The Contribution Ratio of Autotrophic and Heterotrophic Metabolism during a Mixotrophic Culture of Chlorella sorokiniana


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgae Strain, Culture Medium and Cultivation
2.2. Analysis of Organic Carbon, Inorganic Carbon, Cell Density and Chlorophyll-a
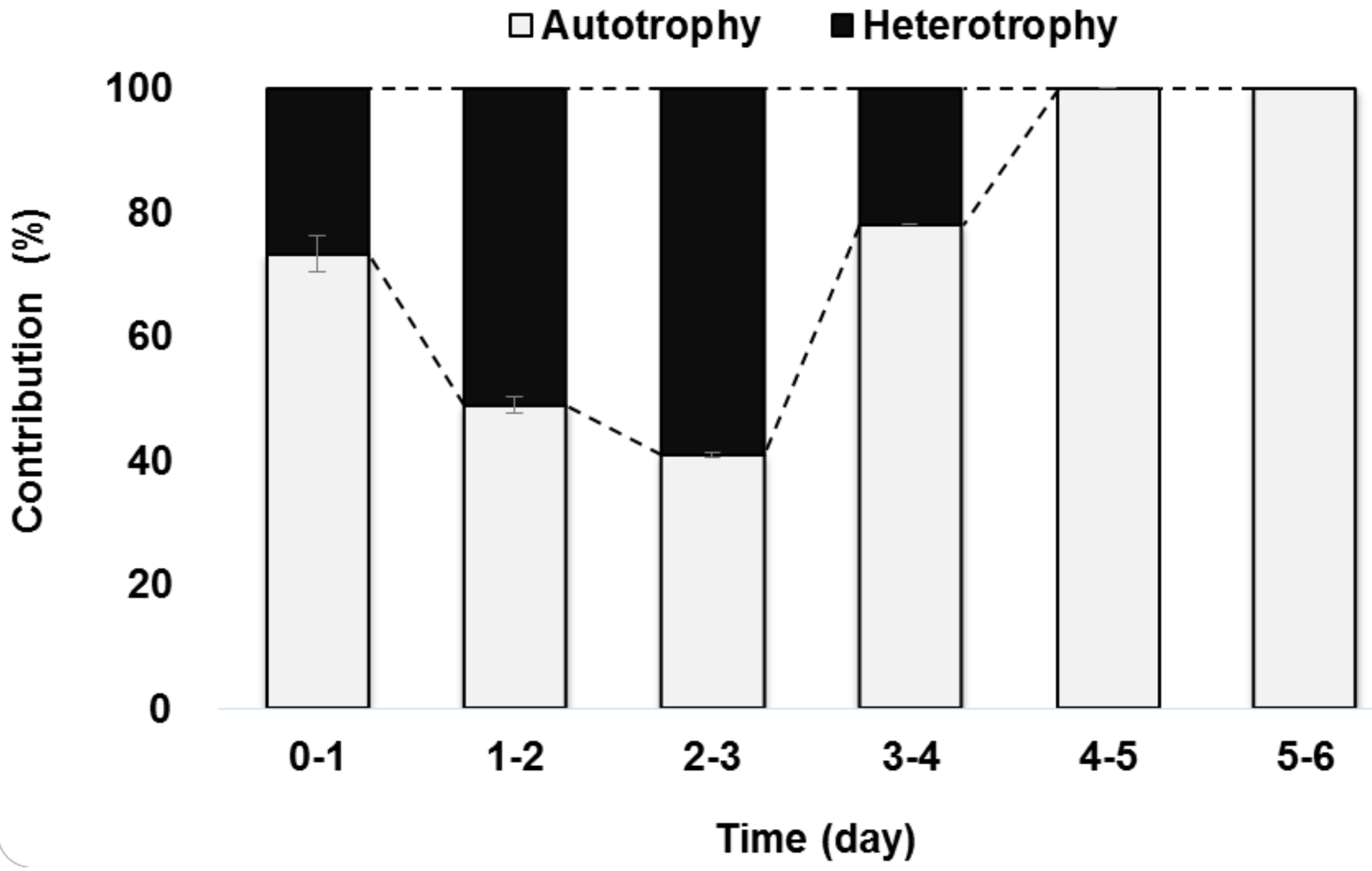
2.3. Contribution Ratio of Autotrophy and Heterotrophy in the Mixotrophic Culture
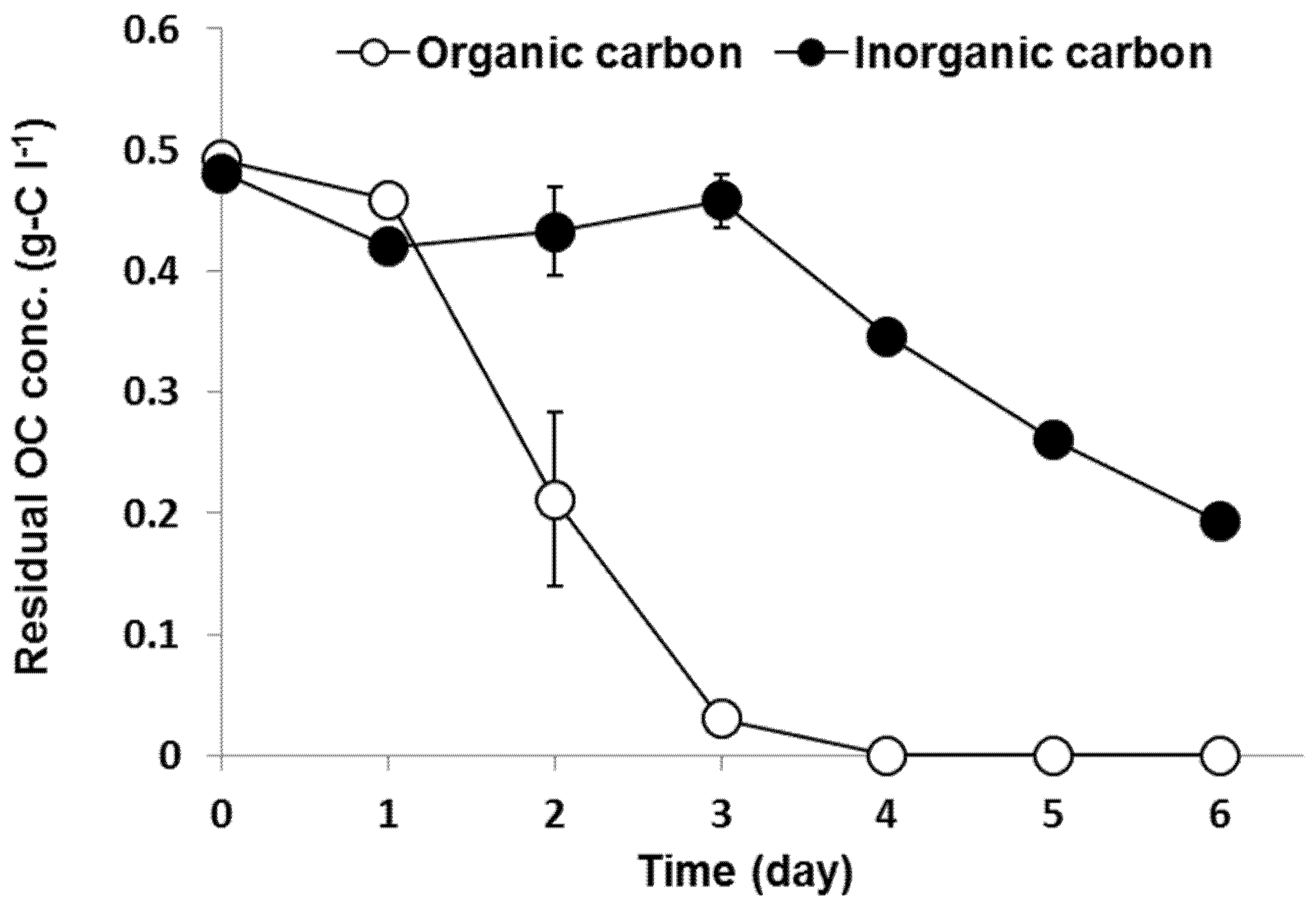
3. Results
3.1. Autotrophy and Heterotrophy Contribution in Mixotrophic Metabolism
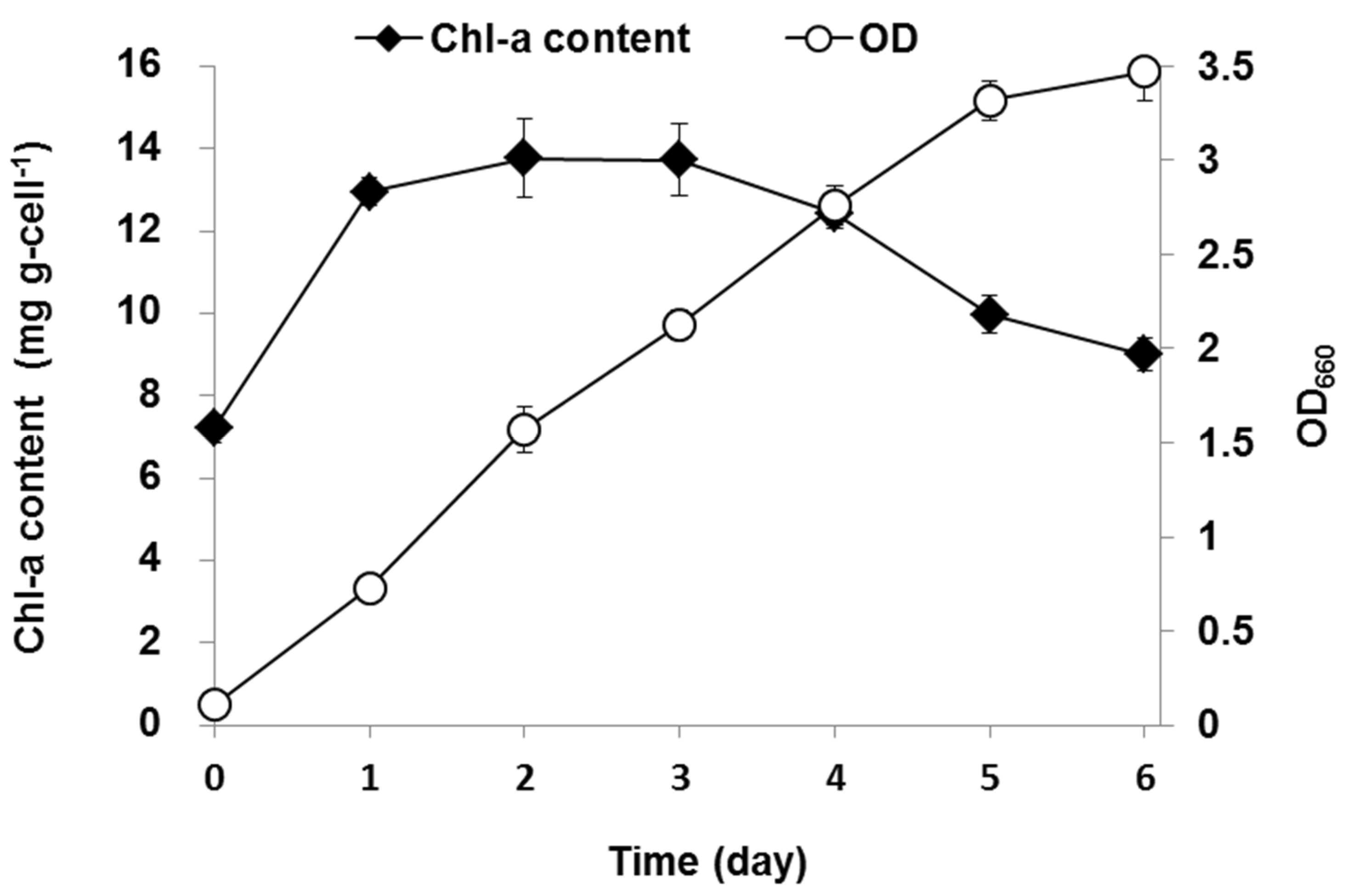
3.2. Variation in Chlorophyll-a Content During the Mixotrophic Culture
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chisti, Y. Biodiesel from microalgae beats bioethanol. Trends Biotechnol. 2008, 26, 126–131. [Google Scholar] [CrossRef]
- Lee, K.; Lee, C.G. Nitrogen removal from wastewaters by microalgae without consuming organic carbon sources. J. Microbiol. Biotechnol. 2002, 12, 979–985. [Google Scholar]
- Kim, S.; Lee, Y.; Hwang, S.J. Removal of nitrogen and phosphorus by Chlorella sorokiniana cultured heterotrophically in ammonia and nitrate. Int. Biodeterior. Biodegrad. 2013, 85, 511–516. [Google Scholar] [CrossRef]
- Han, T.H.; Zhang, S.; Cho, M.H.; Hwang, S.J. Enhancement of volatile fatty acids removal by a co-culture of microalgae and activated sludge. KSCE J. Civ. Eng. 2017, 21, 2106–2112. [Google Scholar] [CrossRef]
- Wollmann, F.; Dietze, S.; Ackermann, J.-U.; Bley, T.; Walther, T.; Steingroewer, J.; Krujatz, F. Microalgae wastewater treatment: Biological and technological approaches. Eng. life Sci. 2019, 19, 860–871. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Nagarajan, D.; Zhang, Q.; Chang, J.-S.; Lee, D.-J. Heterotrophic cultivation of microalgae for pigment production: A review. Biotechnol. Adv. 2018, 36, 54–67. [Google Scholar] [CrossRef]
- Subashchandrabose, S.R.; Ramakrishnan, B.; Megharaj, M.; Venkateswarlu, K.; Naidu, R. Mixotrophic cyanobacteria and microalgae as distinctive biological agents for organic pollutant degradation. Environ. Int. 2013, 51, 59–72. [Google Scholar] [CrossRef]
- Chojnacka, K.; Noworyta, A. Evaluation of Spirulina sp. growth in photoautotrophic, heterotrophic and mixotrophic cultures. Enzyme Microb. Technol. 2004, 34, 461–465. [Google Scholar] [CrossRef]
- Burkholder, J.M.; Glibert, P.M.; Skelton, H.M. Mixotrophy, a major mode of nutrition for harmful algal species in eutrophic waters. Harmful Algae. 2008, 8, 77–93. [Google Scholar] [CrossRef]
- Wan, M.; Liu, P.; Xia, J.; Rosenberg, J.N.; Oyler, G.A.; Betenbaugh, M.J.; Nie, Z.; Qiu, G. The effect of mixotrophy on microalgal growth, lipid content, and expression levels of three pathway genes in Chlorella sorokiniana. Appl. Microbiol. Biotechnol. 2011, 91, 835–844. [Google Scholar] [CrossRef]
- Lee, Y.K.; Ding, S.Y.; Hoe, C.H.; Low, C.S. Mixotrophic growth of Chlorella sorokiniana in outdoor enclosed photobioreactor. J. Appl. Phycol. 1996, 8, 163–169. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, W.; Li, Y.; Yang, W.; Shen, G. Mixotrophic cultivation of Botryococcus braunii. Biomass Bioenergy 2011, 35, 1710–1715. [Google Scholar] [CrossRef]
- Adolf, J.E.; Stoecker, D.K.; Harding, L.W., Jr. The balance of autotrophy and heterotrophy during mixotrophic growth of Carlodinium micrum (Dinophyceae). J. Plankton Res. 2006, 28, 737–751. [Google Scholar] [CrossRef]
- Li, T.; Zheng, Y.; Yu, L.; Chen, S. Mixotrophic cultivation of a Chlorella sorokiniana strain for enhanced biomass and lipid production. Biomass Bioenergy 2014, 66, 204–213. [Google Scholar] [CrossRef]
- Asadi, P.; Rad, H.A.; Qaderi, F. Comparison of Chlorella vulgaris and Chlorella sorokiniana in post treatment of dairy wastewater treatment plant effluents. Environ. Sci. Pollut. Res. 2019, 26, 29473–29489. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Yang, C.; Hua, Q.; Shimizu, K. Energetics and carbon metabolism during growth of microalgal cells under photoautotrophic, mixotrophic and cyclic light-autotrophic/dark-heterotrophic conditions. Biochem. Eng. J. 2000, 6, 87–102. [Google Scholar] [CrossRef]
- Fisher, T.; Berner, T.; Iluz, D.; Dubinsky, Z. The kinetics of the photoacclimation response of Nannochloropsis sp. (Eustigmatophyceae): A study of changes in ultrastructure and PSU density. J. Phycol. 2002, 34, 818–824. [Google Scholar] [CrossRef]
- Aslan, S.; Kapdan, I.K. Batch kinetics of nitrogen and phosphorus removal from synthetic wastewater by algae. Ecol. Eng. 2006, 28, 64–70. [Google Scholar] [CrossRef]
- Dorling, M.; McAuley, P.J.; Hodge, H. Effect of pH on growth and carbon metabolism of maltose-releasing Chlorella (Chlorophyta). Eur. J. Phycol. 1997, 32, 19–24. [Google Scholar] [CrossRef]
- Kamiya, A.; Kowallik, W. Photoinhibition of glucose uptake in Chlorella. Plant. Cell Physiol. 1987, 28, 611–619. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Parameter | Detail |
|---|---|
| Microalgae strain | C. sorokiniana |
| Culture type Organic carbon Inorganic carbon | Mixotrophic culture Glucose: 0.5 g-C L−1 Na2CO3: 0.5 g-C L−1 |
| Artificial wastewater | Modified BG11 (NaNO3: 150 mg-N L−1, K2HPO4: 30 mg-P L−1) |
| pH and Temp. | 8 ± 0.3 and 25 ± 2 °C |
| PPFD | 120 μmolm−2s−1 |
| Working volume | 2 L |
| Mixing rate | 100 rpm |
| Initial inoculum | 0.1 OD |
| Light source | White LED |
| Light/Dark cycle | 24 : 0 (h) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.-E.; Zhang, S.; Han, T.H.; Hwang, S.-J. The Contribution Ratio of Autotrophic and Heterotrophic Metabolism during a Mixotrophic Culture of Chlorella sorokiniana. Int. J. Environ. Res. Public Health 2021, 18, 1353. https://doi.org/10.3390/ijerph18031353
Park J-E, Zhang S, Han TH, Hwang S-J. The Contribution Ratio of Autotrophic and Heterotrophic Metabolism during a Mixotrophic Culture of Chlorella sorokiniana. International Journal of Environmental Research and Public Health. 2021; 18(3):1353. https://doi.org/10.3390/ijerph18031353
Chicago/Turabian StylePark, Jeong-Eun, Shan Zhang, Thi Hiep Han, and Sun-Jin Hwang. 2021. "The Contribution Ratio of Autotrophic and Heterotrophic Metabolism during a Mixotrophic Culture of Chlorella sorokiniana" International Journal of Environmental Research and Public Health 18, no. 3: 1353. https://doi.org/10.3390/ijerph18031353