
Commercial Honeybush (Cyclopia spp.) Tea Extract Inhibits Osteoclast Formation and Bone Resorption in RAW264.7 Murine Macrophages—An in vitro Study

 and
and
Abstract
:1. Introduction
2. Experimental Section
2.1. Reagents and Materials
2.2. Tea Extract Preparation
2.3. Analysis of Known Flavonoids by HPLC-DAD
2.4. Cell Culture and Maintenance
2.5. Alamar Blue Assay
2.6. Investigation of Osteoclast Formation and Activity
2.6.1. Cell Morphology: PlasDIC
2.6.2. TRAP Staining for Multinucleated Osteoclasts
2.6.3. TRAP Activity in Conditioned Media
2.6.4. Bone Resorption Assay
2.6.5. Visualisation of Multinucleated Cells with Actin Rings
2.6.6. Gene Expression Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5’ – 3’) | Genbank Accession Number |
|---|---|---|
| MMP9 | GTCATCCAGTTTGGTGTCGCG AGGGGAAGACGCACAGCTC | NM_013599.3 |
| TRAP | CCACCCTGAGATTTGTGGCT ACATACCAGGGGATGTTGCG | NM_008084.2 |
| Cathepsin K | CTGGAGGGCCAACTCAAGA CCTCTGCATTTAGCTGCCTT | NM_007802.4 |
| GAPDH | GATGACATCAAGAAGGTGGTGAAGC ATACCAGGAAATGAGCTTGACAAAG | NM_001102404.1 |
2.7. Statistical Analysis
3. Results
3.1. HPLC-DAD
| Compound | Iriflophenone -3-C-Glucoside | Mangiferin | Isomangiferin | Hesperidin | Unknown 13 |
|---|---|---|---|---|---|
| Concentration | 9.665 | 10.009 | 9.952 | 6.426 | 6.092 |
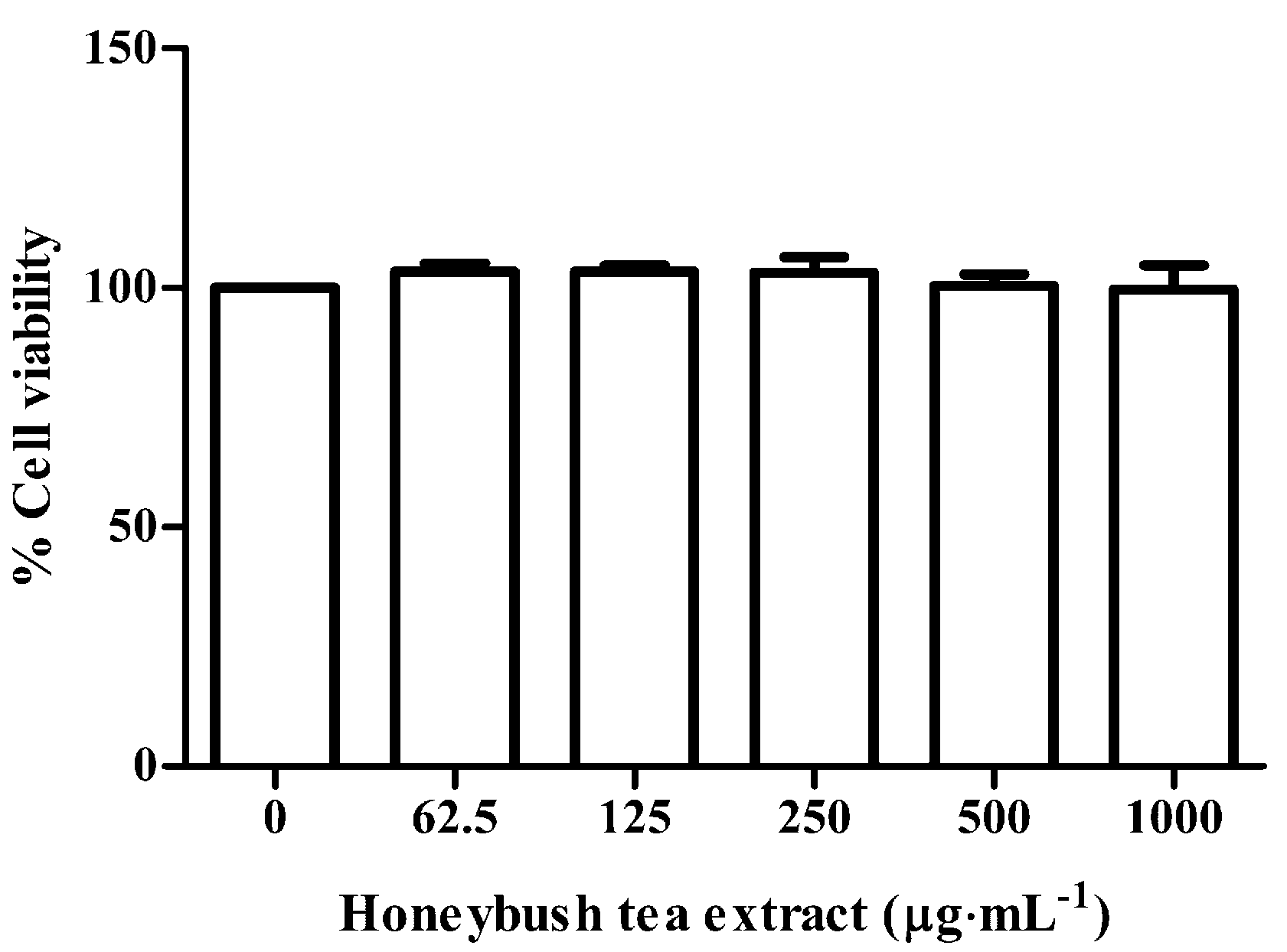
3.2. Effect of Honeybush Tea Extract on Cell Viability

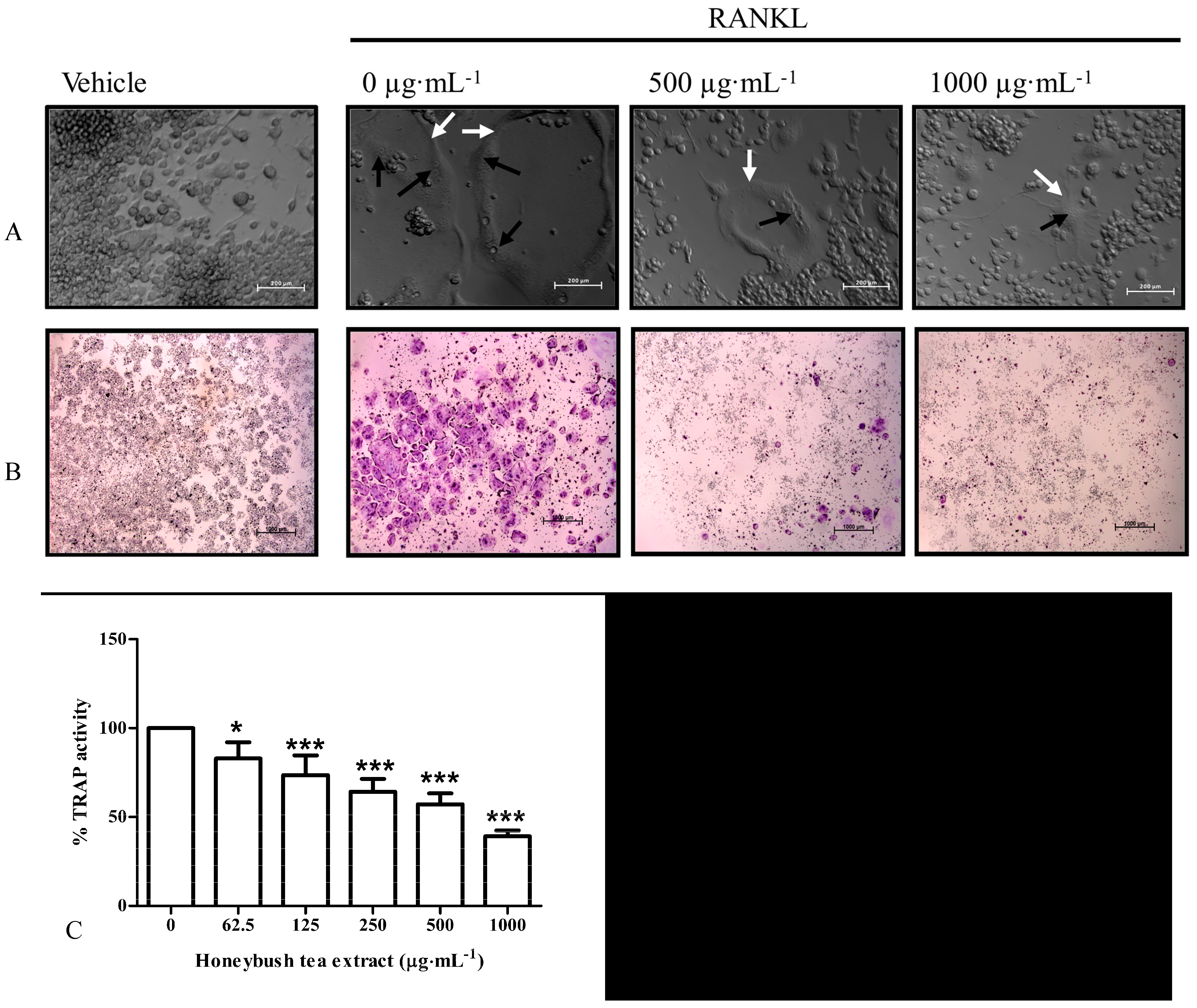
3.3. Effect of Honeybush Tea Extract on Cell Morphology

3.4. Effect of Honeybush Tea Extract on RANKL-Induced Osteoclast Formation and TRAP Activity in RAW264.7 Macrophages
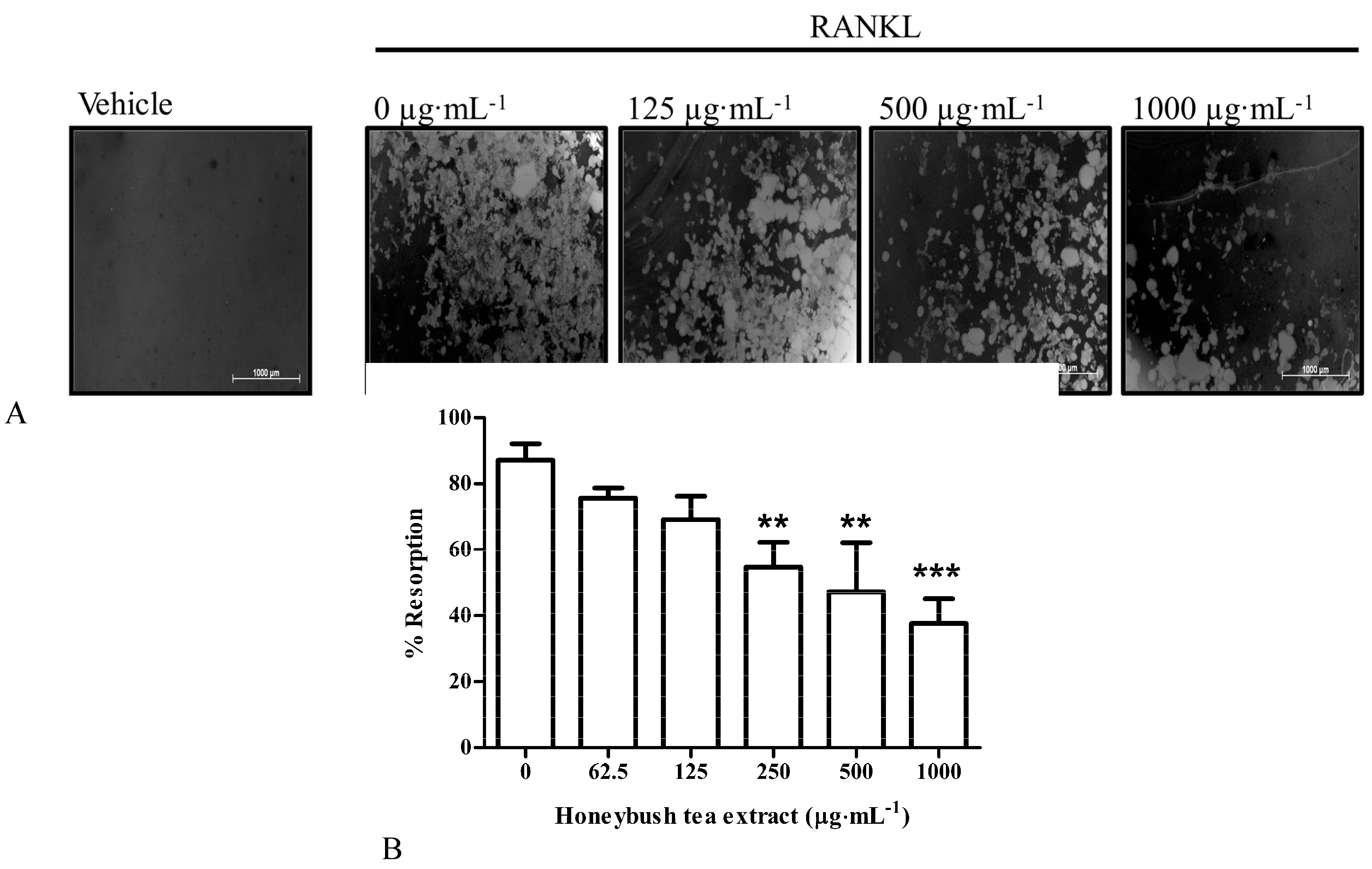
3.5. Effect of Honeybush Tea Extract on Bone Resorption in RAW264.7 Macrophages
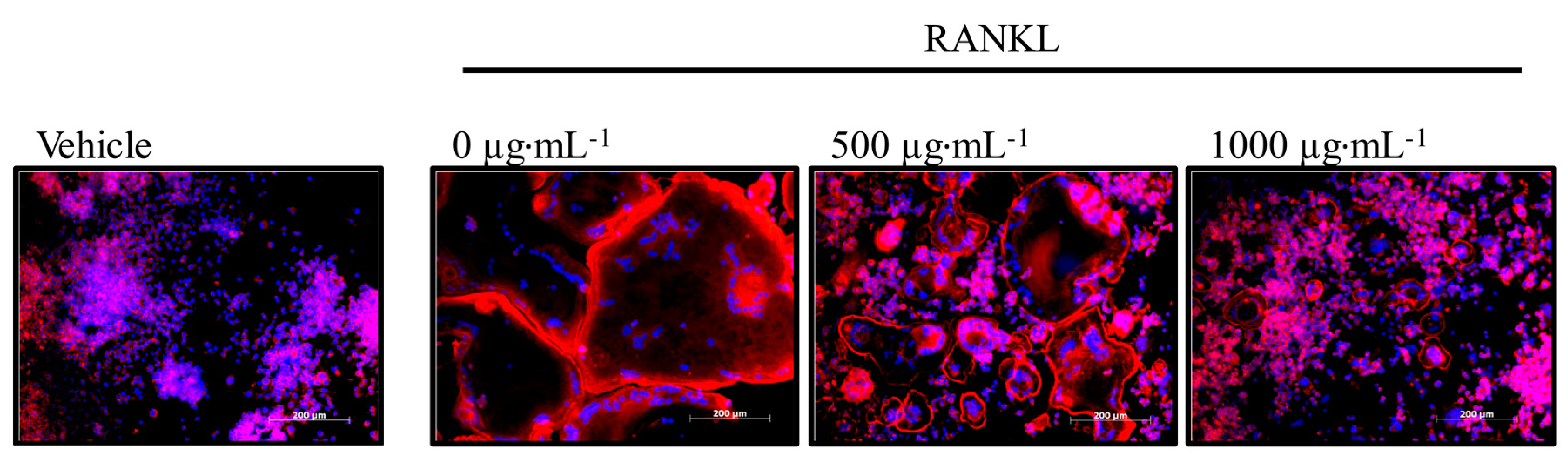
3.6. Effect of Honeybush Tea Extract on Actin Ring Formation in RAW264.7 Macrophages
3.7. Effect of Honeybush Tea Extract on Osteoclast Specific Gene Expression in RAW264.7 Macrophages



4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Agrawal, V.; Gupta, D. Recent update on osteoporosis. Int. J. Med. Sci. 2013, 2, 164–168. [Google Scholar]
- Vӓӓnӓnen, H.K.; Laitala-Leinonen, T. Osteoclast lineage and function. Arch. Biochem. Biophys. 2008, 473, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Vӓӓnӓnen, H.K.; Zhao, H.; Mulari, M.; Halleen, J.M. The cell biology of osteoclast function. J. Cell Sci. 2000, 113, 377–381. [Google Scholar] [PubMed]
- Halleen, J.M.; Tiitinen, S.L.; Ylipahkala, H.; Fagerlund, K.M.; Vӓӓnӓnen, H.K. Tartrate-resistant acid phosphatase 5b (TRACP 5b) as a marker of bone resorption. Clin. Lab. 2006, 52, 499–509. [Google Scholar] [PubMed]
- Kikuta, J.; Ishii, M. Osteoclast migration, differentiation and function: Novel therapeutic targets for rheumatic diseases. Rheumatology 2013, 52, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Goltzman, D. Discoveries, drugs and skeletal disorders. Nat. Rev. Drug Discov. 2002, 1, 784–796. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Udagawa, N.; Suda, T. Vitamin D endocrine system and osteoclasts. Bonekey Rep. 2014. [Google Scholar] [CrossRef]
- Lee, N.K. Molecular understanding of osteoclast differentiation and physiology. Endocr. Metab. 2010, 25, 264–269. [Google Scholar] [CrossRef]
- Joubert, E.; Joubert, M.E.; Bester, C.; de Beer, D.; de Lange, J.H. Honeybush (Cyclopia spp.): From local cottage industry to global markets—The catalytic and supporting role of research. S. Afr. J. Bot. 2011, 77, 887–907. [Google Scholar] [CrossRef]
- McKay, D.L.; Blumberg, J.B. A review of the bioactivity of South African herbal teas: Rooibos (Aspalathus linearis) and honeybush (Cyclopia intermedia). Phytother. Res. 2007, 21, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kokotkiewicz, A.; Luczkiewicz, M. Honeybush (Cyclopia spp.)—A rich source of compounds with high antimutagenic properties. Fitoterapia 2008, 80, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Marnewick, J.L.; Gelderblom, W.C.; Joubert, E. An investigation on the antimutagenic properties of South African herbal teas. Mutat. Res. 2000, 471, 157–166. [Google Scholar] [CrossRef]
- Marnewick, J.; Joubert, E.; Joseph, S.; Swanevelder, S.; Swart, P.; Gelderblom, W. Inhibition of tumour promotion in mouse skin by extracts of Rooibos (Aspalathus linearis) and Honeybush (Cyclopia intermedia), unique South African herbal teas. Cancer Lett. 2005, 224, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.L.; Yeh, J.K.; Samathanam, C.; Cao, J.J.; Stoecker, B.J.; Dagda, R.Y.; Chyu, M.C.; Wang, J.S. Protective actions of green tea polyphenols and alfacalcidol on bone microstructure in female rats with chronic inflammation. J. Nutr. Biochem. 2011, 22, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.L.; Wang, P.; Guerrieri, J.; Yeh, J.K.; Wang, J.S. Protective effect of green tea polyphenols on bone loss in middle-aged female rats. Osteoporos. Int. 2008, 19, 979–990. [Google Scholar] [CrossRef] [PubMed]
- Hendricks, R.; Pool, E.J. The in vitro effects of Rooibos and black tea on immune pathways. J. Immunoass. Immunochem. 2010, 31, 169–180. [Google Scholar] [CrossRef] [PubMed]
- De Beer, D.; Joubert, E. Development of HPLC method for Cyclopia subternata phenolic compound analysis and application to other Cyclopia spp. J. Food Compos. Anal. 2010, 23, 289–297. [Google Scholar] [CrossRef]
- Deepak, V.; Kasonga, A.; Kruger, M.C.; Coetzee, M. Inhibitory effects of eugenol on RANKL-induced osteoclast formation via attenuation of NF-kappaB and MAPK pathways. Connect. Tissue Res. 2015, 56, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Kasonga, A.E.; Deepak, V.; Kruger, M.C.; Coetzee, M. Arachidonic acid and docosahexaenoic acid suppress osteoclast formation and activity in human CD14+ monocytes, in vitro. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Bar-Shavit, Z. The osteoclast: A multinucleated, hematopoietic-origin, bone-resorbing osteoimmune cell. J. Cell Biochem. 2007, 102, 1130–1139. [Google Scholar] [CrossRef] [PubMed]
- Nash, L.A.; Ward, W.E. Tea and bone health: Findings from human studies, potential mechanisms, and identification of knowledge gaps. Crit. Rev. Food Sci. Nutr. 2015. [Google Scholar] [CrossRef] [PubMed]
- Devine, A.; Hodgson, J.M.; Dick, I.M.; Prince, R.L. Tea drinking is associated with benefits on bone density in older women. Am. J. Clin. Nutr. 2007, 86, 1243–1247. [Google Scholar] [PubMed]
- Wu, C.H.; Yang, Y.C.; Yao, W.J.; Lu, F.H.; Wu, J.S.; Chang, C.J. Epidemiological evidence of increased bone mineral density in habitual tea drinkers. Arch. Intern. Med. 2002, 162, 1001–1006. [Google Scholar] [CrossRef] [PubMed]
- Hossein-Nezhad, A.; Maghbooli, Z.; Shafaei, A.R.; Javadi, E.; Larijani, B. Relationship between tea drinking and bone mineral density in Iranian population. Iran. J. Public Health 2007, 13, 57–62. [Google Scholar]
- Hsiao, M.C.; Liu, C.Y.; Wang, C.J. Factors associated with low bone density among women with major depressive disorder. Int. J. Psychiat. Med. 2012, 44, 77–90. [Google Scholar] [CrossRef]
- Leibbrandt, A.; Penninger, J.M. RANK(L) as a key target for controlling bone loss. Adv. Exp. Med. Biol. 2009, 647, 130–145. [Google Scholar] [PubMed]
- Collin-Osdoby, P.; Osdoby, P. RANKL-mediated osteoclast formation from murine RAW 264.7 cells. Methods Mol. Biol. 2012, 816, 187–202. [Google Scholar] [PubMed]
- Kokotkiewicz, A.; Luczkiewicz, M.; Sowinski, P.; Glod, D.; Gorynski, K.; Bucinski, A. Isolation and structure elucidation of phenolic compounds from Cyclopia subternata Vogel (Honeybush) intact plant and in vitro cultures. Food Chem. 2012, 133, 1373–1382. [Google Scholar] [CrossRef]
- Schulze, A.E.; Beelders, T.; Koch, I.S.; Erasmus, L.M.; de Beer, D.; Joubert, E. Honeybush herbal teas (Cyclopia spp.) contribute to high levels of dietary exposure to xanthones, benzophenones, dihydrochalcones and other bioactive phenolics. J. Food Compos. Anal. 2015, 44, 139–148. [Google Scholar] [CrossRef]
- Chiba, H.; Uehara, M.; Wu, J.; Wang, X.; Masuyama, R.; Suzuki, K.; Kanazawa, K.; Ishimi, Y. Hesperidin, a citrus flavonoid, inhibits bone loss and decreases serum and hepatic lipids in ovariectomized mice. J. Nutr. 2003, 133, 1892–1897. [Google Scholar] [PubMed]
- Chiba, H.; Kim, H.; Matsumoto, A.; Akiyama, S.; Ishimi, Y.; Suzuki, K.; Uehara, M. Hesperidin prevents androgen deficiency-induced bone loss in male mice. Phytother. Res. 2014, 28, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Miyahara, T.; Tezuka, Y.; Namba, T.; Nemoto, N.; Tonami, S.; Seto, H.; Tada, T.; Kadota, S. The effect of Kampo formulae on bone resorption in vitro and in vivo. I. Active constituents of Tsu-kan-gan. Biol. Pharm. Bull. 1998, 21, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Ang, E.; Liu, Q.; Qi, M.; Liu, H.G.; Yang, X.; Chen, H.; Zheng, M.H.; Xu, J. Mangiferin attenuates osteoclastogenesis, bone resorption, and RANKL-induced activation of NF-kappaB and ERK. J. Cell Biochem. 2011, 112, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Teitelbaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef] [PubMed]
- Dudhia, Z.; Louw, J.; Muller, C.; Joubert, E.; de Beer, D.; Kinnear, C.; Pheiffer, C. Cyclopia maculata and Cyclopia subternata (honeybush tea) inhibits adipogenesis in 3T3-L1 pre-adipocytes. Phytomedicine 2013, 20, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Yamaza, T.; Tanaka, T. Cathepsins in the osteoclast. J. Electron. Microsc. 2003, 52, 551–558. [Google Scholar] [CrossRef]
- Oddie, G.W.; Schenk, G.; Angel, N.Z.; Walsh, N.; Guddat, L.W.; de Jersey, J.; Cassady, A.I.; Hamilton, S.E.; Hume, D.A. Structure, function, and regulation of tartrate-resistant acid phosphatase. Bone 2000, 27, 575–584. [Google Scholar] [CrossRef]
- Saftig, P.; Hunziker, E.; Everts, V.; Jones, S.; Boyde, A.; Wehmeyer, O.; Suter, A.; von Figura, K. Functions of cathepsin K in bone resorption. Lessons from cathepsin K deficient mice. Adv. Exp. Med. Biol. 2000, 477, 293–303. [Google Scholar] [PubMed]
- Hollberg, K.; Hultenby, K.; Hayman, A.; Cox, T.; Andersson, G. Osteoclasts from mice deficient in tartrate-resistant acid phosphatase have altered ruffled borders and disturbed intracellular vesicular transport. Exp. Cell Res. 2002, 279, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, K.; Nishimura, R.; Senn, J.; Youssef, R.F.; London, S.D.; Reddy, S.V. RANK ligand signaling modulates the matrix metalloproteinase-9 gene expression during osteoclast differentiation. Exp. Cell Res. 2007, 313, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.H.; Pang, E.K.; Kim, C.S.; Yoo, Y.J.; Cho, K.S.; Chai, J.K.; Kim, C.K.; Choi, S.H. Inhibitory effects of green tea polyphenol (−)-epigallocatechin gallate on the expression of matrix metalloproteinase-9 and on the formation of osteoclasts. J. Periodontal Res. 2004, 39, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Muraki, S.; Yamamoto, S.; Ishibashi, H.; Oka, H.; Yoshimura, N.; Kawaguchi, H.; Nakamura, K. Diet and lifestyle associated with increased bone mineral density: Cross-sectional study of Japanese elderly women at an osteoporosis outpatient clinic. J. Orthop. Sci. 2007, 12, 317–320. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Visagie, A.; Kasonga, A.; Deepak, V.; Moosa, S.; Marais, S.; Kruger, M.C.; Coetzee, M. Commercial Honeybush (Cyclopia spp.) Tea Extract Inhibits Osteoclast Formation and Bone Resorption in RAW264.7 Murine Macrophages—An in vitro Study. Int. J. Environ. Res. Public Health 2015, 12, 13779-13793. https://doi.org/10.3390/ijerph121113779
Visagie A, Kasonga A, Deepak V, Moosa S, Marais S, Kruger MC, Coetzee M. Commercial Honeybush (Cyclopia spp.) Tea Extract Inhibits Osteoclast Formation and Bone Resorption in RAW264.7 Murine Macrophages—An in vitro Study. International Journal of Environmental Research and Public Health. 2015; 12(11):13779-13793. https://doi.org/10.3390/ijerph121113779
Chicago/Turabian StyleVisagie, Amcois, Abe Kasonga, Vishwa Deepak, Shaakirah Moosa, Sumari Marais, Marlena C. Kruger, and Magdalena Coetzee. 2015. "Commercial Honeybush (Cyclopia spp.) Tea Extract Inhibits Osteoclast Formation and Bone Resorption in RAW264.7 Murine Macrophages—An in vitro Study" International Journal of Environmental Research and Public Health 12, no. 11: 13779-13793. https://doi.org/10.3390/ijerph121113779