
The Influence of HLA Alleles on the Affective Distress Profile
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Instruments
2.3. DNA Extraction
2.4. Identification of HLA Alleles
2.5. Statistical Analysis
3. Results
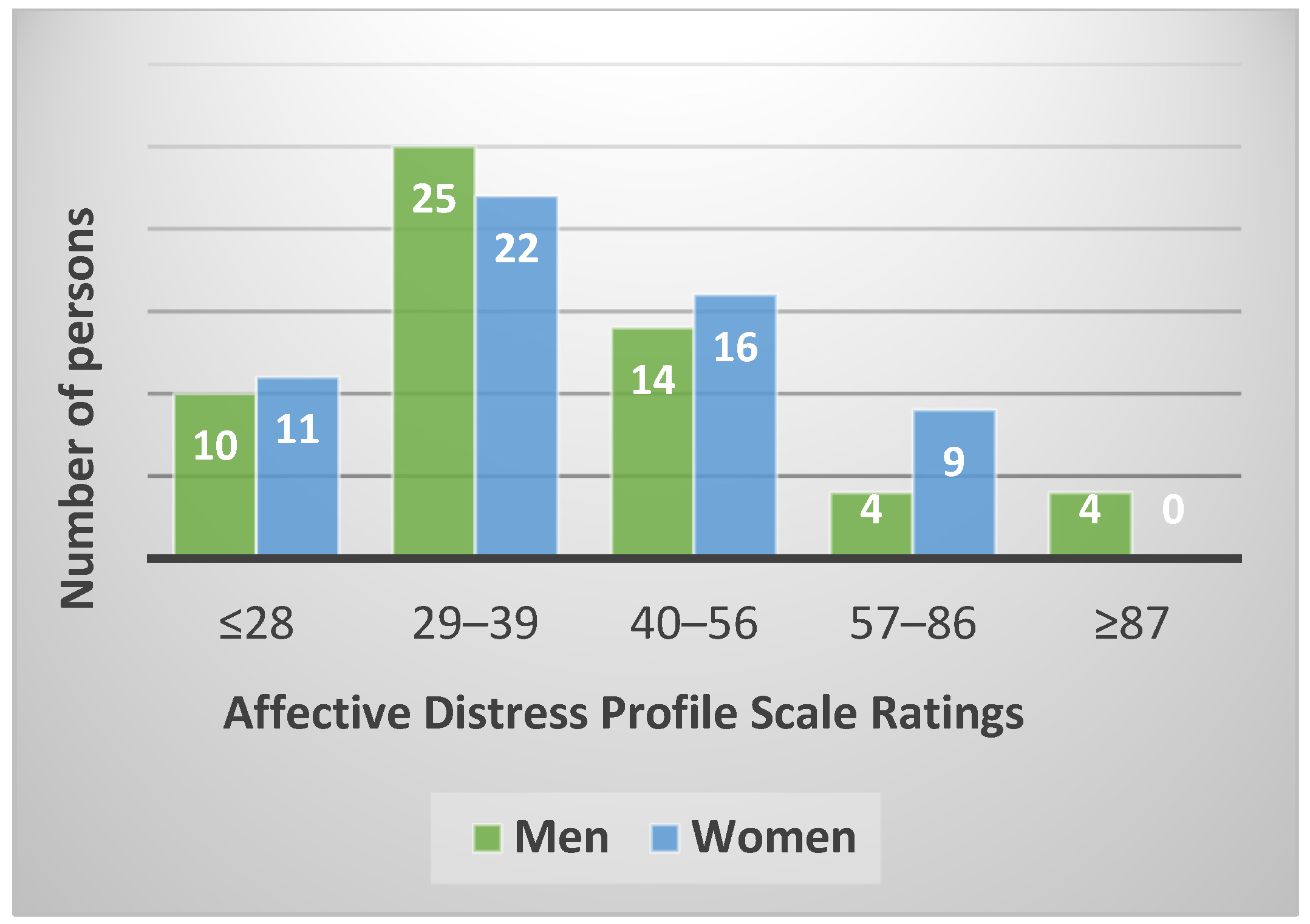
3.1. Affective Distress Profile (PDA)
3.2. Correlations between the HLA Profile and PDA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lazarus, R.S.; Folkman, S. Stress, Appraisal, and Coping; Springer: New York, NY, USA, 1984; pp. 268–286. [Google Scholar]
- Nash, M.W.; Sugden, K.; Huezo-Diaz, P.; Williamson, R.; Sterne, A.; Purcell, S.; Sham, P.C.; Craig, I.W. Association analysis of monoamine genes with measures of depression and anxiety in a selected community sample of siblings. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2005, 135B, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, R.; Keßler, M.S.; Bunck, M.; Murgatroyd, C.; Spengler, D.; Zimbelmann, M.; Nußbaumer, M.; Czibere, L.; Turck, C.W.; Singewald, N.; et al. Candidate genes of anxiety-related behavior in HAB/LAB rats and mice: Focus on vasopressin and glyoxalase-I. Neurosci. Biobehav. Rev. 2007, 31, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Boehm, S.L.; Reed, C.L.; McKinnon, C.S.; Phillips, T.J. Shared genes influence sensitivity to the effects of ethanol on locomotor and anxiety-like behaviors, and the stress axis. Psychopharmacology 2002, 161, 54–63. [Google Scholar] [CrossRef]
- Horton, R.; Wilming, L.; Rand, V.; Lovering, R.C.; Bruford, E.A.; Khodiyar, V.K.; Lush, M.J.; Povey, S.; Talbot, C.C., Jr.; Wright, M.W.; et al. Gene map of the extended human MHC. Nat. Rev. Genet. 2004, 5, 889–899. [Google Scholar] [CrossRef]
- Xie, T.; Rowen, L.; Aguado, B.; Ahearn, M.E.; Madan, A.; Qin, S.; Campbell, R.D.; Hood, L. Analysis of the gene-dense major histocompatibility complex class III region and its comparison to mouse. Genome Res. 2003, 13, 2621–2636. [Google Scholar] [CrossRef] [Green Version]
- Amarapurpar, D.N.; Patel, N.D.; Kankonkar, S.R. HLA class II genotyping in chronic hepatitis B infection. J. Assoc. Physicians India 2003, 51, 779–781. [Google Scholar]
- Thorsby, E. Invited anniversary review: HLA associated diseases. Hum. Immunol. 1997, 53, 1–11. [Google Scholar] [CrossRef]
- Matei, H.V.; Vică, M.L.; Siserman, C.V. Association between HLA class II alleles and hepatitis B virus infection in Transylvania, Romania. Immunol. Investig. 2018, 47, 735–744. [Google Scholar] [CrossRef]
- McCanlies, E.C.; Kreiss, K.; Andrew, M.; Weston, A. HLA-DPB1 and chronic beryllium disease: A huge review. Am. J. Epidemiol. 2003, 157, 388–398. [Google Scholar] [CrossRef] [Green Version]
- Du, R.; Litonjua, A.A.; Tantisira, K.G.; Lasky-Su, J.; Sunyaev, S.R.; Klanderman, B.J.; Celedón, J.C.; Avila, L.; Soto-Quiros, M.E.; Weiss, S.T. Genome-wide association study reveals class I MHC-restricted T cell-associated molecule gene (CRTAM) variants interact with vitamin D levels to affect asthma exacerbations. J. Allergy. Clin. Immunol. 2012, 129, 368–373.e5. [Google Scholar] [CrossRef] [Green Version]
- Matei, H.V.; Vică, M.L.; Ciucă, I.; Coman, H.G.; Nicula, G.Z.; Siserman, C.V. Correlations Among the HLA-DQB1 Alleles and Suicidal Behavior. J. Forensic Sci. 2020, 65, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Katrinli, S.; Lori, A.; Bradley, B.; Jovanovic, T.; Ressler, K.; Smith, A. F95. Effect of HLA alleles on suicide attempts and chronic pain in women. Biol. Psychiatry 2019, 85, S249–S250. [Google Scholar] [CrossRef]
- Hollar, D.W. Risk for intentional violent death associated with HLA genotypes: A preliminary survey of deceased American organ donors. Genetica 2009, 137, 253. [Google Scholar] [CrossRef] [PubMed]
- Corvin, A.; Morris, D.W. Genome-wide association studies: Findings at the major histocompatibility complex locus in psychosis. Biol. Psychiatry 2014, 75, 276–283. [Google Scholar] [CrossRef]
- Shimada-Sugimoto, M.; Otowa, T.; Miyagawa, T.; Khor, S.S.; Omae, Y.; Toyo-Oka, L.; Sugaya, N.; Kawamura, Y.; Umekage, T.; Miyashita, A.; et al. Polymorphisms in the TMEM132D region are associated with panic disorder in HLA-DRB1*13:02-negative individuals of a Japanese population. Hum. Genome Var. 2016, 3, 16001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldowitz, D.; Matthews, D.B.; Hamre, K.M.; Mittleman, G.; Chesler, E.J.; Becker, H.C.; Lopez, M.F.; Jones, S.R.; Mathews, T.A.; Miles, M.F.; et al. Progress in using mouse inbred strains, consomics, and mutants to identify genes related to stress, anxiety, and alcohol phenotypes. Alcohol. Clin. Exp. Res. 2006, 30, 1066–1078. [Google Scholar] [CrossRef]
- Middeldorp, C.M.; Slof-Op ‘t Landt, M.C.; Medland, S.E.; van Beijsterveldt, C.E.; Bartels, M.; Willemsen, G.; Hottenga, J.J.; de Geus, E.J.; Suchiman, H.E.; Dolan, C.V.; et al. Anxiety and depression in children and adults: Influence of serotonergic and neurotrophic genes? Genes Brain Behav. 2010, 9, 808–816. [Google Scholar] [CrossRef] [Green Version]
- Arias, B.; Aguilera, M.; Moya, J.; Sáiz, P.A.; Villa, H.; Ibáñez, M.I.; García-Portillo, M.P.; Bobes, J.; Ortet, G.; Fañanás, L. The role of genetic variability in the SLC6A4, BDNF and GABRA6 genes in anxiety-related traits. Acta Psychiatr. Scand. 2012, 125, 194–202. [Google Scholar] [CrossRef] [Green Version]
- Bourgeois, A.; LeUnes, A.; Meyers, M. Full-scale and short-form of the Profile of Mood States: A factor analytic comparison. J. Sport Behav. 2010, 33, 355–376. [Google Scholar]
- MathWorks. Available online: https://uk.mathworks.com/matlabcentral/answers/96572-how-can-i-perform-a-chi-square-test-to-determine-how-statistically-different-two-proportions-are-in (accessed on 9 August 2022).
- Perneger, T.V. What’s wrong with Bonferroni’s adjustment. BMJ 1998, 316, 1236. [Google Scholar] [CrossRef]
- Gluck, A. Metode Matematice în Industria Chimică; Tehică: Bucuresti, Romania, 1971; pp. 48–52. (In Romanian) [Google Scholar]
- Vică, M.L.; Matei, H.V.; Bondor, C.I.; Nicula, G.Z.; Siserman, C.V.; Loga, L.; Dican, L. HLA Polymorphisms and Haplotype Diversity in Transylvania, Romania. J. Immunol. Res. 2019, 2019, 1342762. [Google Scholar] [CrossRef] [PubMed]
- Scherrer, J.F.; True, W.R.; Xian, H.; Lyons, M.J.; Eisen, S.A.; Goldberg, J.; Lin, N.; Tsuang, M.T. Evidence for genetic influences common and specific to symptoms of generalized anxiety and panic. J. Affect. Disord. 2000, 57, 25–35. [Google Scholar] [CrossRef]
- Marks, I.M. Genetics of fear and anxiety disorders. Br. J. Psychiatry 1986, 149, 406–418. [Google Scholar] [CrossRef] [PubMed]
- Hettema, J.M.; Neale, M.C.; Kendler, K.S. A review and meta-analysis of the genetic epidemiology of anxiety disorders. Am. J. Psychiatry 2001, 158, 1568–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, R.; Zamyatin, V.; Klimenko, A.; Matushkin, Y.; Savostyanov, A.; Lashin, S. Reconstruction and Analysis of Gene Networks of Human Neurotransmitter Systems Reveal Genes with Contentious Manifestation for Anxiety, Depression, and Intellectual Disabilities. Genes 2019, 10, 699. [Google Scholar] [CrossRef] [Green Version]
- CONVERGE Consortium. Sparse whole-genome sequencing identifies two loci for major depressive disorder. Nature 2015, 523, 588–591. [Google Scholar] [CrossRef] [Green Version]
- Hyde, C.L.; Nagle, M.W.; Tian, C.; Chen, X.; Paciga, S.A.; Wendland, J.R.; Tung, J.Y.; Hinds, D.A.; Perlis, R.H.; Winslow, A.R. Identification of 15 genetic loci associated with risk of major depression in individuals of European descent. Nat. Genet. 2016, 48, 1031–1036. [Google Scholar] [CrossRef] [Green Version]
- Na, H.R.; Kang, E.H.; Lee, J.H.; Yu, B.H. The genetic basis of panic disorder. J. Korean Med. Sci. 2011, 26, 701–710. [Google Scholar] [CrossRef] [Green Version]
- Otowa, T.; Kawamura, Y.; Nishida, N.; Sugaya, N.; Koike, A.; Yoshida, E.; Inoue, K.; Yasuda, S.; Nishimura, Y.; Liu, X.; et al. Meta-analysis of genome-wide association studies for panic disorder in the Japanese population. Transl. Psychiatry 2012, 2, e186. [Google Scholar] [CrossRef] [Green Version]
- Shimada-Sugimoto, M.; Otowa, T.; Miyagawa, T.; Khor, S.S.; Kashiwase, K.; Sugaya, N.; Kawamura, Y.; Umekage, T.; Kojima, H.; Saji, H.; et al. Immune-related pathways including HLA-DRB1* 13:02 are associated with panic disorder. Brain Behav. Immun. 2015, 46, 96–103. [Google Scholar] [CrossRef]
- Andreassen, O.A.; Harbo, H.F.; Wang, Y.; Thompson, W.K.; Schork, A.J.; Mattingsdal, M.; Zuber, V.; Bettella, F.; Ripke, S.; Kelsoe, J.R.; et al. Genetic pleiotropy between multiple sclerosis and schizophrenia but not bipolar disorder: Differential involvement of immune-related gene loci. Mol. Psychiatry 2015, 20, 207–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Schizophrenia Consortium. Common polygenic variation contributes to risk of schizophrenia and bipolar disorder. Nature 2009, 460, 748–752. [Google Scholar] [CrossRef] [PubMed]
- Katrinli, S.; Lori, A.; Kilaru, V.; Carter, S.; Powers, A.; Gillespie, C.F.; Wingo, A.P.; Michopoulos, V.; Jovanovic, T.; Ressler, K.J.; et al. Association of HLA locus alleles with posttraumatic stress disorder. Brain Behav. Immun. 2019, 81, 655–658. [Google Scholar] [CrossRef] [PubMed]
- Sayeh, A.; Ben Cheikh, C.; Mardessi, A.; Mrad, M.; Nsiri, B.; Oumaya, A.; Fekih-Mrissa, N. HLA DRB1*03 as a possible common etiology of schizophrenia, Graves’ disease, and type 2 diabetes. Ann. Gen. Psychiatry 2017, 16, 7. [Google Scholar] [CrossRef] [Green Version]
- Debnath, M.; Busson, M.; Jamain, S.; Etain, B.; Hamdani, N.; Oliveira, J.; Boukouaci, W.; Amokrane, K.; Moins-Teisserenc, H.; Lajnef, M.; et al. The HLA-G low expressor genotype is associated with protection against bipolar disorder. Hum. Immunol. 2013, 74, 593–597. [Google Scholar] [CrossRef] [Green Version]
- Siserman, C.; Delcea, C.; Matei, H.V.; Vică, M.L. Major affective distress in testing forensic paternity. Rom. J. Leg. Med. 2019, 27, 292–296. [Google Scholar] [CrossRef]
- Michopoulos, V.; Rothbaum, A.O.; Jovanovic, T.; Almli, L.M.; Bradley, B.; Rothbaum, B.O.; Gillespie, C.F.; Ressler, K.J. Association of CRP genetic variation and CRP level with elevated PTSD symptoms and physiological responses in a civilian population with high levels of trauma. Am. J. Psychiatry 2015, 172, 353–362. [Google Scholar] [CrossRef]


{kind=link}
| HLA-A (n = 103) | Low PDA (n = 64) | Medium + High PDA (n = 39) | χ2 | p-Value | Adj α | ||
|---|---|---|---|---|---|---|---|
| n | % | n | % | ||||
| A*01 | 26 | 40.63 | 4 | 10.26 | 10.8265 | 0.0010 | 0.003333 |
| A*02 | 36 | 56.25 | 21 | 53.85 | 0.0567 | 0.8119 | |
| A*03 | 10 | 15.63 | 9 | 23.08 | 0.8945 | 0.3443 | |
| A*11 | 7 | 1094 | 2 | 5.13 | 1.0256 | 0.3112 | |
| A*23 | 6 | 9.38 | 2 | 5.13 | 0.6101 | 0.4348 | |
| A*24 | 9 | 14.06 | 3 | 7.69 | 0.9554 | 0.3284 | |
| A*25 | 6 | 9.38 | 1 | 2.56 | 1.7747 | 0.1828 | |
| A*26 | 9 | 14.06 | 5 | 12.82 | 0.0318 | 0.8584 | |
| A*29 | 4 | 6.25 | 6 | 15.38 | 2.3066 | 0.1288 | |
| A*30 | 6 | 9.38 | 0 | 0.00 | 3.8824 | 0.0488 | |
| A*31 | 1 | 1.56 | 2 | 5.13 | 1.0896 | 0.2966 | |
| A*32 | 4 | 6.25 | 8 | 20.51 | 4.7893 | 0.0286 | |
| A*33 | 1 | 1.56 | 2 | 5.13 | 1.0896 | 0.2966 | |
| A*68 | 4 | 6.25 | 3 | 7.69 | 0.0796 | 0.7779 | |
| A*69 | 0 | 0.00 | 1 | 2.56 | 1.6571 | 0.1980 | |
| HLA-B (n = 103) | Low PDA (n = 64) | Medium + High PDA (n = 39) | χ2 | p-Value | Adj α | ||
|---|---|---|---|---|---|---|---|
| n | % | n | % | ||||
| B*07 | 8 | 12.50 | 6 | 15.38 | 0.1717 | 0.6786 | 0.002173 |
| B*08 | 14 | 21.88 | 2 | 5.13 | 5.1797 | 0.0229 | |
| B*13 | 3 | 4.69 | 1 | 2.56 | 0.2927 | 0.5885 | |
| B*14 | 1 | 1.56 | 3 | 7.69 | 2.4394 | 0.1183 | |
| B*15 | 7 | 10.94 | 2 | 5.13 | 1.0256 | 0.3112 | |
| B*18 | 9 | 14.06 | 6 | 15.38 | 0.0340 | 0.8536 | |
| B*27 | 7 | 10.94 | 4 | 10.26 | 0.0118 | 0.9136 | |
| B*35 | 19 | 29.69 | 10 | 25.64 | 0.1962 | 0.6578 | |
| B*37 | 2 | 3.13 | 0 | 0.00 | 1.2429 | 0.2649 | |
| B*38 | 3 | 4.69 | 3 | 7.69 | 0.3988 | 0.5277 | |
| B*39 | 5 | 7.81 | 0 | 0.00 | 3.2023 | 0.0735 | |
| B*40 | 7 | 10.94 | 7 | 17.95 | 1.0143 | 0.3139 | |
| B*41 | 2 | 3.13 | 1 | 2.56 | 0.0270 | 0.8696 | |
| B*44 | 9 | 14.06 | 7 | 17.95 | 0.2789 | 0.5974 | |
| B*45 | 0 | 0.00 | 1 | 2.56 | 1.6571 | 0.1980 | |
| B*47 | 0 | 0.00 | 2 | 5.13 | 3.3470 | 0.0673 | |
| B*49 | 3 | 4.69 | 3 | 7.69 | 0.3988 | 0.5277 | |
| B*50 | 1 | 1.56 | 1 | 2.56 | 0.1277 | 0.7209 | |
| B*51 | 10 | 15.63 | 5 | 12.82 | 0.1532 | 0.6955 | |
| B*52 | 1 | 1.56 | 5 | 12.82 | 5.5986 | 0.0180 | |
| B*55 | 1 | 1.56 | 1 | 2.56 | 0.1277 | 0.7209 | |
| B*56 | 3 | 4.69 | 0 | 0.00 | 1.8830 | 0.1700 | |
| B*57 | 8 | 12.50 | 3 | 7.69 | 0.5872 | 0.4435 | |
| HLA-C (n = 103) | Low PDA (n = 64) | Medium + High PDA (n = 39) | χ2 | p-Value | Adj α | ||
|---|---|---|---|---|---|---|---|
| n | % | n | % | ||||
| C*01 | 9 | 14.06 | 3 | 7.69 | 0.9554 | 0.3284 | 0.003846 |
| C*02 | 7 | 10.94 | 7 | 17.95 | 1.0143 | 0.3139 | |
| C*03 | 6 | 9.38 | 5 | 12.82 | 0.3016 | 0.5829 | |
| C*04 | 20 | 31.25 | 9 | 23.08 | 0.8002 | 0.3710 | |
| C*05 | 1 | 1.56 | 3 | 7.69 | 2.4394 | 0.1183 | |
| C*06 | 19 | 29.69 | 6 | 15.38 | 2.6971 | 0.1005 | |
| C*07 | 27 | 42.19 | 11 | 28.21 | 2.0349 | 0.1537 | |
| C*08 | 3 | 4.69 | 3 | 7.69 | 0.3988 | 0.5277 | |
| C*12 | 11 | 17.19 | 15 | 38.46 | 5.8119 | 0.0159 | |
| C*14 | 0 | 0.00 | 1 | 2.56 | 1.6571 | 0.1980 | |
| C*15 | 8 | 12.50 | 3 | 7.69 | 0.5872 | 0.4435 | |
| C*16 | 3 | 4.69 | 2 | 5.13 | 0.0102 | 0.9196 | |
| C*17 | 1 | 1.56 | 1 | 2.56 | 0.1277 | 0.7209 | |
| HLA-DRB1 (n = 99) | Low PDA (n = 59) | Medium + High PDA (n = 40) | χ2 | p-Value | Adj α | ||
|---|---|---|---|---|---|---|---|
| n | % | n | % | ||||
| DRB1*01 | 12 | 20.34 | 7 | 17.50 | 0.1239 | 0.7249 | 0.004166 |
| DRB1*03 | 12 | 20.34 | 6 | 15.00 | 0.4568 | 0.4991 | |
| DRB1*04 | 15 | 25.42 | 11 | 27.50 | 0.0531 | 0.8178 | |
| DRB* 05 | 0 | 0.00 | 1 | 2.50 | 1.4901 | 0.2222 | |
| DRB1*07 | 8 | 13.56 | 8 | 20.00 | 0.7298 | 0.3929 | |
| DRB1*08 | 3 | 5.08 | 3 | 7.50 | 0.2443 | 0.6212 | |
| DRB1*10 | 1 | 1.69 | 1 | 2.50 | 0.0781 | 0.7799 | |
| DRB1*11 | 28 | 47.46 | 8 | 20.00 | 7.7666 | 0.0053 | |
| DRB1*13 | 9 | 15.25 | 6 | 15.00 | 0.0012 | 0.9724 | |
| DRB1*14 | 5 | 8.47 | 7 | 17.50 | 1.8230 | 0.1770 | |
| DRB1*15 | 6 | 10.17 | 6 | 15.00 | 0.5222 | 0.4699 | |
| DRB1*16 | 10 | 16.95 | 7 | 17.50 | 0.0051 | 0.9431 | |
| HLA-DQB1 (n = 99) | Low PDA (n = 59) | Medium + High PDA (n = 40) | χ2 | p-Value | Adj α | ||
|---|---|---|---|---|---|---|---|
| n | % | n | % | ||||
| DQB1*01 | 0 | 0.00 | 1 | 2.50 | 1.4901 | 0.2222 | |
| DQB1*02 | 16 | 27.12 | 11 | 27.50 | 0.0017 | 0.9667 | 0.00625 |
| DQB1*03 | 41 | 69.49 | 21 | 52.50 | 2.9405 | 0.0864 | |
| DQB1*04 | 3 | 5.08 | 3 | 7.50 | 0.2443 | 0.6212 | |
| DQB1*05 | 28 | 47.46 | 21 | 52.50 | 0.2425 | 0.6224 | |
| DQB1*06 | 12 | 20.34 | 13 | 32.50 | 1.8677 | 0.1717 | |
| DQB1*11 | 0 | 0.00 | 1 | 2.50 | 1.4901 | 0.2222 | |
| DQB1*15 | 1 | 1.69 | 0 | 0.00 | 0.6849 | 0.4079 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vică, M.L.; Delcea, C.; Dumitrel, G.A.; Vușcan, M.E.; Matei, H.V.; Teodoru, C.A.; Siserman, C.V. The Influence of HLA Alleles on the Affective Distress Profile. Int. J. Environ. Res. Public Health 2022, 19, 12608. https://doi.org/10.3390/ijerph191912608
Vică ML, Delcea C, Dumitrel GA, Vușcan ME, Matei HV, Teodoru CA, Siserman CV. The Influence of HLA Alleles on the Affective Distress Profile. International Journal of Environmental Research and Public Health. 2022; 19(19):12608. https://doi.org/10.3390/ijerph191912608
Chicago/Turabian StyleVică, Mihaela Laura, Cristian Delcea, Gabriela Alina Dumitrel, Mihaela Elvira Vușcan, Horea Vladi Matei, Cosmin Adrian Teodoru, and Costel Vasile Siserman. 2022. "The Influence of HLA Alleles on the Affective Distress Profile" International Journal of Environmental Research and Public Health 19, no. 19: 12608. https://doi.org/10.3390/ijerph191912608