
New Targets and New Technologies in the Treatment of Parkinson’s Disease: A Narrative Review


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
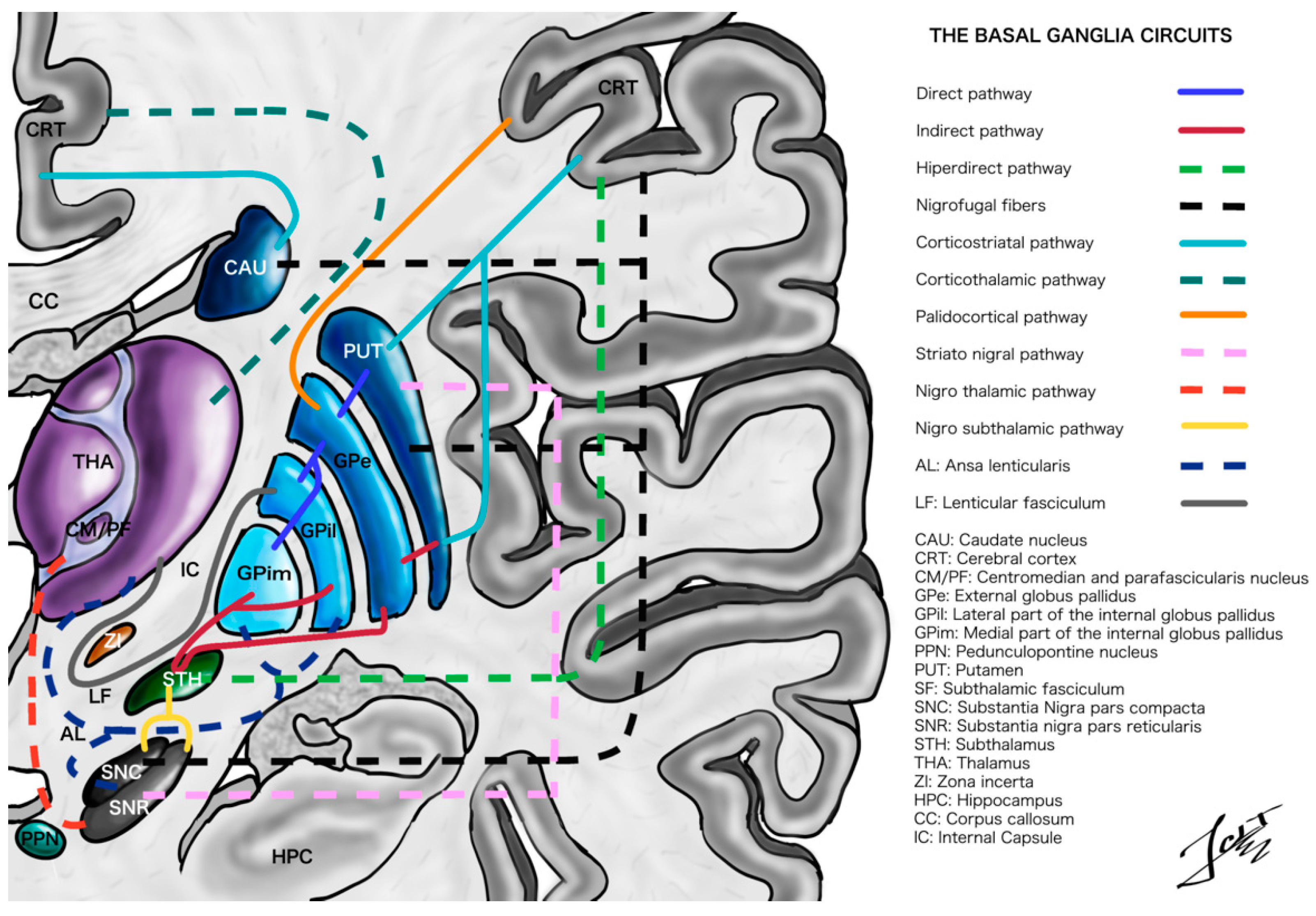
2. Overview of Basal Ganglia Functional Anatomy
3. New DBS Targets
3.1. Pedunculopontine Nucleus
3.2. Zona Incerta
3.3. Thalamic CM/Pf Complex
3.4. Substantia Nigra Pars Reticulata (SNr)
3.5. Multitarget
4. Non-Invasive and Minimally Invasive Treatment
4.1. Endovascular Approach
4.2. Non-Invasive Transcranial Stimulation
4.2.1. Transcranial Magnetic Stimulation (TMS)
4.2.2. Transcranial Current Stimulation—Transcranial Direct Current Stimulation (tDCS)
4.2.3. Transcranial Current Stimulation—Transcranial Stimulation with Alternating Current (tACS)
4.3. Non-Invasive Focused Ultrasound
5. Regenerative Medicine in Parkinson Disease
5.1. Gene, Cell, and Tissue Regenerative Therapies
5.2. Optogenetic Therapy
6. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Ascherio, A.; Schwarzschild, M.A. The epidemiology of Parkinson’s disease: Risk factors and prevention. Lancet Neurol. 2016, 15, 1257–1272. [Google Scholar] [CrossRef]
- Foffani, G.; Obeso, J.A. A Cortical Pathogenic Theory of Parkinson’s Disease. Neuron 2018, 99, 1116–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obeso, J.A.; Rodriguez-Oroz, M.C.; Goetz, C.G.; Marin, C.; Kordower, J.H.; Rodriguez, M.; Hirsch, E.C.; Farrer, M.; Schapira, A.H.; Halliday, G. Missing pieces in the Parkinson’s disease puzzle. Nat. Med. 2010, 16, 653–661. [Google Scholar] [CrossRef]
- Gelb, D.J.; Oliver, E.; Gilman, S. Diagnostic criteria for Parkinson disease. Arch. Neurol. 1999, 56, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.J.; Daniel, S.E.; Kilford, L.; Lees, A.J. Accuracy of clinical diagnosis of idiopathic Parkinson’s disease: A clinico-pathological study of 100 cases. J. Neurol. Neurosurg. Psychiatry 1992, 55, 181–184. [Google Scholar] [CrossRef] [Green Version]
- Langston, J.W.; Widner, H.; Goetz, C.G.; Brooks, D.; Fahn, S.; Freeman, T.; Watts, R. Core assessment program for intracerebral transplantations (CAPIT). Mov. Disord. 1992, 7, 2–13. [Google Scholar] [CrossRef]
- Ward, C.D.; Gibb, W.R. Research diagnostic criteria for Parkinson’s disease. Adv. Neurol. 1990, 53, 245–249. [Google Scholar]
- Postuma, R.B.; Berg, D.; Stern, M.; Poewe, W.; Olanow, C.W.; Oertel, W.; Obeso, J.; Marek, K.; Litvan, I.; Lang, A.E.; et al. MDS clinical diagnostic criteria for Parkinson’s disease. Mov. Disord. 2015, 30, 1591–1601. [Google Scholar] [CrossRef]
- Kalia, L.V.; Lang, A.E. Parkinson’s disease. Lancet. 2015, 386, 896–912. [Google Scholar] [CrossRef]
- Hornykiewicz, O. The discovery of dopamine deficiency in the parkinsonian brain. J. Neural. Transm. Suppl. 2006, 70, 9–15. [Google Scholar]
- Burbaud, P.; Gross, C.; Bioulac, B. Effect of subthalamic high frequency stimulation on substantia nigra pars reticulata and globus pallidus neurons in normal rats. J. Physiol. Paris 1994, 88, 359–361. [Google Scholar] [CrossRef]
- Filali, M.; Hutchison, W.D.; Palter, V.N.; Lozano, A.M.; Dostrovsky, J.O. Stimulation-induced inhibition of neuronal firing in human subthalamic nucleus. Exp. Brain Res. 2004, 156, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Welter, M.-L.; Houeto, J.-L.; Bonnet, A.-M.; Bejjani, P.-B.; Mesnage, V.; Dormont, D.; Navarro, S.; Cornu, P.; Agid, Y.; Pidoux, B. Effects of high-frequency stimulation on subthalamic neuronal activity in parkinsonian patients. Arch. Neurol. 2004, 61, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.H.; Luo, F.; Woodward, D.J.; Chang, J.Y. Basal ganglia neural responses during behaviorally effective deep brain stimulation of the subthalamic nucleus in rats performing a treadmill locomotion test. Synapse 2006, 59, 445–457. [Google Scholar] [CrossRef]
- Tai, C.-H.; Boraud, T.; Bezard, E.; Bioulac, B.; Gross, C.; Benazzouz, A. Electrophysiological and metabolic evidence that high-frequency stimulation of the subthalamic nucleus bridles neuronal activity in the subthalamic nucleus and the substantia nigra reticulata. FASEB J. 2003, 17, 1820–1830. [Google Scholar] [CrossRef] [Green Version]
- McIntyre, C.C.; Anderson, R.W. Deep brain stimulation mechanisms: The control of network activity via neurochemistry modulation. J. Neurochem. 2016, 139, 338–345. [Google Scholar] [CrossRef] [Green Version]
- Bosch, C.; Degos, B.; Deniau, J.M.; Venance, L. Subthalamic nucleus high-frequency stimulation generates a concomitant synaptic excitation–inhibition in substantia nigra pars reticulata. J. Physiol. 2011, 589, 4189–4207. [Google Scholar] [CrossRef]
- Johnson, M.D.; Miocinovic, S.; McIntyre, C.C.; Vitek, J.L. Mechanisms and targets of deep brain stimulation in movement disorders. Neurotherapeutics 2008, 5, 294–308. [Google Scholar] [CrossRef] [Green Version]
- Creed, M.; Pascoli, V.J.; Lüscher, C. Addiction therapy. Refining deep brain stimulation to emulate optogenetic treatment of synaptic pathology. Science 2015, 347, 659–664. [Google Scholar] [CrossRef]
- Hescham, S.; Temel, Y.; Schipper, S.; Lagiere, M.; Schönfeld, L.M.; Blokland, A.; Jahanshahi, A. Fornix deep brain stimulation induced long-term spatial memory independent of hippocampal neurogenesis. Brain Struct. Funct. 2017, 222, 1069–1075. [Google Scholar] [CrossRef] [Green Version]
- Ni, Z.; Kim, S.J.; Phielipp, N.; Ghosh, S.; Udupa, K.; Gunraj, C.A.; Saha, U.; Hodaie, M.; Kalia, S.K.; Lozano, A.M. Pallidal deep brain stimulation modulates cortical excitability and plasticity. Ann. Neurol. 2018, 83, 352–362. [Google Scholar] [CrossRef]
- Cheng, Y.; Xie, X.; Lu, J.; Gangal, H.; Wang, W.; Melo, S.; Wang, X.; Jerger, J.; Woodson, K.; Garr, E. Optogenetic induction of orbitostriatal long-term potentiation in the dorsomedial striatum elicits a persistent reduction of alcohol-seeking behavior in rats. Neuropharmacology 2021, 191, 108560. [Google Scholar] [CrossRef] [PubMed]
- Stefani, A.; Trendafilov, V.; Liguori, C.; Fedele, E.; Galati, S. Subthalamic nucleus deep brain stimulation on motor-symptoms of Parkinson’s disease: Focus on neurochemistry. Prog. Neurobiol. 2017, 151, 157–174. [Google Scholar] [CrossRef] [PubMed]
- Simonyan, K. Recent advances in understanding the role of the basal ganglia. F1000Research 2019, 8, F1000 Faculty Rev-122. [Google Scholar] [CrossRef]
- Parent, M.; Parent, A. The pallidofugal motor fiber system in primates. Parkinsonism Relat. Disord. 2004, 10, 203–211. [Google Scholar] [CrossRef]
- Obeso, J.A.; Stamelou, M.; Goetz, C.G.; Poewe, W.; Lang, A.E.; Weintraub, D.; Burn, D.; Halliday, G.M.; Bezard, E.; Przedborski, S.J.M.D. Past, present, and future of Parkinson’s disease: A special essay on the 200th Anniversary of the Shaking Palsy. Mov. Disord. 2017, 32, 1264–1310. [Google Scholar] [CrossRef] [PubMed]
- Trutti, A.C.; Mulder, M.J.; Hommel, B.; Forstmann, B.U. Functional neuroanatomical review of the ventral tegmental area. NeuroImage 2019, 191, 258–268. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Richard, S.; Parent, A. The organization of the striatal output system: A single-cell juxtacellular labeling study in the rat. Neurosci. Res. 2000, 38, 49–62. [Google Scholar] [CrossRef]
- Simonyan, K.; Cho, H.; Hamzehei Sichani, A.; Rubien-Thomas, E.; Hallett, M. The direct basal ganglia pathway is hyperfunctional in focal dystonia. Brain 2017, 140, 3179–3190. [Google Scholar] [CrossRef] [Green Version]
- Voorn, P.; Vanderschuren, L.J.; Groenewegen, H.J.; Robbins, T.W.; Pennartz, C.M. Putting a spin on the dorsal–ventral divide of the striatum. Trends Neurosci. 2004, 27, 468–474. [Google Scholar] [CrossRef]
- Grabli, D.; McCairn, K.; Hirsch, E.C.; Agid, Y.; Féger, J.; François, C.; Tremblay, L. Behavioural disorders induced by external globus pallidus dysfunction in primates: I. Behavioural study. Brain 2004, 127, 2039–2054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geibl, F.F.; Henrich, M.T.; Oertel, W.H. Mesencephalic and extramesencephalic dopaminergic systems in Parkinson’s disease. J. Neural. Transm. 2019, 126, 377–396. [Google Scholar] [CrossRef]
- Parent, A.; Cossette, M.; Lévesque, M. Anatomical Considerations in Basal Ganglia Surgery. In Movement Disorder Surgery; Karger Publishers: Basel, Switzerland, 2000; Volume 15, pp. 21–30. [Google Scholar]
- Sato, F.; Lavallée, P.; Lévesque, M.; Parent, A. Single-axon tracing study of neurons of the external segment of the globus pallidus in primate. J. Comp. Neurol. 2000, 417, 17–31. [Google Scholar] [CrossRef]
- Avecillas-Chasin, J.M.; Honey, C.R. Modulation of nigrofugal and pallidofugal pathways in deep brain stimulation for parkinson disease. Neurosurgery 2020, 86, E387–E397. [Google Scholar] [CrossRef] [PubMed]
- Saunders, A.; Oldenburg, I.A.; Berezovskii, V.K.; Johnson, C.A.; Kingery, N.D.; Elliott, H.L.; Xie, T.; Gerfen, C.R.; Sabatini, B.L. A direct GABAergic output from the basal ganglia to frontal cortex. Nature 2015, 521, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Nambu, A.; Tokuno, H.; Takada, M. Functional significance of the cortico–subthalamo–pallidal ‘hyperdirect’pathway. Neurosci. Res. 2002, 43, 111–117. [Google Scholar] [CrossRef]
- Apps, R.; Hawkes, R.; Aoki, S.; Bengtsson, F.; Brown, A.M.; Chen, G.; Ebner, T.J.; Isope, P.; Jörntell, H.; Lackey, E.P. Cerebellar modules and their role as operational cerebellar processing units. Cerebellum 2018, 17, 654–682. [Google Scholar] [CrossRef] [Green Version]
- Narain, D.; Remington, E.D.; De Zeeuw, C.I.; Jazayeri, M. A cerebellar mechanism for learning prior distributions of time intervals. Nat. Commun. 2018, 9, 469. [Google Scholar] [CrossRef] [Green Version]
- Dum, R.P.; Strick, P.L. An unfolded map of the cerebellar dentate nucleus and its projections to the cerebral cortex. J. Neurophysiol. 2003, 89, 634–639. [Google Scholar] [CrossRef]
- Bostan, A.C.; Strick, P.L. The basal ganglia and the cerebellum: Nodes in an integrated network. Nat. Rev. Neurosci. 2018, 19, 338–350. [Google Scholar] [CrossRef]
- Lehéricy, S.; Benali, H.; Van de Moortele, P.-F.; Pélégrini-Issac, M.; Waechter, T.; Ugurbil, K.; Doyon, J. Distinct basal ganglia territories are engaged in early and advanced motor sequence learning. Proc. Natl. Acad. Sci. USA 2005, 102, 12566–12571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshi, E.; Tremblay, L.; Féger, J.; Carras, P.L.; Strick, P.L. The cerebellum communicates with the basal ganglia. Nat. Neurosci. 2005, 8, 1491–1493. [Google Scholar] [CrossRef] [PubMed]
- Buisseret-Delmas, C.; Angaut, P. The cerebellar olivo-corticonuclear connections in the rat. Prog. Neurobiol. 1993, 40, 63–87. [Google Scholar] [CrossRef]
- Hilker, R.; Voges, J.; Weisenbach, S.; Kalbe, E.; Burghaus, L.; Ghaemi, M.; Lehrke, R.; Koulousakis, A.; Herholz, K.; Sturm, V. Subthalamic nucleus stimulation restores glucose metabolism in associative and limbic cortices and in cerebellum: Evidence from a FDG-PET study in advanced Parkinson’s disease. J. Cereb. Blood Flow Metab. 2004, 24, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Castrioto, A.; Moro, E. New targets for deep brain stimulation treatment of Parkinson’s disease. Expert Rev. Neurother. 2013, 13, 1319–1328. [Google Scholar] [CrossRef]
- Pahapill, P.A.; Lozano, A.M. The pedunculopontine nucleus and Parkinson’s disease. Brain 2000, 123, 1767–1783. [Google Scholar] [CrossRef]
- Alam, M.; Schwabe, K.; Krauss, J.K. The pedunculopontine nucleus area: Critical evaluation of interspecies differences relevant for its use as a target for deep brain stimulation. Brain 2011, 134, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Nandi, D.; Stein, J.; Aziz, T. Exploration of the role of the upper brainstem in motor control. Stereotact. Funct. Neurosurg. 2002, 78, 158–167. [Google Scholar] [CrossRef]
- Jenkinson, N.; Nandi, D.; Miall, R.C.; Stein, J.F.; Aziz, T.Z. Pedunculopontine nucleus stimulation improves akinesia in a Parkinsonian monkey. Neuroreport 2004, 15, 2621–2624. [Google Scholar] [CrossRef] [Green Version]
- Goetz, L.; Bhattacharjee, M.; Ferraye, M.U.; Fraix, V.; Maineri, C.; Nosko, D.; Fenoy, A.J.; Piallat, B.; Torres, N.; Krainik, A. Deep brain stimulation of the pedunculopontine nucleus area in Parkinson disease: MRI-based anatomoclinical correlations and optimal target. Neurosurgery 2019, 84, 506–518. [Google Scholar] [CrossRef] [Green Version]
- Ferraye, M.U.; Debû, B.; Fraix, V.; Goetz, L.; Ardouin, C.; Yelnik, J.; Henry-Lagrange, C.; Seigneuret, E.; Piallat, B.; Krack, P.; et al. Effects of pedunculopontine nucleus area stimulation on gait disorders in Parkinson’s disease. Brain 2010, 133, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Tykocki, T.; Mandat, T.; Nauman, P. Pedunculopontine nucleus deep brain stimulation in Parkinson’s disease. Arch. Med. Sci. AMS 2011, 7, 555–564. [Google Scholar] [CrossRef]
- Stefani, A.; Lozano, A.M.; Peppe, A.; Stanzione, P.; Galati, S.; Tropepi, D.; Pierantozzi, M.; Brusa, L.; Scarnati, E.; Mazzone, P. Bilateral deep brain stimulation of the pedunculopontine and subthalamic nuclei in severe Parkinson’s disease. Brain 2007, 130, 1596–1607. [Google Scholar] [CrossRef] [PubMed]
- Plaha, P.; Javed, S.; Agombar, D.; O’Farrell, G.; Khan, S.; Whone, A.; Gill, S. Bilateral caudal zona incerta nucleus stimulation for essential tremor: Outcome and quality of life. J. Neurol. Neurosurg. Psychiatry 2011, 82, 899–904. [Google Scholar] [CrossRef] [PubMed]
- Godinho, F.; Thobois, S.; Magnin, M.; Guenot, M.; Polo, G.; Benatru, I.; Xie, J.; Salvetti, A.; Garcia-Larrea, L.; Broussolle, E.; et al. Subthalamic nucleus stimulation in Parkinson’s disease. J. Neurol. 2006, 253, 1347–1355. [Google Scholar] [CrossRef] [PubMed]
- Miocinovic, S.; Parent, M.; Butson, C.R.; Hahn, P.J.; Russo, G.S.; Vitek, J.L.; McIntyre, C.C. Computational analysis of subthalamic nucleus and lenticular fasciculus activation during therapeutic deep brain stimulation. J. Neurophysiol. 2006, 96, 1569–1580. [Google Scholar] [CrossRef] [Green Version]
- Butson, C.R.; Cooper, S.E.; Henderson, J.M.; McIntyre, C.C. Patient-specific analysis of the volume of tissue activated during deep brain stimulation. Neuroimage 2007, 34, 661–670. [Google Scholar] [CrossRef] [Green Version]
- Hägglund, P.; Sandström, L.; Blomstedt, P.; Karlsson, F. Voice Tremor in Patients with Essential Tremor: Effects of Deep Brain Stimulation of Caudal Zona Incerta. J. Voice 2016, 30, 228–233. [Google Scholar] [CrossRef]
- Wang, X.; Chou, X.L.; Zhang, L.I.; Tao, H.W. Zona Incerta: An Integrative Node for Global Behavioral Modulation. Trends Neurosci. 2020, 43, 82–87. [Google Scholar] [CrossRef]
- dos Santos Ghilardi, M.G.; Cury, R.G.; dos Ângelos, J.S.; Barbosa, D.C.; Barbosa, E.R.; Teixeira, M.J.; Fonoff, E.T. Long-term improvement of tremor and ataxia after bilateral DBS of VoP/zona incerta in FXTAS. Neurology 2015, 84, 1904–1906. [Google Scholar] [CrossRef]
- Ossowska, K. Zona incerta as a therapeutic target in Parkinson’s disease. J. Neurol. 2020, 267, 591–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisinger, R.S.; Wong, J.; Almeida, L.; Ramirez-Zamora, A.; Cagle, J.N.; Giugni, J.C.; Ahmed, B.; Bona, A.R.; Monari, E.; Wagle Shukla, A.; et al. Ventral Intermediate Nucleus Versus Zona Incerta Region Deep Brain Stimulation in Essential Tremor. Mov. Disord. Clin. Pract. 2017, 5, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Sasagawa, A.; Enatsu, R.; Kitagawa, M.; Mikami, T.; Nakayama-Kamada, C.; Kuribara, T.; Hirano, T.; Arihara, M.; Mikuni, N. Target Selection of Directional Lead in Patients with Parkinson’s Disease. Neurol. Med. Chir. 2020, 60, 622–628. [Google Scholar] [CrossRef]
- Benabid, A.L.; Pollak, P.; Hoffmann, D.; Gervason, C.; Hommel, M.; Perret, J.E.; De Rougemont, J.; Gao, D.M. Long-term suppression of tremor by chronic stimulation of the ventral intermediate thalamic nucleus. Lancet. 1991, 337, 403–406. [Google Scholar] [CrossRef]
- Warren, A.E.; Dalic, L.J.; Thevathasan, W.; Roten, A.; Bulluss, K.J.; Archer, J. Targeting the centromedian thalamic nucleus for deep brain stimulation. J. Neurol. Neurosurg. Psychiatry 2020, 91, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Kerkerian-Le Goff, L.; Jouve, L.; Melon, C.; Salin, P. Rationale for targeting the thalamic centre-median parafascicular complex in the surgical treatment of Parkinson’s disease. Parkinsonism Relat. Disord. 2009, 15, S167–S170. [Google Scholar] [CrossRef]
- Caparros-Lefebvre, D.; Blond, S.; Feltin, M.-P.; Pollak, P.; Benabid, A.L. Improvement of levodopa induced dyskinesias by thalamic deep brain stimulation is related to slight variation in electrode placement: Possible involvement of the centre median and parafascicularis complex. J. Neurol. Neurosurg. Psychiatry 1999, 67, 308–314. [Google Scholar] [CrossRef]
- Peppe, A.; Gasbarra, A.; Stefani, A.; Chiavalon, C.; Pierantozzi, M.; Fermi, E.; Stanzione, P.; Caltagirone, C.; Mazzone, P. Deep brain stimulation of CM/PF of thalamus could be the new elective target for tremor in advanced Parkinson’s Disease? Parkinsonism Relat. Disord. 2008, 14, 501–504. [Google Scholar] [CrossRef]
- Stefani, A.; Peppe, A.; Pierantozzi, M.; Galati, S.; Moschella, V.; Stanzione, P.; Mazzone, P. Multi-target strategy for Parkinsonian patients: The role of deep brain stimulation in the centromedian–parafascicularis complex. Brain Res. Bull. 2009, 78, 113–118. [Google Scholar] [CrossRef]
- Jouve, L.; Salin, P.; Melon, C.; Kerkerian-Le Goff, L. Deep brain stimulation of the center median–parafascicular complex of the thalamus has efficient anti-parkinsonian action associated with widespread cellular responses in the basal ganglia network in a rat model of Parkinson’s disease. J. Neurosci. 2010, 30, 9919–9928. [Google Scholar] [CrossRef] [Green Version]
- Takakusaki, K.; Habaguchi, T.; Ohtinata-Sugimoto, J.; Saitoh, K.; Sakamoto, T. Basal ganglia efferents to the brainstem centers controlling postural muscle tone and locomotion: A new concept for understanding motor disorders in basal ganglia dysfunction. Neuroscience 2003, 119, 293–308. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.; Beecher, G.; Ba, F. Deep brain stimulation in Parkinson’s disease: New and emerging targets for refractory motor and nonmotor symptoms. Parkinsons Dis. 2017, 5, 5124328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chastan, N.; Westby, G.W.M.; Yelnik, J.; Bardinet, E.; Do, M.C.; Agid, Y.; Welter, M.L. Effects of nigral stimulation on locomotion and postural stability in patients with Parkinson’s disease. Brain 2009, 132, 172–184. [Google Scholar] [CrossRef] [PubMed]
- Weiss, D.; Walach, M.; Meisner, C.; Fritz, M.; Scholten, M.; Breit, S.; Plewnia, C.; Bender, B.; Gharabaghi, A.; Wächter, T.; et al. Nigral stimulation for resistant axial motor impairment in Parkinson’s disease? A randomized controlled trial. Brain 2013, 136, 2098–2108. [Google Scholar] [CrossRef] [Green Version]
- Valldeoriola, F.; Muñoz, E.; Rumià, J.; Roldán, P.; Cámara, A.; Compta, Y.; Martí, M.J.; Tolosa, E. Simultaneous low-frequency deep brain stimulation of the substantia nigra pars reticulata and high-frequency stimulation of the subthalamic nucleus to treat levodopa unresponsive freezing of gait in Parkinson’s disease: A pilot study. Parkinsonism Relat. Disord. 2019, 60, 153–157. [Google Scholar] [CrossRef] [Green Version]
- Parker, T.; Raghu, A.L.; FitzGerald, J.J.; Green, A.L.; Aziz, T.Z. Multitarget deep brain stimulation for clinically complex movement disorders. J. Neurosurg. 2020, 1, 351–356. [Google Scholar] [CrossRef]
- Slotty, P.J.; Poologaindran, A.; Honey, C.R. A prospective, randomized, blinded assessment of multitarget thalamic and pallidal deep brain stimulation in a case of hemidystonia. Clin. Neurol. Neurosurg. 2015, 138, 16–19. [Google Scholar] [CrossRef]
- Mazzone, P. Deep brain stimulation in Parkinson’s disease: Bilateral implantation of globus pallidus and subthalamic nucleus. J. Neurosurg. Sci. 2003, 47, 47–51. [Google Scholar]
- Mazzone, P.; Brown, P.; DiLazzaro, V.; Stanzione, P.; Oliviero, A.; Peppe, A.; Santilli, V.; Insola, A.; Altibrandi, M. Bilateral implantation in globus pallidus internus and in subthalamic nucleus in Parkinson’s disease. Neuromodul. Technol. Neural Interface 2005, 8, 1–6. [Google Scholar] [CrossRef]
- Follett, K.A.; Weaver, F.M.; Stern, M.; Hur, K.; Harris, C.L.; Luo, P.; Marks, W.J., Jr.; Rothlind, J.; Sagher, O.; Moy, C.; et al. Pallidal versus subthalamic deep-brain stimulation for Parkinson’s disease. N. Engl. J. Med. 2010, 362, 2077–2091. [Google Scholar] [CrossRef] [Green Version]
- Oxley, T.J.; Opie, N.L.; John, S.E.; Rind, G.S.; Ronayne, S.M.; Wheeler, T.L.; Judy, J.W.; McDonald, A.J.; Dornom, A.; Lovell, T.J.; et al. Minimally invasive endovascular stent-electrode array for high-fidelity, chronic recordings of cortical neural activity. Nat. Biotechnol. 2016, 34, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Rajah, G.; Saber, H.; Singh, R.; Rangel-Castilla, L. Endovascular delivery of leads and stentrodes and their applications to deep brain stimulation and neuromodulation: A review. Neurosurg. Focus. 2018, 45, E19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oxley, T.J.; Yoo, P.E.; Rind, G.S.; Ronayne, S.M.; Lee, C.S.; Bird, C.; Hampshire, V.; Sharma, R.P.; Morokoff, A.; Williams, D.L.; et al. Motor neuroprosthesis implanted with neurointerventional surgery improves capacity for activities of daily living tasks in severe paralysis: First in-human experience. J. Neurointerv. Surg. 2021, 13, 102–108. [Google Scholar] [CrossRef]
- Turk, A.; Niemann, D.; Ahmed, A.; Aagaard-Kienitz, B. Use of self-expanding stents in distal small cerebral vessels. Am. J. Neuroradiol. 2007, 28, 533–536. [Google Scholar]
- Bernier, M.; Cunnane, S.C.; Whittingstall, K. The morphology of the human cerebrovascular system. Hum. Brain Mapp. 2018, 39, 4962–4975. [Google Scholar] [CrossRef] [Green Version]
- Neudorfer, C.; Bhatia, K.; Boutet, A.; Germann, J.; Elias, G.J.; Loh, A.; Paff, M.; Krings, T.; Lozano, A.M. Endovascular deep brain stimulation: Investigating the relationship between vascular structures and deep brain stimulation targets. Brain Stimul. 2020, 13, 1668–1677. [Google Scholar] [CrossRef] [PubMed]
- Rangel-Castilla, L.; Cress, M.C.; Munich, S.A.; Sonig, A.; Krishna, C.; Gu, E.Y.; Snyder, K.V.; Hopkins, L.N.; Siddiqui, A.H.; Levy, E.I. Feasibility, safety, and periprocedural complications of pipeline embolization for intracranial aneurysm treatment under conscious sedation: University at Buffalo Neurosurgery Experience. Oper. Neurosurg. 2015, 11, 426–430. [Google Scholar] [CrossRef]
- Orlov, K.; Kislitsin, D.; Strelnikov, N.; Berestov, V.; Gorbatykh, A.; Shayakhmetov, T.; Seleznev, P.; Tasenko, A. Experience using pipeline embolization device with Shield Technology in a patient lacking a full postoperative dual antiplatelet therapy regimen. Interv. Neuroradiol. 2018, 24, 270–273. [Google Scholar] [CrossRef]
- Jenkinson, N.; Brown, P. New insights into the relationship between dopamine, beta oscillations and motor function. Trends Neurosci. 2011, 34, 611–618. [Google Scholar] [CrossRef]
- Hammond, C.; Bergman, H.; Brown, P. Pathological synchronization in Parkinson’s disease: Networks, models and treatments. Trends Neurosci. 2007, 30, 357–364. [Google Scholar] [CrossRef]
- Brittain, J.S.; Brown, P. Oscillations and the basal ganglia: Motor control and beyond. Neuroimage 2014, 85, 637–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kühn, A.A.; Kupsch, A.; Schneider, G.H.; Brown, P. Reduction in subthalamic 8–35 Hz oscillatory activity correlates with clinical improvement in Parkinson’s disease. Eur. J. Neurosci. 2006, 23, 1956–1960. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, J.; Kang, Y.; Ghori, M.B. Emergence of Beta Oscillations of a Resonance Model for Parkinson’s Disease. Neural Plast. 2020, 2020, 8824760. [Google Scholar] [CrossRef] [PubMed]
- Davis, N.J.; van Koningsbruggen, M.G. “Non-invasive” brain stimulation is not non-invasive. Front. Syst. Neurosci. 2013, 7, 76. [Google Scholar] [CrossRef] [PubMed]
- Schulz, R.; Gerloff, C.; Hummel, F.C. Non-invasive brain stimulation in neurological diseases. Neuropharmacology 2013, 64, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Priori, A.; Hallett, M.; Rothwell, J.C. Repetitive transcranial magnetic stimulation or transcranial direct current stimulation? Brain Stimul. 2009, 2, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Liebetanz, D.; Fauser, S.; Michaelis, T.; Czéh, B.; Watanabe, T.; Paulus, W.; Frahm, J.; Fuchs, E. Safety aspects of chronic low-frequency transcranial magnetic stimulation based on localized proton magnetic resonance spectroscopy and histology of the rat brain. J. Psychiatr. Res. 2003, 37, 277–286. [Google Scholar] [CrossRef]
- Rezayat, E.; Toostani, I.G. A Review on Brain Stimulation Using Low Intensity Focused Ultrasound. Basic Clin. Neurosci. 2016, 7, 187–194. [Google Scholar]
- Montemurro, N.; Perrini, P.; Marani, W.; Chaurasia, B.; Corsalini, M.; Scarano, A.; Rapone, B. Multiple Brain Abscesses of Odontogenic Origin. May Oral Microbiota Affect Their Development? A Review of the Current Literature. Appl. Sci. 2021, 11, 3316. [Google Scholar] [CrossRef]
- Bronstein, J.M.; Tagliati, M.; Alterman, R.L.; Lozano, A.M.; Volkmann, J.; Stefani, A.; Horak, F.B.; Okun, M.S.; Foote, K.D.; Krack, P. Deep brain stimulation for Parkinson disease: An expert consensus and review of key issues. Arch. Neurol. 2011, 68, 165. [Google Scholar] [CrossRef]
- Das, S.; Montemurro, N.; Ashfaq, M.; Ghosh, D.; Sarker, A.C.; Khan, A.H.; Dey, S.; Chaurasia, B. Resolution of Papilledema Following Ventriculoperitoneal Shunt or Endoscopic Third Ventriculostomy for Obstructive Hydrocephalus: A Pilot Study. Medicina 2022, 58, 281. [Google Scholar] [CrossRef]
- Fregni, F.; Simon, D.K.; Wu, A.; Pascual-Leone, A. Non-invasive brain stimulation for Parkinson’s disease: A systematic review and meta-analysis of the literature. J. Neurol. Neurosurg. Psychiatry 2005, 76, 1614–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benninger, D.H.; Lomarev, M.; Lopez, G.; Wassermann, E.M.; Li, X.; Considine, E.; Hallett, M. Transcranial direct current stimulation for the treatment of Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 2011, 82, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latorre, A.; Rocchi, L.; Berardelli, A.; Bhatia, K.P.; Rothwell, J.C. The use of transcranial magnetic stimulation as a treatment for movement disorders: A critical review. Mov. Disord. 2019, 34, 769–782. [Google Scholar] [CrossRef] [PubMed]
- Chen, R. Guideline on therapeutic use of repetitive transcranial magnetic stimulation: Useful but know the methods and limitations. Clin. Neurophysiol. 2020, 131, 461–462. [Google Scholar] [CrossRef]
- Brandt, S.; Ploner, C.; Meyer, B.U. Repetitive transkranielle Magnetstimulation. Der Nervenarzt 1997, 68, 778–784. [Google Scholar] [CrossRef]
- Paulus, W. Transcranial brain stimulation: Potential and limitations. e-Neuroforum 2014, 5, 29–36. [Google Scholar] [CrossRef]
- Woods, A.J.; Antal, A.; Bikson, M.; Boggio, P.S.; Brunoni, A.R.; Celnik, P.; Cohen, L.G.; Fregni, F.; Herrmann, C.S.; Kappenman, E.S.; et al. A technical guide to tDCS, and related non-invasive brain stimulation tools. Clin. Neurophysiol. 2016, 127, 1031–1048. [Google Scholar] [CrossRef] [Green Version]
- Fritsch, B.; Reis, J.; Martinowich, K.; Schambra, H.M.; Ji, Y.; Cohen, L.G.; Lu, B. Direct current stimulation promotes BDNF-dependent synaptic plasticity: Potential implications for motor learning. Neuron 2010, 66, 198–204. [Google Scholar] [CrossRef] [Green Version]
- Brittain, J.S.; Cagnan, H. Recent trends in the use of electrical neuromodulation in Parkinson’s disease. Curr. Behav. Neurosci. Rep. 2018, 5, 170–178. [Google Scholar] [CrossRef] [Green Version]
- Krause, V.; Wach, C.; Südmeyer, M.; Ferrea, S.; Schnitzler, A.; Pollok, B. Cortico-muscular coupling and motor performance are modulated by 20 Hz transcranial alternating current stimulation (tACS) in Parkinson’s disease. Front. Hum. Neurosci. 2014, 7, 928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brittain, J.S.; Cagnan, H.; Mehta, A.R.; Saifee, T.A.; Edwards, M.J.; Brown, P. Distinguishing the central drive to tremor in Parkinson’s disease and essential tremor. J. Neurosci. 2015, 35, 795–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thut, G.; Bergmann, T.O.; Fröhlich, F.; Soekadar, S.R.; Brittain, J.S.; Valero-Cabré, A.; Sack, A.T.; Miniussi, C.; Antal, A.; Siebner, H.R.; et al. Guiding transcranial brain stimulation by EEG/MEG to interact with ongoing brain activity and associated functions: A position paper. Clin. Neurophysiol. 2017, 128, 843–857. [Google Scholar] [CrossRef] [Green Version]
- Del Felice, A.; Castiglia, L.; Formaggio, E.; Cattelan, M.; Scarpa, B.; Manganotti, P.; Tenconi, E.; Masiero, S. Personalized transcranial alternating current stimulation (tACS) and physical therapy to treat motor and cognitive symptoms in Parkinson’s disease: A randomized cross-over trial. NeuroImage Clin. 2019, 22, 101768. [Google Scholar] [CrossRef]
- Shill, H.A.; Obradov, S.; Katsnelson, Y.; Pizinger, R. A randomized, double-blind trial of transcranial electrostimulation in early Parkinson’s disease. Mov. Disord. 2011, 26, 1477–1480. [Google Scholar] [CrossRef] [PubMed]
- Moosa, S.; Martínez-Fernández, R.; Elias, W.J.; Del Alamo, M.; Eisenberg, H.M.; Fishman, P.S. The role of high-intensity focused ultrasound as a symptomatic treatment for Parkinson’s disease. Mov. Disord. 2019, 34, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, I.; Sinai, A.; Zaaroor, M. MRI-guided focused ultrasound in Parkinson’s disease: A review. Parkinsons Dis. 2017, 2017, 8124624. [Google Scholar] [CrossRef]
- Quadri, S.A.; Waqas, M.; Khan, I.; Khan, M.A.; Suriya, S.S.; Farooqui, M.; Fiani, B. High-intensity focused ultrasound: Past, present, and future in neurosurgery. Neurosurg. Focus 2018, 44, E16. [Google Scholar] [CrossRef] [Green Version]
- Zaaroor, M.; Sinai, A.; Goldsher, D.; Eran, A.; Nassar, M.; Schlesinger, I. Magnetic resonance-guided focused ultrasound thalamotomy for tremor: A report of 30 Parkinson’s disease and essential tremor cases. J Neurosurg. 2018, 128, 202–210. [Google Scholar] [CrossRef] [Green Version]
- Chung, A.H.; Jolesz, F.A.; Hynynen, K. Thermal dosimetry of a focused ultrasound beam in vivo by magnetic resonance imaging. Med. Phys. 1999, 26, 2017–2026. [Google Scholar] [CrossRef]
- Burks, S.R.; Ziadloo, A.; Hancock, H.A.; Chaudhry, A.; Dean, D.D.; Lewis, B.K.; Frenkel, V.; Frank, J.A. Investigation of cellular and molecular responses to pulsed focused ultrasound in a mouse model. PLoS ONE 2011, 6, e24730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishna, V.; Sammartino, F.; Rezai, A. A review of the current therapies, challenges, and future directions of transcranial focused ultrasound technology: Advances in diagnosis and treatment. JAMA Neurol. 2018, 75, 246–254. [Google Scholar] [CrossRef] [PubMed]
- LeWitt, P.A.; Lipsman, N.; Kordower, J.H. Focused ultrasound opening of the blood-brain barrier for treatment of Parkinson’s disease. Mov. Disord. 2019, 34, 1274–1278. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Fernández, R.; Máñez-Miró, J.U.; Rodríguez-Rojas, R.; Del Álamo, M.; Shah, B.B.; Hernández-Fernández, F.; Pineda-Pardo, J.A.; Monje, M.H.; Fernández-Rodríguez, B.; Sperling, S.A.; et al. Randomized trial of focused ultrasound subthalamotomy for Parkinson’s disease. N. Engl. J. Med. 2020, 383, 2501–2513. [Google Scholar] [CrossRef]
- Jung, N.Y.; Park, C.K.; Kim, M.; Lee, P.H.; Sohn, Y.H.; Chang, J.W. The efficacy and limits of magnetic resonance-guided focused ultrasound pallidotomy for Parkinson’s disease: A Phase I clinical trial. J Neurosurg. 2018, 130, 1853–1861. [Google Scholar] [CrossRef] [Green Version]
- Tan, E.K.; Chan, L.L. A Trial of focused ultrasound thalamotomy for essential tremor. N. Engl. J. Med. 2016, 375, 2202–2203. [Google Scholar]
- Martin, E.; Jeanmonod, D.; Morel, A.; Zadicario, E.; Werner, B. High-intensity focused ultrasound for noninvasive functional neurosurgery. Ann. Neurol. 2009, 66, 858–861. [Google Scholar] [CrossRef] [Green Version]
- Bond, A.E.; Shah, B.B.; Huss, D.S.; Dallapiazza, R.F.; Warren, A.; Harrison, M.B.; Sperling, S.A.; Wang, X.Q.; Gwinn, R.; Witt, J.; et al. Safety and Efficacy of focused ultrasound thalamotomy for patients with medication-refractory, tremor-dominant Parkinson disease: A randomized clinical trial. JAMA Neurol. 2017, 74, 1412–1418. [Google Scholar] [CrossRef]
- Ohye, C.; Higuchi, Y.; Shibazaki, T.; Hashimoto, T.; Koyama, T.; Hirai, T.; Matsuda, S.; Serizawa, T.; Hori, T.; Hayashi, M.; et al. Gamma knife thalamotomy for Parkinson disease and essential tremor: A prospective multicenter study. Neurosurgery 2012, 70, 526–536. [Google Scholar] [CrossRef]
- Dauer, W.; Przedborski, S. Parkinson’s disease: Mechanisms and models. Neuron 2003, 39, 889–909. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.P.; Burrell, J.C.; Struzyna, L.A.; Chen, H.I.; Serruya, M.D.; Wolf, J.A.; Duda, J.E.; Cullen, D.K. Emerging regenerative medicine and tissue engineering strategies for Parkinson’s disease. NPJ Parkinsons Dis. 2020, 6, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, S.J.; Diener, M.D.; Kaltenboeck, A.; Birnbaum, H.G.; Siderowf, A.D. An economic model of Parkinson’s disease: Implications for slowing progression in the United States. Mov. Disord. 2013, 28, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Khademhosseini, A.; Langer, R. A decade of progress in tissue engineering. Nat. Protoc. 2016, 11, 1775–1781. [Google Scholar] [CrossRef] [PubMed]
- Petersen, S.E.; Sporns, O. Brain Networks and Cognitive Architectures. Neuron 2015, 88, 207–219. [Google Scholar] [CrossRef] [Green Version]
- Björklund, A.; Schmidt, R.H.; Stenevi, U. Functional reinnervation of the neostriatum in the adult rat by use of intraparenchymal grafting of dissociated cell suspensions from the substantia nigra. Cell Tissue Res. 1980, 212, 39–45. [Google Scholar] [CrossRef]
- Isacson, O.; Bjorklund, L.M.; Schumacher, J.M. Toward full restoration of synaptic and terminal function of the dopaminergic system in Parkinson’s disease by stem cells. Ann. Neurol. 2003, 53, S135–S148. [Google Scholar] [CrossRef]
- Rivetti Di Val Cervo, P.; Romanov, R.A.; Spigolon, G.; Masini, D.; Martín-Montañez, E.; Toledo, E.M.; La Manno, G.; Feyder, M.; Pifl, C.; Ng, Y.H.; et al. Induction of functional dopamine neurons from human astrocytes in vitro and mouse astrocytes in a Parkinson’s disease model. Nat. Biotechnol. 2017, 35, 444–452. [Google Scholar] [CrossRef]
- Harris, J.P.; Struzyna, L.A.; Murphy, P.L.; Adewole, D.O.; Kuo, E.; Cullen, D.K. Advanced biomaterial strategies to transplant preformed micro-tissue engineered neural networks into the brain. J. Neural Eng. 2016, 13, 016019. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Chen, G. In Vivo Reprogramming for CNS Repair: Regenerating Neurons from Endogenous Glial Cells. Neuron 2016, 91, 728–738. [Google Scholar] [CrossRef] [Green Version]
- Dunnett, S.B.; Bjorklund, A.; Schmidt, R.H.; Stenevi, U.; Iversen, S.D. Intracerebral grafting of neuronal cell suspensions. IV. Behavioural recovery in rats with unilateral 6-OHDA lesions following implantation of nigral cell suspensions in different forebrain sites. Acta Physiol. Scand 1983, 522, 29–37. [Google Scholar]
- Björklund, A.; Lindvall, O. Cell replacement therapies for central nervous system disorders. Nat. Neurosci. 2000, 3, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Stoker, T.B.; Torsney, K.M.; Barker, R.A. Emerging Treatment Approaches for Parkinson’s Disease. Front. Neurosci. 2018, 12, 693. [Google Scholar] [CrossRef] [Green Version]
- Brundin, P.; Nilsson, O.G.; Strecker, R.E.; Lindvall, O.; Astedt, B.; Björklund, A. Behavioural effects of human fetal dopamine neurons grafted in a rat model of Parkinson’s disease. Exp. Brain Res. 1986, 65, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Kordower, J.H.; Chu, Y.; Hauser, R.A.; Freeman, T.B.; Olanow, C.W. Lewy body-like pathology in long-term embryonic nigral transplants in Parkinson’s disease. Nat. Med. 2008, 14, 504–506. [Google Scholar] [CrossRef] [PubMed]
- Barker, R.A.; Barrett, J.; Mason, S.L.; Björklund, A. Fetal dopaminergic transplantation trials and the future of neural grafting in Parkinson’s disease. Lancet Neurol. 2013, 12, 84–91. [Google Scholar] [CrossRef]
- Bolam, J.P.; Freund, T.F.; Björklund, A.; Dunnett, S.B.; Smith, A.D. Synaptic input and local output of dopaminergic neurons in grafts that functionally reinnervate the host neostriatum. Exp. Brain Res. 1987, 68, 131–146. [Google Scholar] [CrossRef]
- Kordower, J.H.; Freeman, T.B.; Chen, E.Y.; Mufson, E.J.; Sanberg, P.R.; Hauser, R.A.; Snow, B.; Warren Olanow, C. Fetal nigral grafts survive and mediate clinical benefit in a patient with Parkinson’s disease. Mov. Disord. 1998, 13, 383–393. [Google Scholar] [CrossRef]
- Lindvall, O.; Björklund, A. Cell therapy in Parkinson’s disease. NeuroRx 2004, 1, 382–393. [Google Scholar] [CrossRef]
- Olanow, C.W.; Kordower, J.H.; Freeman, T.B. Fetal nigral transplantation as a therapy for Parkinson’s disease. Trends Neurosci. 1996, 19, 102–109. [Google Scholar] [CrossRef]
- Li, W.; Englund, E.; Widner, H.; Mattsson, B.; van Westen, D.; Lätt, J.; Rehncrona, S.; Brundin, P.; Björklund, A.; Lindvall, O.; et al. Extensive graft-derived dopaminergic innervation is maintained 24 years after transplantation in the degenerating parkinsonian brain. Proc. Natl. Acad. Sci. USA 2016, 113, 6544–6549. [Google Scholar] [CrossRef] [Green Version]
- Brederlau, A.; Correia, A.S.; Anisimov, S.V.; Elmi, M.; Paul, G.; Roybon, L.; Morizane, A.; Bergquist, F.; Riebe, I.; Nannmark, U.; et al. Transplantation of human embryonic stem cell-derived cells to a rat model of Parkinson’s disease: Effect of in vitro differentiation on graft survival and teratoma formation. Stem Cells 2006, 24, 1433–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindvall, O.; Björklund, A. Cell therapeutics in Parkinson’s disease. Neurotherapeutics 2011, 8, 539–548. [Google Scholar] [CrossRef] [Green Version]
- Bezard, E.; Brotchie, J.M.; Gross, C.E. Pathophysiology of levodopa-induced dyskinesia: Potential for new therapies. Nat. Rev. Neurosci. 2001, 2, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Barker, R.A.; Drouin-Ouellet, J.; Parmar, M. Cell-based therapies for Parkinson disease—Past insights and future potential. Nat. Rev. Neurol. 2015, 11, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Parmar, M.; Torper, O.; Drouin-Ouellet, J. Cell-based therapy for Parkinson’s disease: A journey through decades toward the light side of the Force. Eur. J. Neurosci. 2019, 49, 463–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grealish, S.; Diguet, E.; Kirkeby, A.; Mattsson, B.; Heuer, A.; Bramoulle, Y.; Van Camp, N.; Perrier, A.L.; Hantraye, P.; Björklund, A.; et al. Human ESC-derived dopamine neurons show similar preclinical efficacy and potency to fetal neurons when grafted in a rat model of Parkinson’s disease. Cell Stem Cell 2014, 15, 653–665. [Google Scholar] [CrossRef] [Green Version]
- Yi, B.R.; Kim, S.U.; Choi, K.C. Development and application of neural stem cells for treating various human neurological diseases in animal models. Lab. Anim. Res. 2013, 29, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Hellmann, M.A.; Panet, H.; Barhum, Y.; Melamed, E.; Offen, D. Increased survival and migration of engrafted mesenchymal bone marrow stem cells in 6-hydroxydopamine-lesioned rodents. Neurosci. Lett. 2006, 395, 124–128. [Google Scholar] [CrossRef]
- Struzyna, L.A.; Katiyar, K.; Cullen, D.K. Living scaffolds for neuroregeneration. Curr. Opin. Solid State Mater Sci. 2014, 18, 308–318. [Google Scholar] [CrossRef] [Green Version]
- Montemurro, N.; Scerrati, A.; Ricciardi, L.; Trevisi, G. The Exoscope in Neurosurgery: An Overview of the Current Lit-erature of Intraoperative Use in Brain and Spine Surgery. J. Clin. Med. 2021, 11, 223. [Google Scholar] [CrossRef]
- Mishra, R.; Narayanan, M.K.; Umana, G.E.; Montemurro, N.; Chaurasia, B.; Deora, H. Virtual Reality in Neurosurgery: Beyond Neurosurgical Planning. Int. J. Environ. Res. Public Health 2022, 19, 1719. [Google Scholar] [CrossRef] [PubMed]
- Montemurro, N. Telemedicine: Could it represent a new problem for spine surgeons to solve? Global Spine J. 2022, 12, 1306–1307. [Google Scholar] [CrossRef] [PubMed]
- Winter, C.C.; Katiyar, K.S.; Hernandez, N.S.; Song, Y.J.; Struzyna, L.A.; Harris, J.P.; Cullen, D.K. Transplantable living scaffolds comprised of micro-tissue engineered aligned astrocyte networks to facilitate central nervous system regeneration. Acta Biomater. 2016, 38, 44–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, B.; Zhang, C.; Smith, G.M. Bridging between transplantation therapy and neurotrophic factors in Parkinson’s disease. Front. Biosci. 2014, 6, 225–235. [Google Scholar] [CrossRef]
- Ricciardi, L.; Pucci, R.; Piazza, A.; Lofrese, G.; Scerrati, A.; Montemurro, N.; Raco, A.; Miscusi, M.; Ius, T.; Zeppieri, M.; et al. Role of stem cells-based in facial nerve reanimation: A meta-analysis of histological and neurophysiological outcomes. World J. Stem Cells 2022, 14, 420–428. [Google Scholar] [CrossRef]
- Montemurro, N.; Condino, S.; Carbone, M.; Cattari, N.; D’Amato, R.; Cutolo, F.; Ferrari, V. Brain Tumor and Augmented Reality: New Technologies for the Future. Int. J. Environ. Res. Public Health 2022, 19, 6347. [Google Scholar] [CrossRef]
- Gaillard, A.; Jaber, M. Rewiring the brain with cell transplantation in Parkinson’s disease. Trends Neurosci. 2011, 34, 124–133. [Google Scholar] [CrossRef]
- Weng, S.J.; Li, I.H.; Huang, Y.S.; Chueh, S.H.; Chou, T.K.; Huang, S.Y.; Shiue, C.Y.; Cheng, C.Y.; Ma, K.H. KA-bridged transplantation of mesencephalic tissue and olfactory ensheathing cells in a Parkinsonian rat model. J. Tissue Eng. Regen. Med. 2017, 11, 2024–2033. [Google Scholar] [CrossRef]
- Cullen, D.K.; Tang-Schomer, M.D.; Struzyna, L.A.; Patel, A.R.; Johnson, V.E.; Wolf, J.A.; Smith, D.H. Microtissue engineered constructs with living axons for targeted nervous system reconstruction. Tissue Eng. Part A 2012, 18, 2280–2289. [Google Scholar] [CrossRef] [Green Version]
- Ward, R.J.; Zucca, F.A.; Duyn, J.H.; Crichton, R.R.; Zecca, L. The role of iron in brain ageing and neurodegenerative disorders. Lancet Neurol. 2014, 13, 1045–1060. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Li, C.; Zhang, T.; Ge, Y.; Han, X.; Sun, S.; Ding, J.; Lu, M.; Hu, G. Deletion of Kir6.2/SUR1 potassium channels rescues diminishing of DA neurons via decreasing iron accumulation in PD. Mol. Cell Neurosci. 2018, 92, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Cassar, I.R.; Sambangi, J.; Grill, W.M. Frequency-specific optogenetic deep brain stimulation of subthalamic nucleus improves Parkinsonian motor behaviors. J. Neurosci. 2020, 40, 4323–4334. [Google Scholar] [CrossRef] [PubMed]
- Gittis, A.H.; Yttri, E.A. Translating Insights from Optogenetics to Therapies for Parkinson’s Disease. Curr. Opin. Biomed Eng. 2018, 8, 14–19. [Google Scholar] [CrossRef]
- Deisseroth, K.; Feng, G.; Majewska, A.K.; Miesenböck, G.; Ting, A.; Schnitzer, M.J. Next-generation optical technologies for illuminating genetically targeted brain circuits. J. Neurosci. 2006, 26, 10380–10386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gradinaru, V.; Mogri, M.; Thompson, K.R.; Henderson, J.M.; Deisseroth, K. Optical deconstruction of parkinsonian neural circuitry. Science 2009, 324, 354–359. [Google Scholar] [CrossRef] [Green Version]
- Rossi, M.A.; Calakos, N.; Yin, H.H. Spotlight on movement disorders: What optogenetics has to offer. Mov. Disord. 2015, 30, 624–631. [Google Scholar] [CrossRef] [Green Version]
- Vazey, E.M.; Aston-Jones, G. New tricks for old dogmas: Optogenetic and designer receptor insights for Parkinson’s disease. Brain Res. 2013, 1511, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Herman, A.M.; Huang, L.; Murphey, D.K.; Garcia, I.; Arenkiel, B.R. Cell type-specific and time-dependent light exposure contribute to silencing in neurons expressing Channelrhodopsin-2. Elife 2014, 3, e01481. [Google Scholar] [CrossRef]
- Raimondo, J.V.; Kay, L.; Ellender, T.J.; Akerman, C.J. Optogenetic silencing strategies differ in their effects on inhibitory synaptic transmission. Nat. Neurosci. 2012, 15, 1102–1104. [Google Scholar] [CrossRef] [Green Version]
- Edgerton, J.R.; Jaeger, D. Optogenetic activation of nigral inhibitory inputs to motor thalamus in the mouse reveals classic inhibition with little potential for rebound activation. Front. Cell. Neurosci. 2014, 8, 36. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montemurro, N.; Aliaga, N.; Graff, P.; Escribano, A.; Lizana, J. New Targets and New Technologies in the Treatment of Parkinson’s Disease: A Narrative Review. Int. J. Environ. Res. Public Health 2022, 19, 8799. https://doi.org/10.3390/ijerph19148799
Montemurro N, Aliaga N, Graff P, Escribano A, Lizana J. New Targets and New Technologies in the Treatment of Parkinson’s Disease: A Narrative Review. International Journal of Environmental Research and Public Health. 2022; 19(14):8799. https://doi.org/10.3390/ijerph19148799
Chicago/Turabian StyleMontemurro, Nicola, Nelida Aliaga, Pablo Graff, Amanda Escribano, and Jafeth Lizana. 2022. "New Targets and New Technologies in the Treatment of Parkinson’s Disease: A Narrative Review" International Journal of Environmental Research and Public Health 19, no. 14: 8799. https://doi.org/10.3390/ijerph19148799