
Effect of Increasing C/N Ratio on Performance and Microbial Community Structure in a Membrane Bioreactor with a High Ammonia Load

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Set-Up
2.2. Batch Experiments
2.3. Analytical Methods
2.4. DNA Extraction and PCR
2.5. Sequencing and Analysis
3. Results and Discussion
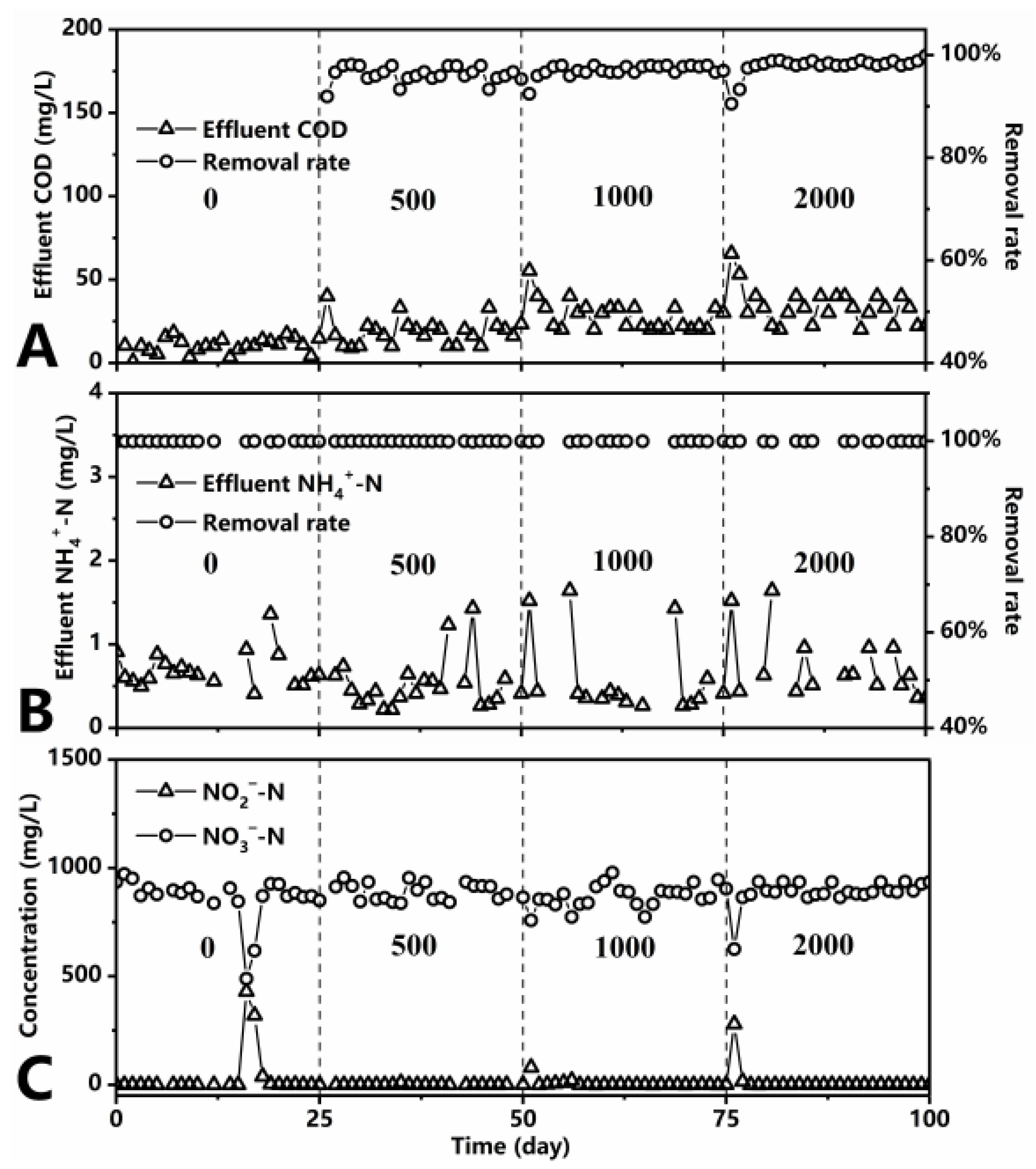
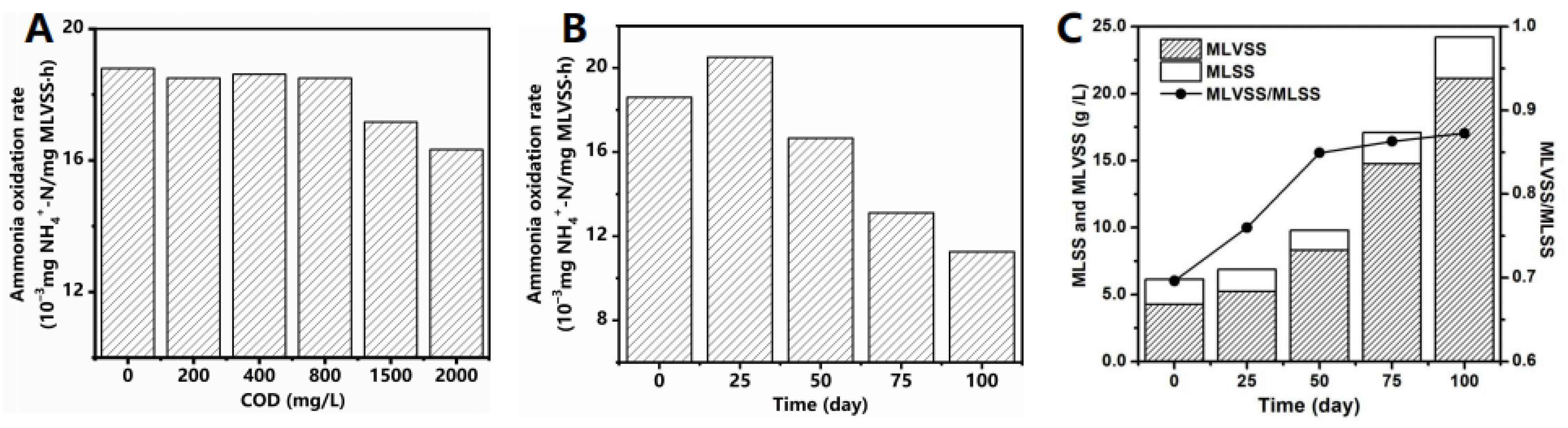
3.1. Effect of C/N Ratio on Performance and Sludge Characteristics of MBR
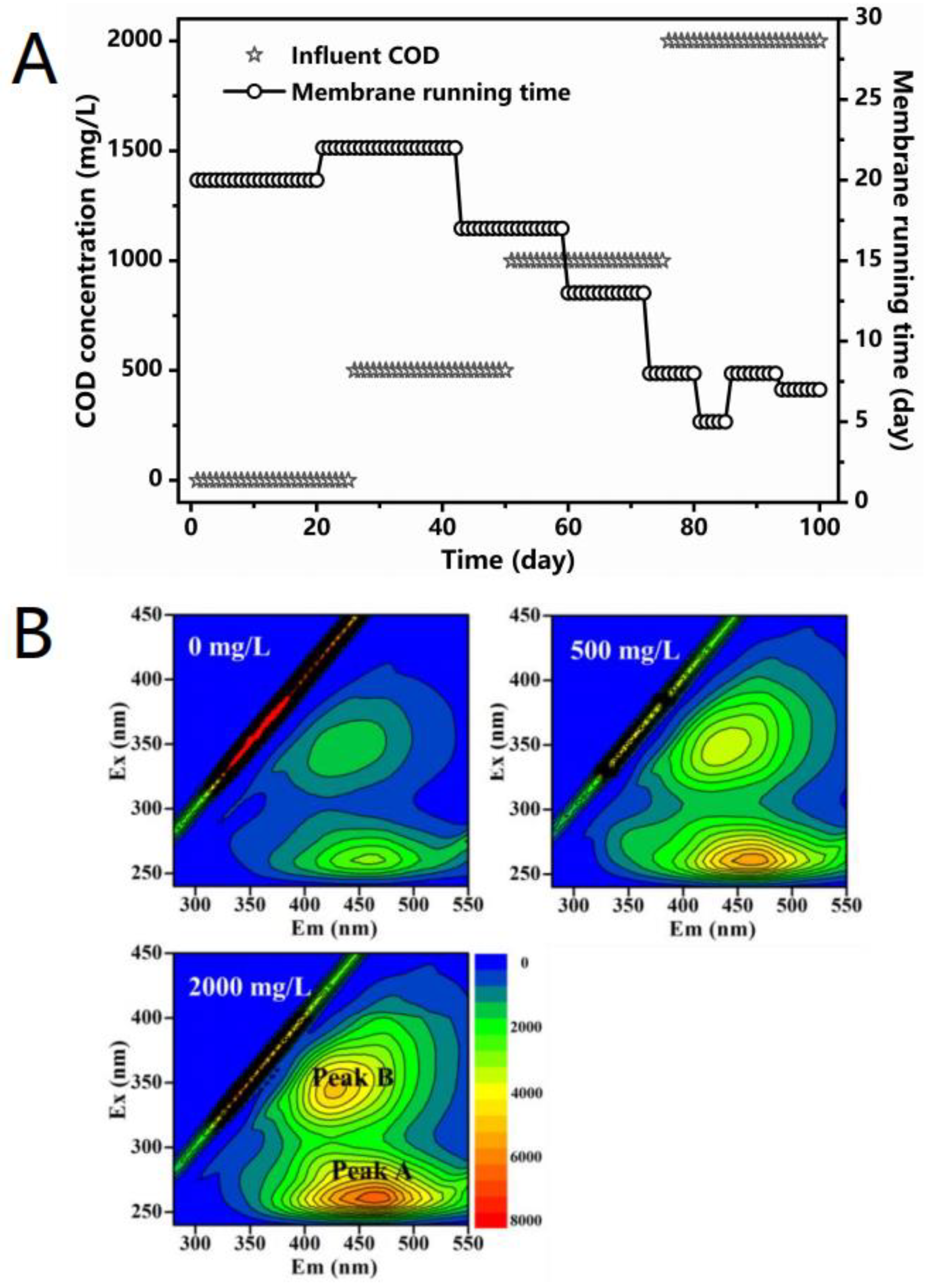
3.2. Membrane Fouling and SMP
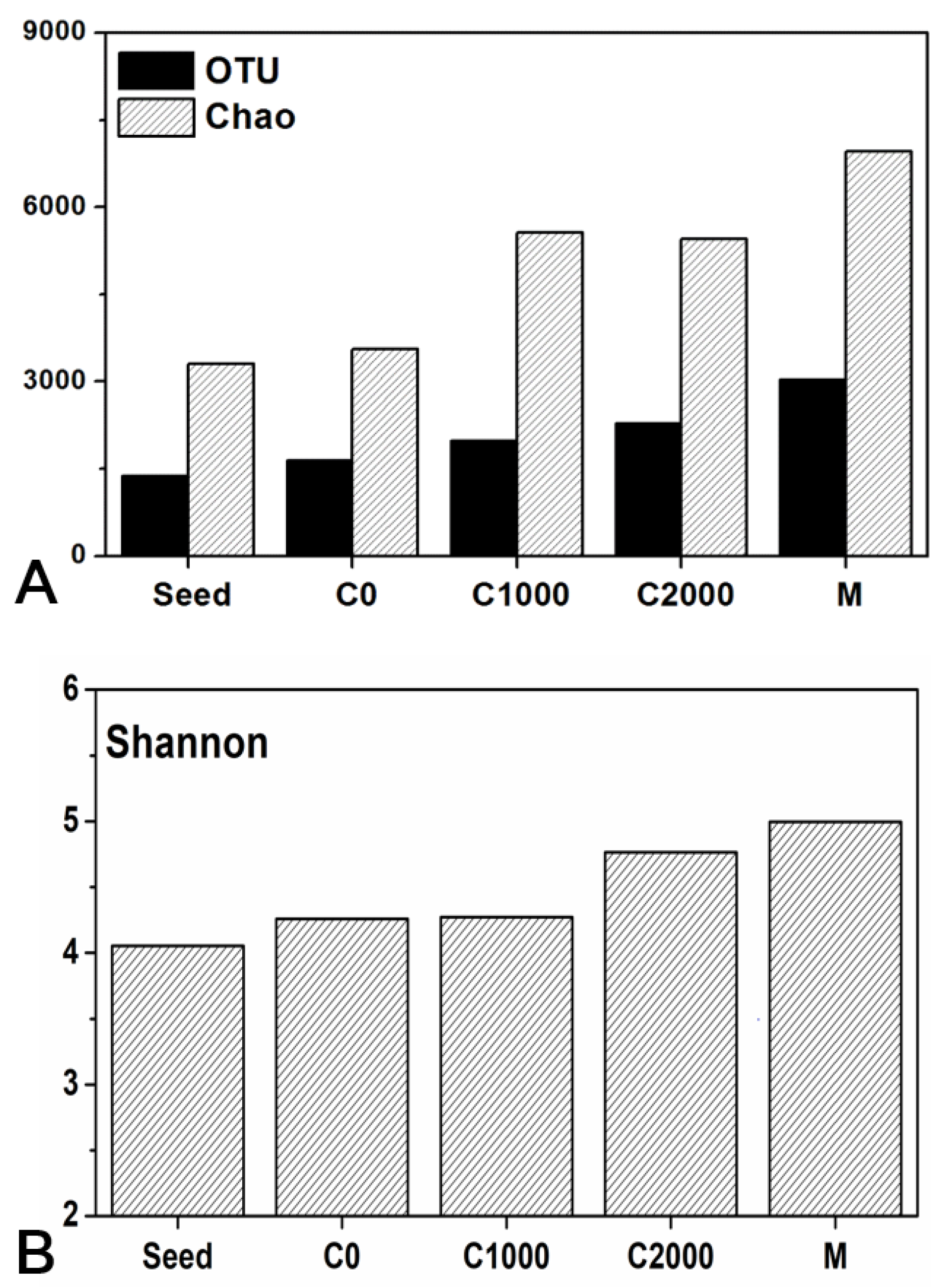
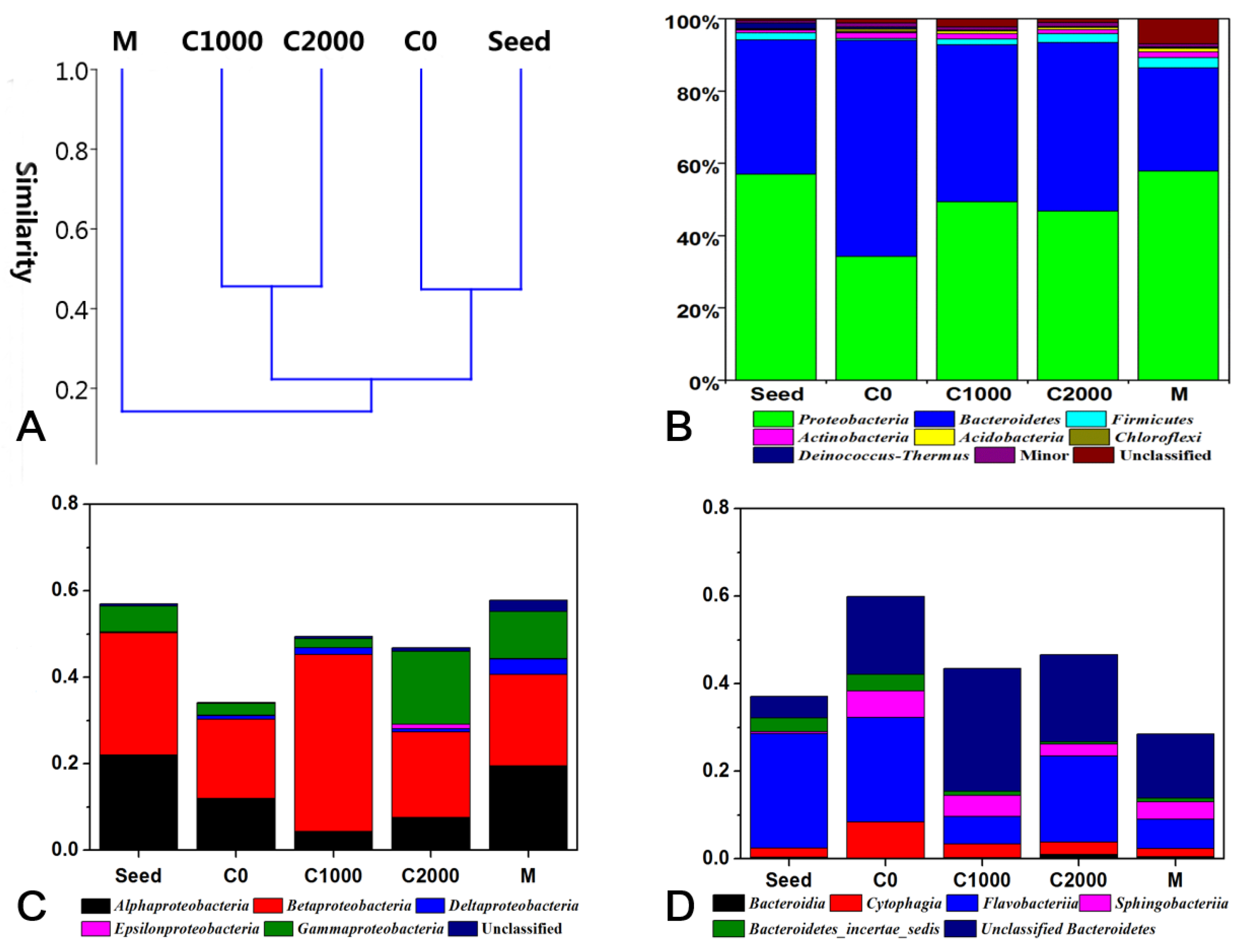
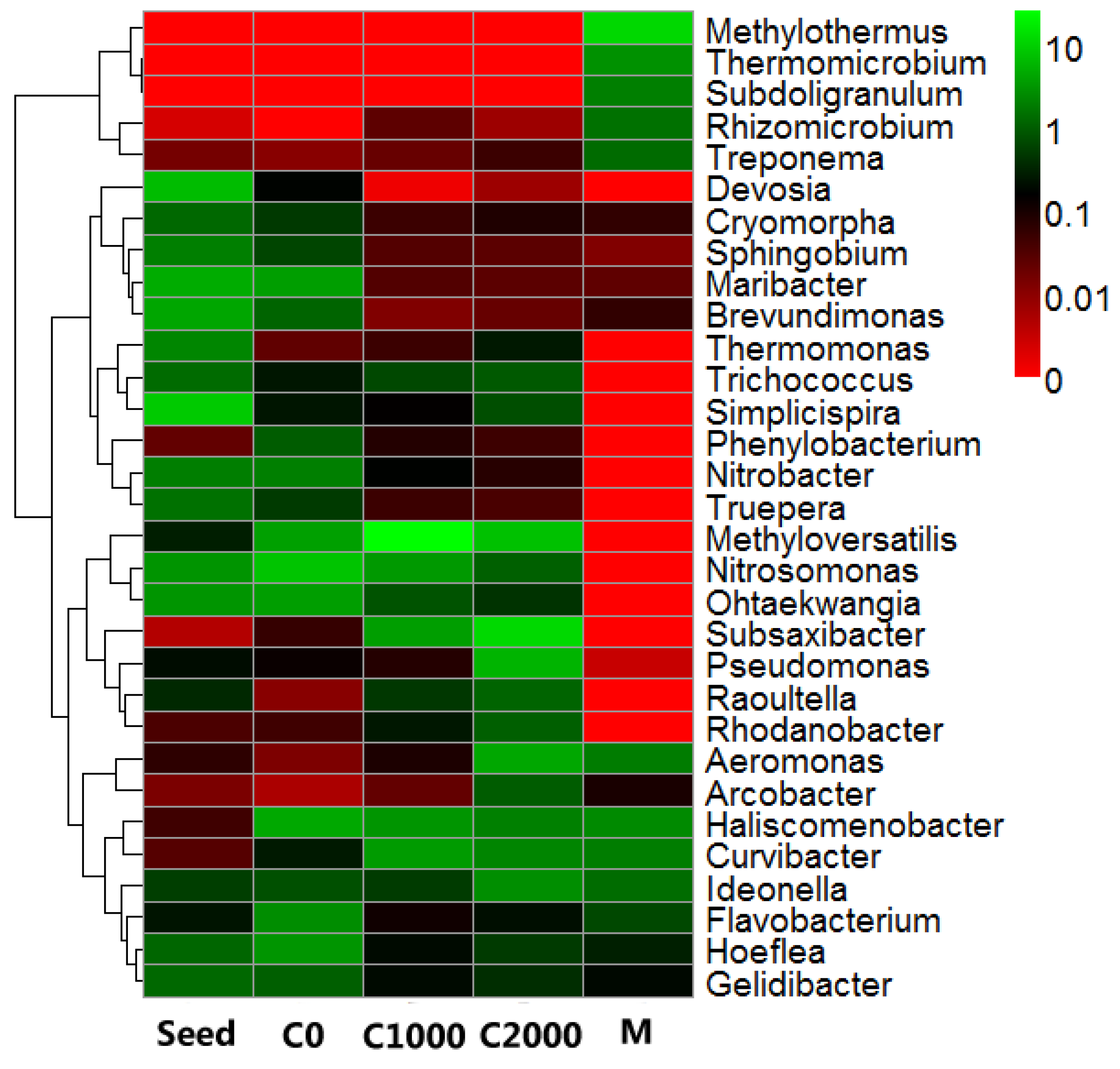
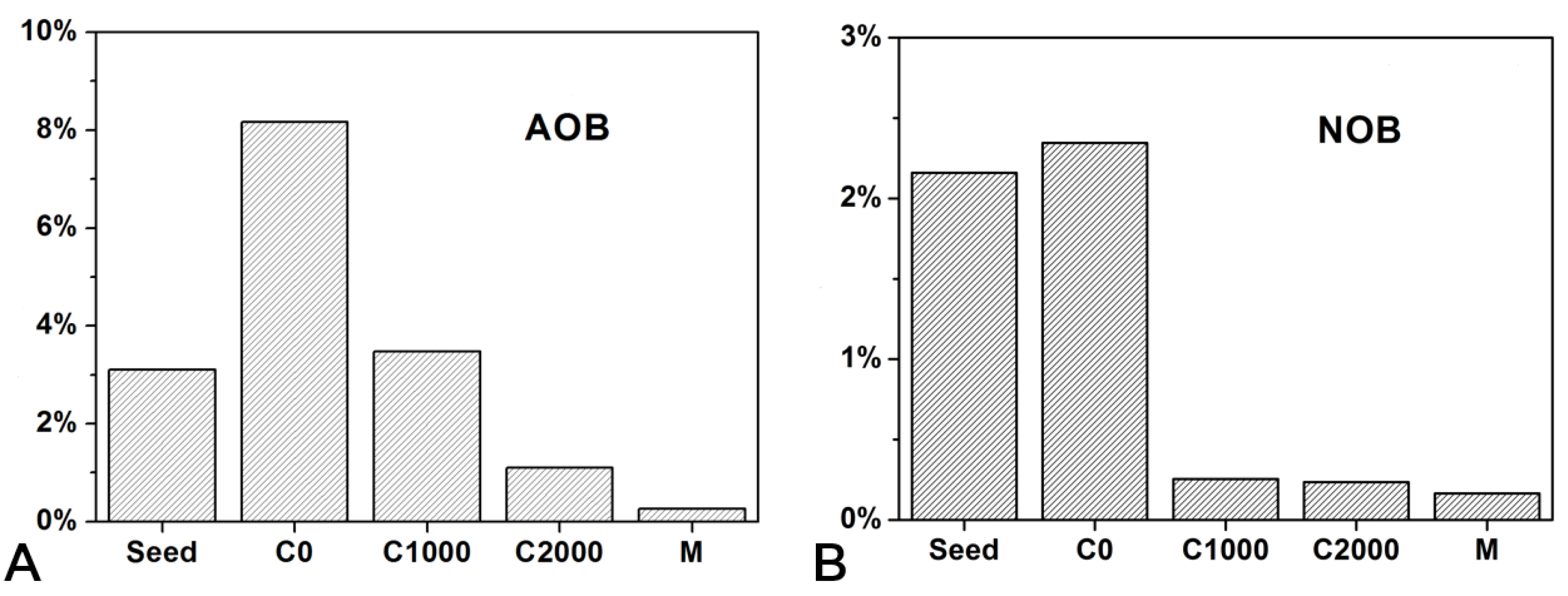
3.3. Microbial Community Structure
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gao, J.; Zhu, T.; Liu, C.; Zhang, J.; Gao, J.; Zhang, J.; Cai, M.; Li, Y. Ammonium removal characteristics of heterotrophic nitrifying bacterium Pseudomonas stutzeri GEP-01 with potential for treatment of ammonium-rich wastewater. Bioprocess Biosyst. Eng. 2020, 43, 959–969. [Google Scholar] [CrossRef]
- Chai, L.Y.; Ali, M.; Min, X.B.; Song, Y.X.; Tang, C.J.; Wang, H.Y.; Yu, C.; Yang, Z.H. Partial nitrification in an air-lift reactor with long-term feeding of increasing ammonium concentrations. Bioresour. Technol. 2015, 185, 134–142. [Google Scholar] [CrossRef]
- Camargo, J.A.; Alonso, A. Ecological and toxicological effects of inorganic nitrogen pollution in aquatic ecosystems: A global assessment. Environ. Int. 2006, 32, 831–849. [Google Scholar] [CrossRef] [PubMed]
- Rabalais, N.N. Nitrogen in aquatic ecosystems. Ambio 2002, 31, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Martinez, A.; Pesciaroli, C.; Martinez-Toledo, M.V.; Hontoria, E.; Gonzalez-Lopez, J.; Osorio, F. Study of nitrifying microbial communities in a partial-nitritation bioreactor. Ecol. Eng. 2014, 64, 443–450. [Google Scholar] [CrossRef]
- Yao, Q.; Peng, D.; Wang, B.; Chen, Y.; Li, J.; Zhao, Q.; Wang, B. Effect of free ammonium and free nitrous acid on the activity, aggregate morphology and extracellular polymeric substance distribution of ammonium oxidizing bacteria in partial nitrification. J. Biosci. Bioeng. 2017, 124, 319–326. [Google Scholar] [CrossRef]
- Vadivelu, V.M.; Keller, J.; Yuan, Z. Effect of free ammonia on the respiration and growth processes of an enriched Nitrobacter culture. Water Res. 2007, 41, 826–834. [Google Scholar] [CrossRef]
- Gil, K.I.; Choi, E.S. Modelling of inhibition of nitrite oxidation in biological nitritation processes by free ammonia. Biotechnol. Lett. 2001, 23, 2021–2026. [Google Scholar] [CrossRef]
- Campos, J.L.; Garrido, J.M.; Mosquera-Corral, A.; Mendez, R. Stability of a nitrifying activated sludge reactor. Biochem. Eng. J. 2007, 35, 87–92. [Google Scholar] [CrossRef]
- Chen, F.Y.; Liu, Y.Q.; Tay, J.H.; Ning, P. Rapid formation of nitrifying granules treating high-strength ammonium wastewater in a sequencing batch reactor. Appl. Microbiol. Biotechnol. 2015, 99, 4445–4452. [Google Scholar] [CrossRef] [PubMed]
- Vejmelkova, D.; Sorokin, D.Y.; Abbas, B.; Kovaleva, O.L.; Kleerebezem, R.; Kampschreur, M.J.; Muyzer, G.; van Loosdrecht, M.C. Analysis of ammonia-oxidizing bacteria dominating in lab-scale bioreactors with high ammonium bicarbonate loading. Appl. Microbiol. Biotechnol. 2012, 93, 401–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Zhu, G. Biological nitrogen removal with nitrification and denitrification via nitrite pathway. Appl. Microbiol. Biotechnol. 2006, 73, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Shiskowski, D.M.; Mavinic, D.S. The influence of nitrite and pH (nitrous acid) on aerobic-phase, autotrophic N2O generation in a wastewater treatment bioreactor. J. Environ. Eng. Sci. 2006, 5, 273–283. [Google Scholar] [CrossRef]
- Wang, Z.; Luo, G.; Li, J.; Chen, S.Y.; Li, Y.; Li, W.T.; Li, A.M. Response of performance and ammonia oxidizing bacteria community to high salinity stress in membrane bioreactor with elevated ammonia loading. Bioresour. Technol. 2016, 216, 714–721. [Google Scholar] [CrossRef]
- Anthonisen, A.C.; Loehr, R.C.; Prakasam, T.B.; Srinath, E.G. Inhibition of nitrification by ammonia and nitrous acid. J. Water Pollut. Control. Fed. 1976, 48, 835–852. [Google Scholar]
- Kim, D.J.; Lee, D.I.; Keller, J. Effect of temperature and free ammonia on nitrification and nitrite accumulation in landfill leachate and analysis of its nitrifying bacterial community by FISH. Bioresour. Technol. 2006, 97, 459–468. [Google Scholar] [CrossRef]
- Liu, Y.; Ngo, H.H.; Guo, W.; Peng, L.; Wang, D.; Ni, B. The roles of free ammonia (FA) in biological wastewater treatment processes: A review. Environ. Int. 2019, 123, 10–19. [Google Scholar] [CrossRef]
- Le-Clech, P.; Chen, V.; Fane, T.A.G. Fouling in membrane bioreactors used in wastewater treatment. J. Membr. Sci. 2006, 284, 17–53. [Google Scholar] [CrossRef]
- Meng, F.; Zhang, S.; Oh, Y.; Zhou, Z.; Shin, H.S.; Chae, S.R. Fouling in membrane bioreactors: An updated review. Water Res. 2017, 114, 151–180. [Google Scholar] [CrossRef]
- Dvorak, L.; Svojitka, J.; Wanner, J.; Wintgens, T. Nitrification performance in a membrane bioreactor treating industrial wastewater. Water Res. 2013, 47, 4412–4421. [Google Scholar] [CrossRef] [PubMed]
- Ji, G.D.; Zhi, W.; Tan, Y.F. Association of nitrogen micro-cycle functional genes in subsurface wastewater infiltration systems. Ecol. Eng. 2012, 44, 269–277. [Google Scholar] [CrossRef]
- Zhi, W.; Ji, G. Quantitative response relationships between nitrogen transformation rates and nitrogen functional genes in a tidal flow constructed wetland under C/N ratio constraints. Water Res. 2014, 64, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Li, J.; Wang, R. Nitrogen removal performance and microbial community structure dynamics response to carbon nitrogen ratio in a compact suspended carrier biofilm reactor. Ecol. Eng. 2008, 32, 256–262. [Google Scholar] [CrossRef]
- Ballinger, S.J.; Head, I.M.; Curtis, T.P.; Godley, A.R. The effect of C/N ratio on ammonia oxidising bacteria community structure in a laboratory nitrification-denitrification reactor. Water Sci. Technol. 2002, 46, 543–550. [Google Scholar] [CrossRef]
- Ma, J.; Wang, Z.; Zhu, C.; Liu, S.; Wang, Q.; Wu, Z. Analysis of nitrification efficiency and microbial community in a membrane bioreactor fed with low COD/N-ratio wastewater. PLoS ONE 2013, 8, e63059. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.F.; Tay, J.H.; Liu, Y. Respirometric activities of heterotrophic and nitrifying populations in aerobic granules developed at different substrate N/COD ratios. Curr. Microbiol. 2004, 49, 42–46. [Google Scholar] [CrossRef]
- Di Trapani, D.; Di Bella, G.; Mannina, G.; Torregrossa, M.; Viviani, G. Effect of C/N shock variation on the performances of a moving bed membrane bioreactor. Bioresour. Technol. 2015, 189, 250–257. [Google Scholar] [CrossRef] [Green Version]
- Cydzik-Kwiatkowska, A.; Zielinska, M.; Bernat, K.; Wojnowska-Baryla, I.; Truchan, T. Treatment of high-ammonium anaerobic digester supernatant by aerobic granular sludge and ultrafiltration processes. Chemosphere 2013, 90, 2208–2215. [Google Scholar] [CrossRef]
- Kim, J.H.; Guo, X.J.; Park, H.S. Comparison study of the effects of temperature and free ammonia concentration on nitrification and nitrite accumulation. Process Biochem. 2008, 43, 154–160. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, H.; Yang, F.; Liu, S.; Fu, Z.; Chen, H. Start-up of the Anammox process from the conventional activated sludge in a membrane bioreactor. Bioresour. Technol. 2009, 100, 2501–2506. [Google Scholar] [CrossRef]
- Yang, C.H.; Crowley, D.E. Rhizosphere microbial community structure in relation to root location and plant iron nutritional status. Appl. Environ. Microbiol. 2000, 66, 345–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; McLenachan, P.A.; Biggs, P.J.; Winder, L.H.; Schoenfeld, B.I.; Narayan, V.V.; Phiri, B.J.; Lockhart, P.J. Environmental bio-monitoring with high-throughput sequencing. Brief. Bioinform. 2013, 14, 575–588. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Zhang, Y.; Ren, H.Q.; Geng, J.J.; Xu, K.; Huang, H.; Ding, L.L. Physicochemical characteristics and microbial community evolution of biofilms during the start-up period in a moving bed biofilm reactor. Bioresour. Technol. 2015, 180, 345–351. [Google Scholar] [CrossRef]
- Zheng, H.L.; Liu, M.Z.; Lu, Q.; Wu, X.D.; Ma, Y.W.; Cheng, Y.L.; Addy, M.; Liu, Y.H.; Ruan, R. Balancing carbon/nitrogen ratio to improve nutrients removal and algal biomass production in piggery and brewery wastewaters. Bioresour. Technol. 2018, 249, 479–486. [Google Scholar] [CrossRef]
- Zhao, C.Q.; Chen, W.Y. A review for tannery wastewater treatment: Some thoughts under stricter discharge requirements. Environ. Sci. Pollut. Res. 2019, 26, 26102–26111. [Google Scholar] [CrossRef]
- Jang, D.; Hwang, Y.; Shin, H.; Lee, W. Effects of salinity on the characteristics of biomass and membrane fouling in membrane bioreactors. Bioresour. Technol. 2013, 141, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.P.; Zhang, T. Characterization of soluble microbial products (SMP) under stressful conditions. Water Res. 2010, 44, 5499–5509. [Google Scholar] [CrossRef]
- Harris, J.K.; El Kasmi, K.C.; Anderson, A.L.; Devereaux, M.W.; Fillon, S.A.; Robertson, C.E.; Wagner, B.D.; Stevens, M.J.; Pace, N.R.; Sokol, R.J. Specific microbiome changes in a mouse model of parenteral nutrition associated liver injury and intestinal inflammation. PLoS ONE 2014, 9, e110396. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Miao, Y.; Wu, B.; Ye, L.; Yu, H.; Liu, S.; Zhang, X.X. Correlation between microbial community structure and biofouling as determined by analysis of microbial community dynamics. Bioresour. Technol. 2015, 197, 99–105. [Google Scholar] [CrossRef]
- Zhang, T.; Shao, M.F.; Ye, L. 454 pyrosequencing reveals bacterial diversity of activated sludge from 14 sewage treatment plants. ISME J. 2012, 6, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, X.X.; Lu, X.; Liu, B.; Li, Y.; Long, C.; Li, A. Abundance and diversity of bacterial nitrifiers and denitrifiers and their functional genes in tannery wastewater treatment plants revealed by high-throughput sequencing. PLoS ONE 2014, 9, e113603. [Google Scholar] [CrossRef] [PubMed]
- Wiesmann, U. Biological nitrogen removal from wastewater. Adv. Biochem. Eng. Biotechnol. 1994, 51, 113–154. [Google Scholar] [CrossRef]
- Blackburne, R.; Yuan, Z.; Keller, J. Partial nitrification to nitrite using low dissolved oxygen concentration as the main selection factor. Biodegradation 2008, 19, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.W.; Yao, Y.; Meng, F.G. Reactor performance and microbial ecology of a nitritation membrane bioreactor. J. Membr. Sci. 2014, 462, 139–146. [Google Scholar] [CrossRef]
- Guo, J.; Peng, Y.; Wang, S.; Zheng, Y.; Huang, H.; Wang, Z. Long-term effect of dissolved oxygen on partial nitrification performance and microbial community structure. Bioresour. Technol. 2009, 100, 2796–2802. [Google Scholar] [CrossRef]
- He, Z.J.; Mahmud, S.; Zhao, S.F.; Yang, Y.; Zhu, L.J.; Zhao, Y.B.; Zeng, Q.Y.; Xiong, Z.; Hu, C. Hierarchically Active Poly(vinylidene fluoride) Membrane Fabricated by In Situ Generated Zero-Valent Iron for Fouling Reduction. ACS Appl. Mater. Interfaces 2020, 12, 10993–11004. [Google Scholar] [CrossRef]
- Chen, Z.; Mahmud, S.; Cai, L.; He, Z.; Yang, Y.; Zhang, L.; Zhao, S.; Xiong, Z. Hierarchical poly(vinylidene fluoride)/active carbon composite membrane with self-confining functional carbon nanotube layer for intractable wastewater remediation. J. Membr. Sci. 2020, 603, 118041. [Google Scholar] [CrossRef]
- Li, C.; Cabassud, C.; Reboul, B.; Guigui, C. Effects of pharmaceutical micropollutants on the membrane fouling of a submerged MBR treating municipal wastewater: Case of continuous pollution by carbamazepine. Water Res. 2015, 69, 183–194. [Google Scholar] [CrossRef]
- Feng, S.P.; Zhang, N.N.; Liu, H.C.; Du, X.L.; Liu, Y.L.; Lin, H. The effect of COD/N ratio on process performance and membrane fouling in a submerged bioreactor. Desalination 2012, 285, 232–238. [Google Scholar] [CrossRef]
- Barker, D.J.; Stuckey, D.C. A review of soluble microbial products (SMP) in wastewater treatment systems. Water Res. 1999, 33, 3063–3082. [Google Scholar] [CrossRef]
- Liang, S.; Liu, C.; Song, L. Soluble microbial products in membrane bioreactor operation: Behaviors, characteristics, and fouling potential. Water Res. 2007, 41, 95–101. [Google Scholar] [CrossRef]
- Ni, B.J.; Rittmann, B.E.; Yu, H.Q. Soluble microbial products and their implications in mixed culture biotechnology. Trends Biotechnol. 2011, 29, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Jarusutthirak, C.; Amy, G. Role of soluble microbial products (SMP) in membrane fouling and flux decline. Environ. Sci. Technol. 2006, 40, 969–974. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.K.; Lin, C.F.; Lateef, S.K.; Panchangam, S.C.; Hong, P.K.A.; Yang, P.Y. The effect of soluble microbial products on membrane fouling in a fixed carrier biological system. Sep. Purif. Technol. 2010, 72, 98–104. [Google Scholar] [CrossRef]
- Shi, Y.H.; Huang, J.H.; Zeng, G.M.; Gu, Y.L.; Hu, Y.; Tang, B.; Zhou, J.X.; Yang, Y.; Shi, L.X. Evaluation of soluble microbial products (SMP) on membrane fouling in membrane bioreactors (MBRs) at the fractional and overall level: A review. Rev. Environ. Sci. Bio. 2018, 17, 71–85. [Google Scholar] [CrossRef]
- Kimura, K.; Naruse, T.; Watanabe, Y. Changes in characteristics of soluble microbial products in membrane bioreactors associated with different solid retention times: Relation to membrane fouling. Water Res. 2009, 43, 1033–1039. [Google Scholar] [CrossRef]
- Head, I.M.; Hiorns, W.D.; Embley, T.M.; McCarthy, A.J.; Saunders, J.R. The phylogeny of autotrophic ammonia-oxidizing bacteria as determined by analysis of 16S ribosomal RNA gene sequences. J. Gen. Microbiol. 1993, 139 Pt 6, 1147–1153. [Google Scholar] [CrossRef] [Green Version]
- Teske, A.; Alm, E.; Regan, J.M.; Toze, S.; Rittmann, B.E.; Stahl, D.A. Evolutionary relationships among ammonia- and nitrite-oxidizing bacteria. J. Bacteriol. 1994, 176, 6623–6630. [Google Scholar] [CrossRef] [Green Version]
- Thomas, F.; Hehemann, J.H.; Rebuffet, E.; Czjzek, M.; Michel, G. Environmental and gut bacteroidetes: The food connection. Front. Microbiol. 2011, 2, 93. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhang, C.; Hu, C.; Liu, H.; Bai, Y.; Qu, J. Sulfur-based mixotrophic denitrification corresponding to different electron donors and microbial profiling in anoxic fluidized-bed membrane bioreactors. Water Res. 2015, 85, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Koops, H.P.; Pommerening-Roser, A. Distribution and ecophysiology of the nitrifying bacteria emphasizing cultured species. FEMS Microbiol. Ecol. 2001, 37, 1–9. [Google Scholar] [CrossRef]
- Liang, Y.; Li, D.; Zhang, X.; Zeng, H.; Yang, Z.; Cui, S.; Zhang, J. Nitrogen removal and microbial characteristics in CANON biofilters fed with different ammonia levels. Bioresour. Technol. 2014, 171, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Ye, L.; Tong, A.H.; Shao, M.F.; Lok, S. Ammonia-oxidizing archaea and ammonia-oxidizing bacteria in six full-scale wastewater treatment bioreactors. Appl. Microbiol. Biotechnol. 2011, 91, 1215–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Gedalanga, P.B.; Asvapathanagul, P.; Olson, B.H. Influence of physicochemical and operational parameters on Nitrobacter and Nitrospira communities in an aerobic activated sludge bioreactor. Water Res. 2010, 44, 4351–4358. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Time (Day) | COD (mg/L) | NH4+-N (mg/L) | C/N Ratio | ALR (gNH4+-N/L·Day) | HRT (h) | DO (mg/L) |
|---|---|---|---|---|---|---|---|
| I | 1–25 | 0 | 1000 | 0 | 1.71 | 14 | 2–6 |
| II | 26–50 | 500 | 1000 | 0.5 | 1.71 | 14 | 2–6 |
| III | 51–75 | 1000 | 1000 | 1 | 1.71 | 14 | 2–6 |
| IV | 75–100 | 2000 | 1000 | 2 | 1.71 | 14 | 2–6 |
| Effluent Parameters (mg/L) | Mean ± SD | p-Value | |||
|---|---|---|---|---|---|
| C/N = 0 | C/N = 0.5 | C/N = 1 | C/N = 2 | ||
| COD | 10.39 ± 4.54 | 18.88 ± 7.94 | 28.53 ± 8.67 | 32.20 ± 8.56 | 0.000 |
| NH4+-N | 0.70 ± 0.21 | 0.52 ± 0.30 | 0.59 ± 0.47 | 0.68 ± 0.34 | 0.332 |
| NO2−-N | 2.69 ± 8.21 | 1.01 ± 1.95 | 5.25 ± 16.20 | 1.31± 3.07 | 0.394 |
| NO3−-N | 911.98 ± 34.32 | 911.08 ± 38.38 | 890.50 ± 52.34 | 919.84 ± 25.95 | 0.065 |
| COD (mg/L) | Peak A | Peak B | ||
|---|---|---|---|---|
| Ex/Em (nm) 1 | Intensity 2 | Ex/Em (nm) | Intensity | |
| 0 | 260/460 | 3197 | 345/432 | 1924 |
| 500 | 260/461 | 5880 | 350/437 | 3900 |
| 2000 | 260/464 | 6923 | 345/426 | 5211 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, H.; Deng, Y.; Li, X.; Liu, Y.; Huang, S.; Yang, Y.; Wang, Z.; Hu, C. Effect of Increasing C/N Ratio on Performance and Microbial Community Structure in a Membrane Bioreactor with a High Ammonia Load. Int. J. Environ. Res. Public Health 2021, 18, 8070. https://doi.org/10.3390/ijerph18158070
Xu H, Deng Y, Li X, Liu Y, Huang S, Yang Y, Wang Z, Hu C. Effect of Increasing C/N Ratio on Performance and Microbial Community Structure in a Membrane Bioreactor with a High Ammonia Load. International Journal of Environmental Research and Public Health. 2021; 18(15):8070. https://doi.org/10.3390/ijerph18158070
Chicago/Turabian StyleXu, Huaihao, Yuepeng Deng, Xiuying Li, Yuxian Liu, Shuangqiu Huang, Yunhua Yang, Zhu Wang, and Chun Hu. 2021. "Effect of Increasing C/N Ratio on Performance and Microbial Community Structure in a Membrane Bioreactor with a High Ammonia Load" International Journal of Environmental Research and Public Health 18, no. 15: 8070. https://doi.org/10.3390/ijerph18158070