
Enhancement of X-ray Induced Apoptosis by Mobile Phone-Like Radio-Frequency Electromagnetic Fields in Mouse Spermatocyte-Derived Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. RF Field and X-ray Exposure
2.3. MTT Assay
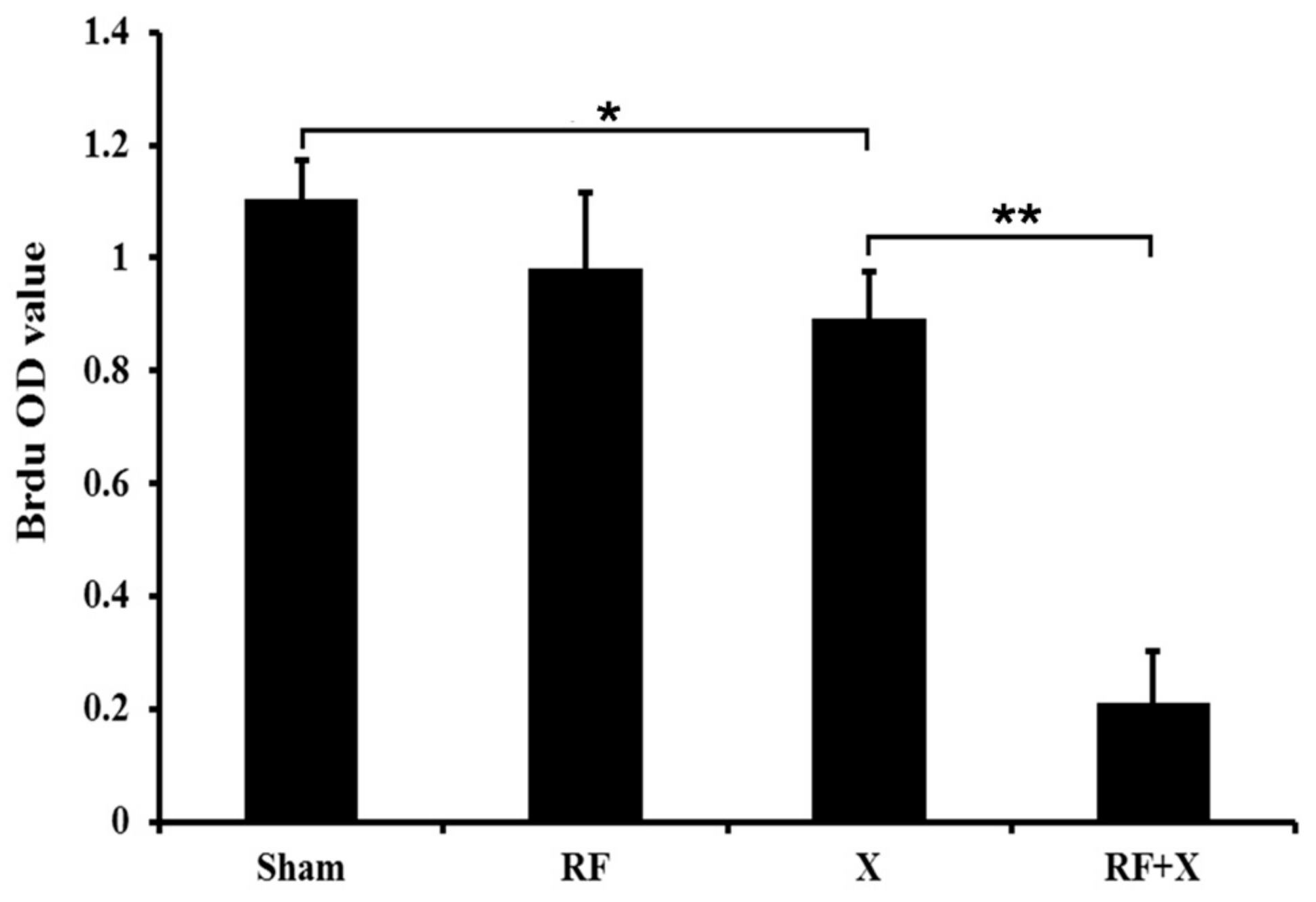
2.4. BrdU ELISA Assay
2.5. Flow Cytometry Analysis for Apoptosis
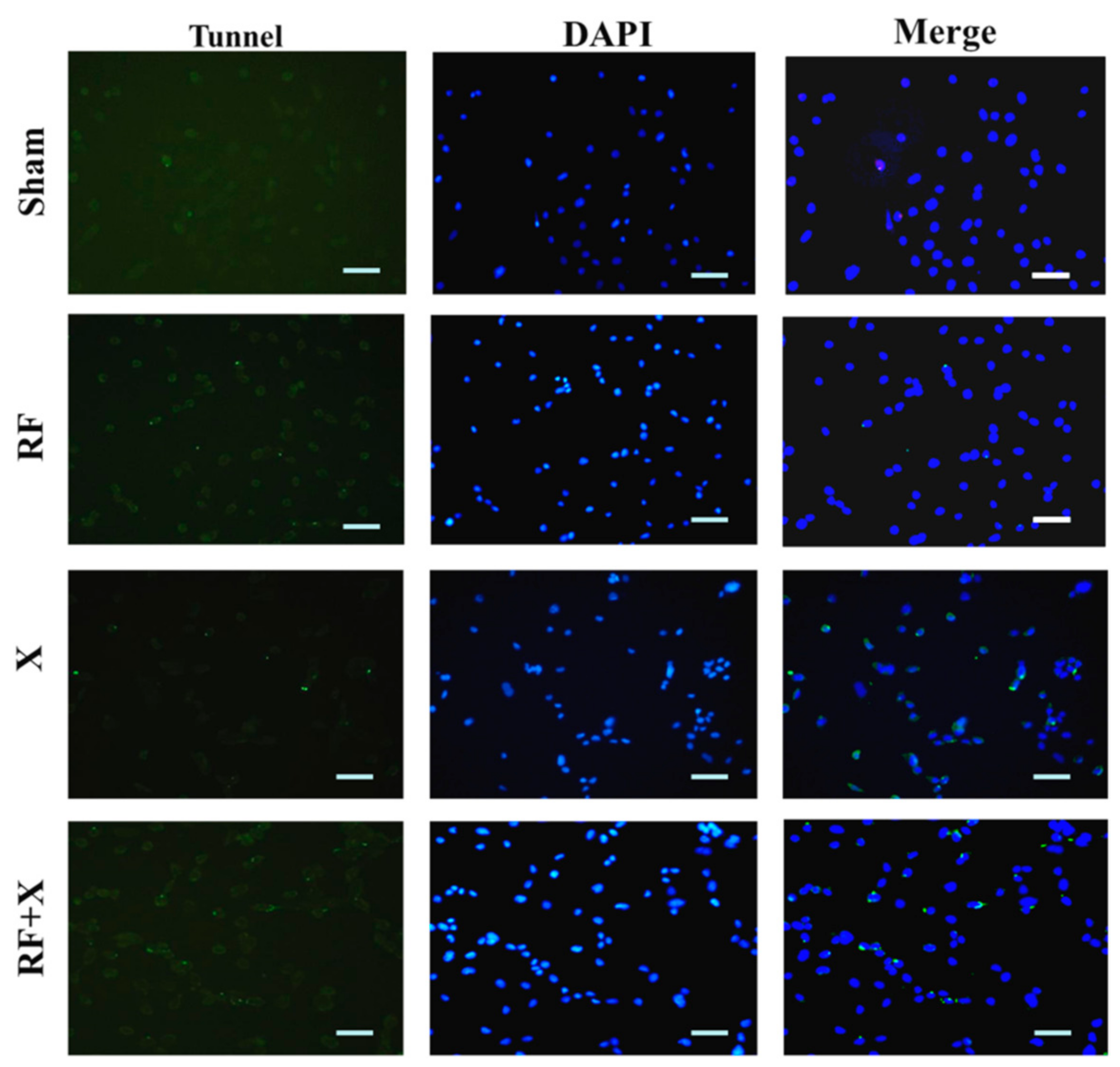
2.6. TUNEL Assay
2.7. Caspase-3 Activity Assay
2.8. Western Blot Analysis and Antibodies
2.9. Statistical Analysis
3. Results
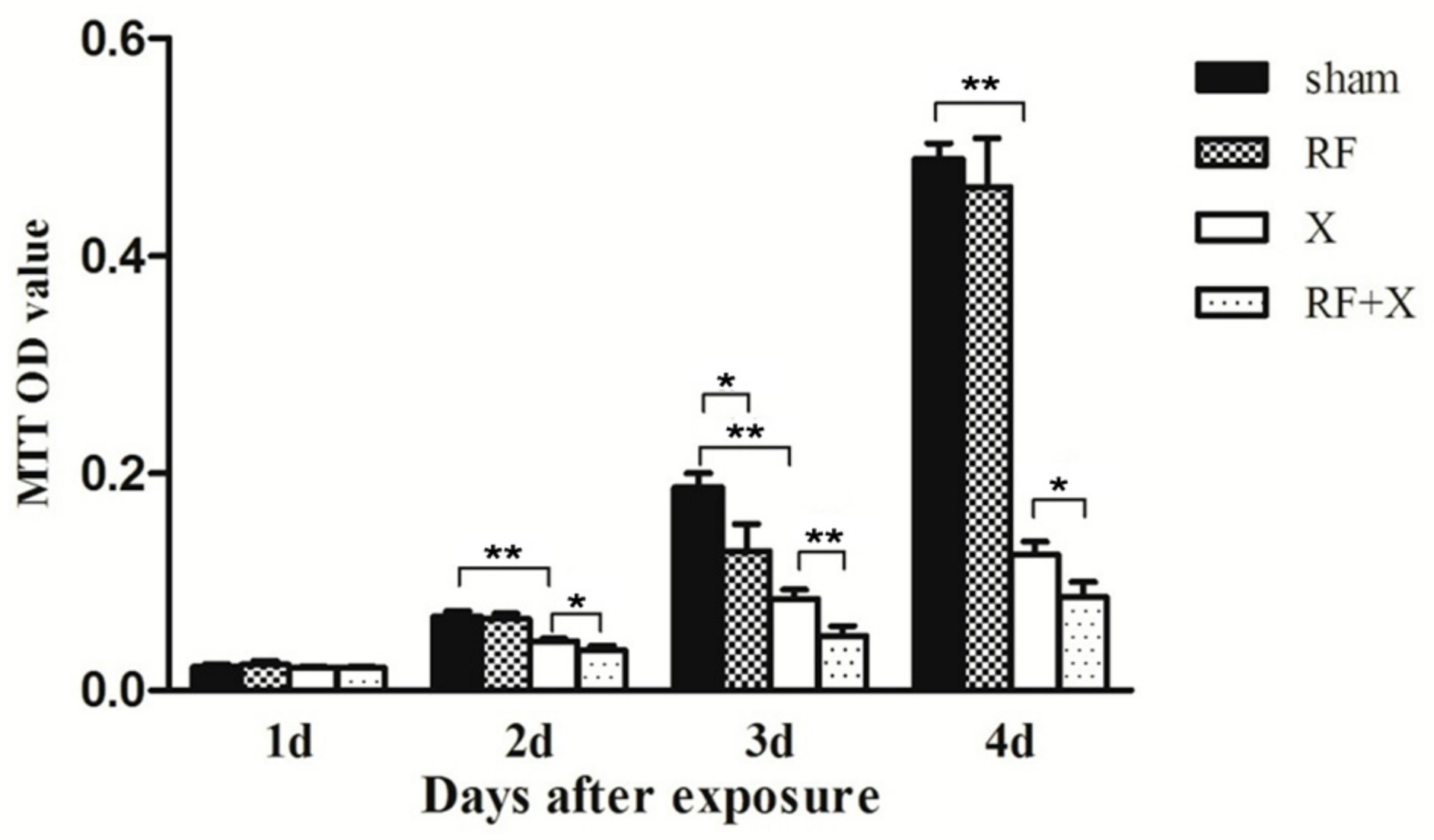
3.1. Cell Proliferation of GC-1 Cells after Exposure to RF and/or X-ray Determined by MTT Assay
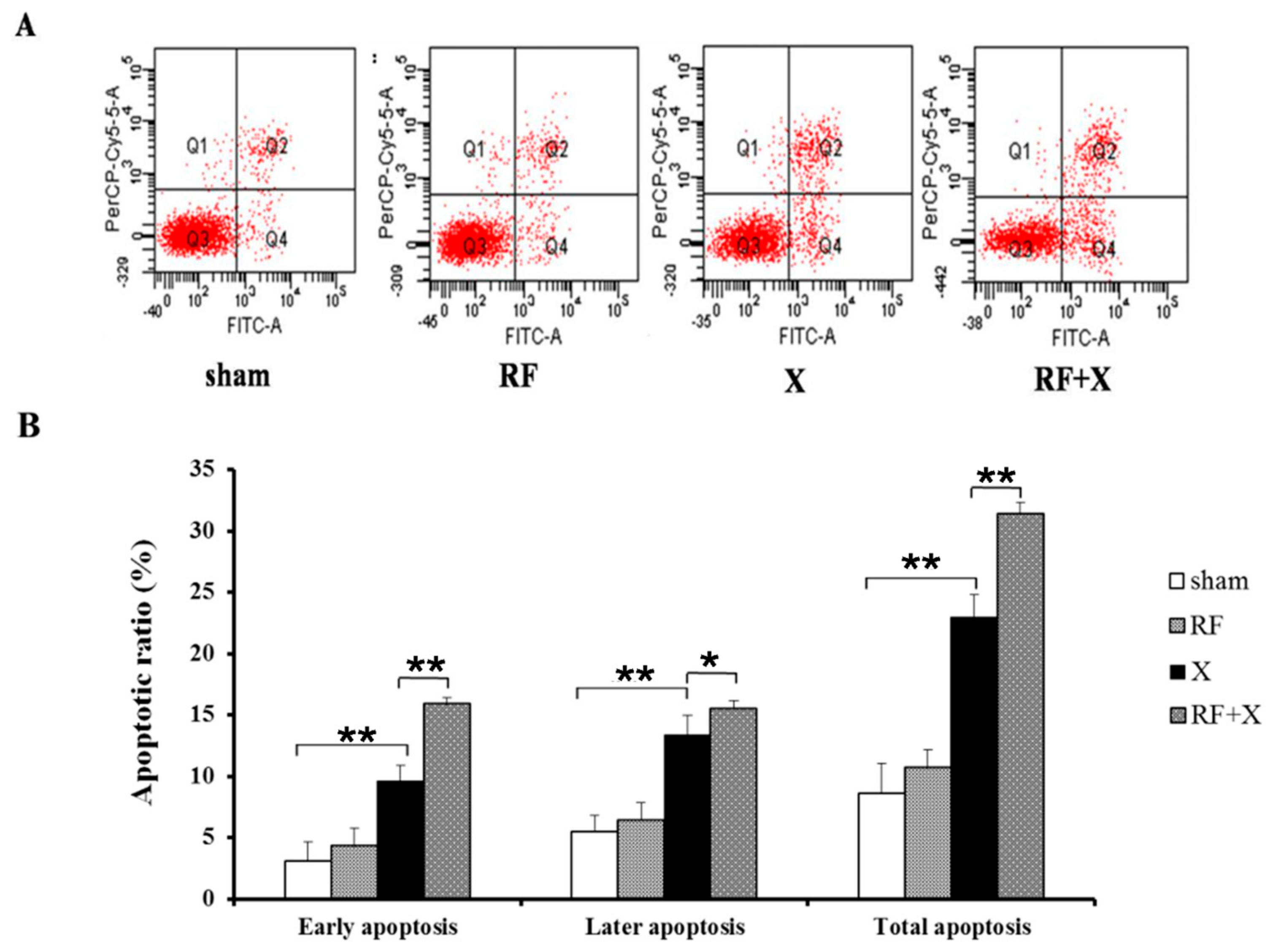
3.2. The Apoptosis Level of GC-1 Cells after Exposure to RF and/or X-ray
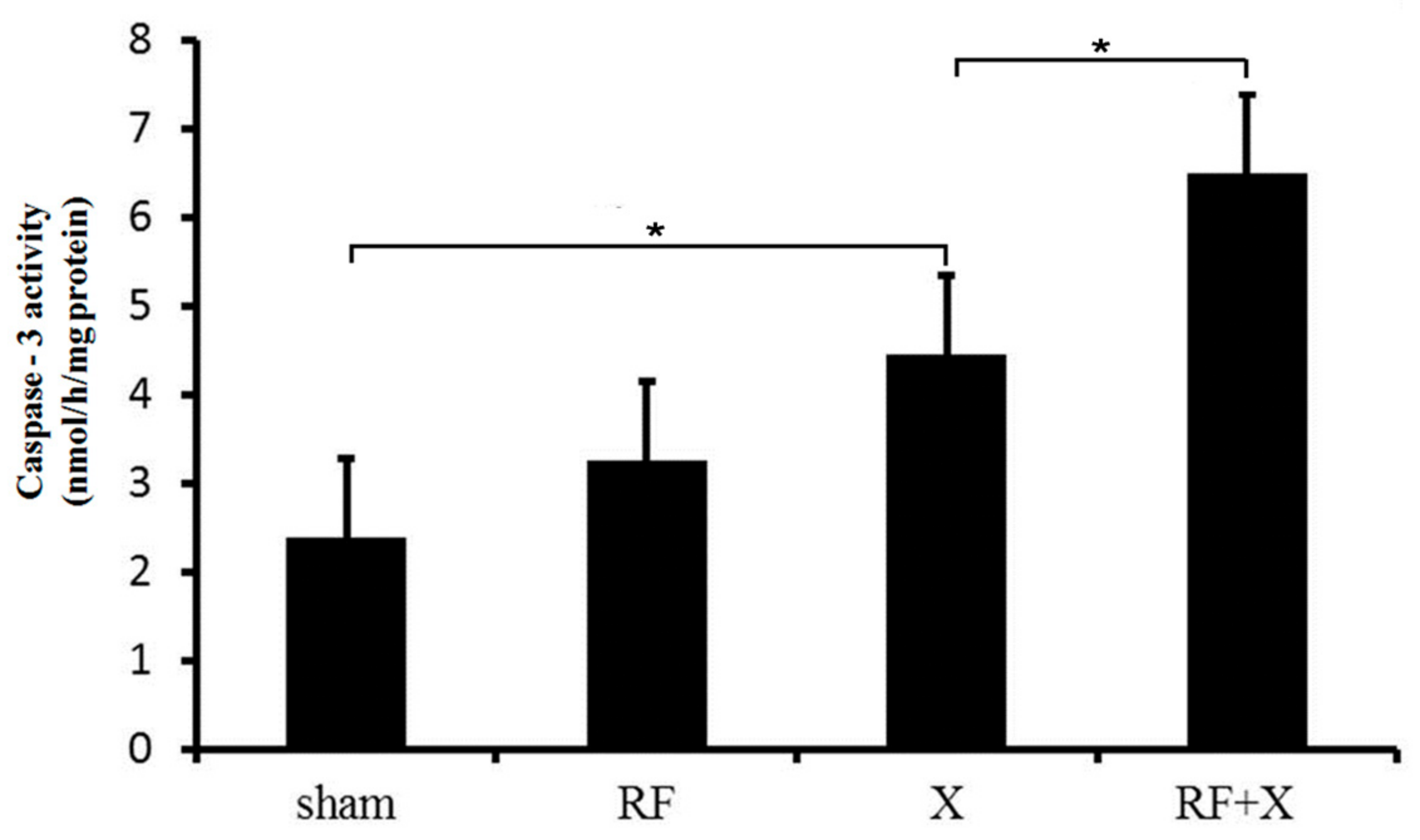
3.3. The Activity of Caspase-3 in GC-1 Cells after Treatment with RF and/or X-ray
3.4. Protein Expression after Treatment with RF and/or X-ray
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wdowiak, A.; Wdowiak, L.; Wiktor, H. Evaluation of the effect of using mobile phones on male fertility. Ann. Agric. Environ. Med. 2007, 14, 169–172. [Google Scholar] [PubMed]
- La-Vignera, S.; Condorelli, R.A.; Vicari, E.; D’Agata, R.; Calogero, A.E. Effects of the exposure to mobile phones on male reproduction: A review of the literature. J. Androl. 2012, 33, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Deepinder, F.; Sharma, R.K.; Ranga, G.; Li, J. Effect of cell phone usage on semen analysis in men attending infertility clinic: An observational study. Fertil. Steril. 2008, 89, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Gutschi, T.; Mohamad, A.-A.B.; Shamloul, R.; Pummer, K.; Trummer, H. Impact of cell phone use on men’s semen parameters. Andrologia 2011, 43, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.C.; Minguez-Alarcon, L.; Meeker, J.D.; Williams, P.L.; Mezei, G.; Ford, J.B.; Hauser, R.; EARTH Study Team. Self-reported mobile phone use and semen parameters among men from a fertility clinic. Reprod. Toxicol. 2016, 67, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Erogul, O.; Oztas, E.; Yildirim, I.; Kir, T.; Aydur, E.; Komesli, G.; Irkilata, H.C.; Irmak, M.K.; Peker, A.F. Effects of electromagnetic radiation from a cellular phone on human sperm motility: An in vitro study. Arch. Med. Res. 2006, 37, 840–843. [Google Scholar] [CrossRef] [PubMed]
- Dasdag, S.; Tas, M.; Akdag, M.Z.; Yegin, K. Effect of long-term exposure of 2.4 GHz radiofrequency radiation emitted from Wi-Fi equipment on testes functions. Electromagn. Biol. Med. 2015, 34, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Tas, M.; Dasdag, S.; Akdag, M.Z.; Cirit, U.; Yegin, K.; Seker, U.; Ozmen, M.F.; Eren, L.B. Long-term effects of 900 MHz radiofrequency radiation emitted from mobile phone on testicular tissue and epididymal semen quality. Electromagn. Biol. Med. 2014, 33, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Kesari, K.K.; Behari, J. Influence of microwave exposure on fertility of male rats. Fertil. Steril. 2011, 95, 1500–1502. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Duan, W.; Xu, S.; Chen, C.; He, M.; Zhang, L.; Yu, Z.; Zhou, Z. Exposure to 1800 MHz radiofrequency electromagnetic radiation induces oxidative DNA base damage in a mouse spermatocyte-derived cell line. Toxicol. Lett. 2013, 218, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Jin, Y.B.; Kim, T.H.; Pack, J.K.; Kim, N.; Choi, H.D.; Lee, J.S.; Lee, Y.S. The effects of simultaneous combined exposure to CDMA and WCDMA electromagnetic fields on rat testicular function. Bioelectromagnetics 2012, 33, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.B.; Choi, H.D.; Kim, B.C.; Pack, J.K.; Kim, N.; Lee, Y.S. Effects of simultaneous combined exposure to CDMA and WCDMA electromagnetic fields on serum hormone levels in rats. J. Radiat. Res. 2013, 54, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Dasdag, S.; Akdag, M.Z.; Ulukaya, E.; Uzunlar, A.K.; Yegin, D. Mobile phone exposure does not induce apoptosis on spermatogenesis in rats. Arch. Med. Res. 2008, 39, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Bonde, J.P. Male reproductive organs are at risk from environmental hazards. Asian J. Androl. 2010, 12, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Aghamohammadi, A.; Hosseinimehr, S.J.; Ghasemi, A.; Azadbakht, M.; Pourfallah, T.A. Radiosensitization Effects of a Zataria multiflora Extract on Human Glioblastoma Cells. Asian Pac. J. Cancer Prev. 2015, 16, 7285–7290. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.J.; Li, J.H.; Zhang, Y.J.; Zhao, S.T. Optimal combination of radiotherapy and endocrine drugs in breast cancer treatment. Cancer Radiother. 2013, 17, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.; Wilson, G.; Russell, L.D.; Meistrich, M.L. Radiation-induced cell death in the mouse testis: Relationship to apoptosis. Radiat. Res. 1997, 147, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Saygin, M.; Asci, H.; Ozmen, O.; Cankara, F.N.; Dincoglu, D.; Ilhan, I. Impact of 2.45 GHz microwave radiation on the testicular inflammatory pathway biomarkers in young rats: The role of gallic acid. Environ. Toxicol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, S.; Cabot, E.; Christ, A.; Capstick, M.; Kuster, N. Assessment of the radio-frequency electromagnetic fields induced in the human body from mobile phones used with hands-free kits. Phys. Med. Biol. 2009, 54, 5493–5508. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Tang, X.L.; Miao, S.Y.; Zong, S.D.; Wang, L.F. Regulation of the G2/M phase of the cell cycle by sperm associated antigen 8 (SPAG8) protein. Cell Biochem. Funct. 2009, 27, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Kanter, M.; Topcu-Tarladacalisir, Y.; Parlar, S. Antiapoptotic effect of L-carnitine on testicular irradiation in rats. J. Mol. Histol. 2010, 41, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Burrer, C.M.; Foight, G.W.; Keating, A.E.; Chan, G.C. Selective peptide inhibitors of antiapoptotic cellular and viral Bcl-2 proteins lead to cytochrome C release during latent Kaposi’s sarcoma-associated herpesvirus infection. Virus Res. 2016, 211, 86–88. [Google Scholar] [CrossRef] [PubMed]
- Ryu, B.; Ahn, B.N.; Kang, K.H.; Kim, Y.S.; Li, Y.X.; Kong, C.S.; Kim, S.K.; Kim, D.G. Dioxinodehydroeckol protects human keratinocyte cells from UVB-induced apoptosis modulated by related genes Bax/Bcl-2 and caspase pathway. J. Photochem. Photobiol. B 2015, 153, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Shalini, S.; Dorstyn, L.; Dawar, S.; Kumar, S. Old, new and emerging functions of caspases. Cell Death Differ. 2015, 22, 526–539. [Google Scholar] [CrossRef] [PubMed]
- Thomas, L.W.; Lam, C.; Edwards, S.W. Mcl-1; the molecular regulation of protein function. FEBS Lett. 2010, 584, 2981–2989. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.; Watari, H.; AbuAlmaaty, A.; Ohba, Y.; Sakuragi, N. Apoptosis and molecular targeting therapy in cancer. Biomed. Res. Int. 2014, 2014, 150845. [Google Scholar] [CrossRef] [PubMed]
- Sannino, A.; Zeni, O.; Romeo, S.; Massa, R.; Gialanella, G.; Grossi, G.; Manti, L.; Vijayalaxmi; Scarfi, M.R. Adaptive response in human blood lymphocytes exposed to non-ionizing radiofrequency fields: Resistance to ionizing radiation-induced damage. J. Radiat. Res. 2014, 55, 210–217. [Google Scholar] [CrossRef]
- Yongxin, J.; Qina, H.; Yulong, S.; Jian, T.; Yi, C. Adaptive response in mouse bone-marrow stromal cells exposed to 900-MHz radiofrequency fields: Gamma-radiation-induced DNA strand breaks and Repair. J. Toxicol. Environ. Health A 2016, 79, 419–426. [Google Scholar] [CrossRef]
- International Commission on Non-Ionizing Radiation Protection. Guidelines for limiting exposure to time-varying electric magnetic and electromagnetic fields (up to 300 GHz). Health Phys. 1998, 74, 494–522. [Google Scholar]
- Institute of Electrical and Electronics Engineers. IEEE Standard for Safety Levels with Respect to Human Exposure to Radio Frequency Electromagnetic Fields, 3 kHz to 300 GHz; IEEE Std C95.1™-2005; IEEE: New York, NY, USA, 2006. [Google Scholar]






© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, K.-Y.; Xu, H.; Du, L.; Xing, J.-L.; Zhang, B.; Bai, Q.-S.; Xu, Y.-Q.; Zhou, Y.-C.; Zhang, J.-P.; Zhou, Y.; et al. Enhancement of X-ray Induced Apoptosis by Mobile Phone-Like Radio-Frequency Electromagnetic Fields in Mouse Spermatocyte-Derived Cells. Int. J. Environ. Res. Public Health 2017, 14, 616. https://doi.org/10.3390/ijerph14060616
Zhang K-Y, Xu H, Du L, Xing J-L, Zhang B, Bai Q-S, Xu Y-Q, Zhou Y-C, Zhang J-P, Zhou Y, et al. Enhancement of X-ray Induced Apoptosis by Mobile Phone-Like Radio-Frequency Electromagnetic Fields in Mouse Spermatocyte-Derived Cells. International Journal of Environmental Research and Public Health. 2017; 14(6):616. https://doi.org/10.3390/ijerph14060616
Chicago/Turabian StyleZhang, Ke-Ying, Hui Xu, Le Du, Jun-Ling Xing, Bin Zhang, Qiang-Shan Bai, Yu-Qiao Xu, Yong-Chun Zhou, Jun-Ping Zhang, Yan Zhou, and et al. 2017. "Enhancement of X-ray Induced Apoptosis by Mobile Phone-Like Radio-Frequency Electromagnetic Fields in Mouse Spermatocyte-Derived Cells" International Journal of Environmental Research and Public Health 14, no. 6: 616. https://doi.org/10.3390/ijerph14060616