
Human-Based New Approach Methodologies in Developmental Toxicity Testing: A Step Ahead from the State of the Art with a Feto–Placental Organ-on-Chip Platform
 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. The Regulatory Context: The OECD TG 414
3. Human-Relevant NAMs for Developmental Toxicity Testing
3.1. Human-Relevant NAMs for Placenta Toxicology
3.1.1. Ex Vivo Dual Side Human Placenta Perfusion
3.1.2. Human Placenta Ex Vivo Explant Cultures
3.2. Human-Relevant NAMs of Amniotic Membranes
3.3. Human-Relevant NAMs for Embryonic/Fetal Toxicology
3.3.1. In Vitro Cultures of ESCs and iPSCs and Derived 3D Organotypic Models
3.3.2. Primary Fetal Cell Culture Models
3.3.3. Organotypic Fetal Models
3.3.4. Remarks
3.4. Immortalization of Cellular Models in Developmental Toxicology Testing
3.5. In Silico Models for Developmental Toxicity
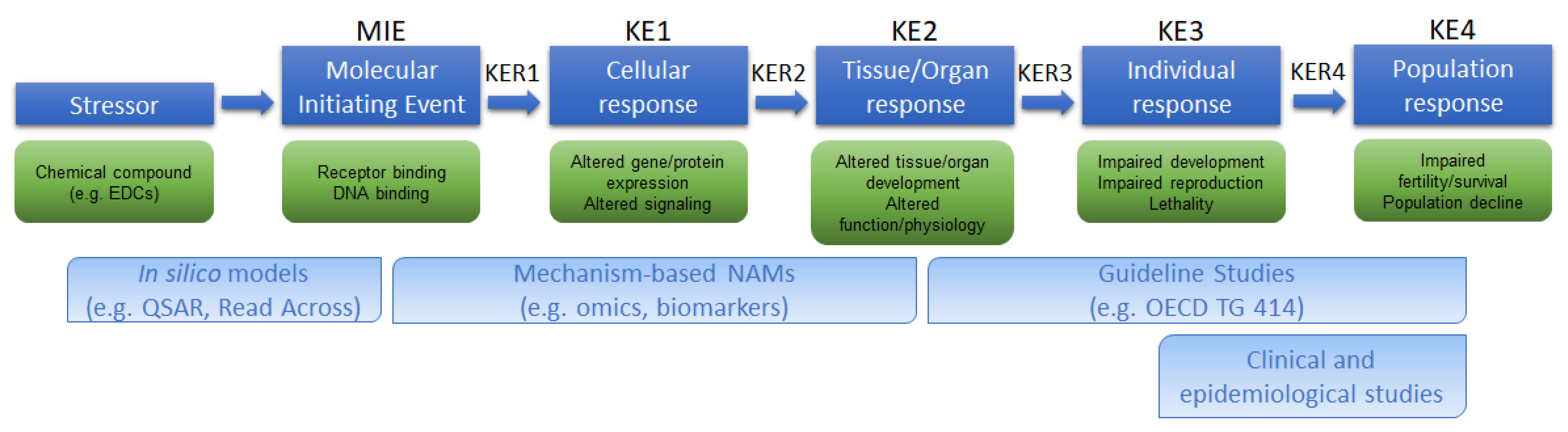
4. Available AOPs Related to TG 414
4.1. Neurodevelopmental Toxicity
4.2. Reproductive Toxicity
4.3. Cardiovascular Toxicity
4.4. Liver Toxicity
4.5. Other Soft Organs’ Toxicity
4.6. Skeletal Malformations
4.7. Immunotoxicity
4.8. Other Developmental Outcomes
4.9. Placental Toxicity
4.10. Remarks
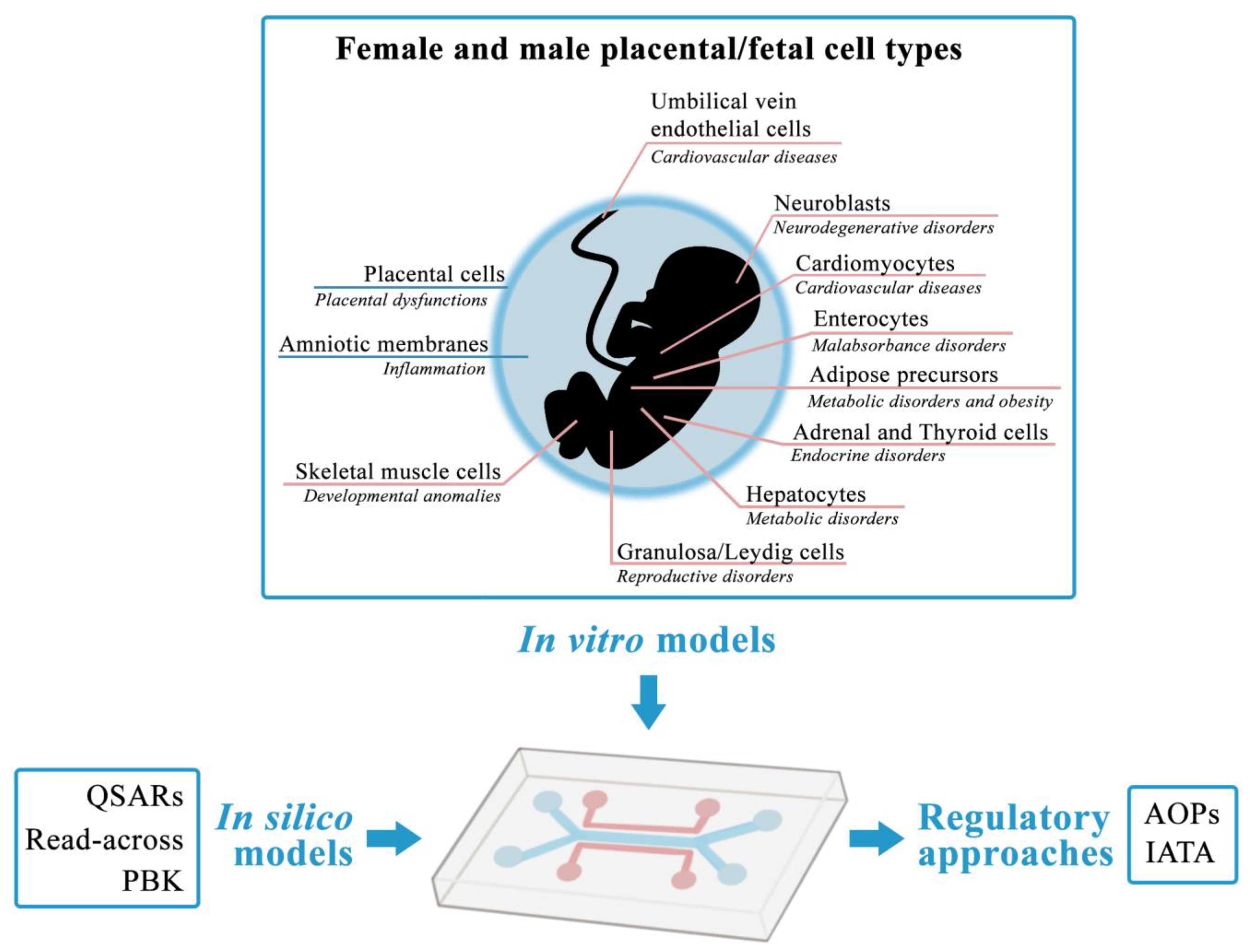
5. Future Perspective of a Feto–Placental OoC System: Advantages and Potential
- To have a strong human relevance due to the choice of cellular models more closely recapitulating normal physiology and multicellular complexity.
- To be based on human cells representative of different fetal target organs, at the sensitive window of development, relevant for EDCs identification and characterization.
- To reconstruct a more physiological fetal environment, by adding placental and amniotic barriers to the system, thus mimicking metabolic, hormonal and immune crosstalk occurring during gestation.
- To enable a modular configuration of the placental downstream fetal organs to reproduce relevant in vivo axes.
- To be sex-specifically assembled, coupling placental cells from male or female pregnancies to amniotic and fetal cells of matched sex, for the identification of sex-specific effects of stressors, especially EDCs, and sex-related sensitivities to develop chronic diseases later in life.
- To represent an advanced, miniaturized NAM platform based on the integration of in vitro and in silico models, enabling high throughput and high content screening, as well as set up of fit-for-purpose readouts linked to regulatory relevant AOPs for the inclusion in an IATA approach.
- To strongly support the reduction of animal studies due to the human relevance and the transferability of the obtained results.
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barker, D.J. The developmental origins of chronic disease. In Families and Child Health; Landale, N.S., McHale, S.M., Booth, A., Eds.; Springer: New York, NY, USA, 2013; pp. 3–11. [Google Scholar]
- European Commission. Regulation (EC) no 1907/2006 of the European Parliament and of the Council of 18 December 2006 Concerning the Registration, Evaluation, Authorisation and Restriction of Chemicals (REACH); Establishing a European Chemicals Agency, Amending Directive 1999/45/EC and Repealing Council Regulation (EEC) no 793/93 and Commission Regulation (EC) no 1488/94 as Well as Council Directive 76/769/EEC and Commission Directives 91/155/EEC, 93/67/EEC, 93/105/EC and 2000/21/EC; European Union: Brussels, Belgium, 2006; pp. 1–849. [Google Scholar]
- Bourguignon, J.; Parent, A.; Kleinjans, J.; Nawrot, T.S.; Schoeters, G.; Van Larebeke, N. Rationale for Environmental Hygiene Towards Global Protection of Fetuses and Young Children from Adverse Lifestyle Factors. Environ. Health 2018, 17, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Attina, T.M.; Hauser, R.; Sathyanarayana, S.; Hunt, P.A.; Bourguignon, J.; Myers, J.P.; DiGangi, J.; Zoeller, R.T.; Trasande, L. Exposure to Endocrine-Disrupting Chemicals in the USA: A Population-Based Disease Burden and Cost Analysis. Lancet Diabetes Endocrinol. 2016, 4, 996–1003. [Google Scholar] [CrossRef] [PubMed]
- Malits, J.; Naidu, M.; Trasande, L. Exposure to Endocrine Disrupting Chemicals in Canada: Population-Based Estimates of Disease Burden and Economic Costs. Toxics 2022, 10, 146. [Google Scholar] [CrossRef] [PubMed]
- Grandjean, P.; Bellanger, M. Calculation of the Disease Burden Associated with Environmental Chemical Exposures: Application of Toxicological Information in Health Economic Estimation. Environ. Health 2017, 16, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, G.J.; Fowden, A.L.; Thornburg, K.L. Placental Origins of Chronic Disease. Physiol. Rev. 2016, 96, 1509–1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, A.; Schmidt, A.; Markert, U.R. The Road (Not) taken—Placental Transfer and Interspecies Differences. Placenta 2021, 115, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Rovida, C.; Hartung, T. Re-Evaluation of Animal Numbers and Costs for in Vivo Tests to Accomplish REACH Legislation Requirements for Chemicals—A Report by the Transatlantic Think Tank for Toxicology (T4). ALTEX-Altern. Anim. Exp. 2009, 26, 187–208. [Google Scholar] [CrossRef] [Green Version]
- Interagency Coordinating Committee on the Validation of Alternative Methods. A Strategic Roadmap for Establishing New Approaches to Evaluate the Safety of Chemicals and Medical Products in the United States; Interagency Coordinating Committee on the Validation of Alternative Methods: Research Triangle, NC, USA, 2018.
- US Environmental Protection Agency (EPA). Strategic Plan to Promote the Development and Implementation of Alternative Test Methods within the TSCA Program; USEPA: Washington, DC, USA, 2018.
- European Chemicals Agency (ECHA). New Approach Methodologies in Regulatory Science. In Proceedings of the Scientific Workshop, Helsinki, Finland, 19–20 April 2016. [Google Scholar]
- Escher, S.E.; Partosch, F.; Konzok, S.; Jennings, P.; Luijten, M.; Kienhuis, A.; de Leeuw, V.; Reuss, R.; Lindemann, K.; Bennekou, S.H. Development of a Roadmap for Action on New Approach Methodologies in Risk Assessment. EFSA Support. Publ. 2022, 19, 7341E. [Google Scholar] [CrossRef]
- OECD. Prenatal Developmental Toxicity Study (OECD TG 414); OECD: Paris, France, 2018. [Google Scholar]
- OECD. Guidance Document for the use of Adverse Outcome Pathways in Developing Integrated Approaches to Testing and Assessment (IATA); OECD: Paris, France, 2017. [Google Scholar]
- Estevan, C.; Pamies, D.; Sogorb, M.A.; Vilanova, E. Reproductive toxicity: In vivo testing guidelines from OECD. In Reproductive and Developmental Toxicology; Gupta, R.C., Ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 165–178. [Google Scholar]
- OECD. Test no. 421: Reproduction/Developmental Toxicity Screening Test; OECD: Paris, France, 2016. [Google Scholar]
- OECD. Test no. 422: Combined Repeated Dose Toxicity Study with the Reproduction/Developmental Toxicity Screening Test; OECD: Paris, France, 2015. [Google Scholar]
- Reuter, U.; Heinrich-Hirsch, B.; Hellwig, J.; Holzum, B.; Welsch, F. Evaluation of OECD Screening Tests 421 (Reproduction/Developmental Toxicity Screening Test) and 422 (Combined Repeated Dose Toxicity Study with the Reproduction/Developmental Toxicity Screening Test). Regul. Toxicol. Pharmacol. 2003, 38, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Fleischer, M. Testing Costs and Testing Capacity According to the REACH Requirements—Results of a Survey of Independent and Corporate GLP Laboratories in the EU and Switzerland. J. Bus. Chem. 2007, 4, 96–114. [Google Scholar]
- Höfer, T.; Gerner, I.; Gundert-Remy, U.; Liebsch, M.; Schulte, A.; Spielmann, H.; Vogel, R.; Wettig, K. Animal Testing and Alternative Approaches for the Human Health Risk Assessment Under the Proposed New European Chemicals Regulation. Arch. Toxicol. 2004, 78, 549–564. [Google Scholar] [CrossRef]
- Busquet, F.; Kleensang, A.; Rovida, C.; Herrmann, K.; Leist, M.; Hartung, T. New European Union Statistics on Laboratory Animal Use—What Really Counts! ALTEX-Altern. Anim. Exp. 2020, 37, 167–186. [Google Scholar] [CrossRef] [Green Version]
- Petetta, F.; Ciccocioppo, R. Public Perception of Laboratory Animal Testing: Historical, Philosophical, and Ethical View. Addict. Biol. 2021, 26, e12991. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Fowden, A.L. The Placenta: A Multifaceted, Transient Organ. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140066. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.; Morales-Prieto, D.M.; Pastuschek, J.; Fröhlich, K.; Markert, U.R. Only Humans have Human Placentas: Molecular Differences between Mice and Humans. J. Reprod. Immunol. 2015, 108, 65–71. [Google Scholar] [CrossRef]
- Schneider, H.; Albrecht, C.; Ahmed, M.S.; Broekhuizen, M.; Aengenheister, L.; Buerki-Thurnherr, T.; Danser, A.J.; Gil, S.; Hansson, S.R.; Greupink, R. Ex Vivo Dual Perfusion of an Isolated Human Placenta Cotyledon: Towards Protocol Standardization and Improved Inter-Centre Comparability. Placenta 2022, 126, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Schneider, H.; Panigel, M.; Dancis, J. Transfer Across the Perfused Human Placenta of Antipyrine, Sodium, and Leucine. Obstet. Gynecol. 1972, 114, 822–828. [Google Scholar] [CrossRef]
- Mathiesen, L.; Buerki-Thurnherr, T.; Pastuschek, J.; Aengenheister, L.; Knudsen, L.E. Fetal Exposure to Environmental Chemicals; Insights from Placental Perfusion Studies. Placenta 2021, 106, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Kummu, M.; Sieppi, E.; Koponen, J.; Laatio, L.; Vähäkangas, K.; Kiviranta, H.; Rautio, A.; Myllynen, P. Organic Anion Transporter 4 (OAT 4) Modifies Placental Transfer of Perfluorinated Alkyl Acids PFOS and PFOA in Human Placental Ex Vivo Perfusion System. Placenta 2015, 36, 1185–1191. [Google Scholar] [CrossRef] [PubMed]
- Pidoux, G.; Gerbaud, P.; Guibourdenche, J.; Thérond, P.; Ferreira, F.; Simasotchi, C.; Evain-Brion, D.; Gil, S. Formaldehyde Crosses the Human Placenta and Affects Human Trophoblast Differentiation and Hormonal Functions. PLoS ONE 2015, 10, e0133506. [Google Scholar] [CrossRef] [Green Version]
- Conings, S.; Amant, F.; Annaert, P.; Van Calsteren, K. Integration and Validation of the Ex Vivo Human Placenta Perfusion Model. J. Pharmacol. Toxicol. Methods 2017, 88, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.; Genbacev, O.; Turner, M.; Aplin, J.; Caniggia, I.; Huppertz, B. Human Placental Explants in Culture: Approaches and Assessments. Placenta 2005, 26, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Gundacker, C.; Ellinger, I. The Unique Applicability of the Human Placenta to the Adverse Outcome Pathway (AOP) Concept: The Placenta Provides Fundamental Insights into Human Organ Functions at Multiple Levels of Biological Organization. Reprod. Toxicol. 2020, 96, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.; Berndt, W. Characterization of Neutral Amino Acid Accumulation by Human Term Placental Slices. Am. J. Physiol.-Leg. Content 1974, 227, 1236–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kupper, N.; Pritz, E.; Siwetz, M.; Guettler, J.; Huppertz, B. Placental Villous Explant Culture 2.0: Flow Culture Allows Studies Closer to the in Vivo Situation. Int. J. Mol. Sci. 2021, 22, 7464. [Google Scholar] [CrossRef]
- Mai, P.; Hampl, J.; Baca, M.; Brauer, D.; Singh, S.; Weise, F.; Borowiec, J.; Schmidt, A.; Küstner, J.M.; Klett, M. MatriGrid® Based Biological Morphologies: Tools for 3D Cell Culturing. Bioengineering 2022, 9, 220. [Google Scholar] [CrossRef] [PubMed]
- James, J.L.; Lissaman, A.; Nursalim, Y.N.; Chamley, L.W. Modelling Human Placental Villous Development: Designing Cultures that Reflect Anatomy. Cell. Mol. Life Sci. 2022, 79, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Siman, C.; Sibley, C.; Jones, C.; Turner, M.; Greenwood, S. The Functional Regeneration of Syncytiotrophoblast in Cultured Explants of Term Placenta. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2001, 280, R1116–R1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gingrich, J.; Ticiani, E.; Veiga-Lopez, A. Placenta Disrupted: Endocrine Disrupting Chemicals and Pregnancy. Trends Endocrinol. Metab. 2020, 31, 508–524. [Google Scholar] [CrossRef] [PubMed]
- Morck, T.J.; Sorda, G.; Bechi, N.; Rasmussen, B.S.; Nielsen, J.B.; Ietta, F.; Rytting, E.; Mathiesen, L.; Paulesu, L.; Knudsen, L.E. Placental Transport and in Vitro Effects of Bisphenol, A. Reprod. Toxicol. 2010, 30, 131–137. [Google Scholar] [CrossRef]
- Boadi, W.Y.; Urbach, J.; Barnea, E.R.; Brand, J.M.; Yannai, S. Enzyme Activities in the Term Human Placenta: In Vitro Effect of Cadmium. Pharmacol. Toxicol. 1992, 71, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Boadi, W.Y.; Urbach, J.; Brandes, J.M.; Yannai, S. In Vitro Exposure to Mercury and Cadmium Alters Term Human Placental Membrane Fluidity. Toxicol. Appl. Pharmacol. 1992, 116, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Wójtowicz, A.K.; Honkisz, E.; Ziąba-Przybylska, D.; Milewicz, T.; Kajta, M. Effects of Two Isomers of DDT and their Metabolite DDE on CYP1A1 and AhR Function in Human Placental Cells. Pharmacol. Rep. 2011, 63, 1460–1468. [Google Scholar] [CrossRef] [PubMed]
- Bechi, N.; Ietta, F.; Romagnoli, R.; Jantra, S.; Cencini, M.; Galassi, G.; Serchi, T.; Corsi, I.; Focardi, S.; Paulesu, L. Environmental Levels of Para-Nonylphenol are Able to Affect Cytokine Secretion in Human Placenta. Environ. Health Perspect. 2010, 118, 427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, S.V.; Atala, A. Amniotic Fluid and Placental Membranes: Unexpected Sources of Highly Multipotent Cells. Semin. Reprod. Med. 2013, 31, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Miwa, H.; Yasui, M. Inflammatory Mediators Weaken the Amniotic Membrane Barrier through Disruption of Tight Junctions. J. Physiol. 2010, 588, 4859–4869. [Google Scholar] [CrossRef] [PubMed]
- Brant, K.A.; Caruso, R.L. PCB 50 Stimulates Release of Arachidonic Acid and Prostaglandins from Late Gestation Rat Amnion Fibroblast Cells. Reprod. Toxicol. 2006, 22, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Behnia, F.; Peltier, M.R.; Saade, G.R.; Menon, R. Environmental Pollutant Polybrominated Diphenyl Ether, a Flame Retardant, Induces Primary Amnion Cell Senescence. Am. J. Reprod. Immunol. 2015, 74, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Xing, M.; Lou, J.; Wang, X.; Fu, W.; Xu, L. Apoptotic Related Biochemical Changes in Human Amnion Cells Induced by Tributyltin. Toxicology 2007, 230, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhao, M.; Zhang, C.; Ma, Y.; Liu, W. Enantioselective Cytotoxicity of the Insecticide Bifenthrin on a Human Amnion Epithelial (FL) Cell Line. Toxicology 2008, 253, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Gaggi, G.; Di Credico, A.; Izzicupo, P.; Antonucci, I.; Crescioli, C.; Di Giacomo, V.; Di Ruscio, A.; Amabile, G.; Alviano, F.; Di Baldassarre, A. Epigenetic Features of Human Perinatal Stem Cells Redefine their Stemness Potential. Cells 2020, 9, 1304. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.F.; Loch-Caruso, R. Comparison of LPS-Stimulated Release of Cytokines in Punch Versus Transwell Tissue Culture Systems of Human Gestational Membranes. Reprod. Biol. Endocrinol. 2010, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Gnecco, J.S.; Anders, A.P.; Cliffel, D.; Pensabene, V.; Rogers, L.M.; Osteen, K.; Aronoff, D.M. Instrumenting a Fetal Membrane on a Chip as Emerging Technology for Preterm Birth Research. Curr. Pharm. Des. 2017, 23, 6115–6124. [Google Scholar] [CrossRef]
- Banerjee, A.; Weidinger, A.; Hofer, M.; Steinborn, R.; Lindenmair, A.; Hennerbichler-Lugscheider, S.; Eibl, J.; Redl, H.; Kozlov, A.V.; Wolbank, S. Different Metabolic Activity in Placental and Reflected Regions of the Human Amniotic Membrane. Placenta 2015, 36, 1329–1332. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Lindenmair, A.; Hennerbichler, S.; Steindorf, P.; Steinborn, R.; Kozlov, A.V.; Redl, H.; Wolbank, S.; Weidinger, A. Cellular and Site-Specific Mitochondrial Characterization of Vital Human Amniotic Membrane. Cell Transplant. 2018, 27, 3–11. [Google Scholar] [CrossRef]
- Banerjee, A.; Lindenmair, A.; Steinborn, R.; Dumitrescu, S.D.; Hennerbichler, S.; Kozlov, A.V.; Redl, H.; Wolbank, S.; Weidinger, A. Oxygen Tension Strongly Influences Metabolic Parameters and the Release of Interleukin-6 of Human Amniotic Mesenchymal Stromal Cells in Vitro. Stem Cells Int. 2018, 2018, 9502451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poženel, L.; Lindenmair, A.; Schmidt, K.; Kozlov, A.V.; Grillari, J.; Wolbank, S.; Banerjee, A.; Weidinger, A. Critical Impact of Human Amniotic Membrane Tension on Mitochondrial Function and Cell Viability in Vitro. Cells 2019, 8, 1641. [Google Scholar] [CrossRef] [Green Version]
- Kuske, B.; Pulyanina, P.Y.; Nieden, N.I.Z. Embryonic stem cell test: Stem cell use in predicting developmental cardiotoxicity and osteotoxicity. In Developmental Toxicology; Harris, C., Hansen, J.M., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 147–179. [Google Scholar]
- Palmer, J.A.; Smith, A.M.; Egnash, L.A.; Conard, K.R.; West, P.R.; Burrier, R.E.; Donley, E.L.; Kirchner, F.R. Establishment and Assessment of a New Human Embryonic Stem Cell-based Biomarker Assay for Developmental Toxicity Screening. Birth Defects Res. Part B Dev. Reprod. Toxicol. 2013, 98, 343–363. [Google Scholar] [CrossRef]
- Theunissen, P.T.; Pennings, J.L.; van Dartel, D.A.; Robinson, J.F.; Kleinjans, J.C.; Piersma, A.H. Complementary Detection of Embryotoxic Properties of Substances in the Neural and Cardiac Embryonic Stem Cell Tests. Toxicol. Sci. 2013, 132, 118–130. [Google Scholar] [CrossRef] [Green Version]
- Marikawa, Y.; Chen, H.; Menor, M.; Deng, Y.; Alarcon, V.B. Exposure-Based Assessment of Chemical Teratogenicity using Morphogenetic Aggregates of Human Embryonic Stem Cells. Reprod. Toxicol. 2020, 91, 74–91. [Google Scholar] [CrossRef]
- Pratten, M.; Ahir, B.K.; Smith-Hurst, H.; Memon, S.; Mutch, P.; Cumberland, P. Primary cell and micromass culture in assessing developmental toxicity. In Developmental Toxicology; Harris, C., Hansen, J.M., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 115–146. [Google Scholar]
- Di Paolo, C.; Hoffmann, S.; Witters, H.; Carrillo, J. Minimum Reporting Standards Based on a Comprehensive Review of the Zebrafish Embryo Teratogenicity Assay. Regul. Toxicol. Pharmacol. 2021, 127, 105054. [Google Scholar] [CrossRef]
- Yamanaka, S. Elite and Stochastic Models for Induced Pluripotent Stem Cell Generation. Nature 2009, 460, 49–52. [Google Scholar] [CrossRef]
- Kugler, J.; Huhse, B.; Tralau, T.; Luch, A. Embryonic Stem Cells and the Next Generation of Developmental Toxicity Testing. Expert Opin. Drug Metab. Toxicol. 2017, 13, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Shinozawa, T. Human Organoids for Predictive Toxicology Research and Drug Development. Front. Genet. 2021, 12, 2119. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; McKnight, K.D.; Wong, D.J.; Rodriguez, R.T.; Sugiyama, T.; Gu, X.; Ghodasara, A.; Qu, K.; Chang, H.Y.; Kim, S.K. A Molecular Signature for Purified Definitive Endoderm Guides Differentiation and Isolation of Endoderm from Mouse and Human Embryonic Stem Cells. Stem Cells Dev. 2012, 21, 2273–2287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baillie-Benson, P.; Moris, N.; Arias, A.M. Pluripotent Stem Cell Models of Early Mammalian Development. Curr. Opin. Cell Biol. 2020, 66, 89–96. [Google Scholar] [CrossRef] [PubMed]
- van den Brink, S.C.; van Oudenaarden, A. 3D Gastruloids: A Novel Frontier in Stem Cell-Based in Vitro Modeling of Mammalian Gastrulation. Trends Cell Biol. 2021, 31, 747–759. [Google Scholar] [CrossRef] [PubMed]
- Marikawa, Y. Toward Better Assessments of Developmental Toxicity using Stem Cell-based in Vitro Embryogenesis Models. Birth Defects Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency. ICH S5 (R3) Guideline on Reproductive Toxicology: Detection of Toxicity to Reproduction for Human Pharmaceuticals; European Medicines Agency: Amsterdam, The Netherlands, 2020.
- Mennen, R.; Oldenburger, M.; Piersma, A. Endoderm and Mesoderm Derivatives in Embryonic Stem Cell Differentiation and their use in Developmental Toxicity Testing. Reprod. Toxicol. 2022, 107, 44–59. [Google Scholar] [CrossRef]
- Zhang, S.; Wernig, M.; Duncan, I.D.; Brüstle, O.; Thomson, J.A. In Vitro Differentiation of Transplantable Neural Precursors from Human Embryonic Stem Cells. Nat. Biotechnol. 2001, 19, 1129–1133. [Google Scholar] [CrossRef] [PubMed]
- Rice, D.; Barone, S., Jr. Critical Periods of Vulnerability for the Developing Nervous System: Evidence from Humans and Animal Models. Environ. Health Perspect. 2000, 108, 511–533. [Google Scholar]
- Pașca, S.P. The Rise of Three-Dimensional Human Brain Cultures. Nature 2018, 553, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Chesnut, M.; Hartung, T.; Hogberg, H.; Pamies, D. Human Oligodendrocytes and Myelin in Vitro to Evaluate Developmental Neurotoxicity. Int. J. Mol. Sci. 2021, 22, 7929. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Xu, R.; Padmashri, R.; Dunaevsky, A.; Liu, Y.; Dreyfus, C.F.; Jiang, P. Pluripotent Stem Cell-Derived Cerebral Organoids Reveal Human Oligodendrogenesis with Dorsal and Ventral Origins. Stem Cell Rep. 2019, 12, 890–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madhavan, M.; Nevin, Z.S.; Shick, H.E.; Garrison, E.; Clarkson-Paredes, C.; Karl, M.; Clayton, B.L.; Factor, D.C.; Allan, K.C.; Barbar, L. Induction of Myelinating Oligodendrocytes in Human Cortical Spheroids. Nat. Methods 2018, 15, 700–706. [Google Scholar] [CrossRef]
- Marton, R.M.; Miura, Y.; Sloan, S.A.; Li, Q.; Revah, O.; Levy, R.J.; Huguenard, J.R.; Pașca, S.P. Differentiation and Maturation of Oligodendrocytes in Human Three-Dimensional Neural Cultures. Nat. Neurosci. 2019, 22, 484–491. [Google Scholar] [CrossRef]
- James, O.G.; Selvaraj, B.T.; Magnani, D.; Burr, K.; Connick, P.; Barton, S.K.; Vasistha, N.A.; Hampton, D.W.; Story, D.; Smigiel, R. iPSC-Derived Myelinoids to Study Myelin Biology of Humans. Dev. Cell 2021, 56, 1346–1358.e6. [Google Scholar] [CrossRef] [PubMed]
- Sandström, J.; Eggermann, E.; Charvet, I.; Roux, A.; Toni, N.; Greggio, C.; Broyer, A.; Monnet-Tschudi, F.; Stoppini, L. Development and Characterization of a Human Embryonic Stem Cell-Derived 3D Neural Tissue Model for Neurotoxicity Testing. Toxicol. In Vitro 2017, 38, 124–135. [Google Scholar] [CrossRef]
- Pamies, D.; Barreras, P.; Block, K.; Makri, G.; Kumar, A.; Wiersma, D.; Smirnova, L.; Zang, C.; Bressler, J.; Christian, K.M.; et al. A Human Brain Microphysiological System Derived from Induced Pluripotent Stem Cells to Study Neurological Diseases and Toxicity. ALTEX-Altern. Anim. Exp. 2017, 34, 362–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pamies, D.; Block, K.; Lau, P.; Gribaldo, L.; Pardo, C.A.; Barreras, P.; Smirnova, L.; Wiersma, D.; Zhao, L.; Harris, G. Rotenone Exerts Developmental Neurotoxicity in a Human Brain Spheroid Model. Toxicol. Appl. Pharmacol. 2018, 354, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Harris, G.; Smirnova, L.; Zufferey, V.; Baldino Russo, F.; Braga, B.B.; Cristina, P.; Chestnut, M.; Zurich, M.; Hogberg, H.T.; et al. Antidepressant Paroxetine Exerts Developmental Neurotoxicity in an iPSC-Derived 3D Human Brain Model. Front. Cell. Neurosci. 2020, 14, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chesnut, M.; Paschoud, H.; Repond, C.; Smirnova, L.; Hartung, T.; Zurich, M.; Hogberg, H.T.; Pamies, D. Human Ipsc-Derived Model to Study Myelin Disruption. Int. J. Mol. Sci. 2021, 22, 9473. [Google Scholar] [CrossRef] [PubMed]
- Pamies, D.; Wiersma, D.; Katt, M.E.; Zhao, L.; Burtscher, J.; Harris, G.; Smirnova, L.; Searson, P.C.; Hartung, T.; Hogberg, H.T. Human IPSC 3D Brain Model as a Tool to Study Chemical-Induced Dopaminergic Neuronal Toxicity. Neurobiol. Dis. 2022, 169, 105719. [Google Scholar] [CrossRef] [PubMed]
- Lauschke, K.; Rosenmai, A.K.; Meiser, I.; Neubauer, J.C.; Schmidt, K.; Rasmussen, M.A.; Holst, B.; Taxvig, C.; Emnéus, J.K.; Vinggaard, A.M. A Novel Human Pluripotent Stem Cell-Based Assay to Predict Developmental Toxicity. Arch. Toxicol. 2020, 94, 3831–3846. [Google Scholar] [CrossRef] [PubMed]
- Marx-Stoelting, P.; de LM Solano, M.; Aoyama, H.; Adams, R.H.; Bal-Price, A.; Buschmann, J.; Chahoud, I.; Clark, R.; Fang, T.; Fujiwara, M. 25th Anniversary of the Berlin Workshop on Developmental Toxicology: DevTox Database Update, Challenges in Risk Assessment of Developmental Neurotoxicity and Alternative Methodologies in Bone Development and Growth. Reprod. Toxicol. 2021, 100, 155–162. [Google Scholar] [CrossRef]
- Kang, H.Y.; Choi, Y.; Jo, N.R.; Lee, J.; Ahn, C.; Ahn, I.Y.; Kim, T.S.; Kim, K.; Choi, K.; Lee, J.K. Advanced Developmental Toxicity Test Method Based on Embryoid Body’s Area. Reprod. Toxicol. 2017, 72, 74–85. [Google Scholar] [CrossRef]
- Easley, C.A. Induced pluripotent stem cells (iPSCs) in developmental toxicology. In Developmental Toxicology; Harris, C., Hansen, J.M., Eds.; Humana Press: Totowa, NJ, USA, 2019; pp. 19–34. [Google Scholar]
- Buser, M.; Pohl, H.; Abadin, H. Windows of Sensitivity to Toxic Chemicals in the Development of the Endocrine System: An Analysis of ATSDR’s Toxicological Profile Database. Int. J. Environ. Health Res. 2022, 32, 437–454. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Yin, N.; Yang, R.; Liang, S.; Liang, S.; Faiola, F. Silver Nanoparticles (AgNPs) and AgNO3 Perturb the Specification of Human Hepatocyte-Like Cells and Cardiomyocytes. Sci. Total Environ. 2020, 725, 138433. [Google Scholar] [CrossRef] [PubMed]
- Teino, I.; Matvere, A.; Pook, M.; Varik, I.; Pajusaar, L.; Uudeküll, K.; Vaher, H.; Trei, A.; Kristjuhan, A.; Org, T. Impact of AHR Ligand TCDD on Human Embryonic Stem Cells and Early Differentiation. Int. J. Mol. Sci. 2020, 21, 9052. [Google Scholar] [CrossRef]
- Fu, H.; Wang, L.; Wang, J.; Bennett, B.D.; Li, J.; Zhao, B.; Hu, G. Dioxin and AHR Impairs Mesoderm Gene Expression and Cardiac Differentiation in Human Embryonic Stem Cells. Sci. Total Environ. 2019, 651, 1038–1046. [Google Scholar] [CrossRef]
- Feutz, A.; De Geyter, C. Accuracy, Discriminative Properties and Reliability of a Human ESC-Based in Vitro Toxicity Assay to Distinguish Teratogens Responsible for Neural Tube Defects. Arch. Toxicol. 2019, 93, 2375–2384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, R.; Mamidi, M.K.; Kumar Das, A.; Bhonde, R. Human Embryonic Stem Cell Proliferation and Differentiation as Parameters to Evaluate Developmental Toxicity. J. Cell. Physiol. 2011, 226, 1583–1595. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Yamazaki, D.; Kanda, Y. 5-Fluorouracil Inhibits Neural Differentiation Via Mfn1/2 Reduction in Human Induced Pluripotent Stem Cells. J. Toxicol. Sci. 2018, 43, 727–734. [Google Scholar] [CrossRef] [Green Version]
- Davidsen, N.; Rosenmai, A.K.; Lauschke, K.; Svingen, T.; Vinggaard, A.M. Developmental Effects of PFOS, PFOA and GenX in a 3D Human Induced Pluripotent Stem Cell Differentiation Model. Chemosphere 2021, 279, 130624. [Google Scholar] [CrossRef] [PubMed]
- Guarnieri, G.; Becatti, M.; Comeglio, P.; Vignozzi, L.; Maggi, M.; Vannelli, G.B.; Morelli, A. Benzo[a]Pyrene Impairs the Migratory Pattern of Human Gonadotropin-Releasing-Hormone-Secreting Neuroblasts. Eur. J. Histochem. 2021, 65, 3282. [Google Scholar] [CrossRef] [PubMed]
- Tait, S.; La Rocca, C.; Mantovani, A. Exposure of Human Fetal Penile Cells to Different PCB Mixtures: Transcriptome Analysis Points to Diverse Modes of Interference on External Genitalia Programming. Reprod. Toxicol. 2011, 32, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, T.; Gassmann, K.; Gotz, C.; Hubenthal, U.; Moors, M.; Krause, G.; Merk, H.F.; Nguyen, N.H.; Scanlan, T.S.; Abel, J.; et al. Polybrominated Diphenyl Ethers Induce Developmental Neurotoxicity in a Human in Vitro Model: Evidence for Endocrine Disruption. Environ. Health Perspect. 2010, 118, 572–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barni, T.; Maggi, M.; Fantoni, G.; Granchi, S.; Mancina, R.; Gulisano, M.; Marra, F.; Macorsini, E.; Luconi, M.; Rotella, C. Sex Steroids and Odorants Modulate Gonadotropin-Releasing Hormone Secretion in Primary Cultures of Human Olfactory Cells. J. Clin. Endocrinol. Metab. 1999, 84, 4266–4273. [Google Scholar] [CrossRef]
- Romanelli, R.G.; Barni, T.; Maggi, M.; Luconi, M.; Failli, P.; Pezzatini, A.; Pelo, E.; Torricelli, F.; Crescioli, C.; Ferruzzi, P.; et al. Expression and Function of Gonadotropin-Releasing Hormone (GnRH) Receptor in Human Olfactory GnRH-Secreting Neurons: An Autocrine GnRH Loop Underlies Neuronal Migration. J. Biol. Chem. 2004, 279, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Romanelli, R.G.; Barni, T.; Maggi, M.; Luconi, M.; Failli, P.; Pezzatini, A.; Morelli, A.; Maggi, R.; Zaninetti, R.; Salerno, R. Role of Endothelin-1 in the Migration of Human Olfactory Gonadotropin-Releasing Hormone-Secreting Neuroblasts. Endocrinology 2005, 146, 4321–4330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morelli, A.; Marini, M.; Mancina, R.; Luconi, M.; Vignozzi, L.; Fibbi, B.; Filippi, S.; Pezzatini, A.; Forti, G.; Vannelli, G.B. Sex Steroids and Leptin Regulate the “first Kiss”(KiSS 1/G-Protein-Coupled Receptor 54 System) in Human Gonadotropin-Releasing-Hormone-Secreting Neuroblasts. J. Sex. Med. 2008, 5, 1097–1113. [Google Scholar] [CrossRef] [PubMed]
- Sarchielli, E.; Comeglio, P.; Squecco, R.; Ballerini, L.; Mello, T.; Guarnieri, G.; Idrizaj, E.; Mazzanti, B.; Vignozzi, L.; Gallina, P. Tumor Necrosis Factor-A Impairs Kisspeptin Signaling in Human Gonadotropin-Releasing Hormone Primary Neurons. J. Clin. Endocrinol. Metab. 2017, 102, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Benvenuti, S.; Luciani, P.; Vannelli, G.B.; Gelmini, S.; Franceschi, E.; Serio, M.; Peri, A. Estrogen and Selective Estrogen Receptor Modulators Exert Neuroprotective Effects and Stimulate the Expression of Selective Alzheimer’s Disease Indicator-1, a Recently Discovered Antiapoptotic Gene, in Human Neuroblast Long-Term Cell Cultures. J. Clin. Endocrinol. Metab. 2005, 90, 1775–1782. [Google Scholar] [CrossRef] [Green Version]
- Benvenuti, S.; Luciani, P.; Cellai, I.; Deledda, C.; Baglioni, S.; Saccardi, R.; Urbani, S.; Francini, F.; Squecco, R.; Giuliani, C. Thyroid Hormones Promote Cell Differentiation and Up-Regulate the Expression of the Seladin-1 Gene in in Vitro Models of Human Neuronal Precursors. J. Endocrinol. 2008, 197, 437. [Google Scholar] [CrossRef] [Green Version]
- Morelli, A.; Fibbi, B.; Marini, M.; Silvestrini, E.; De Vita, G.; Chavalmane, A.K.; Vignozzi, L.; Filippi, S.; Forti, G.; Vannelli, G.B. Dihydrotestosterone and Leptin Regulate Gonadotropin-Releasing Hormone (GnRH) Expression and Secretion in Human GnRH-Secreting Neuroblasts. J. Sex. Med. 2009, 6, 397–407. [Google Scholar] [CrossRef]
- Coppi, E.; Cherchi, F.; Sarchielli, E.; Fusco, I.; Guarnieri, G.; Gallina, P.; Corradetti, R.; Pedata, F.; Vannelli, G.B.; Pugliese, A.M. Acetylcholine Modulates K and Na Currents in Human Basal Forebrain Cholinergic Neuroblasts through an Autocrine/Paracrine Mechanism. J. Neurochem. 2021, 157, 1182–1195. [Google Scholar] [CrossRef]
- Rapizzi, E.; Benvenuti, S.; Deledda, C.; Martinelli, S.; Sarchielli, E.; Fibbi, B.; Luciani, P.; Mazzanti, B.; Pantaleo, M.; Marroncini, G. A Unique Neuroendocrine Cell Model Derived from the Human Foetal Neural Crest. J. Endocrinol. Investig. 2020, 43, 1259–1269. [Google Scholar] [CrossRef]
- Sottili, M.; Filardi, T.; Cantini, G.; Cosmi, L.; Morano, S.; Luconi, M.; Lenzi, A.; Crescioli, C. Human Cell-Based Anti-Inflammatory Effects of Rosiglitazone. J. Endocrinol. Investig. 2022, 45, 105–114. [Google Scholar] [CrossRef]
- Crescioli, C.; Sottili, M.; Bonini, P.; Cosmi, L.; Chiarugi, P.; Romagnani, P.; Vannelli, G.B.; Colletti, M.; Isidori, A.M.; Serio, M. Inflammatory Response in Human Skeletal Muscle Cells: CXCL10 as a Potential Therapeutic Target. Eur. J. Cell Biol. 2012, 91, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Crescioli, C.; Maggi, M.; Vannelli, G.; Ferruzzi, P.; Granchi, S.; Mancina, R.; Muratori, M.; Forti, G.; Serio, M.; Luconi, M. Expression of Functional Estrogen Receptors in Human Fetal Male External Genitalia. J. Clin. Endocrinol. Metab. 2003, 88, 1815–1824. [Google Scholar] [CrossRef] [Green Version]
- Di Franco, A.; Guasti, D.; Squecco, R.; Mazzanti, B.; Rossi, F.; Idrizaj, E.; Gallego-Escuredo, J.M.; Villarroya, F.; Bani, D.; Forti, G. Searching for Classical Brown Fat in Humans: Development of a Novel Human Fetal Brown Stem Cell Model. Stem Cells 2016, 34, 1679–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poli, G.; Sarchielli, E.; Guasti, D.; Benvenuti, S.; Ballerini, L.; Mazzanti, B.; Armignacco, R.; Cantini, G.; Lulli, M.; Chortis, V. Human Fetal Adrenal Cells Retain Age-related Stem-and Endocrine-differentiation Potential in Culture. FASEB J. 2019, 33, 2263–2277. [Google Scholar] [CrossRef] [PubMed]
- Melau, C.; Nielsen, J.E.; Perlman, S.; Lundvall, L.; Langhoff Thuesen, L.; Juul Hare, K.; Schou Hammerum, M.; Frederiksen, H.; Mitchell, R.T.; Juul, A. Establishment of a Novel Human Fetal Adrenal Culture Model that Supports De Novo and Manipulated Steroidogenesis. J. Clin. Endocrinol. Metab. 2021, 106, 843–857. [Google Scholar] [CrossRef] [PubMed]
- Melau, C.; Riis, M.L.; Nielsen, J.E.; Perlman, S.; Lundvall, L.; Thuesen, L.L.; Hare, K.J.; Hammerum, M.S.; Mitchell, R.T.; Frederiksen, H. The Effects of Selected Inhibitors on Human Fetal Adrenal Steroidogenesis Differs Under Basal and ACTH-Stimulated Conditions. BMC Med. 2021, 19, 1–17. [Google Scholar] [CrossRef]
- Harpelunde Poulsen, K.; Nielsen, J.; Frederiksen, H.; Melau, C.; Juul Hare, K.; Langhoff Thuesen, L.; Perlman, S.; Lundvall, L.; Mitchell, R.; Juul, A. Dysregulation of FGFR Signalling by a Selective Inhibitor Reduces Germ Cell Survival in Human Fetal Gonads of both Sexes and Alters the Somatic Niche in Fetal Testes. Hum. Reprod. 2019, 34, 2228–2243. [Google Scholar] [CrossRef]
- Scialli, A.R.; Daston, G.; Chen, C.; Coder, P.S.; Euling, S.Y.; Foreman, J.; Hoberman, A.M.; Hui, J.; Knudsen, T.; Makris, S.L. Rethinking Developmental Toxicity Testing: Evolution or Revolution? Birth Defects Res. 2018, 110, 840–850. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Levy, M.Z.; Allsopp, R.C.; Futcher, A.B.; Greider, C.W.; Harley, C.B. Telomere End-Replication Problem and Cell Aging. J. Mol. Biol. 1992, 225, 951–960. [Google Scholar] [CrossRef]
- Ishii, T.; Eto, K. Fetal Stem Cell Transplantation: Past, Present, and Future. World J. Stem Cells 2014, 6, 404–420. [Google Scholar] [CrossRef]
- Todaro, G.J.; Green, H. Quantitative Studies of the Growth of Mouse Embryo Cells in Culture and their Development into Established Lines. J. Cell Biol. 1963, 17, 299–313. [Google Scholar] [CrossRef]
- Pipas, J.M. SV40: Cell Transformation and Tumorigenesis. Virology 2009, 384, 294–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Dakic, A.; Chen, R.; Disbrow, G.L.; Zhang, Y.; Dai, Y.; Schlegel, R. Cell-Restricted Immortalization by Human Papillomavirus Correlates with Telomerase Activation and Engagement of the hTERT Promoter by Myc. J. Virol. 2008, 82, 11568–11576. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.D.; Berk, A.J. Adenovirus Proteins from both E1B Reading Frames are Required for Transformation of Rodent Cells by Viral Infection and DNA Transfection. Virology 1987, 156, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, A.G.; Ouellette, M.; Frolkis, M.; Holt, S.E.; Chiu, C.; Morin, G.B.; Harley, C.B.; Shay, J.W.; Lichtsteiner, S.; Wright, W.E. Extension of Life-Span by Introduction of Telomerase into Normal Human Cells. Science 1998, 279, 349–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Nagavarapu, U.; Relloma, K.; Sjaastad, M.D.; Moss, W.C.; Passaniti, A.; Herron, G.S. Telomerized Human Microvasculature is Functional in Vivo. Nat. Biotechnol. 2001, 19, 219–224. [Google Scholar] [CrossRef]
- Simonsen, J.L.; Rosada, C.; Serakinci, N.; Justesen, J.; Stenderup, K.; Rattan, S.I.; Jensen, T.G.; Kassem, M. Telomerase Expression Extends the Proliferative Life-Span and Maintains the Osteogenic Potential of Human Bone Marrow Stromal Cells. Nat. Biotechnol. 2002, 20, 592–596. [Google Scholar] [CrossRef] [PubMed]
- Deurholt, T.; van Til, N.P.; Chhatta, A.A.; ten Bloemendaal, L.; Schwartlander, R.; Payne, C.; Plevris, J.N.; Sauer, I.M.; Chamuleau, R.A.; Elferink, R.P. Novel Immortalized Human Fetal Liver Cell Line, cBAL111, has the Potential to Differentiate into Functional Hepatocytes. BMC Biotechnol. 2009, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, N.; Fujiwara, T.; Westerman, K.A.; Inoue, Y.; Sakaguchi, M.; Noguchi, H.; Miyazaki, M.; Cai, J.; Tanaka, N.; Fox, I.J. Prevention of Acute Liver Failure in Rats with Reversibly Immortalized Human Hepatocytes. Science 2000, 287, 1258–1262. [Google Scholar] [CrossRef] [PubMed]
- Tátrai, P.; Szepesi, Á.; Matula, Z.; Szigeti, A.; Buchan, G.; Mádi, A.; Uher, F.; Német, K. Combined Introduction of Bmi-1 and hTERT Immortalizes Human Adipose Tissue-Derived Stromal Cells with Low Risk of Transformation. Biochem. Biophys. Res. Commun. 2012, 422, 28–35. [Google Scholar] [CrossRef]
- Blasi, E.; Radzioch, D.; Durum, S.K.; Varesio, L. A Murine Macrophage Cell Line, Immortalized by V-raf and V-myc Oncogenes, Exhibits Normal Macrophage Functions. Eur. J. Immunol. 1987, 17, 1491–1498. [Google Scholar] [CrossRef]
- Alani, R.M.; Hasskarl, J.; Grace, M.; Hernandez, M.; Israel, M.A.; Münger, K. Immortalization of Primary Human Keratinocytes by the Helix-loop-helix Protein, Id-1. Proc. Natl. Acad. Sci. USA 1999, 96, 9637–9641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwakkenbos, M.J.; Diehl, S.A.; Yasuda, E.; Bakker, A.Q.; Van Geelen, C.M.; Lukens, M.V.; Van Bleek, G.M.; Widjojoatmodjo, M.N.; Bogers, W.M.; Mei, H. Generation of Stable Monoclonal Antibody-producing B Cell Receptor-positive Human Memory B Cells by Genetic Programming. Nat. Med. 2010, 16, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Lipps, C.; Klein, F.; Wahlicht, T.; Seiffert, V.; Butueva, M.; Zauers, J.; Truschel, T.; Luckner, M.; Köster, M.; MacLeod, R. Expansion of Functional Personalized Cells with Specific Transgene Combinations. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heiss, M.; Hellström, M.; Kalén, M.; May, T.; Weber, H.; Hecker, M.; Augustin, H.G.; Korff, T. Endothelial Cell Spheroids as a Versatile Tool to Study Angiogenesis in Vitro. FASEB J. 2015, 29, 3076–3084. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, A.; Kletting, S.; de Souza Carvalho-Wodarz, C.; Repnik, U.; Griffiths, G.; Fischer, U.; Meese, E.; Huwer, H.; Wirth, D.; May, T. Human Alveolar Epithelial Cells Expressing Tight Junctions to Model the Air-Blood Barrier. ALTEX-Altern. Anim. Exp. 2016. [Google Scholar] [CrossRef]
- Lodes, N.; Seidensticker, K.; Perniss, A.; Nietzer, S.; Oberwinkler, H.; May, T.; Walles, T.; Hebestreit, H.; Hackenberg, S.; Steinke, M. Investigation on Ciliary Functionality of Different Airway Epithelial Cell Lines in Three-Dimensional Cell Culture. Tissue Eng. Part A 2020, 26, 432–440. [Google Scholar] [CrossRef]
- López-Gálvez, R.; Fleurot, I.; Chamero, P.; Trapp, S.; Olivier, M.; Chevaleyre, C.; Barc, C.; Riou, M.; Rossignol, C.; Guillon, A. Airway Administration of Flagellin Regulates the Inflammatory Response to Pseudomonas Aeruginosa. Am. J. Respir. Cell Mol. Biol. 2021, 65, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Campo, F.M.; May, T.; Zauers, J.; Sañudo, C.; Delgado-Calle, J.; Arozamena, J.; Berciano, M.T.; Lafarga, M.; Riancho, J.A. Generation and Characterization of Two Immortalized Human Osteoblastic Cell Lines Useful for Epigenetic Studies. J. Bone Miner. Metab. 2017, 35, 150–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burk, J.; Holland, H.; Lauermann, A.F.; May, T.; Siedlaczek, P.; Charwat, V.; Kasper, C. Generation and Characterization of a Functional Human Adipose-derived Multipotent Mesenchymal Stromal Cell Line. Biotechnol. Bioeng. 2019, 116, 1417–1426. [Google Scholar] [CrossRef] [PubMed]
- Oymans, J.; van Keulen, L.; Vermeulen, G.M.; Wichgers Schreur, P.J.; Kortekaas, J. Shuni Virus Replicates at the Maternal-Fetal Interface of the Ovine and Human Placenta. Pathogens 2020, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Hopperstad, K.; Truschel, T.; Wahlicht, T.; Stewart, W.; Eicher, A.; May, T.; Deisenroth, C. Characterization of Novel Human Immortalized Thyroid Follicular Epithelial Cell Lines. Appl. In Vitro Toxicol. 2021, 7, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Inoue, H.; Wu, J.C.; Yamanaka, S. Induced Pluripotent Stem Cell Technology: A Decade of Progress. Nat. Rev. Drug Discov. 2017, 16, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Schutgens, F.; Clevers, H. Human Organoids: Tools for Understanding Biology and Treating Diseases. Annu. Rev. Pathol. Mech. Dis. 2020, 15, 211–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madden, J.C.; Enoch, S.J.; Paini, A.; Cronin, M.T. A Review of in Silico Tools as Alternatives to Animal Testing: Principles, Resources and Applications. Altern. Lab. Anim. 2020, 48, 146–172. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Tong, W.; Roberts, R.; Liu, Z.; Thakkar, S. DeepCarc: Deep Learning-Powered Carcinogenicity Prediction using Model-Level Representation. Front. Artif. Intell. 2021, 4, 757780. [Google Scholar] [CrossRef]
- Marzo, M.; Roncaglioni, A.; Kulkarni, S.; Barton-Maclaren, T.S.; Benfenati, E. In silico models for developmental toxicity. In In Silico Methods for Predicting Drug Toxicity; Benfenati, E., Ed.; Humana Press: New York, NY, USA, 2022; pp. 217–240. [Google Scholar]
- Cassano, A.; Manganaro, A.; Martin, T.; Young, D.; Piclin, N.; Pintore, M.; Bigoni, D.; Benfenati, E. CAESAR Models for Developmental Toxicity. Chem. Cent. J. 2010, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Fisher, J.; Naciff, J.; Laufersweiler, M.; Lester, C.; Daston, G.; Blackburn, K. Framework for Identifying Chemicals with Structural Features Associated with the Potential to Act as Developmental or Reproductive Toxicants. Chem. Res. Toxicol. 2013, 26, 1840–1861. [Google Scholar] [CrossRef] [PubMed]
- Matthews, E.J.; Kruhlak, N.L.; Benz, R.D.; Contrera, J.F. A Comprehensive Model for Reproductive and Developmental Toxicity Hazard Identification: I. Development of a Weight of Evidence QSAR Database. Regul. Toxicol. Pharmacol. 2007, 47, 115–135. [Google Scholar] [CrossRef] [PubMed]
- Matthews, E.J.; Kruhlak, N.L.; Benz, R.D.; Ivanov, J.; Klopman, G.; Contrera, J.F. A Comprehensive Model for Reproductive and Developmental Toxicity Hazard Identification: II. Construction of QSAR Models to Predict Activities of Untested Chemicals. Regul. Toxicol. Pharmacol. 2007, 47, 136–155. [Google Scholar] [CrossRef]
- Ciallella, H.L.; Russo, D.P.; Sharma, S.; Li, Y.; Sloter, E.; Sweet, L.; Huang, H.; Zhu, H. Predicting Prenatal Developmental Toxicity Based on the Combination of Chemical Structures and Biological Data. Environ. Sci. Technol. 2022, 56, 5984–5998. [Google Scholar] [CrossRef] [PubMed]
- Bolčič-Tavčar, M.; Vračko, M. Prediction of Mutagenicity, Carcinogenicity, Developmental Toxicity, and Skin Sensitisation with CAESAR Program for a Set of Conazoles. Arh. Hig. Rada. Toksikol. 2012, 63, 283–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzo, M.; Kulkarni, S.; Manganaro, A.; Roncaglioni, A.; Wu, S.; Barton-Maclaren, T.S.; Lester, C.; Benfenati, E. Integrating in Silico Models to Enhance Predictivity for Developmental Toxicity. Toxicology 2016, 370, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Manganelli, S.; Schilter, B.; Scholz, G.; Benfenati, E.; Lo Piparo, E. Value and Limitation of Structure-Based Profilers to Characterize Developmental and Reproductive Toxicity Potential. Arch. Toxicol. 2020, 94, 939–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contrera, J.F.; Kruhlak, N.L.; Matthews, E.J.; Benz, R.D. Comparison of MC4PC and MDL-QSAR Rodent Carcinogenicity Predictions and the Enhancement of Predictive Performance by Combining QSAR Models. Regul. Toxicol. Pharmacol. 2007, 49, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Marzo, M.; Roncaglioni, A.; Kulkarni, S.; Barton-Maclaren, T.S.; Benfenati, E. In silico model for developmental toxicity: How to use QSAR models and interpret their results. In In Silico Methods for Predicting Drug Toxicity; Benfenati, E., Ed.; Humana Press: New York, NY, USA, 2016; pp. 139–161. [Google Scholar]
- Vračko, M.; Stanojević, M.; Sollner Dolenc, M. Comparison of Predictions of Developmental Toxicity for Compounds of Solvent Data Set. SAR QSAR Environ. Res. 2022, 33, 35–48. [Google Scholar] [CrossRef]
- Kumar, K.K.; Devi, B.U.; Neeraja, P. Integration of in Silico Approaches to Determination of Endocrine-Disrupting Perfluorinated Chemicals Binding Potency with Steroidogenic Acute Regulatory Protein. Biochem. Biophys. Res. Commun. 2017, 491, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.K.; Devi, B.U.; Neeraja, P. Elucidation of Endocrine-Disrupting Polychlorinated Biphenyls Binding Potency with Steroidogenic Genes: Integration of in Silico Methods and Ensemble Docking Approaches. Ecotoxicol. Environ. Saf. 2018, 165, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.K.; Yugandhar, P.; Devi, B.U.; Kumar, T.S.; Savithramma, N.; Neeraja, P. Applications of in Silico Methods to Analyze the Toxicity and Estrogen Receptor-Mediated Properties of Plant-Derived Phytochemicals. Food Chem. Toxicol. 2019, 125, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, M.; Ellison, C.; Enoch, S.; Madden, J.; Cronin, M. Integrating (Q)SAR Models, Expert Systems and Read-Across Approaches for the Prediction of Developmental Toxicity. Reprod. Toxicol. 2010, 30, 147–160. [Google Scholar] [CrossRef]
- ECHA. The Use of Alternatives to Testing on Animals for the REACH Regulation; Fourth Report Under Article 117 (3) of the REACH Regulation; ECHA: Helsinki, Finland, 2020.
- Hewitt, M.; Madden, J.; Rowe, P.; Cronin, M. Structure-Based Modelling in Reproductive Toxicology: (Q)SARs for the Placental Barrier. SAR QSAR Environ. Res. 2007, 18, 57–76. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xia, Z.; Yan, L.; Liu, S. Prediction of Placental Barrier Permeability: A Model Based on Partial Least Squares Variable Selection Procedure. Molecules 2015, 20, 8270–8286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giaginis, C.; Zira, A.; Theocharis, S.; Tsantili-Kakoulidou, A. Application of Quantitative Structure-Activity Relationships for Modeling Drug and Chemical Transport Across the Human Placenta Barrier: A Multivariate Data Analysis Approach. J. Appl. Toxicol. 2009, 29, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Takaku, T.; Nagahori, H.; Sogame, Y.; Takagi, T. Quantitative Structure-Activity Relationship Model for the Fetal-Maternal Blood Concentration Ratio of Chemicals in Humans. Biol. Pharm. Bull. 2015, 38, 930–934. [Google Scholar] [CrossRef] [Green Version]
- Eguchi, A.; Hanazato, M.; Suzuki, N.; Matsuno, Y.; Todaka, E.; Mori, C. Maternal-Fetal Transfer Rates of PCBs, OCPs, PBDEs, and Dioxin-Like Compounds Predicted through Quantitative Structure-Activity Relationship Modeling. Environ. Sci. Pollut. Res. 2018, 25, 7212–7222. [Google Scholar] [CrossRef] [PubMed]
- Madden, J.C.; Pawar, G.; Cronin, M.T.; Webb, S.; Tan, Y.; Paini, A. In Silico Resources to Assist in the Development and Evaluation of Physiologically-Based Kinetic Models. Comput. Toxicol. 2019, 11, 33–49. [Google Scholar] [CrossRef]
- Boyce, M.; Meyer, B.; Grulke, C.; Lizarraga, L.; Patlewicz, G. Comparing the Performance and Coverage of Selected in Silico (Liver) Metabolism Tools Relative to Reported Studies in the Literature to Inform Analogue Selection in Read-Across: A Case Study. Comput. Toxicol. 2022, 21, 100208. [Google Scholar] [CrossRef] [PubMed]
- Leung, M.C.; Hutson, M.S.; Seifert, A.W.; Spencer, R.M.; Knudsen, T.B. Computational Modeling and Simulation of Genital Tubercle Development. Reprod. Toxicol. 2016, 64, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Hutson, M.S.; Leung, M.C.; Baker, N.C.; Spencer, R.M.; Knudsen, T.B. Computational Model of Secondary Palate Fusion and Disruption. Chem. Res. Toxicol. 2017, 30, 965–979. [Google Scholar] [CrossRef]
- OECD. Revised Guidance Document on Developing and Assessing Adverse Outcome Pathways. Series on Testing and Assessment; OECD: Paris, France, 2017; pp. 1–32. [Google Scholar]
- Sachana, M.; Munn, S.; Bal-Price, A. Adverse Outcome Pathway on Chronic Binding of Antagonist to N-Methyl-D-Aspartate Receptors (NMDARs) during Brain Development Induces Impairment of Learning and Memory Abilities; OECD: Paris, France, 2016. [Google Scholar]
- Tschudi-Monnet, F.; FitzGerald, R. Adverse Outcome Pathway on Chronic Binding of Antagonist to N-Methyl-D-Aspartate Receptors during Brain Development Leading to Neurodegeneration with Impairment in Learning and Memory in Aging; OECD: Paris, France, 2018. [Google Scholar]
- Rolaki, A.; Pistollato, F.; Munn, S.; Bal-Price, A. Adverse Outcome Pathway on Inhibition of Na/I-Symporter (NIS) Leads to Learning and Memory Impairment; OECD: Paris, France, 2019. [Google Scholar]
- Spinu, N.; Bal-Price, A.; Cronin, M.T.; Enoch, S.J.; Madden, J.C.; Worth, A.P. Development and Analysis of an Adverse Outcome Pathway Network for Human Neurotoxicity. Arch. Toxicol. 2019, 93, 2759–2772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crofton, K.M.; Gilbert, M.; Friedman, K.P.; Demeneix, B.; Marty, M.S.; Zoeller, R.T. Adverse Outcome Pathway on Inhibition of Thyroperoxidase and Subsequent Adverse Neurodevelopmental Outcomes in Mammals; OECD: Paris, France, 2019. [Google Scholar]
- Knudsen, T.B.; Kleinstreuer, N.C. Disruption of Embryonic Vascular Development in Predictive Toxicology. Birth Defects Res. Part C Embryo Today Rev. 2011, 93, 312–323. [Google Scholar] [CrossRef]
- Farhat, A.; Kennedy, S.W. Adverse Outcome Pathway on Aryl Hydrocarbon Receptor Activation Leading to Early Life Stage Mortality, Via Reduced VEGF; OECD: Paris, France, 2019. [Google Scholar]
- Ellis-Hutchings, R.G.; Settivari, R.S.; McCoy, A.T.; Kleinstreuer, N.; Franzosa, J.; Knudsen, T.B.; Carney, E.W. Embryonic Vascular Disruption Adverse Outcomes: Linking High Throughput Signaling Signatures with Functional Consequences. Reprod. Toxicol. 2017, 70, 82–96. [Google Scholar] [CrossRef]
- Webster, F.; Lambert, I.B.; Yauk, C.L. Adverse Outcome Pathway on Cyp2E1 Activation Leading to Liver Cancer; OECD: Paris, France, 2021. [Google Scholar]
- Schneider, M.R.; Oelgeschlaeger, M.; Burgdorf, T.; van Meer, P.; Theunissen, P.; Kienhuis, A.S.; Piersma, A.H.; Vandebriel, R.J. Applicability of Organ-on-Chip Systems in Toxicology and Pharmacology. Crit. Rev. Toxicol. 2021, 51, 540–554. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Romero, R.; Han, Y.M.; Kim, H.C.; Kim, C.J.; Hong, J.; Huh, D. Placenta-on-a-Chip: A Novel Platform to Study the Biology of the Human Placenta. J. Matern.-Fetal Neonatal Med. 2016, 29, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Blundell, C.; Yi, Y.; Ma, L.; Tess, E.R.; Farrell, M.J.; Georgescu, A.; Aleksunes, L.M.; Huh, D. Placental Drug Transport-on-a-chip: A Microengineered in Vitro Model of Transporter-mediated Drug Efflux in the Human Placental Barrier. Adv. Healthc. Mater. 2018, 7, 1700786. [Google Scholar] [CrossRef]
- Yin, F.; Zhu, Y.; Zhang, M.; Yu, H.; Chen, W.; Qin, J. A 3D Human Placenta-on-a-Chip Model to Probe Nanoparticle Exposure at the Placental Barrier. Toxicol. In Vitro 2019, 54, 105–113. [Google Scholar] [CrossRef]
- Richardson, L.; Kim, S.; Menon, R.; Han, A. Organ-on-Chip Technology: The Future of Feto-Maternal Interface Research? Front. Physiol. 2020, 11, 715. [Google Scholar] [CrossRef]
- Richardson, L.S.; Kammala, A.K.; Costantine, M.M.; Fortunato, S.J.; Radnaa, E.; Kim, S.; Taylor, R.N.; Han, A.; Menon, R. Testing of Drugs using Human Feto-Maternal Interface Organ-on-Chips Provide Insights into Pharmacokinetics and Efficacy. Lab Chip 2022, 22, 4574–4592. [Google Scholar] [CrossRef]
- Kim, S.; Richardson, L.; Radnaa, E.; Chen, Z.; Rusyn, I.; Menon, R.; Han, A. Molecular Mechanisms of Environmental Toxin Cadmium at the Feto-Maternal Interface Investigated using an Organ-on-Chip (FMi-OOC) Model. J. Hazard. Mater. 2022, 422, 126759. [Google Scholar] [CrossRef]
- Richardson, L.; Gnecco, J.; Ding, T.; Osteen, K.; Rogers, L.M.; Aronoff, D.M.; Menon, R. Fetal Membrane Organ-on-Chip: An Innovative Approach to Study Cellular Interactions. Reprod. Sci. 2020, 27, 1562–1569. [Google Scholar] [CrossRef]
- Tait, S.; Luconi, M.; Straccia, M.; Sogorb, M. A Proposal of an Innovative Microfluidic Platform for Testing Developmental Toxicity. Toxicol. Lett. 2021, 350, S143–S144. [Google Scholar] [CrossRef]
- Amadei, G.; Handford, C.E.; Qiu, C.; De Jonghe, J.; Greenfeld, H.; Tran, M.; Martin, B.K.; Chen, D.; Aguilera-Castrejon, A.; Hanna, J.H. Synthetic Embryos Complete Gastrulation to Neurulation and Organogenesis. Nature 2022, 610, 143–153. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission Regulation (EU) 2018/605 of 19 April 2018 Amending Annex II to Regulation (EC) no 1107/2009 by Setting Out Scientific Criteria for the Determination of Endocrine Disrupting Properties; European Union: Brussels, Belgium, 2018; pp. 33–36. [Google Scholar]



{kind=link}
{kind=link}
| Maternal Toxicity | Developmental Endpoints for Litters with Implants | Developmental Endpoints for Litters with Live Fetuses |
|---|---|---|
| Number of animals: at the start of the test, surviving, pregnant, aborting and delivering early | Number of corpora lutea | Number and percent of live offspring |
| Day of death during the study or whether animals survived to termination | Number of implantations | Sex ratio |
| Day of observation of each abnormal clinical sign and its subsequent course | Number and percent of live and dead fetuses and resorptions | Fetal body weight |
| Body weight, body weight change and gravid uterine weight, including body weight change corrected for gravid uterine weight | Number and percent of pre- and post-implantation losses | Anogenital distance of all rodent fetuses |
| Food and water consumption | External, soft tissue and skeletal malformations and other relevant alterations | |
| For rat dams: thyroid hormones T4, T3 and thyroid-stimulating hormone | Total number and percent of fetuses and litters with any external, soft tissue or skeletal alteration | |
| Thyroid histopathology assessment | ||
| Necropsy findings, including uterine weight | ||
| No Observed Adverse Effect Level (NOAEL) |
| Substance | Type of Placental Explant/Culture Period | Results | Ref. |
|---|---|---|---|
| BPA | Placental explant cultures | Increase of beta-hCG secretion and caspase-3 expression | [40] |
| Cadmium chloride | Term placenta explants, 24 h | Toxic effects | [41] |
| Cadmium chloride Mercury chloride | Term placenta explants, 24 h | Decrease in membrane fluidity and accumulation of cadmium in the membrane | [42] |
| DDT | Third-trimester human placental tissue explants, up to 72 h | Inhibition of CYP1A1 activity | [43] |
| Para-nonylphenol | First trimester chorionic villous explants, 24 h | Increase of cytokines at extremely low doses of p-NP | [44] |
| NAMs | PROs | CONs | |
|---|---|---|---|
| 1a | Differentiated embryonic stem cell (ESC) |
|
|
| 1b | Differentiated iPSCs | ||
| 2 | Fetal/embryo primary cell organ cultures |
|
|
| Toxicants | hESC-or ihPSC- Induced Lineage | Main Outcomes | Endpoints | Ref. |
|---|---|---|---|---|
| hESC-derived cardiomyocytes and hepatocytes | Perturbed specification of two endoderm and mesoderm primary germ layers but not of ectoderm | Decreased expression of liver and cardiac markers | [92] |
| hESC towards neural progenitor, early mesoderm and definitive endoderm cells | Lineage-specific regulation of Aryl Hydrocarbon Receptor (AhR) expression during human embryonic stem cell differentiation | Gene expression patterns of the lineage | [93] |
| hESC-derived cardiomyocytes | Inhibition of cardiomyocyte differentiation, mediated by AhR | Gene expression patterns of the lineage | [94] |
| hESC-derived neural precursors | Alterations of neurogenic differentiation of hESC | Neural tube-like structures (rosettes) Expression of specific neuronal markers | [95] |
| hESC and embryoid bodies | Downregulation of markers associated with stemness, cardiac mesoderm, hepatic endoderm and neuroectoderm, indicating abnormal differentiation | Cell adherence Morphology Viability Apoptosis Lineage-specific expression markers | [96] |
| iPSCs-derived neural precursors | Neurotoxicity via mitofusin-mediated mitochondria dynamics | Expression of neural differentiation marker genes Intracellular ATP content Mitochondrial fragmentation Mitochondrial fusion | [97] |
| BrainSpheres from hiPSC | Myelination process oligodendrocyte differentiation No toxicity | Gene expression analysis of myelination markers, cytotoxicity; myelin quantification | [85] |
| BrainSpheres from hiPSC combined with endothelial barrier cells | Dopaminergic neuron toxicity | Gene expression analysis; metabolomics; ROS production; cytotoxicity | [86] |
| hiPSC line BiONi010-C-derived cardiomyocytes hiPSC line IMR90-1-derived cardiomyocytes | Reduced cardiac differentiation | PluriBeat assay to evaluate cardiac differentiation and cardiac gene expression markers | [98] |
| Toxicants | Human Fetal Cells | Main Outcomes | Endpoints | Ref. |
| Human fetal olfactory epithelium neuroblasts | Impaired migration of gonadotropin-releasing hormone (GnRH) neurons | Gene expression of neuronal migration markers | [99] |
| Human fetal corpora cavernosa cells | Altered transcriptomic profiles | Toxicogenomics profiles Developmental pathways | [100] |
| Human fetal progenitor cells from 16 weeks of gestation commercially available (NPCs) | Migration, differentiation, gene expression | [101] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luconi, M.; Sogorb, M.A.; Markert, U.R.; Benfenati, E.; May, T.; Wolbank, S.; Roncaglioni, A.; Schmidt, A.; Straccia, M.; Tait, S. Human-Based New Approach Methodologies in Developmental Toxicity Testing: A Step Ahead from the State of the Art with a Feto–Placental Organ-on-Chip Platform. Int. J. Environ. Res. Public Health 2022, 19, 15828. https://doi.org/10.3390/ijerph192315828
Luconi M, Sogorb MA, Markert UR, Benfenati E, May T, Wolbank S, Roncaglioni A, Schmidt A, Straccia M, Tait S. Human-Based New Approach Methodologies in Developmental Toxicity Testing: A Step Ahead from the State of the Art with a Feto–Placental Organ-on-Chip Platform. International Journal of Environmental Research and Public Health. 2022; 19(23):15828. https://doi.org/10.3390/ijerph192315828
Chicago/Turabian StyleLuconi, Michaela, Miguel A. Sogorb, Udo R. Markert, Emilio Benfenati, Tobias May, Susanne Wolbank, Alessandra Roncaglioni, Astrid Schmidt, Marco Straccia, and Sabrina Tait. 2022. "Human-Based New Approach Methodologies in Developmental Toxicity Testing: A Step Ahead from the State of the Art with a Feto–Placental Organ-on-Chip Platform" International Journal of Environmental Research and Public Health 19, no. 23: 15828. https://doi.org/10.3390/ijerph192315828