thiamine and yeast extract), the culture period for maximal enzyme production (14 days) was much higher than that determined for
P. sanguineus RP15, increasing the production costs.
In contrast to
P. sanguineus, the production of laccases by other white-rot fungi species under SSF conditions has been investigated by several authors. From three- to twenty-fold lower production levels, as compared to that obtained for
P. sanguineus RP15, were reported for different species of the genus
Pleurotus [
33,
34,
50,
51] and for
T. versicolor [
16,
51]. In great contrast, a 72-fold higher production, as compared to that obtained in this study, was described for an isolate of
Ganoderma sp. [
38]. Levels about two- and six-fold higher were also found for strains of
F. sclerodermeus [
37] and
Phanerochaete chrysosporium [
15], respectively. However, these higher productivities were obtained in more expensive culture media, as compared to that of
P. sanguineus RP15, and/or the culture times were much higher (from two- to three-fold), contributing to an increase in the production costs.
2.3. Lacps1 Purification and Molecular Properties
Lacps1 was purified by a simple two-step method from the crude culture extract from
P. sanguineus RP15 cultivated under optimized conditions for laccase production (
Table 4). The enzyme was purified about 24.5-fold with an overall yield of 30%, reaching a specific activity of 411.0 U·mg
−1. When maintained in water at 4 °C, the activity of Lacps1 remained constant for at least 60 days.
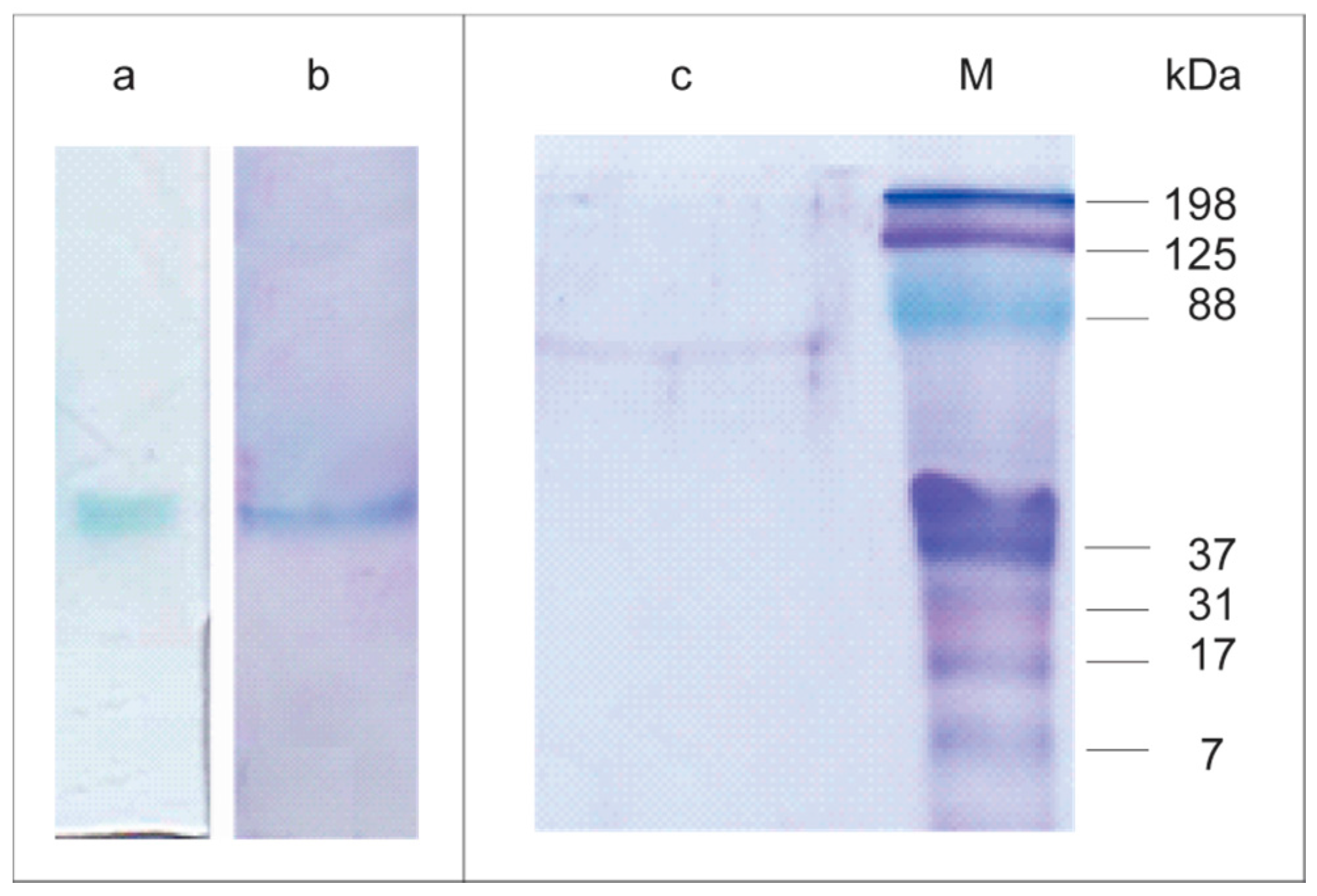
Non-denaturing polyacrylamide gel electrophoresis (PAGE) of the purified Lacps1 revealed a single laccase activity band (
Figure 2a), coincident with a single band after Coomassie Blue staining (
Figure 2b), confirming the homogeneity of the preparation. A single protein band corresponding to 65 kDa was also revealed after sodium dodecyl sulfate-PAGE (SDS-PAGE) analysis (
Figure 2c). The apparent molecular mass of Lacps1 estimated by gel filtration was 67 kDa, in good agreement with that estimated by SDS-PAGE, suggesting that the native enzyme was a monomer. Similar apparent molecular masses were reported for several laccases from other
P. sanguineus strains [
7,
17,
18,
19,
52,
53,
54,
55,
56] and other
Pycnoporus species [
7,
20].
Most extracellular fungal laccases are glycoproteins, and their carbohydrate content usually ranges from 10% to 30%. Laccases with higher saccharide content, however, have been described [
2,
4,
7]. Lacps1 showed a carbohydrate content of about 24%, close to those determined for two laccases from
P. sanguineus CS43 [
19], but about three-fold higher than those estimated for the laccases from
P. sanguineus BRFM66 and
P. sanguineus BRFM902 [
7]. The enzyme-bound carbohydrate moieties of fungal laccases greatly affect their properties, such as proteolytic susceptibility, copper retention ability, kinetic parameters, optimum pH and thermal stability [
19,
56].
Lacps1 was characterized by mass spectrometry (MS) after trypsinolysis. Three tryptic peptides were detected, and their respective precursor ion masses and amino acid sequences were:
m/
z 1,242.682 (SPGTTAADLAVIK),
m/
z 1718.75 (SAGSSEYNYDNPIFR) and
m/
z 1992.69 (ANPSFGNTGFAGGINSAILR). The results of the mass spectrometry analysis and database search demonstrated that the identified peptides shared 100% sequence identity to peptides of two laccases from
Pycnoporus coccineus M85-2 (UNIPROT: Q96TR6 and Q96VA5) [
57]. The amino acid sequences of the tryptic peptides covered about 9.3% of the amino acid sequences of both
P. coccineus laccases.
2.4. Effects of pH and Temperature on Laccase Activity
The effects of pH and temperature on the enzymatic activity of Lacps1 were evaluated using a CCRD and RSM analysis. The experimental conditions and the results of the experimental design are summarized in
Table 5.
The statistical analysis of the data revealed extremely low
p-values for the linear and quadratic effects, and also for the interaction between the pH and temperature, indicating their significant effects on the enzymatic activity (
Table 6).
The quadratic model that describes the laccase activity of Lacps1 as a function of the reaction pH and temperature was expressed by the equation:
The ANOVA for the model (
Table 6) revealed that, based on the
F test, it was predictive of the laccase activity, since the calculated
F value was much higher than the listed
F value. The
R2 coefficient (0.99) confirmed the goodness of the model.
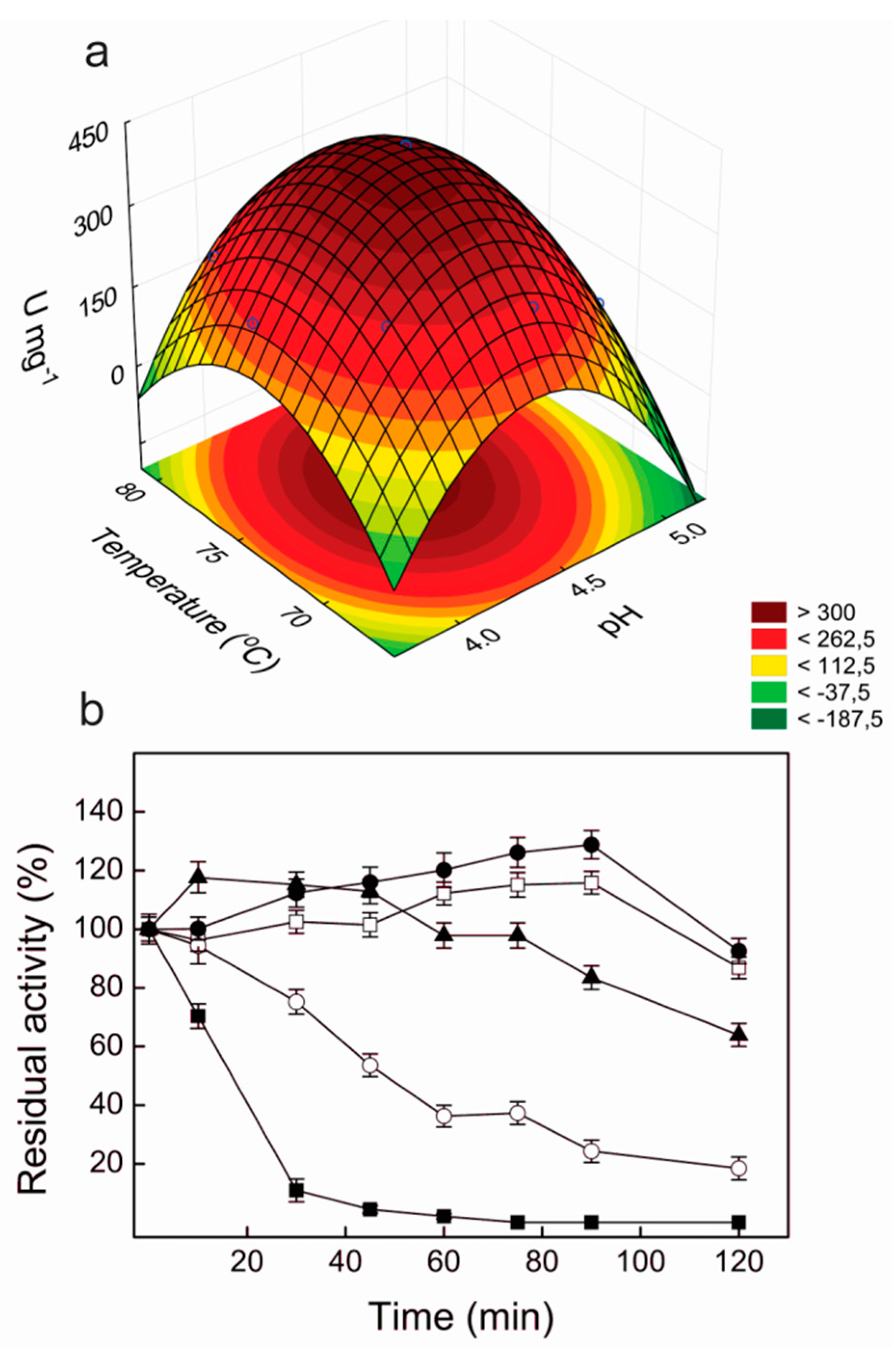
According to the statistical analysis, the optimum pH and temperature for the laccase activity were 4.4 and 74.4 °C, respectively, with a maximum predicted value of 402.4 U·mg
−1. A very close value was determined in the experimental validation (397.1 ± 20.8 U·mg
−1), confirming the validity of the model. The response surface curve for the effects of pH and temperature on the laccase activity of Lacps1 is illustrated in
Figure 3a; useful pH and temperature working ranges corresponded to 4.0–4.75 and 70–79 °C, respectively.
Among the purified laccases from
P. sanguineus described to date, most presented optimum pH for the oxidation of ABTS in the range of 2.0–3.0 [
17,
19,
53,
54,
58], and similar values were determined for two laccases from
Pycnoporus sp. SYBC [
20]. However, the enzymes from
P. sanguineus BRFM66 and
P. sanguineus BRFM902 [
7] exhibited maximum activity at pH 4.0–5.0, similarly as observed for Lacps1. In general, the laccases from other white-rot basidiomycetes exhibit pH optima for the oxidation of ABTS in the acid range (2.2–5.0) [
18,
59,
60,
61,
62,
63,
64,
65,
66].
The optimum temperature for ABTS oxidation by Lacps1 was somewhat higher than those reported for the laccases from other
P. sanguineus strains, that varied from 50 to 70 °C [
7,
17,
19,
53,
54], which is an attractive feature for application in some biotechnological processes. Optimal temperatures in the range 40–70 °C were found for laccases from other white-rot fungi [
59,
60,
61,
62,
63,
64,
65,
66].
2.6. Kinetic Properties and the Effects of Metals and Inhibitors on the Laccase Activity
The stimulation of the activity of Lacps1 by ABTS followed Michaelian kinetics, with a maximum velocity (
VM) of 413.4 ± 21.2 U·mg
−1. The Michaelis-Menten constant (
KM) value corresponded to 147.0 ± 7.4 μmol·L
−1 and the turnover number (
kcat) value was 462.4 ± 2.4 s
−1, resulting in a catalytic efficiency (
kcat/
KM) of about 3140.1 ± 149.6 L·mmol
−1·s
−1. Markedly different parameters were determined for a laccase produced by the same strain in wheat bran and water only, which oxidized ABTS with a
KM about 3.5-fold higher and a 10-fold lower
kcat value, at 25 °C, resulting in a catalytic efficiency around 88 L·mmol
−1·s
−1 [
67]. Lower
kcat values (1.15 to 236.9 s
−1), as compared to that determined for Lacps1, were also found for the laccases from other
P. sanguineus strains [
7,
17,
53,
54,
55,
56] and other white-rot fungi [
61,
62,
63,
68]. Remarkable exceptions are, however, two laccases from
P. sanguineus CS43 [
19], one from
Pycnoporus sp. SYBCL1 [
20] and two from different
Trametes species [
64,
65], with similar or higher
kcat values. The
KM values for the oxidation of ABTS by the laccases from different
P. sanguineus strains are highly variable (12 to 238 μmol·L
−1) [
7,
17,
19,
53,
54,
55,
56]. The value estimated for Lacps1 lies in the upper half of this range, resulting in a relatively low catalytic efficiency for ABTS oxidation, as compared to some other enzymes.
The activity of Lacps1 was practically unaffected by the presence of Ca
2+, Sr
2+, Co
2+, Zn
2+, Ni
2, Mg
2+ and Na
+ at 1 and 5 mmol·L
−1 concentrations. In contrast, the enzyme was highly sensitive to Hg
2+, Ag
+ and Fe
2+, with residual activities of 17.0%, 31.6% and 22.4%, respectively, at 1 mmol·L
−1 concentration and total depletion at 5 mmol·L
−1 concentration. In addition, Pb
2+ completely abolished the activity, at both concentrations tested. Similar inhibition patterns by divalent metal ions were observed for the laccases from
Pycnoporus sp. SYBCL1 [
20] and other white-rot fungi [
60,
69].
Differently, Cu
2+ stimulated about 12.7% and 31.5% the activity of Lacps1, at concentrations of 1 and 5 mmol·L
−1, respectively. Similarly to our results, stimulation by Cu
2+ has been described for two laccases from
Pycnoporus sp. SYBC-L1 [
20], as well as for the laccases from some other white-rot fungi [
59,
66,
70]. The stimulation by Cu
2+ has been attributed to the filling of the T2 copper sites on the enzyme molecules [
66]. However, some Cu
2+-inhibited laccases have also been described [
69].
Lacps1 was fairly resistant to SDS: residual activities of 78% and 17.3% were estimated in the presence of the anionic detergent at 1 and 5 mmol·L
−1 concentrations, respectively. In contrast, most laccases from white-rot fungi were strongly inhibited by SDS [
20,
60]. Ethylenediaminetetraacetic acid (EDTA) and sodium azide completely depleted the activity of Lacps1 at 1 mmol·L
−1 concentration, and a residual activity of about 10% was observed in the presence of 1 mmol·L
−1 dithiothreitol (DTT); the reducing agent completely inhibited the activity at a 5 mmol·L
−1 concentration. Inhibition by thiol compounds, azide and EDTA has been extensively described for white-rot fungi laccases [
1,
20,
60,
66,
69,
71], including two enzymes from
P. sanguineus strains [
17,
52]. However, some EDTA- and DTT-insensitive laccases have been identified [
20,
60,
69,
71]. Azide is considered a true laccase inhibitor and seems to bind to the T2 and T3 copper sites, interfering with the internal electron transfer and decreasing the catalytic activity [
69,
71]. The inhibition of laccases by DTT is less understood. According to some authors, it results from the coordination of the thiol reagent to the copper atoms at the active site [
69]. However, there is also some evidence that the inhibition could be a secondary effect, derived from a reduction of the reaction product, ABTS
+, by DTT [
71].
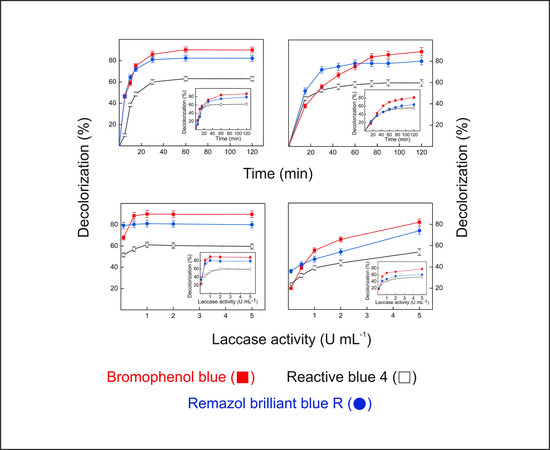
2.9. Dye Decolorization by Lacps1
The ability of the pure Lacps1 and the laccase-rich crude extract from
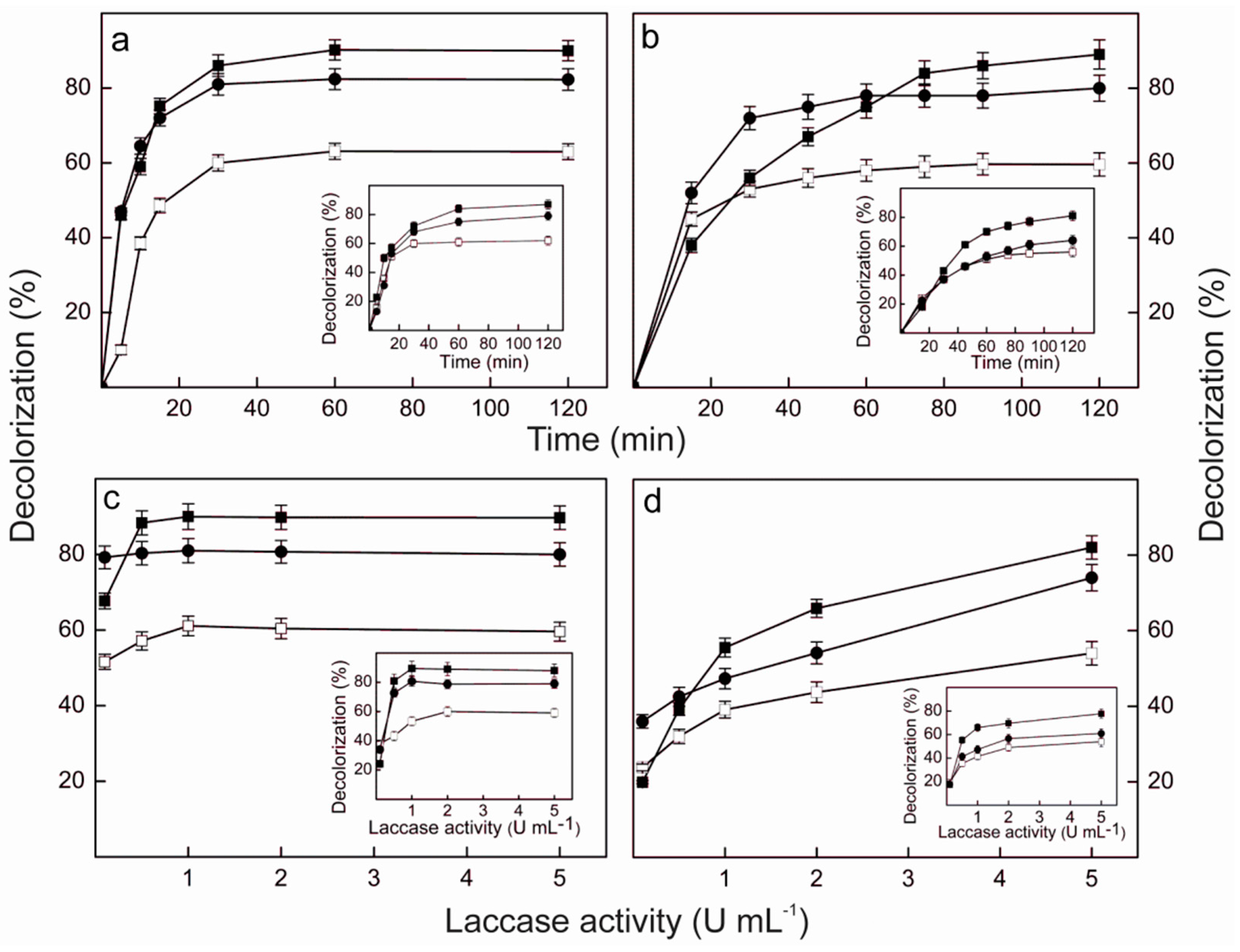
P. sanguineus RP15 to degrade RBBR, RB4 and BPB was analyzed at 25 and 40 °C (
Figure 5), temperatures in which preliminary experiments revealed that the enzyme showed good levels of activity on the dyes and excellent thermal stability. The temperature exerted little effect on the maximum levels of decolorization, when the pure enzyme was employed (
Figure 5a,c). After 120 min reaction with an enzyme load of 5 U·mL
−1, the percentages of decolorization reached about 90%, 80% and 60% for BPB, RBBR and RB4, respectively, either at 25 °C (insets to
Figure 5a,c) or 40 °C. However, at 40 °C the maximum decolorization of BPB and RBBR was attained after 30 min (
Figure 5a), while 60 min were required at 25 °C (inset to
Figure 5a). In contrast, RB4 was maximally decolorized after 30 min at both temperatures. The reaction temperature also affected the enzyme load needed to attain the maximum level of decolorization after 120 min (
Figure 5c). While at 40 °C it corresponded to 0.5 U·mL
−1 for BPB, 0.1 U·mL
−1 for RBBR and 1.0 U·mL
−1 for RB4, at 25 °C about 1.0 U·mL
−1 was required for BPB and RBBR, and 2.0 U·mL
−1 for RB4 (inset to
Figure 5c).
BPB is a triphenylmethane dye extensively used in textile, paper, leather, food and cosmetic industries [
73]. Among the different dye groups, the triphenylmethane dyes are particularly resistant to enzymatic treatments and long reaction times are usually required for, sometimes, even a partial decolorization. To the best of our knowledge, this is the first study on the decolorization of BPB by the action of a laccase from
P. sanguineus, either in pure or crude form. In great contrast to our results, a maximum decolorization level of BPB around 20% was obtained using 8 U·mL
−1 of a pure commercial laccase from
T. versicolor after three hours at 35 °C [
74], while maximum levels of about 90% were obtained with 6.5 U·mL
−1 of a pure laccase from
Trametes trogii 463 at 30 °C, but after 24 h [
75]. However, similarly as observed for Lacps1, a decolorization level of about 90% was achieved after treatment of BPB with 5 U·mL
−1 of a pure laccase from
Lentinula edodes for 90 min at 30 °C [
70].
RB4 and RBBR are typical reactive anthraquinonic dyes, widely used in the textile industry. The dyes of this class are especially resistant to degradation due to their fused aromatic rings, and most are toxic, carcinogenic and mutagenic. Furthermore, under usual dyeing conditions, up to 50% of the reactive dye initially present is discharged, generating highly colored effluents [
12,
20,
76,
77]. In spite of its unquestionable interest, the degradation of RB4 by other crude or pure laccases from
P. sanguineus has not been investigated to date. However, similar to our results, maximum levels of RB4 decolorization of about 69% were attained after 30 min at 30 °C using a laccase-rich crude extract from
T. trogii SYBC-LZ at an enzyme load of 1.1 U·mL
−1 [
78].
In great contrast, the degradation of RBBR by purified laccases has been under intense investigation in the last years. Considering the enzymes produced by different species or strains of
Pycnoporus, maximum levels of decolorization comparable to those determined for Lacps1 were obtained with a laccase from a Chinese isolate of
P. sanguineus, under similar conditions of enzyme load, reaction time and temperature [
17]. In contrast, a maximum decolorization of 20% was reached under the same conditions when a laccase from
Pycnoporus sp. SYBC-L1 was employed [
20], and 52 h were necessary to attain around 90% decolorization using each of three purified laccases from
P. coccineus, at an enzyme load of 0.1 U·mL
−1 [
7]. Among the enzymes from other white-rot fungi, similar maximum percentages of decolorization, as compared to Lacps1, were obtained with the purified laccases from
Trametes sp. SQ01 [
79] and
Cerrena sp. HYB07 [
80], under comparable reaction conditions. Maximum decolorization levels of 80%–90% have also been obtained with the laccases from
Lentinus sp. [
13],
T. versicolor [
74] and
Armillaria sp. F022 [
81], but much higher enzyme loads or reaction times were required. In contrast, a maximum decolorization of about 15% was obtained with a laccase from
Ganoderma sp. rckk-02, even after 12 h with an enzyme load of 10 U·mL
−1 [
82]. Taken together, these comparisons reveal that Lacps1 is among the most efficient white-rot fungi laccases known to date for the decolorization of BPB, RBBR and RB4.
The patterns of decolorization of BPB, RBBR and RB4 obtained using the laccase-rich crude extract from
P. sanguineus RP15 (
Figure 5b,d) were similar to those obtained with the pure Lacps1. The reaction temperature exerted little effect on the maximum levels of decolorization, which were close to those obtained with the pure enzyme: after 120 min with 5 U·mL
−1 of laccase activity the percentages of decolorization reached 80%–90% for BPB, 70%–80% for RBBR and 50%–60% for RB4, both at 25 °C (insets to
Figure 5b,d) and 40 °C. At 40 °C, however, the maximum decolorization of RBBR and RB4 was achieved after 45 min (
Figure 5b), while about 90 min and 75 min were needed at 25 °C, respectively (inset to
Figure 5b). Furthermore, the percentage of decolorization of BPB did not reach a well-defined maximum after 120 min, at both temperatures tested (
Figure 5b and inset to
Figure 5b). Similarly, the percentages of decolorization of the three dyes have not reached maxima at 5 U·mL
−1 (
Figure 5d), suggesting that higher decolorization efficiencies could be achieved using higher laccase loads, both at 25 °C (inset to
Figure 5d) and 40 °C. Similar to our results, maximal decolorization of RBBR and BPB (around 80%) occurred after 30 min at 30 °C using 6.5 U·mL
−1 of a purified laccase from
T. trogii BAFC 463 while a three-fold higher enzyme load was required to obtain a similar level of decolorization using the crude culture extract [
75].
A comparison of the results obtained with the pure Lacps1 and the crude extract strongly suggested that the decolorization of BPB, RBBR and RB4 by the crude extract could be mostly attributed to the action of Lacps1, although other oxidases and natural redox mediators could have been present and contributed to the degradation, as proposed by other authors [
75,
83]. Furthermore, the excellent potential of the
P. sanguineus RP15 crude extract for application in dye decolorization processes was undoubtedly demonstrated. Although somewhat higher enzyme loads and reaction times may be needed to attain the same decolorization efficiencies obtained with the pure enzyme, the use of crude laccase preparations at relatively low temperatures (25–40 °C) is much more economical for practical applications, minimizing energy consumption and avoiding expensive and time-consuming steps of enzyme purification [
75,
83]. In spite of all these considerations, however, the success of a practical application of either the pure Lacps1 or the crude culture extract from
P. sanguineus RP15 for the treatment of industrial effluents containing BPB, RBBR and/or RB4 depends on the results of a toxicity test of the degradation products of these dyes. In fact, the identification of the metabolites produced during the decolorization and/or biodegradation of a certain dye, as well as the evaluation of their toxicity, are essential steps for an eventual application of a laccase, assuring the safety of the treated effluents [
11,
36,
84,
85].


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}