
Characterization and Evaluation of Neuronal Trans-Differentiation with Electrophysiological Properties of Mesenchymal Stem Cells Isolated from Porcine Endometrium

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
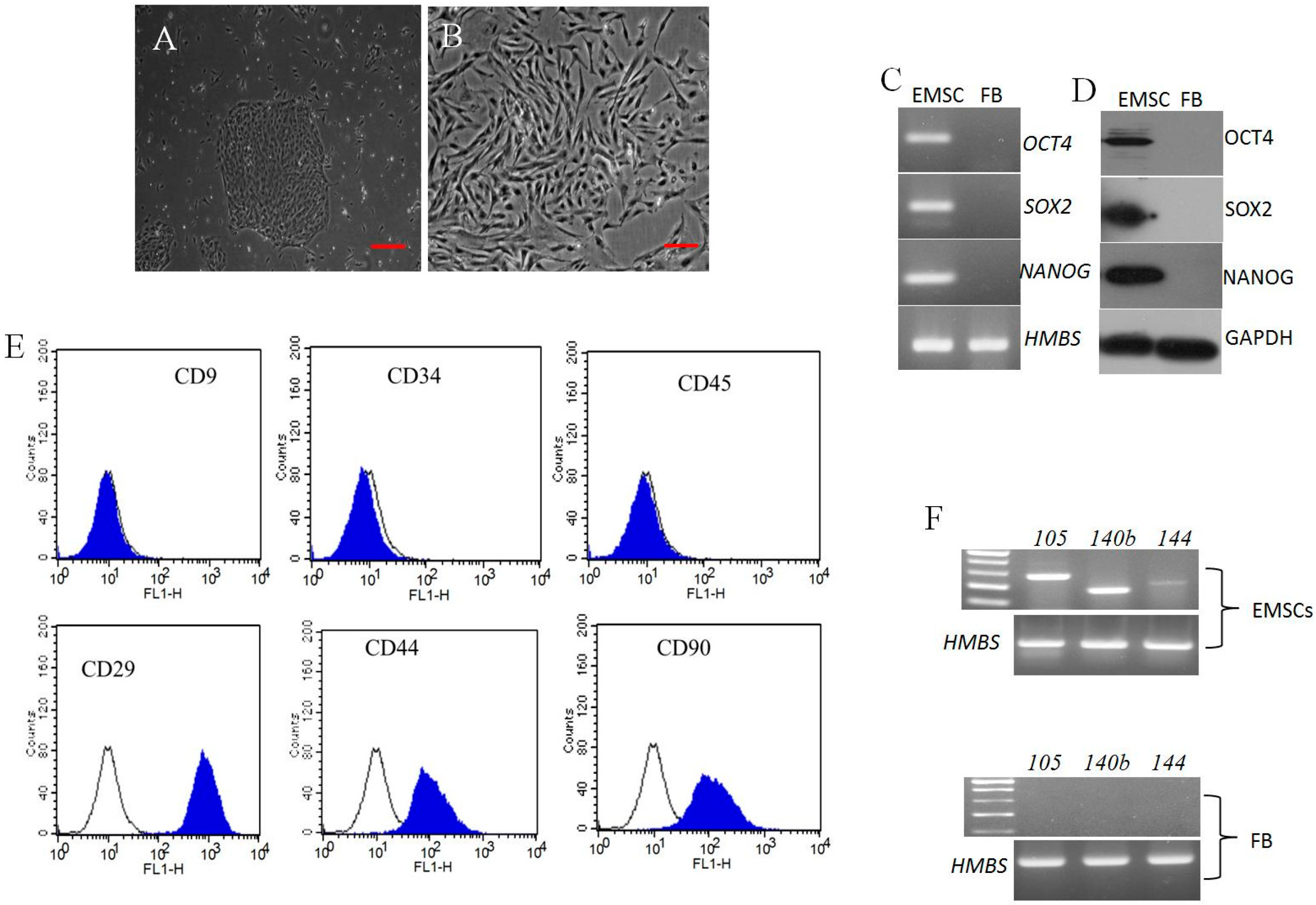
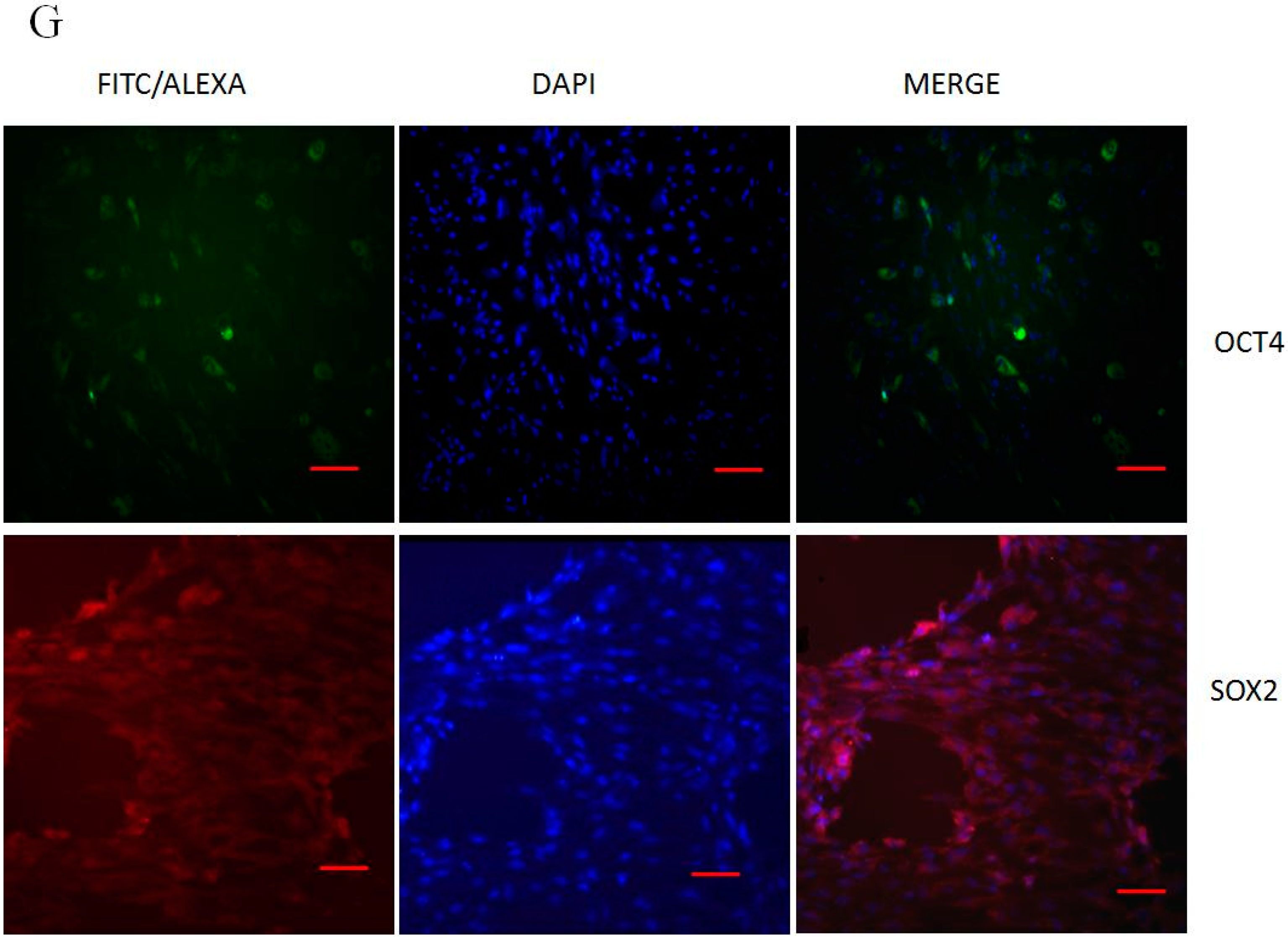
2.1. Morphology, Cell Surface Markers and Pluripotent Markers


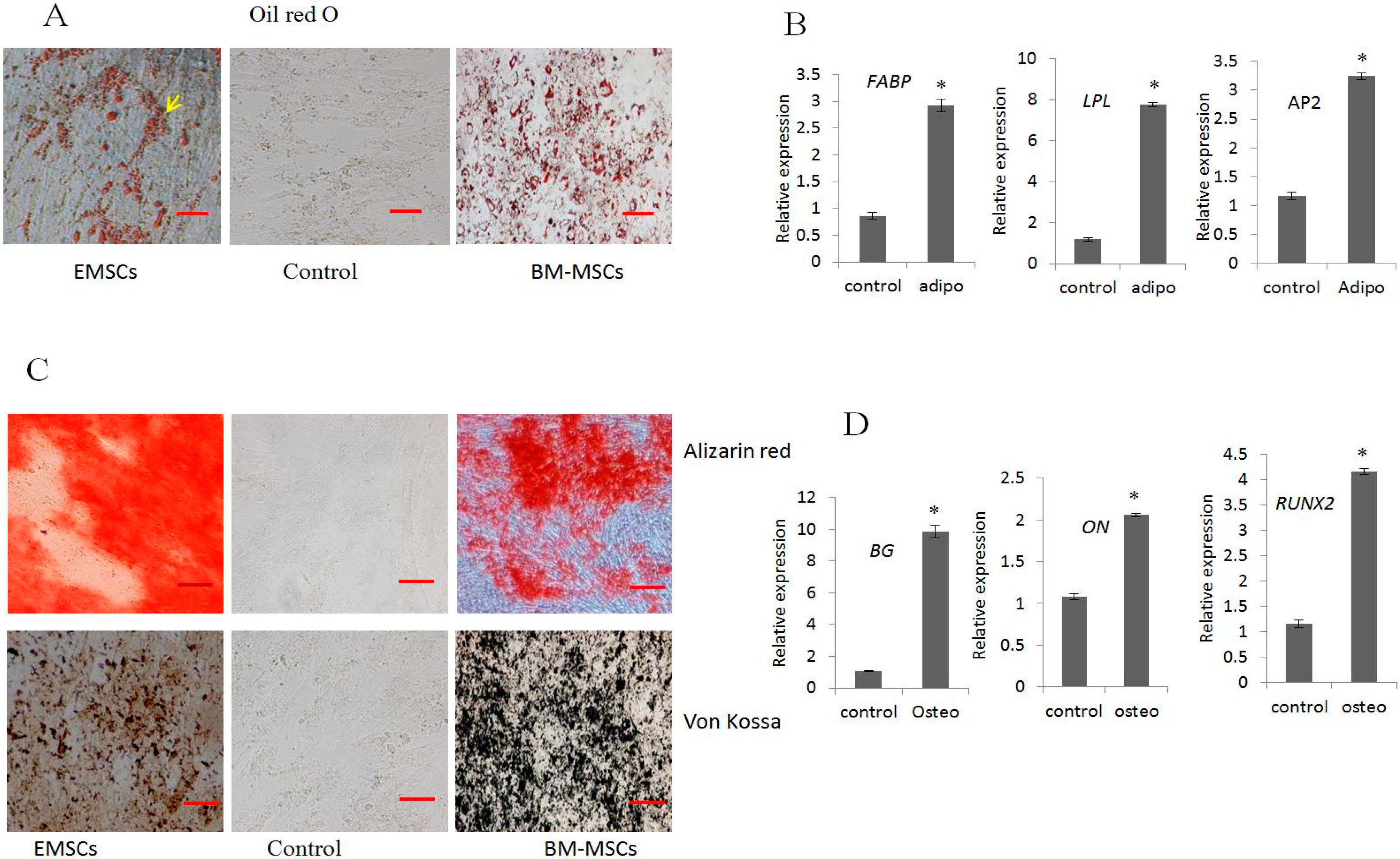
2.2. In Vitro Differentiation into Adipocytes and Osteocytes

2.3. Neurogenic Differentiation

2.4. Functional Analysis of Differentiated EMSCs into Neurogenic Cells by Electrophysiology

3. Discussion
4. Materials and Method
4.1. Chemicals and Media
4.2. Isolation and Culture of Porcine Endometrial Stromal Cell, Bone Marrow Cells and Ear Skin Fibroblast
4.3. CD Markers Analysis by Flow Cytometry and Polymerase Chain Reaction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5'–3') | Products Sizes (bp) | Annealing Tm (°C) | Reference/Accession Number |
|---|---|---|---|---|
| OCT4 | F-AGGTGTTCAGCCAAACGACC | 335 | 60 | Carlin et al., 2006 |
| R-TGATCGTTTGCCCTTCTGGC | ||||
| SOX2 | F-GCCTGGGCGCCGAGTGGA | 443 | 64 | Carlin et al., 2006 |
| R-GGGCGAGCCGTTCATGTAGGTCTG | ||||
| NANOG | F-ATCCAGCTTGTCCCCAAAG | 438 | 60 | Carlin et al., 2006 |
| R-ATTTCATTCGCTGGTTCTGG | ||||
| CD105 | F-CGCTTCAGCTTCCTCCTCCG | 281 | 56 | Miernik & Karasinski, 2012 |
| R-CACCACGGGCTCCCGCTTG | ||||
| CD140b | F-TACGTGCCCATGCTGGACATG | 175 | 54 | Miernik & Karasinski, 2012 |
| R-TGGTAGCTGAAGCCCACGAG | ||||
| CD144 | F-TGCAACGAGCGGGGCGAGTT | 220 | 56 | Miernik & Karasinski, 2012 |
| R-CGCCGCCCTCCTCATCGTA | ||||
| FABP | F-TGGTACAGGTGCAGAAGTGGGA | 100 | 60 | NM_001002817 |
| R-GCCGTGACACCTTTCATGATACA | ||||
| LPL | F-CAAACTTGTGGCTGCCCTAT | 202 | 60 | Kumar et al., 2007 |
| R-AAGGCTGTATCCCAGGAGGT | ||||
| AP2 | F-AACCCAACCTGATCACTG | 192 | 60 | AF102872.1 |
| R-TCTTTCCATCCCACTTCTGC | ||||
| ON | F-TCCGGATCTTTCCTTTGGTTTCTA | 187 | 60 | Kumar et al., 2007 |
| R-CCTTCACATCGTGGCAAGAGTTTG | ||||
| RUNX2 | F-CAGACCAGCAGCACTCCATA | 167 | 60 | XM_003482203 |
| R-AACGCCATCATTCTGGTTAG | ||||
| BG | F-GTCGTCCAGTGCTCTGACCT | 93 | 60 | XM_003135475 |
| R-GGAGCTCGGAGATGTCGTTA | ||||
| B3T | F-CAGAGCAAGAACAGCAGCAGCTACTT | 227 | 60 | XM_003480812 |
| R-GTGAACTCCATCTCATCCATGCCCTC | ||||
| NFM | F-GTCAGACCAGGCAGAAGAGG | 222 | 60 | Kumar et al., 2012 |
| R-GATTTGGGCATAGGGGATTT | ||||
| NES | F-AGGAACCAAAAGAGGCAGGT | 229 | 60 | Kumar et al., 2012 |
| R-TTGGGACCAGGGACTGTTAG | ||||
| NGF | F-CACACCGAGAGCAATGTCCC | 130 | 60 | XM_005674272 |
| R-CCACCCTGGCGGCTATCGCC | ||||
| MBP | F-GAGATGGCTCAACTCAGAACG | 125 | 60 | NM_001001546 |
| R-GGTTAGTATTTGCCGTGAGCA | ||||
| MAP2 | F-GCCATTATTCGTACACCTCCA | 291 | 60 | XM_005672149 |
| R-AGAGCCGCATTTGGATGTCAC | ||||
| HMBS | F-TTCATTCCCTCAAGGACCTG | 101 | 60 | NM_001097412 |
| R-GGGGTGAAAGACAACAGCAT |
4.4. Analysis of Pluripotent Markers by PCR and Western Blotting
4.5. Differentiation into Adipocytes and Osteocytes
4.6. Neurogenic Differentiation
4.7. RNA Isolation, cDNA Synthesis and Real Time Quantitaive PCR (RT-qPCR) Analysis
4.8. Immunofluorescence Staining
4.9. Electrophysiological Study
4.10. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mitchell, K.E.; Weiss, M.L.; Mitchell, B.M.; Martin, P.; Davis, D.; Morales, L.; Helwig, B.; Beerenstrauch, M.; Abou-Easa, K.; Hildreth, T.; et al. Matrix cells from wharton’s jelly form neurons and glia. Stem Cells 2003, 21, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; He, D.; Kleiner, G.; Kuluz, J. Neuron-like differentiation of adipose-derived stem cells from infant piglets in vitro. J. Spinal Cord Med. 2007, 30, S35–S40. [Google Scholar] [PubMed]
- Kumar, B.M.; Yoo, J.G.; Ock, S.A.; Kim, J.G.; Song, H.J.; Kang, E.J.; Cho, S.K.; Lee, S.L.; Cho, J.H.; Balasubramanian, S.; et al. In vitro differentiation of mesenchymal progenitor cells derived from porcine umbilical cord blood. Mol. Cells 2007, 24, 343–350. [Google Scholar] [PubMed]
- Park, B.W.; Kang, D.H.; Kang, E.J.; Byun, J.H.; Lee, J.S.; Maeng, G.H.; Rho, G.J. Peripheral nerve regeneration using autologous porcine skin-derived mesenchymal stem cells. J. Tissue Eng. Regen. Med. 2012, 6, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.M.; Maeng, G.H.; Lee, Y.M.; Kim, T.H.; Lee, J.H.; Jeon, B.G.; Ock, S.A.; Yoo, J.G.; Rho, G.J. Neurogenic and cardiomyogenic differentiation of mesenchymal stem cells isolated from minipig bone marrow. Res. Vet. Sci. 2012, 93, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Donovan, P.J.; Gearhart, J. The end of the beginning for pluripotent stem cells. Nature 2001, 414, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Miernik, K.; Karasinski, J. Porcine uterus contains a population of mesenchymal stem cells. Reproduction 2012, 143, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Chan, R.W.; Schwab, K.E.; Gargett, C.E. Clonogenicity of human endometrial epithelial and stromal cells. Biol. Reprod. 2004, 70, 1738–1750. [Google Scholar] [CrossRef] [PubMed]
- Wolff, E.F.; Wolff, A.B.; Hongling, D.; Taylor, H.S. Demonstration of multipotent stem cells in the adult human endometrium by in vitro chondrogenesis. Reprod. Sci. 2007, 14, 524–533. [Google Scholar] [CrossRef] [PubMed]
- Schwab, K.E.; Chan, R.W.; Gargett, C.E. Putative stem cell activity of human endometrial epithelial and stromal cells during the menstrual cycle. Fertil. Steril. 2005, 84, 1124–1130. [Google Scholar] [CrossRef] [PubMed]
- Wolff, E.F.; Gao, X.B.; Yao, K.V.; Andrews, Z.B.; Du, H.; Elsworth, J.D.; Taylor, H.S. Endometrial stem cell transplantation restores dopamine production in a Parkinson’s disease model. J. Cell. Mol. Med. 2011, 15, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Ikegami, Y.; Miyoshi, S.; Nishiyama, N.; Hida, N.; Okamoto, K.; Miyado, K.; Segawa, K.; Ogawa, S.; Umezawa, A. Serum-independent cardiomyogenic transdifferentiation in human endometrium-derived mesenchymal cells. Artif. Organs 2010, 34, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, X.; Massasa, E.E.; Feng, Y.; Wolff, E.; Taylor, H.S. Derivation of insulin producing cells from human endometrial stromal stem cells and use in the treatment of murine diabetes. Mol. Ther. 2011, 19, 2065–2071. [Google Scholar] [CrossRef] [PubMed]
- Bodek, G.; Bukowska, J.; Wisniewska, J.; Ziecik, A.J. Evidence for the presence of stem/progenitor cells in porcine endometrium. Mol. Reprod. Dev. 2015. [Google Scholar] [CrossRef]
- Gargett, C.E.; Masuda, H. Adult stem cells in the endometrium. Mol. Hum. Reprod. 2010, 16, 818–834. [Google Scholar] [CrossRef] [PubMed]
- Rho, G.J.; Kumar, B.M.; Balasubramanian, S.S. Porcine mesenchymal stem cells—Current technological status and future perspective. Front. Biosci. 2009, 14, 3942–3961. [Google Scholar] [CrossRef]
- Woodbury, D.; Schwarz, E.J.; Prockop, D.J.; Black, I.B. Adult rat and human bone marrow stromal cells differentiate into neurons. J. Neurosci. Res. 2000, 61, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Rismanchi, N.; Floyd, C.L.; Berman, R.F.; Lyeth, B.G. Cell death and long-term maintenance of neuron-like state after differentiation of rat bone marrow stromal cells: A comparison of protocols. Brain Res. 2003, 991, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.X.; He, B.P.; Cao, Q.; Tay, S.S. Protein aggregate-containing neuron-like cells are differentiated from bone marrow mesenchymal stem cells from mice with neurofilament light subunit gene deficiency. Neurosci. Lett. 2007, 417, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Kang, E.J.; Lee, Y.H.; Kim, M.J.; Lee, Y.M.; Kumar, B.M.; Jeon, B.G.; Ock, S.A.; Kim, H.J.; Rho, G.J. Transplantation of porcine umbilical cord matrix mesenchymal stem cells in a mouse model of Parkinson’s disease. J. Tissue Eng. Regen. Med. 2013, 7, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Barnabe, G.F.; Schwindt, T.T.; Calcagnotto, M.E.; Motta, F.L.; Martinez, G., Jr.; de Oliveira, A.C.; Keim, L.M.; D’Almeida, V.; Mendez-Otero, R.; Mello, L.E. Chemically-induced rat mesenchymal stem cells adopt molecular properties of neuronal-like cells but do not have basic neuronal functional properties. PLoS ONE 2009, 4, e5222. [Google Scholar] [CrossRef] [PubMed]
- Mammadov, B.; Karakas, N.; Isik, S. Comparison of long-term retinoic acid-based neural induction methods of bone marrow human mesenchymal stem cells. In vitro cellular & developmental biology. Animal 2011, 47, 484–491. [Google Scholar]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The international society for cellular therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Padykula, H.A. Regeneration in the primate uterus: The role of stem cells. Ann. N. Y. Acad. Sci. 1991, 622, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, R.; Timeva, T.; Kyurkchiev, D.; Stamenova, M.; Shterev, A.; Kostova, P.; Zlatkov, V.; Kehayov, I.; Kyurkchiev, S. Characterization of clonogenic stromal cells isolated from human endometrium. Reproduction 2008, 135, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Gargett, C.E.; Schwab, K.E.; Zillwood, R.M.; Nguyen, H.P.; Wu, D. Isolation and culture of epithelial progenitors and mesenchymal stem cells from human endometrium. Biol. Reprod. 2009, 80, 1136–1145. [Google Scholar] [CrossRef] [PubMed]
- Gargett, C.E. Uterine stem cells: What is the evidence? Hum. Reprod. Update 2007, 13, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.M.; Wu, Y.N.; Guo, X.M.; Hui, J.H.; Lee, E.H.; Lim, B. Effects of ectopic nanog and oct4 overexpression on mesenchymal stem cells. Stem Cells Dev. 2009, 18, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Matthai, C.; Horvat, R.; Noe, M.; Nagele, F.; Radjabi, A.; van Trotsenburg, M.; Huber, J.; Kolbus, A. Oct-4 expression in human endometrium. Mol. Hum. Reprod. 2006, 12, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Song, S.H.; Kumar, B.M.; Kang, E.J.; Lee, Y.M.; Kim, T.H.; Ock, S.A.; Lee, S.L.; Jeon, B.G.; Rho, G.J. Characterization of porcine multipotent stem/stromal cells derived from skin, adipose, and ovarian tissues and their differentiation in vitro into putative oocyte-like cells. Stem Cells Dev. 2011, 20, 1359–1370. [Google Scholar] [CrossRef] [PubMed]
- Carlin, R.; Davis, D.; Weiss, M.; Schultz, B.; Troyer, D. Expression of early transcription factors oct-4, sox-2 and nanog by porcine umbilical cord (puc) matrix cells. Reprod. Biol. Endocrinol. 2006, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Yin, Y.; Zhou, H.; Song, G.; Fan, A.; Tang, B.; Shi, W.; Li, Z. Proliferative capacity and pluripotent characteristics of porcine adult stem cells derived from adipose tissue and bone marrow. Cell. Reprogr. 2012, 14, 342–352. [Google Scholar]
- Kues, W.A.; Petersen, B.; Mysegades, W.; Carnwath, J.W.; Niemann, H. Isolation of murine and porcine fetal stem cells from somatic tissue. Biol. Reprod. 2005, 72, 1020–1028. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Y.; Chen, Y.J.; Chen, S.J.; Kao, C.L.; Tseng, L.M.; Lo, W.L.; Chang, C.M.; Yang, D.M.; Ku, H.H.; Twu, N.F.; et al. Induction of insulin-producing cells derived from endometrial mesenchymal stem-like cells. J. Pharmacol. Exp. Ther. 2010, 335, 817–829. [Google Scholar] [CrossRef] [PubMed]
- Faast, R.; Harrison, S.J.; Beebe, L.F.; McIlfatrick, S.M.; Ashman, R.J.; Nottle, M.B. Use of adult mesenchymal stem cells isolated from bone marrow and blood for somatic cell nuclear transfer in pigs. Cloning Stem Cells 2006, 8, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Vacanti, V.; Kong, E.; Suzuki, G.; Sato, K.; Canty, J.M.; Lee, T. Phenotypic changes of adult porcine mesenchymal stem cells induced by prolonged passaging in culture. J. Cell. Physiol. 2005, 205, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Ringe, J.; Kaps, C.; Schmitt, B.; Buscher, K.; Bartel, J.; Smolian, H.; Schultz, O.; Burmester, G.R.; Haupl, T.; Sittinger, M. Porcine mesenchymal stem cells. Induction of distinct mesenchymal cell lineages. Cell Tissue Res. 2002, 307, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Ono, M.; Maruyama, T.; Masuda, H.; Kajitani, T.; Nagashima, T.; Arase, T.; Ito, M.; Ohta, K.; Uchida, H.; Asada, H.; et al. Side population in human uterine myometrium displays phenotypic and functional characteristics of myometrial stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 18700–18705. [Google Scholar] [CrossRef] [PubMed]
- Tosh, D.; Slack, J.M. How cells change their phenotype. Nature reviews. Mol. Cell. Biol. 2002, 3, 187–194. [Google Scholar]
- Kleeberger, W.; Rothamel, T.; Glockner, S.; Flemming, P.; Lehmann, U.; Kreipe, H. High frequency of epithelial chimerism in liver transplants demonstrated by microdissection and str-analysis. Hepatology 2002, 35, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Blau, H.M.; Brazelton, T.R.; Weimann, J.M. The evolving concept of a stem cell: Entity or function? Cell 2001, 105, 829–841. [Google Scholar] [CrossRef] [PubMed]
- Quaini, F.; Urbanek, K.; Beltrami, A.P.; Finato, N.; Beltrami, C.A.; Nadal-Ginard, B.; Kajstura, J.; Leri, A.; Anversa, P. Chimerism of the transplanted heart. N. Engl. J. Med. 2002, 346, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Raff, M. Adult stem cell plasticity: Fact or artifact? Ann. Rev. Cell Dev. Biol. 2003, 19, 1–22. [Google Scholar] [CrossRef]
- Weiss, M.L.; Mitchell, K.E.; Hix, J.E.; Medicetty, S.; El-Zarkouny, S.Z.; Grieger, D.; Troyer, D.L. Transplantation of porcine umbilical cord matrix cells into the rat brain. Exp. Neurol. 2003, 182, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Anghileri, E.; Marconi, S.; Pignatelli, A.; Cifelli, P.; Galie, M.; Sbarbati, A.; Krampera, M.; Belluzzi, O.; Bonetti, B. Neuronal differentiation potential of human adipose-derived mesenchymal stem cells. Stem Cells Dev. 2008, 17, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Arthur, A.; Rychkov, G.; Shi, S.; Koblar, S.A.; Gronthos, S. Adult human dental pulp stem cells differentiate toward functionally active neurons under appropriate environmental cues. Stem Cells 2008, 26, 1787–1795. [Google Scholar] [CrossRef] [PubMed]
- Chao, M.V. Neurotrophins and their receptors: A convergence point for many signalling pathways. Nat. Rev. Neurosci. 2003, 4, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Jeon, B.G.; Kwack, D.O.; Rho, G.J. Variation of telomerase activity and morphology in porcine mesenchymal stem cells and fibroblasts during prolonged in vitro culture. Anim. Biotechnol. 2011, 22, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subbarao, R.B.; Ullah, I.; Kim, E.-J.; Jang, S.-J.; Lee, W.-J.; Jeon, R.H.; Kang, D.; Lee, S.-L.; Park, B.-W.; Rho, G.-J. Characterization and Evaluation of Neuronal Trans-Differentiation with Electrophysiological Properties of Mesenchymal Stem Cells Isolated from Porcine Endometrium. Int. J. Mol. Sci. 2015, 16, 10934-10951. https://doi.org/10.3390/ijms160510934
Subbarao RB, Ullah I, Kim E-J, Jang S-J, Lee W-J, Jeon RH, Kang D, Lee S-L, Park B-W, Rho G-J. Characterization and Evaluation of Neuronal Trans-Differentiation with Electrophysiological Properties of Mesenchymal Stem Cells Isolated from Porcine Endometrium. International Journal of Molecular Sciences. 2015; 16(5):10934-10951. https://doi.org/10.3390/ijms160510934
Chicago/Turabian StyleSubbarao, Raghavendra Baregundi, Imran Ullah, Eun-Jin Kim, Si-Jung Jang, Won-Jae Lee, Ryoung Hoon Jeon, Dawon Kang, Sung-Lim Lee, Bong-Wook Park, and Gyu-Jin Rho. 2015. "Characterization and Evaluation of Neuronal Trans-Differentiation with Electrophysiological Properties of Mesenchymal Stem Cells Isolated from Porcine Endometrium" International Journal of Molecular Sciences 16, no. 5: 10934-10951. https://doi.org/10.3390/ijms160510934