
Biases in the Detection of Intentionally Poisoned Animals: Public Health and Conservation Implications from a Field Experiment

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
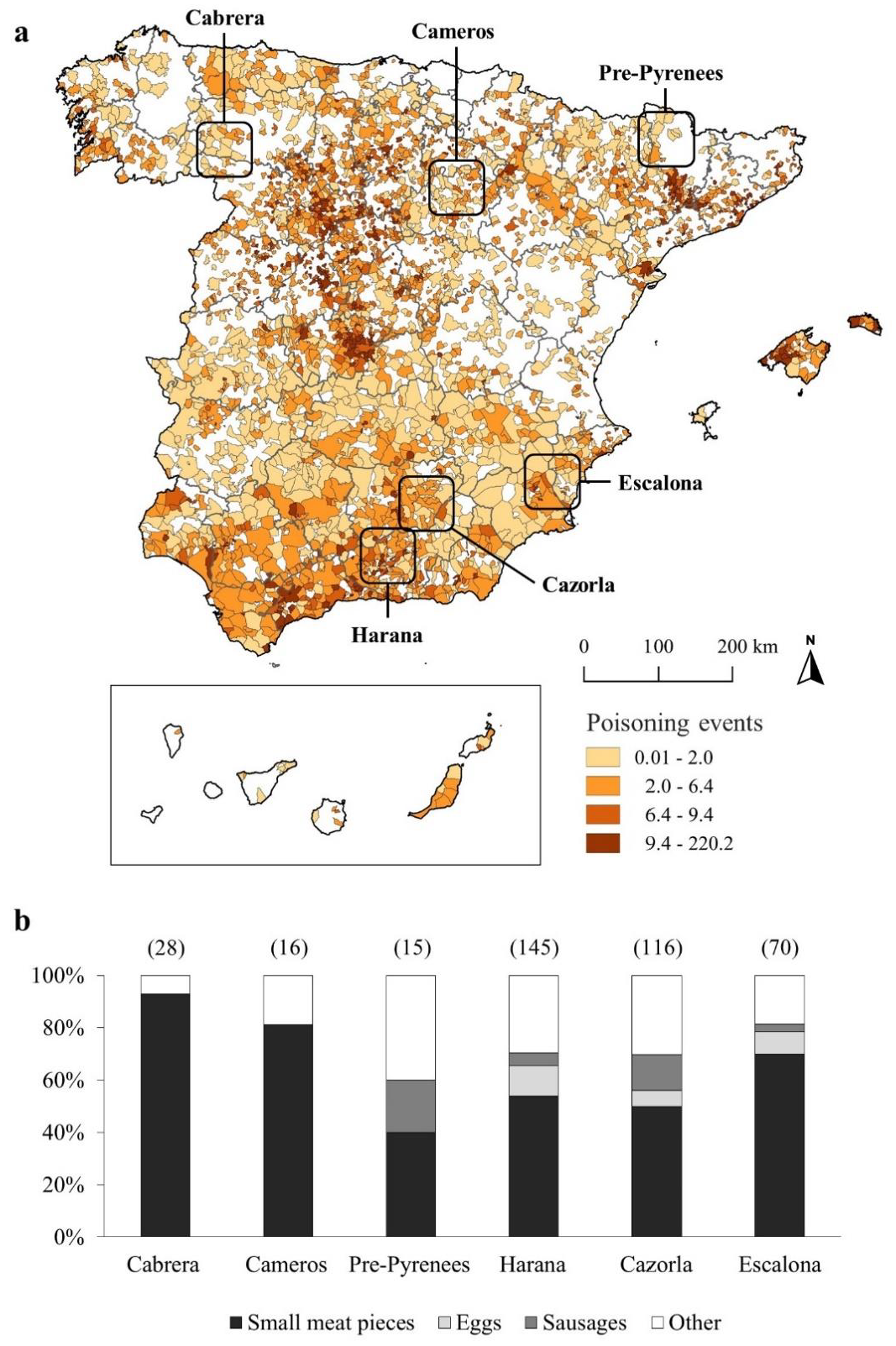
2.1. Study Areas
2.2. Poisoning Database
2.3. Field Experiment
2.4. Ethic Statement
2.5. Statistical Analyses
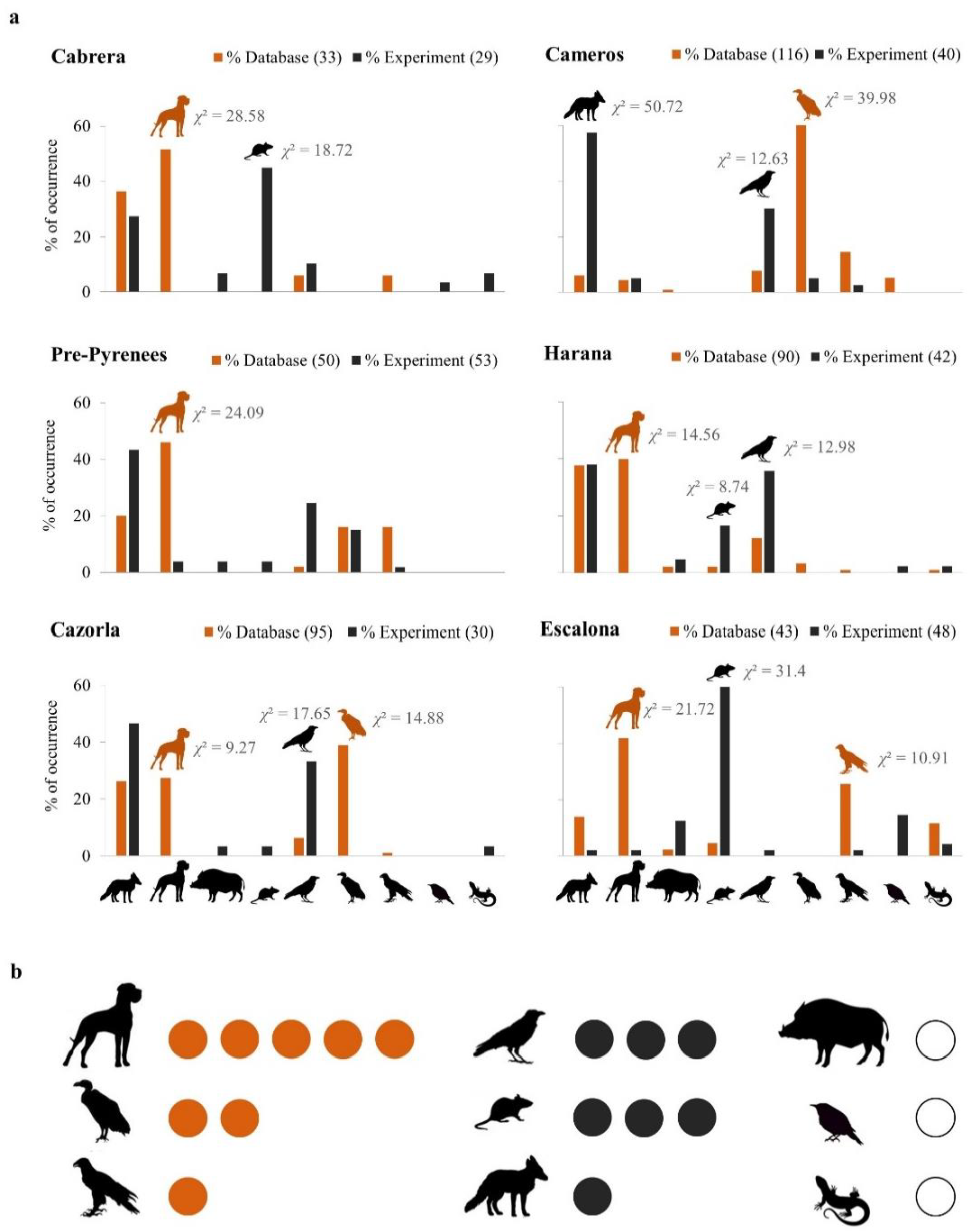
3. Results
4. Discussion
Conclusions and Conservation Implications
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guitart, R.; Sachana, M.; Caloni, F.; Croubels, S.; Vandenbroucke, V.; Berny, P. Animal poisoning in Europe. Part 3: Wildlife. Vet. J. 2010, 183, 260–265. [Google Scholar] [CrossRef]
- Mateo-Tomás, P.; Olea, P.P.; Sánchez-Barbudo, I.S.; Mateo, R. Alleviating human-wildlife conflicts: Identifying the causes and mapping the risk of illegal poisoning of wild fauna. J. Appl. Ecol. 2012, 49, 376–385. [Google Scholar] [CrossRef]
- Ogada, D.L. The power of poison: Pesticide poisoning of Africa’s wildlife. Ann. N. Y. Acad. Sci. 2014, 1322, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Mateo-Tomás, P.; Olea, P.P.; Mínguez, E.; Mateo, R.; Viñuela, J. Direct evidence of poison-driven widespread population decline in a wild vertebrate. Proc. Natl. Acad. Sci. USA 2020, 117, 16418–16423. [Google Scholar] [CrossRef] [PubMed]
- Di Blasio, A.; Bertolini, S.; Gili, M.; Avolio, R.; Leogrande, M.; Ostorero, F.; Ru, G.; Dondo, A.; Zoppi, S. Local context and environment as risk factors for acute poisoning in animals in northwest Italy. Sci. Total. Environ. 2020, 709, 136016. [Google Scholar] [CrossRef] [PubMed]
- Berny, P.; Gaillet, J.R. Acute poisoning of Red Kites (Milvus milvus) in France: Data from the SAGIR network. J. Wildl. Dis. 2008, 44, 417–426. [Google Scholar] [CrossRef] [Green Version]
- Margalida, A. Baits, budget cuts: A deadly mix. Science 2012, 338, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margalida, A.; Mateo, R. Illegal killing of birds in Europe continues. Science 2019, 363, 1161. [Google Scholar] [CrossRef]
- Ogada, D.; Botha, A.; Shaw, P. Ivory poachers and poison: Drivers of Africa’s declining vulture populations. Oryx 2016, 50, 593–596. [Google Scholar] [CrossRef] [Green Version]
- Lozano, J.; Olszańska, A.; Morales-Reyes, Z.; Castro, A.A.; Malo, A.F.; Moleón, M.; Sánchez-Zapata, J.A.; Cortés-Avizanda, A.; von Wehrden, H.; Dorresteijn, I.; et al. Human-carnivore relations: A systematic review. Biol. Conserv. 2019, 237, 480–492. [Google Scholar] [CrossRef]
- Hernández, M.; Margalida, A. Pesticide abuse in Europe: Effects on the Cinereous vulture (Aegypius monachus) population in Spain. Ecotoxicology 2008, 17, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Hernández, M.; Margalida, A. Poison-related mortality effects in the endangered Egyptian vulture (Neophron percnopterus) population in Spain. Eur. J. Wildl. Res. 2009, 55, 415–423. [Google Scholar] [CrossRef]
- Margalida, A.; Ogada, D.; Botha, A. Protect African vultures from poison. Science 2019, 365, 1089–1090. [Google Scholar] [CrossRef] [PubMed]
- Vyas, N.B. Factors influencing estimation of pesticide-related wildlife mortality. Toxicol. Ind. Health 1999, 15, 187–192. [Google Scholar] [CrossRef]
- Mineau, P. Direct losses of birds to pesticides—Beginnings of a quantification. In Bird Conservation Implementation and Integration in the Americas: Proceedings of the Third International Partners in Flight Conference, Asilomar, CA, USA, 20–24 March 2002; Gen. Tech. Rep. PSW-GTR-191; Ralph, C.J., Rich, T.D., Eds.; U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2005; Volume 2, pp. 1065–1070. [Google Scholar]
- Margalida, A.; Heredia, R.; Razin, M.; Hernández, M. Sources of variation in mortality of the Bearded Vulture Gypaetus barbatus in Europe. Bird Conserv. Int. 2008, 18, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Treves, A.; Artelle, K.A.; Darimont, C.T.; Parsons, D.R. Mismeasured mortality: Correcting estimates of wolf poaching in the United States. J. Mammal. 2017, 98, 1256–1264. [Google Scholar] [CrossRef] [Green Version]
- De la Bodega, D.; Cano, C.; Mínguez, E. El Veneno en España. Evolución del Envenenamiento de Fauna Silvestre (1992–2017); SEO/BirdLife, WWF, Eds.; WWF and SEO/BirdLife: Madrid, Spain, 2020; ISBN 978-84-120635-3-0. [Google Scholar]
- Silva, J.P.; Eldridge, J.; Nottinghan, S.; Travagnin, C. Life & Wildlife Crime; Publications Office of the European Union: Luxembourg, 2018. [Google Scholar]
- Durá-Alemañ, C.J.; Morales-Reyes, Z.; Ayerza, P.; De la Bodega, D.; Aguilera-Alcalá, N.; Botella, F.; Jiménez-Peinado, J.; Jiménez, J.; López-Bao, J.V.; Mateo-Tomás, P.; et al. La responsabilidad por el daño ambiental generado en el caso de la lucha contra el uso del veneno en España. Actual. Jurid. Ambient. 2020, 102, 564–576. [Google Scholar]
- Cano, C.; De la Bodega, D.; Ayerza, P.; Mínguez, E. El Veneno en España. Evolución del Envenenamiento de Fauna Silvestre (1992–2013); WWF and SEO/BirdLife: Madrid, Spain, 2016. [Google Scholar]
- González, L.M.; Oria, J.; Sánchez, R.; Margalida, A.; Aranda, A.; Prada, L.; Caldera, J.; Molina, J.I. Status and habitat changes in the endangered Spanish Imperial Eagle Aquila adalberti population during 1974–2004: Implications for its recovery. Bird Conserv. Int. 2008, 18, 242–259. [Google Scholar] [CrossRef] [Green Version]
- Naves, J.; Wiegand, T.; Fernández-Gil, A.; Stephan, T. Riesgo de Extinción del oso Pardo Cantábrico. La Población Occidental; Fundación Oso de Asturias: Oviedo, Spain, 1999. [Google Scholar]
- Carrete, M.; Grande, J.M.; Tella, J.L.; Sánchez-Zapata, J.A.; Donázar, J.A.; Díaz-Delgado, R.; Romo, A. Habitat, human pressure, and social behavior: Partialling out factors affecting large-scale territory extinction in an endangered vulture. Biol. Conserv. 2007, 136, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Sanz-Aguilar, A.; Sánchez-Zapata, J.A.; Carrete, M.; Benítez, J.R.; Ávila, E.; Arenas, R.; Donázar, J.A. Action on multiple fronts, illegal poisoning and wind farm planning, is required to reverse the decline of the Egyptian vulture in southern Spain. Biol. Conserv. 2015, 187, 10–18. [Google Scholar] [CrossRef] [Green Version]
- Berny, P.; Caloni, F.; Croubels, S.; Sachana, M.; Vandenbroucke, V.; Davanzo, F.; Guitart, R. Animal poisoning in Europe. Part 2: Companion animals. Vet. J. 2010, 183, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, D.P.; McLeod, D.R.A.; Watson, J.; Fielding, A.H.; Haworth, P.F. The association of grouse moor in Scotland with the illegal use of poisons to control predators. Biol. Conserv. 2003, 114, 157–163. [Google Scholar] [CrossRef]
- Cano, C. La Lucha Contra el Veneno en España (2011–2016). Clasificación por Comunidades Autónomas; WWF: Madrid, Spain, 2017. [Google Scholar]
- Moleón, M.; Martínez-Carrasco, C.; Muellerklein, O.C.; Getz, W.M.; Muñoz-Lozano, C.; Sánchez-Zapata, J.A. Carnivore carcasses are avoided by carnivores. J. Anim. Ecol. 2017, 86, 1179–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuur, A.; Leno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Blanco, J.C. Mamíferos de España. Vol. I and II; Editorial Planeta: Barcelona, Spain, 1998; ISBN 9788408028260. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. lme4: Linear Mixed-Effects Models Using Eigen and S4. R Package Version 1.0–5. 2013. Available online: https://CRAN.R-project.org/package=lme4 (accessed on 14 January 2021).
- Barton, K. MuMIn: Multi-Model Interference. R Package Version 1.9.13. 2013. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 14 January 2021).
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference. A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Richards, N. Carbofuran and Wildlife Poisoning: Global Perspectives and Forensic Approaches; John Wiley & Sons: Hoboken, NJ, USA, 2012; ISBN 1119951100. [Google Scholar]
- Barrientos, R.; Martins, R.C.; Ascensão, F.; D’Amico, M.; Moreira, F.; Borda-de-Água, L. A review of searcher efficiency and carcass persistence in infrastructure-driven mortality assessment studies. Biol. Conserv. 2018, 222, 146–153. [Google Scholar] [CrossRef]
- Ruxton, G.D.; Houston, D.C. Obligate vertebrate scavengers must be large soaring fliers. J. Theor. Biol. 2004, 228, 431–436. [Google Scholar] [CrossRef]
- Gutiérrez-Cánovas, C.; Moleón, M.; Mateo-Tomás, P.; Olea, P.P.; Sebastián-González, E.; Sánchez-Zapata, J.A. Large home range scavengers support higher rates of carcass removal. bioRxiv 2020, 1–32. [Google Scholar] [CrossRef]
- Brakes, C.R.; Smith, R.H. Exposure of non-target small mammals to rodenticides: Short-term effects, recovery and implications for secondary poisoning. J. Appl. Ecol. 2005, 42, 118–128. [Google Scholar] [CrossRef]
- Moleón, M.; Selva, N.; Quaggiotto, M.M.; Bailey, D.M.; Cortés-Avizanda, A.; DeVault, T.L. Carrion availability in space and time. In Carrion Ecology and Management; Olea, P.P., Mateo-Tomás, P., Sánchez-Zapata, J.A., Eds.; Springer: Cham, Switzerland, 2019; pp. 23–44. [Google Scholar]
- Moleón, M.; Sánchez-Zapata, J.A.; Sebastián-González, E.; Owen-Smith, N. Carcass size shapes the structure and functioning of an African scavenging assemblage. Oikos 2015, 124, 1391–1403. [Google Scholar] [CrossRef]
- Mateo-Tomás, P.; Olea, P.P.; Moleón, M.; Vicente, J.; Botella, F.; Selva, N.; Viñuela, J.; Sánchez-Zapata, J.A. From regional to global patterns in vertebrate scavenger communities subsidized by big game hunting. Divers. Distrib. 2015, 21, 913–924. [Google Scholar] [CrossRef]
- Sebastián-González, E.; Barbosa, J.M.; Pérez-García, J.M.; Morales-Reyes, Z.; Botella, F.; Olea, P.P.; Mateo-Tomás, P.; Moleón, M.; Hiraldo, F.; Arrondo, E.; et al. Scavenging in the Anthropocene: Human impact drives vertebrate scavenger species richness at a global scale. Glob. Chang. Biol. 2019, 25, 3005–3017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emery, N.J.; Clayton, N.S. The mentality of crows: Convergent evolution of intelligence in corvids and apes. Science 2004, 306, 1903–1907. [Google Scholar] [CrossRef] [PubMed]
- Moe, R.O.; Bakken, M.; Kittilsen, S.; Kingsley-Smith, H.; Spruijt, B.M. A note on reward-related behaviour and emotional expressions in farmed silver foxes (Vulpes vulpes)-Basis for a novel tool to study animal welfare. Appl. Anim. Behav. Sci. 2006, 101, 362–368. [Google Scholar] [CrossRef]
- Thomas, R.L.; Baker, P.J.; Fellowes, M.D.E. Ranging characteristics of the domestic cat (Felis catus) in an urban environment. Urban Ecosyst. 2014, 17, 911–921. [Google Scholar] [CrossRef]
- Jarjour, C.; Evans, J.C.; Routh, M.; Morand-Ferron, J. Does city life reduce neophobia? A study on wild black-capped chickadees. Behav. Ecol. 2020, 31, 123–131. [Google Scholar] [CrossRef]
- Bradshaw, J.W.S. The evolutionary basis for the feeding behavior of domestic dogs (Canis familiaris) and cats (Felis catus). J. Nutr. 2006, 136, 1927S–1931S. [Google Scholar] [CrossRef]
- Berny, P.; Vilagines, L.; Cugnasse, J.M.; Mastain, O.; Chollet, J.Y.; Joncour, G.; Razin, M. Vigilance Poison: Illegal poisoning and lead intoxication are the main factors affecting avian scavenger survival in the Pyrenees (France). Ecotoxicol. Environ. Saf. 2015, 118, 71–82. [Google Scholar] [CrossRef]
- Ogada, D.L.; Keesing, F.; Virani, M.Z. Dropping dead: Causes and consequences of vulture population declines worldwide. Ann. N. Y. Acad. Sci. 2012, 1249, 57–71. [Google Scholar] [CrossRef]
- Durá-Alemañ, C.J.; Ayerza, P.; Cano, C.; Jiménez, J.; Sánchez-Zapata, J.A.; Morales-Reyes, Z. Las sentencias contra el veneno en España. Quercus 2021, 419, 30–37. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Area | Habitat Type | Hunting Type/Intensity | Natural Park | Livestock/Density | Vulture Abundance | Wolf Abundance |
|---|---|---|---|---|---|---|
| Cabrera | Mosaic of transitional Mediterranean to Euro-Siberian scrubs, forests and meadows | Small and big game/low | No | Cattle, sheep/high | Low | High |
| Pre-Pyrenees | Mosaic of transitional Mediterranean to Euro-Siberian forests and meadows | Small and big game/low | No | Cattle, sheep/high | High | (Absent) |
| Cameros | Mosaic of transitional Mediterranean to Euro-Siberian forests and meadows | Small and big game/medium | No | Cattle, horses, sheep/high | High | Low |
| Harana | Mosaic of xeric Mediterranean scrubs, forests and crops | Small and big game/high | No | Sheep, goats/low | Low | (Absent) |
| Cazorla | Mediterranean forests | Big game/medium | Yes | Sheep, goats/medium | High | (Absent) |
| Escalona | Mosaic of xeric Mediterranean scrubs, forests and crops | Small game/high | No | Sheep/very low | (Absent) | (Absent) |
| Source | Wild Carnivores | Domestic Carnivores | Suids | Small Mammals | Corvids | Vultures | Other Raptors | Other Birds | Reptiles | N |
|---|---|---|---|---|---|---|---|---|---|---|
| Database | 6 | 2 | 1 | 2 | 4 | 3 | 9 | 1 | 1 | 27 |
| Experiment | 5 | 2 | 1 | 3 | 5 | 3 | 3 | 4 | 2 | 26 |
| Total | 7 | 1 | 5 | 5 | 4 | 9 | 5 | 2 | 38 | |
| Spain | 16 (44%) | 1 (100%) | 34 (15%) | 9 (56%) | 4 (100%) | 31 (29%) | 528 (1%) | 57 (4%) | 680 (6%) |
| Parameter | Estimate | SE |
|---|---|---|
| (Intercept) | 1.154 | 1.325 |
| weight | 3.612 | 0.950 |
| color (conspicuous) | −5.139 | 1.960 |
| mobility (aerial) | 4.289 | 1.863 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gil-Sánchez, J.M.; Aguilera-Alcalá, N.; Moleón, M.; Sebastián-González, E.; Margalida, A.; Morales-Reyes, Z.; Durá-Alemañ, C.J.; Oliva-Vidal, P.; Pérez-García, J.M.; Sánchez-Zapata, J.A. Biases in the Detection of Intentionally Poisoned Animals: Public Health and Conservation Implications from a Field Experiment. Int. J. Environ. Res. Public Health 2021, 18, 1201. https://doi.org/10.3390/ijerph18031201
Gil-Sánchez JM, Aguilera-Alcalá N, Moleón M, Sebastián-González E, Margalida A, Morales-Reyes Z, Durá-Alemañ CJ, Oliva-Vidal P, Pérez-García JM, Sánchez-Zapata JA. Biases in the Detection of Intentionally Poisoned Animals: Public Health and Conservation Implications from a Field Experiment. International Journal of Environmental Research and Public Health. 2021; 18(3):1201. https://doi.org/10.3390/ijerph18031201
Chicago/Turabian StyleGil-Sánchez, José M., Natividad Aguilera-Alcalá, Marcos Moleón, Esther Sebastián-González, Antoni Margalida, Zebensui Morales-Reyes, Carlos J. Durá-Alemañ, Pilar Oliva-Vidal, Juan M. Pérez-García, and José A. Sánchez-Zapata. 2021. "Biases in the Detection of Intentionally Poisoned Animals: Public Health and Conservation Implications from a Field Experiment" International Journal of Environmental Research and Public Health 18, no. 3: 1201. https://doi.org/10.3390/ijerph18031201