
Viral Infection Control in the Essential Oil-Bearing Rose Nursery: Collection Maintenance and Monitoring

 , , ,
, , , 
Abstract
:1. Introduction
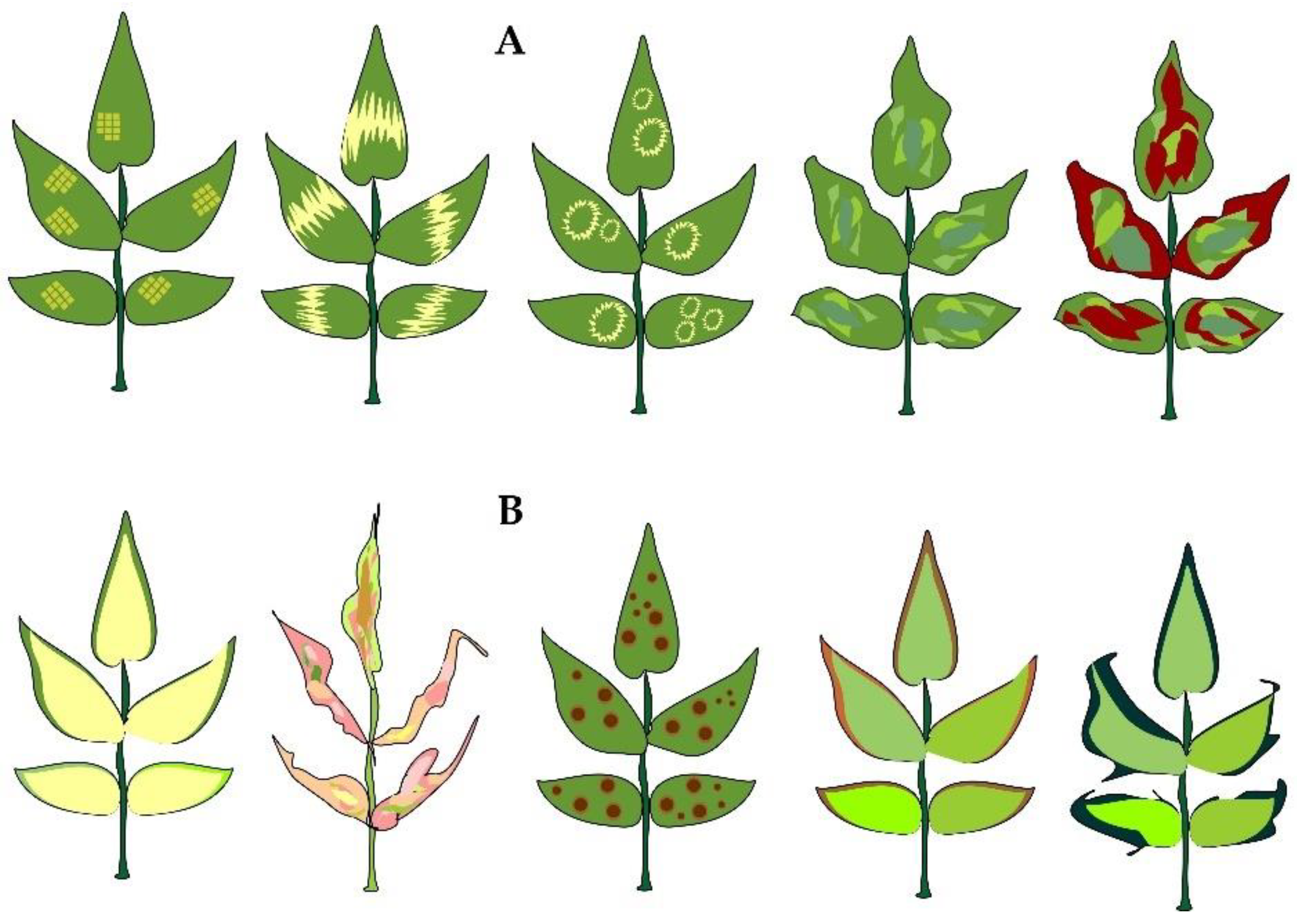
2. Overview of the Rosa L. Species Viruses
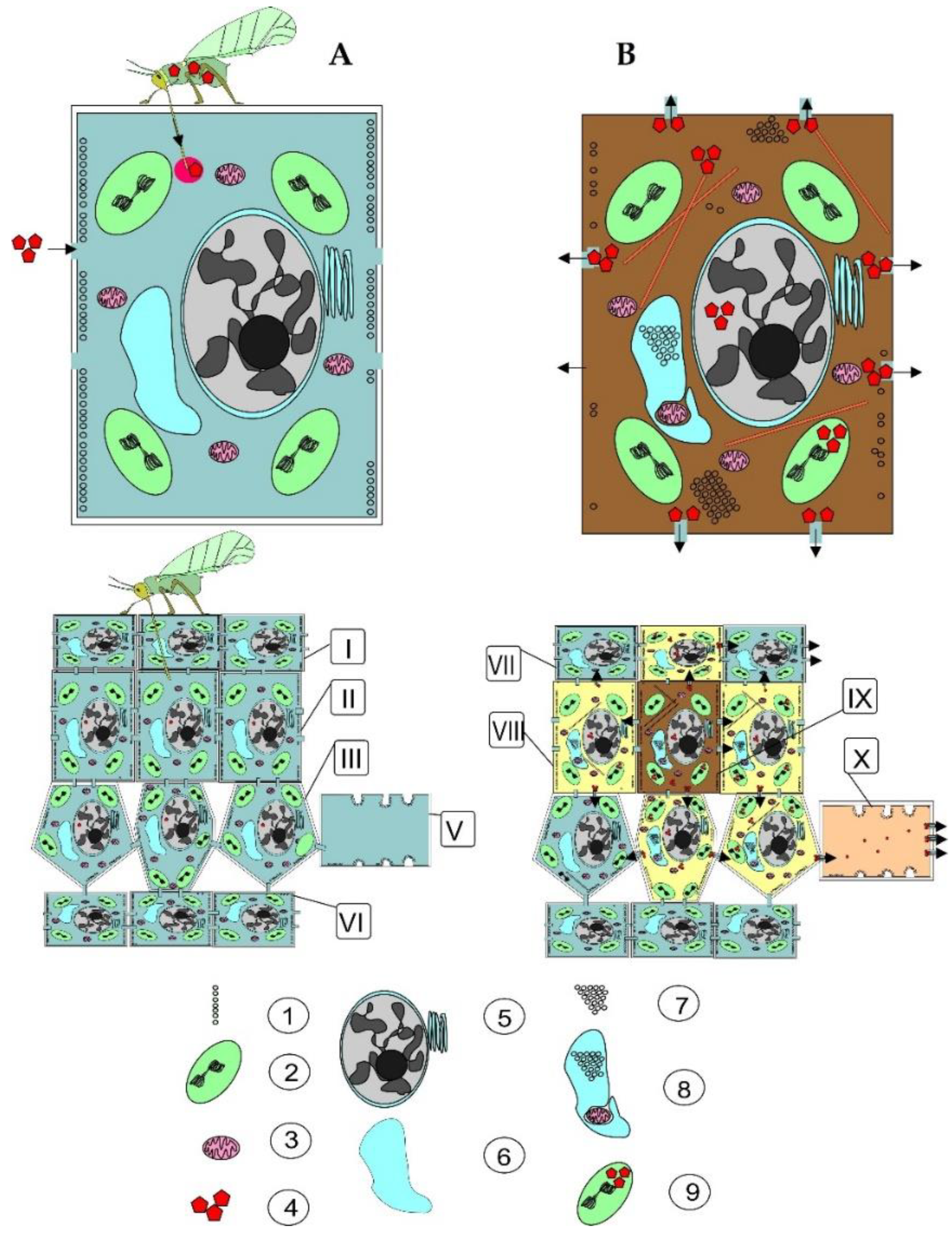
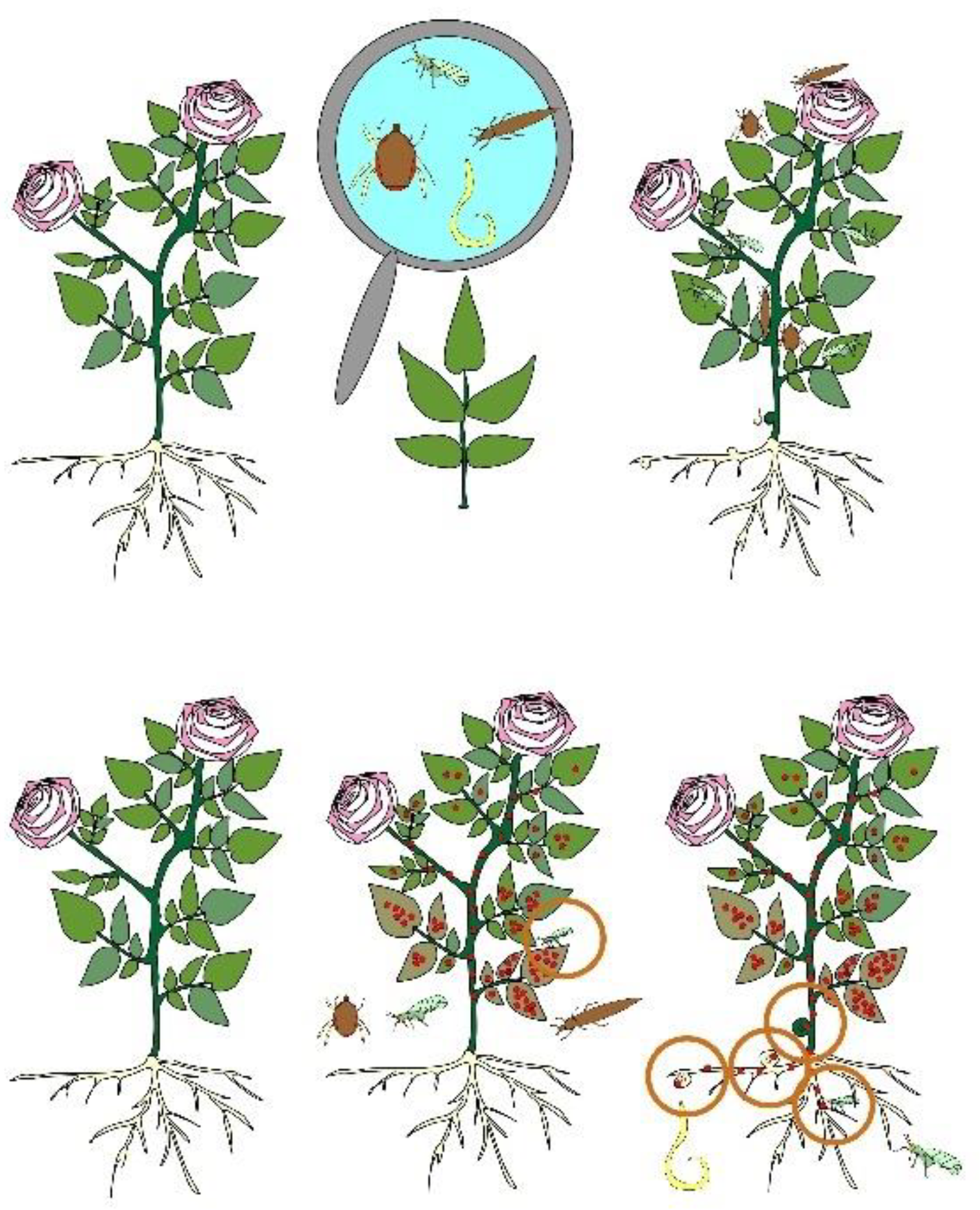
3. Spread of Rosa L. Plants’ Viruses
4. Rosa L. Plants Viruses Control
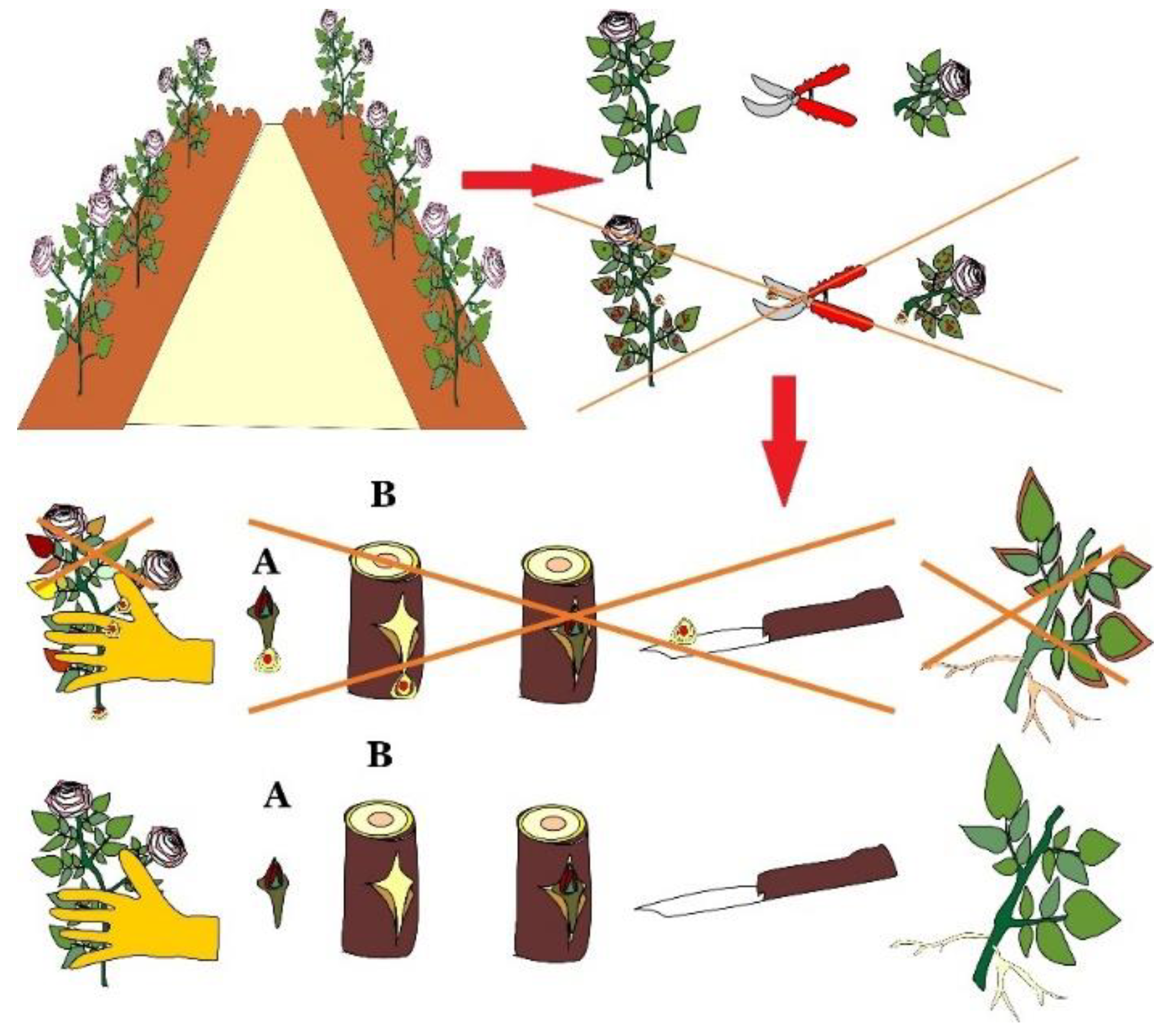
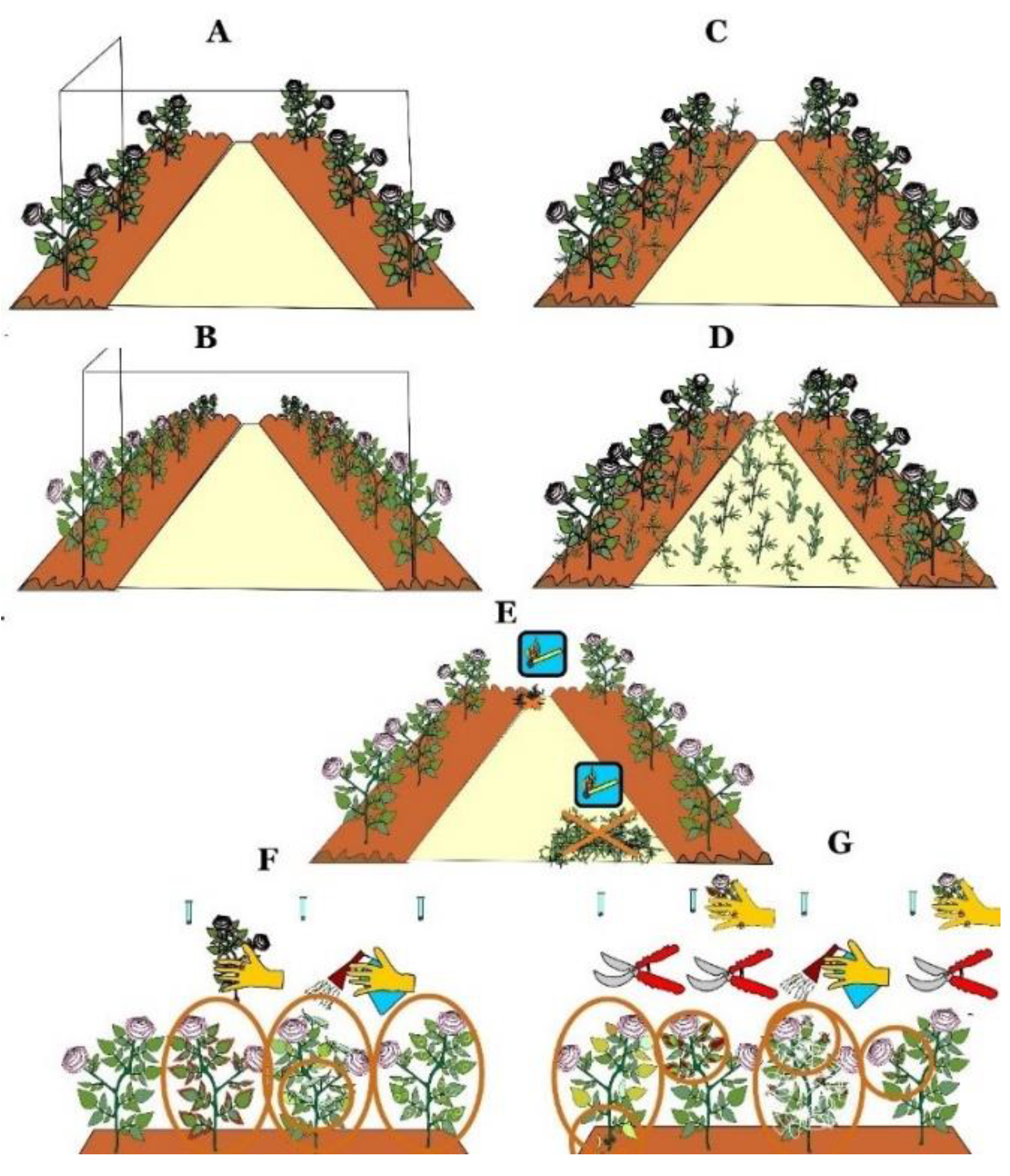
4.1. Traditional Methods of Viruses Control
4.2. Biotechnological Methods for Viruses Control
- Mother plant diagnosis using test plants, electron microscopy, ELISA and PCR techniques.
- In case the plant is infected, thermotherapy in vitro, or in vivo at 37 °C for 4–15 weeks, or chemotherapy with virucides in vitro.
- Plant tissue culture growth and plant regeneration on artificial nutrient media over 14–20 weeks.
- Regenerated plants adaptated in vitro at 15–20 °C over 3–4 weeks.
- The adapted plants retested using the test plants, ELISA and PCR techniques.
- Obtaining of virus-free plants and their certification.
5. Key Points and Current Prospects for Viral Disease Control in Essential Oil Rose Cultivation
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A. Genetic Structure of the Mosaic Viruses
Appendix A.1. Prunus Necrotic Ringspot Virus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RNA Type | RNA Chain Length (Number of Nucleotides) | Encoded Protein |
|---|---|---|
| RNA-1 | 3332 | Replicase P1 protein Methyltransferase/helicase) |
| RNA-2 | 2594 | Replicase P2 protein RNA-dependent RNA polymerase |
| RNA-3 | 1951 | Movement protein P3a and the coat protein |
Appendix A.2. Apple Mosaic Virus
| RNA Type | RNA Chain Length (Number of Nucleotides) | Encoded Protein |
|---|---|---|
| RNA-1 | 3476 | Methyltransferase/helicase) |
| RNA-2 | 2979 | RNA-dependent RNA polymerase |
| RNA-3 | 2056 | Movement protein and the coat protein |
Appendix A.3. Arabis Mosaic Virus
References
- Dobreva, A.; Kovatcheva, N.; Astatkie, T.; Zheljazkov, V.D. Improvement of essential oil yield of oil-bearing (Rosa damascena Mill.) due to surfactant and maceration. Ind. Crops Prod. 2011, 34, 1649–1651. [Google Scholar] [CrossRef]
- Petkova, M.; Tahsin, N.; Yancheva, S.; Yancheva, H. Development of the production of aromatic oil crops in Bulgaria. In China-Bulgaria Rural Revitalization Development Cooperation Forum; Institute of Agricultural Economics: Sofia, Bulgaria, 2018; pp. 71–86. [Google Scholar]
- Nenov, N.; Atanasova, T.; Gochev, V.; Merdzhanov, P.; Girova, T.; Djurkov, T.; Stoyanova, A. New product from Bulgarian rose. World Sci. 2016, 1, 17–22. [Google Scholar]
- Pashtetskiy, V.S.; Nevkrytaya, N.V.; Mishnev, A.V.; Nazarenko, L.G. Essential Oil Sector: Yesterday, Today, Tomorrow, 2nd ed.; IT “Arial”: Simferopol, Russia, 2018; 320p. [Google Scholar]
- Kiani, M.; Zamani, Z.; Khalighi, A.; Fatahi, R.; Byrne, D.H. Microsatellite analysis of Iranian Damask rose (Rosa damascena Mill.) germplasm. Plant Breed. 2010, 129, 551–557. [Google Scholar] [CrossRef]
- Redwan, T.; Nassour, M.; Mahfoud, H. Genetic Diversity of Rosa damascena Mill. in Latakia Province as Reveled by ISSR Analysis. SSRG—IJAES 2018, 5, 18–22. [Google Scholar] [CrossRef]
- Rusanov, K.; Kovacheva, N.; Rusanova, M.; Atanassov, I. Flower phenotype variation, essential oil variation and genetic diversity among Rosa alba L. accessions used for rose oil production in Bulgaria. Sci. Hortic. 2013, 161, 76–80. [Google Scholar] [CrossRef]
- Yardimci, N.; Çulal, H. Occurrence and incidence of Prunus necrotic ringspot virus, Arabis mosaic virus, and Apple mosaic virus on oil rose (Rosa damascena) in the Lakes region of Turkey. N. Z. J. Crop Hortic. Sci. 2009, 37, 95–98. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.M.; Darwish, H.Y.; Alamer, K.H. Microsatellite, inter simple sequence repeat and biochemical analyses of Rosa genotypes from Saudi Arabia. Afr. J. Biotechnol. 2017, 16, 552–557. [Google Scholar]
- Alotaibi, S.S.; Hassan, M.M.; Gaber, A.; Aljuaid, B.S. Genetic relationship and diversity of Taif-roses plant by using three different types of molecular markers. Res. J. Biotech. 2019, 14, 130–138. [Google Scholar]
- Gaurav, A.K.; Raju, D.V.S.; Ramkumar, M.K.; Singh, M.K.; Singh, B.; Krishnan, S.G.; Panwar, S.; Sevanthi, A.M. Genetic diversity analysis of wild and cultivated Rosa species of India using microsatellite markers and their comparison with morphology based diversity. J. Plant Biochem. Biotechnol. 2022, 31, 61–70. [Google Scholar] [CrossRef]
- Zolotilov, V.; Nevkrytaya, N.; Zolotilova, O.; Seitadzhieva, S.; Myagkikh, E.; Pashtetskiy, V.; Karpukhin, M. The Essential-Oil-Bearing Rose Collection Variability Study in Terms of Biochemical Parameters. Agronomy 2022, 12, 529. [Google Scholar] [CrossRef]
- Slavov, A.; Vasileva, I.; Stefanov, L.; Stoyanova, A. Valorization of wastes from the rose oil industry. Rev. Environ. Sci. Bio/Technol. 2017, 16, 309–325. [Google Scholar] [CrossRef]
- Nazarenko, L.G.; Korshunov, V.A.; Kochetkov, E.S. Essential Oil Rose Growing and Breeding; Tavriya: Simferopol, Ukraine, 2006; 216p. [Google Scholar]
- Zolotilov, V.A.; Nevkrytaya, N.V.; Zolotilova, O.M.; Skipor, O.B. The results of the essential oil rose selection for a high yield of concrete. Taurida Her. Agrar. Sci. 2020, 3, 93–104. [Google Scholar] [CrossRef]
- Collection of Essential-Oil, Spicy, Aromatic and Medicinal Plants. Available online: https://ckp-rf.ru/usu/507515/ (accessed on 22 February 2022).
- The State Register of Selection Achievements Authorized for Use (National List); Plant Cultivars (Official Publication); FGBNU “Rosinformagrotekh”: Moscow, Russia, 2021; Volume 1, 719p, Available online: https://gossortrf.ru/ (accessed on 22 February 2022).
- Stancheva, I.; Rosnev, B. Ornamental and Forest Crops Diseases; Pensoft: Sofia, Bulgaria, 2005; pp. 150–162. [Google Scholar]
- Treyvas, L.Y. Roses, Conifers and Other Ornamental Crops Diseases and Pests; Phyton: Moscow, Russia, 2017; pp. 5–115. [Google Scholar]
- Zakubanskiy, A.V.; Chirkov, S.N.; Mitrofanova, O.V.; Mitrofanova, I.V. Viruses of some valuable fruits, essential-oil and ornamental plants (Overview). Bull. State Nikitsk. Bot. Gard. 2016, 121, 7–18. [Google Scholar]
- Keldysh, M.A.; Chervyakova, O.N. Viral diseases of roses and their prevention in protected ground conditions. Gavrish 2008, 2, 16–21. [Google Scholar]
- EPPO Global Database. Available online: https://gd.eppo.int/ (accessed on 22 February 2022).
- Certification scheme for rose. EPPO Bull. 2002, 32, 159–177.
- Milleza, E.J.M.; Ward, L.I.; Delmiglio, C.; Tang, J.Z.; Veerakone, S.; Perez-Egusquiza, Z. Survey of viruses infecting Rosa spp. in New Zealand. Australas. Plant Pathol. 2013, 42, 313–320. [Google Scholar] [CrossRef]
- Moury, B.; Cardin, L.; Onesto, J.P.P.; Candresse, T.; Poupet, A. Survey of Prunus necrotic ringspot virus in Rose and Its Variability in Rose and Prunus spp. Phytopathology 2001, 91, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Pscheidt, J.W.; Rodriguez, T.G. Diseases of Rose. In Handbook of Florists’ Crops Diseases; McGovern, R.J., Elmer, W.H., Eds.; Springer: Cham, Switzerland, 2018; pp. 736–737. [Google Scholar]
- Sastry, S.K.; Mandal, B.; Hammond, J.; Scott, S.W.; Briddon, R.W. Encyclopedia of Plant Viruses and Viroids; Springer: New Delhi, India, 2019; pp. 2092–2106. [Google Scholar]
- Unified Register of the Eurasian Economic Union Quarantine Pests. Available online: https://docs.cntd.ru/document/456047397 (accessed on 22 February 2022).
- Khatri, S.; Nahid, N.; Fauquet, C.M.; Mubin, M.; Nawaz-ul-Rehman, M.S. A betasatellite-dependent begomovirus infects ornamental rose: Characterization of begomovirus infecting rose in Pakistan. Virus Genes 2014, 49, 124–131. [Google Scholar] [CrossRef]
- Sahu, A.K.; Marwal, A.; Shahid, M.S.; Nehra, C.; Gaur, R.K. First report of a begomovirus and associated betasatellite in Rosa indica and in India. Australas. Plant Dis. Notes. 2014, 9, 147. [Google Scholar] [CrossRef] [Green Version]
- Epstein, A.H.; Hill, J.H. Status of Rose Rosette Disease as a Biological Control for Multiflora Rose. Plant Dis. 1999, 83, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, P.; Das, S.; Saha, B.; Karmakar, A.; Saha, D.; Saha, A. Rose rosette virus: An emerging pathogen of garden roses in India. Australas. Plant Pathol. 2017, 46, 223–226. [Google Scholar] [CrossRef]
- Solo, K.M.; Collins, S.B.; Shires, M.K.; Ochoa, R.; Bauchan, G.R.; Schneider, L.G.; Henn, A.; Jacobi, J.C.; Williams-Woodward, J.L.; Hajimorad, M.R.; et al. A Survey of Rose rosette virus and Eriophyid Mites Associated with Roses in the Southeastern United States. HortScience 2020, 55, 1288–1294. [Google Scholar] [CrossRef]
- Salem, N.; Golino, D.A.; Falk, B.W.; Rowhani, A. Identification and Partial Characterization of a New Luteovirus Associated with Rose Spring Dwarf Disease. Plant Dis. 2008, 92, 508–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera, P.A.; Engel, E.A. Presence of rose spring dwarf-associated virus in Chile: Partial genome sequence and detection in roses and their colonizing aphids. Virus Genes 2010, 41, 295–297. [Google Scholar] [CrossRef]
- Barjadze, S.; Karaca, İ.; Yaşar, B.; Japoshvili, G. The yellow rose aphid Rhodobium porosum: A new pest of Damask rose in Turkey. Phytoparasitica 2011, 39, 59–62. [Google Scholar] [CrossRef]
- Xing, F.; Gao, D.; Wang, H.; Zhang, Z.; Habili, N.; Li, S. Molecular characterization of rose spring dwarf-associated virus isolated from China rose (Rosa chinensis Jacq.) in China. Arch. Virol. 2021, 166, 2059–2062. [Google Scholar] [CrossRef]
- Karanfil, A. Prevalence and molecular characterization of Turkish isolates of the rose viruses. Crop Prot. 2021, 143, 105565. [Google Scholar] [CrossRef]
- Mollov, D.; Lockhart, B. Symptoms, Transmission, and Detection of Four New Rose Viruses. Acta Hortic. 2015, 1064, 303–310. [Google Scholar] [CrossRef]
- Karanfil, A.; Randa-Zelyüt, F.; Ertunç, F.; Korkmaz, S. First report of Rose yellow vein virus in Turkey. New Dis. Rep. 2018, 38, 11. [Google Scholar] [CrossRef] [Green Version]
- Lockhart, B.; Zlesak, D.; Fetzer, J. Identification and Partial Characterization of Six New Viruses of Cultivated Roses in the USA. Acta Hortic. 2011, 901, 139–148. [Google Scholar] [CrossRef]
- Sabanadzovic, S.; Ghanem-Sabanadzovic, N.A. Molecular characterization and detection of a tripartite cryptic virus from rose. J. Plant Pathol. 2008, 90, 87–293. [Google Scholar]
- Vazquez-Iglesias, I.; Adams, I.P.; Hodgetts, J.; Fowkes, A.; Forde, S.; Ward, R.; Buxton-Kirk, A.; Kelly, M.; Santin-Azcona, J.; Skelton, A.; et al. High throughput sequencing and RT-qPCR assay reveal the presence of rose cryptic virus-1 in the United Kingdom. J. Plant Pathol. 2019, 101, 1171–1175. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Yang, Z.; Hong, N.; Wang, G.; Ning, G.; Xu, W. Deep sequencing reveals a novel closterovirus associated with wild rose leaf rosette disease. Mol. Plant Pathol. 2015, 16, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Soltani, N.; Golino, D.A.; Al Rwahnih, M. First Report of Rose Leaf Rosette-Associated Virus Infecting Rose (Rosa spp.) in California, USA. Plant Dis. 2021, 105, 2740. [Google Scholar] [CrossRef] [PubMed]
- Phelan, J.; James, D. Complete genome sequences of a putative new alphapartitivirus detected in Rosa spp. Arch. Virol. 2016, 161, 2623–2626. [Google Scholar] [CrossRef] [PubMed]
- Ohata, Y.; Nishio, T.; Tsuda, S. First isolation of rose yellow mosaic virus in Japan. J. Gen. Plant Pathol. 2021, 87, 295–299. [Google Scholar] [CrossRef]
- Diaz-Lara, A.; Mollov, D.; Golino, D.; Al Rwahnih, M. Complete genome sequence of rose virus A, the first carlavirus identified in rose. Arch. Virol. 2020, 165, 241–244. [Google Scholar] [CrossRef]
- Katsiani, A.T.; Maliogka, V.I.; Candresse, T.; Katis, N.I. Host-range studies, genetic diversity and evolutionary relationships of ACLSV isolates from ornamental, wild and cultivated Rosaceous species. Plant Pathol. 2014, 63, 63–71. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Chen, Y.S.; Wang, Z.H.; Wang, L.P.; Wang, G.P.; Hong, N.; Xu, W.X. First Report of Apple stem grooving virus Infecting Rosa chinensis in China. Plant Dis. 2016, 100, 1252. [Google Scholar] [CrossRef]
- Tzanetakis, I.E.; Gergerich, R.C.; Martin, R.R. A new Ilarvirus found in rose. Plant Pathol. 2006, 55, 568. [Google Scholar] [CrossRef]
- Awasthi, P.; Dhyani, D.; Ram, R.; Zaidi, A.A.; Hallan, V. Wild roses as natural reservoirs of Cherry necrotic rusty mottle virus. Eur. J. Plant Pathol. 2015, 142, 403–409. [Google Scholar] [CrossRef]
- Shahraeen, N.; Ghotbi, T.; Mehraban, A.H. Occurrence of Impatiens necrotic spot virus in Ornamentals in Mahallat and Tehran Provinces in Iran. Plant Dis. 2002, 86, 694. [Google Scholar] [CrossRef]
- Rafizadeh, N.; Jafarpour, B.; Falahati Rastegar, M. Detection of Iris yellow spot virus (IYSV) in onion and some of ornamental plants by ELISA and RT-PCR methods in Khorosan Razavi provinces. J. Plant Protect. Res. (Agric. Sci. Technol.) 2013, 27, 149–158. [Google Scholar]
- von Bargen, S.; Demiral, R.; Büttner, C. First detection of Raspberry ringspot virus in mosaic diseased hybrid roses in Germany. New Dis. Rep. 2015, 32, 18. [Google Scholar] [CrossRef] [Green Version]
- Kulshrestha, S.; Hallan, V.; Raikhy, G.; Ram, R.; Zaidi, A.A. Strawberry latent ringspot virus Infecting Roses in India. Plant Dis. 2004, 88, 86. [Google Scholar] [CrossRef]
- Sattary, M.; Rakhshanderoo, F.; Mozafari, J. First report of a mosaic disease caused by Tomato ringspot virus on rose and almond plants in Iran. Plant Pathol. 2015, 97, 393. [Google Scholar]
- Moini, A.A.; Izadpanah, K. New hosts for Tomato spotted wilt virus in Tehran. Iran. J. Plant Pathol. 2000, 36, 104–105. [Google Scholar]
- Matthews, R.E.F. Plant Virology; Mir: Moscow, Russia, 1973; 686p. [Google Scholar]
- Anikina, I.N.; Seitzhanova, L.L. Phytovirusology; Kereku: Pavlodar, Kazakhstan, 2015; pp. 26–27. [Google Scholar]
- Golino, D.A.; Sim, S.T.; Cunningham, M.; Rowhani, A. Transmission of Rose Mosaic Viruses. Acta Hortic. 2007, 751, 217–224. [Google Scholar] [CrossRef]
- Kiliç, H.Ç.; Yardimci, N.; Gübür, Ş. Serological, biological and molecular detection of Prunus necrotic ringspot virus on Rosa damascena Mill. in Turkey. Acta Sci. Pol. Hortorum Cultus. 2017, 16, 145–150. [Google Scholar]
- Sertkaya, G. An nvestigation on Rose Mosaic Disease of Rose in Hatay-Turkey. Julius-Kühn-Archiv 2010, 427, 309–313. [Google Scholar]
- da Silva, S.; Babu, B.; Paret, M.L.; Knox, G.; Iriarte, F.; Riddle, B.; Folimonova, S.Y. Rose Mosaic Virus: A Disease Caused by a Virus Complex and Symptoms on Roses and Management Practices. EDIS 2018, 4, 1–5. [Google Scholar] [CrossRef]
- Valasevich, N.; Cieślińska, M.; Kolbanova, E. Molecular characterization of Apple mosaic virus isolates from apple and rose. Eur. J. Plant Pathol. 2015, 141, 839–845. [Google Scholar] [CrossRef]
- Cui, H.; Hong, N.; Wang, G.; Wang, A. Genomic Segments RNA1 and RNA2 of Prunus necrotic ringspot virus Codetermine Viral Pathogenicity to Adapt to Alternating Natural Prunus Hosts. Mol. Plant-Microbe Interact. 2013, 26, 515–527. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.G.; Liu, H.Z.; Chen, J.; Zhou, J.F.; Qu, L.N.; Su, J.M.; Wang, G.P.; Hong, N. Genetic diversity of Prunus necrotic ringspot virus infecting stone fruit trees grown at seven regions in China and differentiation of three phylogroups by multiplex RT-PCR. Crop Prot. 2015, 74, 30–36. [Google Scholar] [CrossRef]
- Huo, Y.-Y.; Li, G.-F.; Qiu, Y.-H.; Li, W.-M.; Zhang, Y.-J. Rapid Detection of Prunus necrotic ringspot virus by Reverse Transcription-cross-priming Amplification Coupled with Nucleic Acid Test Strip Cassette. Sci. Rep. 2017, 7, 16175. [Google Scholar] [CrossRef] [Green Version]
- Khalid, A.; Adel, R. Prunus necrotic ringspot virus in apricot (Prunus armeniaca) and peach (P. persica) newly reported in Saudi Arabia. New Dis. Rep. 2011, 23, 26. [Google Scholar]
- Mekuria, G.; Ramesh, S.A.; Alberts, E.; Bertozzi, T.; Wirthensohn, M.; Collins, G.; Sedgley, M. Comparison of ELISA and RT-PCR for the detection of Prunus necrotic ring spot virus and prune dwarf virus in almond (Prunus dulcis). J. Virol. Methods. 2003, 114, 65–69. [Google Scholar] [CrossRef]
- Paduch-Cichal, E.; Sala-Rejczak, K. Biological and molecular characterization of Prunus necrotic ringspot virus isolates from three rose cultivars. Acta Physiol. Plant. 2011, 33, 2349–2354. [Google Scholar] [CrossRef] [Green Version]
- Ulubas, C.; Ertunc, F. RT-PCR Detection and Molecular Characterization of Prunus necrotic ringspot virus Isolates Occurring in Turkey. Phytopathology 2004, 152, 498–502. [Google Scholar] [CrossRef]
- Xing, F.; Gao, D.; Liu, H.; Wang, H.; Habili, N.; Li, S. Molecular characterization and pathogenicity analysis of prunus necrotic ringspot virus isolates from China rose (Rosa chinensis Jacq.). Arch. Virol. 2020, 165, 2479–2486. [Google Scholar] [CrossRef]
- Grimová, L.; Winkowska, L.; Konrady, M.; Ryšánek, P. Apple mosaic virus. Phytopathol. Mediterr. 2016, 55, 1–19. [Google Scholar]
- Sánchez-Navarro, J.A.; Aparicio, F.; Herranz, M.C.; Minafra, A.; Myrta, A.; Pallás, V. Simultaneous detection and identification of eight stone fruit viruses by one-step RT-PCR. Eur. J. Plant Pathol. 2005, 111, 77–84. [Google Scholar] [CrossRef]
- Meng, B.; Martelli, G.P.; Golino, D.A.; Fuchs, M. Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer: Cham, Switzerland, 2018; 698p. [Google Scholar]
- Grimová, L.; Winkowska, L.; Ryšánek, P.; Svoboda, P.; Petrzik, K. Reflects the coat protein variability of apple mosaic virus host preference? Virus Genes 2013, 7, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Martelli, G.P.; Taylor, C.E. Distribution of Viruses and Their Nematode Vectors. In Advances in Disease Vector Research; Harrys, K.F., Ed.; Springer: New York, NY, USA, 1990; pp. 151–189. [Google Scholar]
- Wetzel, T.; Beck, A.; Wegener, U.; Krczal, G. Complete nucleotide sequence of the RNA 1 of a grapevine isolate of Arabis mosaic virus. Arch. Virol. 2004, 149, 989–995. [Google Scholar] [PubMed]
- Drobotova, E.N. Pests of essential oil crops grown at the Research Institute of Agriculture of Crimea. In Proceedings of the Current State, Problems and Prospects of Agricultural Science Development, Simferopol, Russia, 5–9 October 2020. [Google Scholar]
- Zolotilov, V.A.; Nevkrytaya, N.V. Laying and Exploitation of Essential Oil Roses Breed Sheds: Methodological Recommendations; FSBSI “Research Institute of Agriculture of Crimea”: Simferopol, Russia, 2017; 28p. [Google Scholar]
- Pashtetskiy, V.S.; Skipor, O.B.; Kravchenko, G.D.; Zolotilova, O.M.; Zolotilov, V.A.; Myagkih, E.F.; Verdysh, M.V.; Popova, A.A.; Polyakova, N.Y.; Kolesnikova, A.V. Technologies of Traditional and Perspective Essential Oil Cultures Breeding in the Republic of Crimea; IT “Arial”: Simferopol, Russia, 2020; 36p. [Google Scholar]
- Vazquez-Iglesias, I.; Ochoa-Corona, F.M.; Tang, J.; Robinson, R.; Clover, G.R.; Fox, A.; Boonham, N. Facing Rose rosette virus: A risk to European rose cultivation. Plant Pathol. 2020, 69, 1603–1617. [Google Scholar] [CrossRef]
- Özbek, H.; Çalmaşur, O. A Review of Insects and Mites Associated with Roses in Turkey. In Proceedings of the I International Rose Hip Conference 690, Erzurum, Turkey, 7 September 2004. [Google Scholar]
- Smitha, R.; Rajendran, P.; Sandhya, P.T.; Aparna, V.S.; Rajees, P.C. Insect pest complex of rose at Regional Agricultural Research Station, Ambalavayal, Wayanad. In Proceedings of the International Symposium on Succulents and Other Ornamentals 1165, Wayanad, India, 24–27 January 2016. [Google Scholar]
- Dogadina, M.A.; Stavtseva, T.I.; Botuz, N.I. Pests of roses of the open ground in the conditions of the Orel region. Hortic. Vinic. 2015, 6, 47–52. [Google Scholar]
- Sergeeva, O.; Barinov, M.; Medvedeva, K. Study of the twospotted spider mite harmfulness on roses in the Pavlovsk State reserve museum. In Proceedings of the Scientific Contribution of Young Researchers to the Preservation of Traditions and Development of Agroindustrial Complex, Saint Petersburg, Russia, 31 March–1 April 2016. [Google Scholar]
- Balykina, E.B.; Klimenko, Z.K.; Zvonareva, L.N.; Plugatar, S.A.; Rybareva, T.S. Pests and diseases of garden roses cultivars from the collection of the Nikitskiy Botanical Garden. Bull. Tver. State Univ. 2017, 4, 92–102. [Google Scholar]
- Pizetta, P.U.C.; Pivetta, K.F.L.; Santos, J.M.; Batista, G.S.; Gimenes, R.; Martins, T.A. Resistance of Rose Rootstocks to Meloidogyne hapla Nematode. In Proceedings of the II International Conference on Landscape and Urban Horticulture 881, Bologna, Italy, 9–13 June 2009. [Google Scholar]
- Meressa, B.H.; Dehne, H.W.; Hallmann, J. Population Dynamics and Damage Potential of Meloidogyne hapla to Rose Rootstock Species. J. Phytopathol. 2016, 164, 711–721. [Google Scholar] [CrossRef]
- Manners, A.G.; Dembowski, B.R.; Healey, M.A. Biological control of western flower thrips, Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae), in gerberas, chrysanthemums and roses. Aust. J. Entomol. 2013, 52, 246–258. [Google Scholar] [CrossRef] [Green Version]
- Avellaneda, J.; Díaz, M.; Coy-Barrera, E.; Rodríguez, D.; Osorio, C. Rose volatile compounds allow the design of new control strategies for the western flower thrips (Frankliniella occidentalis). J. Pest Sci. 2021, 94, 129–142. [Google Scholar] [CrossRef]
- Mitrofanova, I.V.; Mitrofanova, O.V.; Brailko, V.A.; Lesnikova-Sedoshenko, N.P. Biotechnological and physiological features of the essential oil rose valuable genotypes in vitro cultivation. News Univ. Appl. Chem. Biotechnol. 2015, 2, 37–48. [Google Scholar]
- Pati, P.K.; Rath, S.P.; Sharma, M.; Sood, A.; Ahuja, P.S. In vitro propagation of rose—A review. Biotechnol. Adv. 2006, 24, 94–114. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.; Rebek, E.J.; Schnelle, M.A. Rose Rosette Disease. Available online: https://shareok.org/bitstream/handle/11244/319875/oksa_epp_7329_2017-03.pdf?sequence=1 (accessed on 22 February 2022).
- Pashtetskiy, V.S.; Timasheva, L.A.; Pekhova, O.A.; Danilova, I.L.; Serebryakova, O.A. Essential Oils and Their Quality; IT “Arial”: Simferopol, Russia, 2021; 212p. [Google Scholar]
- Panattoni, A.; Luvisi, A.; Triolo, E. Review. Elimination of viruses in plants: Twenty years of progress. Span. J. Agric. Res. 2013, 1, 173–188. [Google Scholar] [CrossRef] [Green Version]
- Mitrofanova, O.V.; Mitrofanova, I.V.; Lesnikova-Sedoshenko, N.P.; Ivanova, N.N. Application of biotechnological methods in plant health improvement and virus-free planting material of promising flower and ornamental crops propagation. Plant Biol. Hortic. Theory Innov. 2014, 138, 5–56. [Google Scholar]
- Yegorova, N.; Stavtzeva, I.; Zolotilov, V. Micropropagation in vitro of essential oil rose hybrids obtained in embryoculture. In Proceedings of the BIO Web of Conferences, Simferopol, Russia, 28 October 2021. [Google Scholar]
- Khosh-Khui, M. Biotechnology of Scented Roses: A Review. Int. J. Hortic. Sci. Technol. 2014, 1, 1–20. [Google Scholar]
- Grondeau, C.; Samson, R.; Sands, D.C. A Review of Thermotherapy to Free Plant Materials from Pathogens, Especially Seeds from Bacteria. CRC Crit. Rev. Plant Sci. 1994, 13, 57–75. [Google Scholar] [CrossRef]
- Lerch, B. On the inhibition of plant virus multiplication by ribavirin. Antivir. Res. 1987, 7, 257–270. [Google Scholar] [CrossRef]
- Yegorova, N.A.; Stavtseva, I.V.; Mitrofanova, I.V. Morphogenesis in the essential rose oil meristem culture during in vitro chemotherapy. Bull. State Nikitsk. Bot. Gard. 2017, 125, 65–72. [Google Scholar]
- Ghorbani, A.; Hadifar, S.; Salari, R.; Izadpanah, K.; Burmistrz, M.; Afsharifar, A.; Eskandari, M.H.; Niazi, A.; Denes, C.E.; Neely, G.G. A short overview of CRISPR-Cas technology and its application in viral disease control. Transgenic Res. 2021, 30, 221–238. [Google Scholar] [CrossRef]
- Khan, Z.A.; Kumar, R.; Dasgupta, I. CRISPR/Cas-Mediated Resistance against Viruses in Plants. Int. J. Mol. Sci. 2022, 23, 2303. [Google Scholar]
- Taliansky, M.; Samarskaya, V.; Zavriev, S.K.; Fesenko, I.; Kalinina, N.O.; Love, A.J. RNA-Based Technologies for Engineering Plant Virus Resistance. Plants 2021, 10, 82. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yang, X.; Zhou, G.; Zhang, T. Engineering plant virus resistance: From RNA silencing to genome editing strategies. Plant Biotechnol. J. 2020, 18, 328–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, D.; Maiss, E.; Adam, G.; Casper, R.M. Prunus necrotic ringspot ilarvirus: Nucleotide sequence of RNA3 and the relationship to other ilarviruses based on coat protein comparison. J. Gen. Virol. 1995, 76, 1073–1079. [Google Scholar] [CrossRef]
- Pallas, V.; Aparicio, F.; Herranz, M.C.; Sanchez-Navarro, J.A.; Scott, S.W. The molecular Biology of Ilarviruses. Adv. Virus Res. 2013, 87, 139–181. [Google Scholar] [PubMed]
- Koper-Zwarthoff, E.C.; Bol, J.F. Nucleotide sequence of the putative recognition site for coat protein in the RNAs of alfalfa mosaic virus and tobacco streak virus. Nucleic Acids Res. 1980, 8, 3307–3318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reusken, C.; Neeleman, L.; Bol, J.F. The 3′-untranslated region of alfalfa mosaic virus RNA 3 contains at least two independent binding sites for viral coat protein. Nucleic Acids Res. 1994, 22, 1346–1353. [Google Scholar] [CrossRef]
- Sehnke, P.C.; Mason, A.M.; Hood, S.J.; Lister, R.M.; Johnson, J.E. A “zinc-finger”-type binding domain in tobacco streak virus coat protein. Virology 1989, 168, 48–56. [Google Scholar] [CrossRef]
- Gao, F.; Lin, W.; Shen, J.; Liao, F. Genetic diversity and molecular evolution of arabis mosaic virus based on the CP gene sequence. Arch. Virol. 2016, 161, 1047–1051. [Google Scholar] [CrossRef]
- Sánchez-Navarro, J.A.; Pallás, V. Evolutionary relationships in the ilarviruses: Nucleotide sequence of prunus necrotic ringspot virus RNA 3. Arch. Virol. 1997, 142, 749–763. [Google Scholar] [CrossRef]
- Sánchez-Navarro, J.A.; Pallás, V. Nucleotide sequence of apple mosaic ilarvirus RNA 4. J. Gen. Virol. 1994, 75, 1441–1445. [Google Scholar] [CrossRef]
- Shiel, P.J.; Alrefai, R.H.; Domier, L.L.; Korban, S.S.; Berger, P.H. The complete nucleotide sequence of apple mosaic virus RNA-3. Arch. Virol. 1995, 140, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
- Shiel, P.J.; Berger, P.H. The complete nucleotide sequence of apple mosaic virus (ApMV) RNA 1 and RNA 2: ApMV is more closely related to alfalfa mosaic virus than to other ilarviruses. J. Gen. Virol. 2000, 81, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, T.; Meunier, L.; Jaeger, U.; Reustle, G.M.; Krczal, G. Complete nucleotide sequences of the RNAS 2 of German isolates of Grapevine fanleaf and Arabis mosaic nepoviruses. Virus Res. 2001, 75, 139–145. [Google Scholar] [CrossRef]





| Virus | Acronym | Reference |
|---|---|---|
| Prunus necrotic ringspot virus | PNRSV | [20,23,24,25,26,27] |
| Apple mosaic virus | ApMV | |
| Arabis mosaic virus | ArMV | |
| Tobacco streak virus | TSV | |
| Tobacco ringspot virus | TRSV | |
| Tomato ringspot virus | ToRSV | |
| Tomato spotted wilt virus | TSWV | |
| Tomato yellow ring virus | TYRV | |
| Strawberry latent ringspot virus | SLRSV | |
| Blackberry chlorotic ringspot virus | BCRV | |
| Raspberry ringspot virus | RpRSV | |
| Cherry necrotic rusty mottle virus | CNRMV | |
| Impatiens necrotic spot virus | INSV | |
| Apple chlorotic leaf spot virus | ACLSV | |
| Apple stem grooving virus | ASGV | |
| Iris yellow spot virus | IYSV | |
| Rose rosette virus | RRV | |
| Rose leaf curl virus | RoLCuV | |
| Rose spring dwarf-associated virus | RSDaV | |
| Rose yellow vein virus | RYVV | |
| Rose cryptic virus-1 | RoCV-1 | |
| Rose yellow mosaic virus | RoYMV | |
| Rosa rugosa leaf distortion virus | RrLDV | |
| Rose yellow leaf virus | RYLV | |
| Rose chlorotic ringspot virus | RoCRSV | |
| Rose necrotic mosaic virus | RoNMV | |
| Rose leaf rosette-associated virus | RLRaV | |
| Rose colour break virus | RCBV |
| Virus | Symptoms | Virus Spread Mechanism | Country | Reference |
|---|---|---|---|---|
| Rose leaf curl virus | Pronounced leaf stunted growth and curling | Not specified | Pakistan | [29] |
| Dwarfing, leaf distortion and leaf curling | Not specified | India | [30] | |
| Rose rosette virus | Shoot elongation and colouring from light pink to dark purple; thorn proliferation; leaf elongation, distortion and red pigmentation; petioles shortening; reduced flowering; lateral buds coming out of dormancy, growing and colouring red. | Eriophyid mite Phyllocoptes fructiphylus | USA | [31] |
| Leaf curling and puckering; flower distortion; persistent red pigmentation | Not specified | India | [32] | |
| Excessive thorn production; “witch’s broom” rosetting; abundance of lateral shoots; shoots coloring red; leaves and flowers mottling or distortion; lateral shoot growth. | Eriophyid mite Phyllocoptes fructiphilus | USA | [33] | |
| Rose spring dwarf-associated virus | Rosetting; leaves shortening with vein clearing or netting; shoot zigzag growth pattern. | Aphids Metapolophium dirhodum and Rhodobium porosum | USA | [34] |
| Yellow vein chlorosis | Aphid Rhodobium porosum | Chile | [35] | |
| Not specified | Aphid Rhodobium porosum | Turkey | [36] | |
| Leaf rosetting | Aphid Metapolophium dirhodum | New Zealand | [24] | |
| Not specified | Not specified | China | [37] | |
| Not specified | Not specified | Turkey | [38] | |
| Rose yellow vein virus | Not specified | Not specified | New Zealand | [24] |
| Mosaic and vein yellowing | Grafting | USA | [39] | |
| Vein banding, central vein chlorosis | Not specified | Turkey | [40] | |
| Rosa rugosa leaf distortion virus | Leaf stunted growth and distortion; pale circular lines appearing only on early spring growth | Not specified | USA | [41] |
| Leaf distortion and stunted growth; pale circular lines appearing only on early spring growth | Grafting | USA | [39] | |
| Vein yellowing | Not specified | Turkey | [38] | |
| Rose color break virus | Deformed, flecked and streaked petals | Sap-transmission and use of infected budwood | Egypt | [27] |
| Rose yellow leaf virus | Leaf premature yellowing and senescence | Not specified | USA | [41] |
| Leaf premature yellowing and senescence | Grafting | USA | [39] | |
| Rose cryptic virus 1 | Not specified | Not specified | USA | [42] |
| Leaf mottling and necrosis | Not specified | New Zealand | [24] | |
| Leaf banding, mottling and distortion | Not specified | UK | [43] | |
| Mottling | Not specified | Turkey | [38] | |
| Rose transient mosaic virus | Leaf mosaic and yellowing | Not specified | Minnesota, the USA | [41] |
| Rose leaf rosette-associated virus | Leaf rosette (“witch’s broom” symptom) formed by dense small leaves on branches; clearly noticeable decay, destruction and, finally, dieback of plants | Not specified | China | [44] |
| Not specified | Not specified | USA | [45] | |
| Rose necrotic mosaic virus | Mosaic, necrotic streaks, leaf distortion | Not specified | USA | [41] |
| Rose partitivirus | Not specified | Not specified | Canada | [46] |
| Rose yellow mosaic virus | Yellow mosaic; ring mosaic; premature leaf senescence and dark-brown rings on canes | Not specified | Minnesota, the USA | [41] |
| Yellow mosaic; premature leaf senescence | Grafting | Minnesota, the USA | [39] | |
| Yellow chlorotic spots | Not specified | Japan | [47] | |
| Rose Chlorotic Ringspot Virus | Chlorotic ringspots and mosaic symptoms | Not specified | Minnesota, the USA | [41] |
| Rose virus A | Leaf distortion; mosaic symptoms | Not specified | California, the USA | [48] |
| Virus | Symptoms | Virus Spread Mechanism | Country | Reference |
|---|---|---|---|---|
| Apple chlorotic leaf spot virus | Not specified | Vegetative propagation and grafting | Greece | [49] |
| Apple stem grooving virus | Leaf rosette | Sap-transmission, transmission on grafting and via infected seed material [27] | China | [50] |
| Blackberry chlorotic ringspot virus | Leaf rosette | Sap-transmission, transmission on grafting [27] | China | [44] |
| Not specified | USA | [51] | ||
| Cherry necrotic rusty mottle virus | Chlorosis and necrotic spots on leaves | Transmission via budding and grafting [27] | India | [52] |
| Impatiens necrotic spot orthotospovirus | Small necrotic spots; leaves yellowing; ringspots; necrotic streaks; wilting and dwarf symptoms | Transmitted by thrips Frankliniella occidentalis; sap-transmission; use of infected budwood [27] | Iran | [53] |
| Iris yellow spot orthotospovirus | Chlorotic and necrotic symptoms | Thrips, sap-transmission [27] | Iran | [54] |
| Raspberry ringspot virus | Mosaic; chlorosis; leaves curling and distortion; stunted growth | Nematodes, sap-transmission [27] | Germany | [55] |
| Strawberry latent ringspot virus | Yellow flecking in young leaves and reduction in leaflet size | Nematodes, sap-transmission; grafting [27] | India | [56] |
| Tomato ringspot virus | Banded chlorosis wrinkling; malformation and chlorotic spots on leaves | Nematodes (Xiphinema spp.), sap-transmission, grafting [27] | Iran | [57] |
| Tomato spotted wilt orthotospovirus | Necrotic spots and leaves marginal necrosis | Thrips, sap-transmission, grafting [27] | Iran | [58] |
| Virus | Taxonomic Affiliation | Host Plants | Geographical Region/Country | Symptoms |
|---|---|---|---|---|
| Prunus necrotic ringspot virus | Family: Bromoviridae Genus: Ilarvirus [22] | Apple tree (Malus domestica); white mulberry (Morus alba); red mulberry (Morus rubra); sour cherry tree (Prunus cerasus); oriental cherry (Prunus serrulata); sweet cherry (Prunus avium); almond (Prunus dulcis); peach tree (Prunus persica); Japanese plum (Prunus salicina); garden plum (Prunus domestica); apricot tree (Prunus armeniaca); Japanese apricot (Prunus mume); common hop (Humulus lupulus); rose (Rosa spp.) [22,25,66,67,68,69,70,71,72,73] | Africa: Algeria, Egypt, Morocco, South Africa, Tunisia. America: Argentina, Brazil, Canada, Chile, Mexico, USA, Uruguay. Asia: China, India, Iran, Israel, Jordan, Japan, Korea, Lebanon, Saudi Arabia, Syria. Oceania: Australia, Fiji, New Zealand. Europe: Albania, Austria, Belgium, Bosnia and Herzegovina, Bulgaria, Croatia, Cyprus, Czech Republic, Denmark, France, Germany, Greece, Hungary, Italy, Latvia, Malta, Moldova, Montenegro, Netherlands, Poland, Russia, Portugal, Romania, Serbia, Slovakia, Slovenia, Spain, Sweden, Turkey, Ukraine, Great Britain [22]. | Chlorotic and necrotic ringspots, mottling and vein banding develop on leaves. Stems are covered with necrotic stripes. Flowers are distorted due to a significant diameter reduction, loss of fresh and dry weight as well as petal discoloration and decrease in number [62,66,70,71,73] |
| Apple mosaic virus | Family: Bromoviridae Genus: Ilarvirus [22] | Some representatives of the chestnut tree genus (Aesculus); birch tree Betula); Prunus (apricot, peach, cherry, plum, cherry plum); Rosa; Rubus (raspberry, blackberry, blackcurrant); hawthorn (Crataegus); wormwood (Artemisia vulgaris); hazel (Corylus avellana); strawberry (Fragaria ananassa); common hop (Humulus lupulus); apple tree (Malus domestica); common pear (Pyrus communis); redcurrant (Ribes rubrum); wild clary (Salvia verbenaca); rowan (Sorbus aucuparia) [8,22,74,75] | Africa: Algeria, Ethiopia, Kenya, Morocco, South Africa, Tunisia, Zimbabwe. America: Argentina, Brazil, Canada, Chile, Mexico, USA, Uruguay; Asia: China, India, Japan, Jordan, Lebanon, Syria. Europe: Austria, Azerbaijan, Belarus, Belgium, Bosnia and Herzegovina, Bulgaria, Cyprus, Czech Republic, Denmark, Finland, France, Germany, Greece, Hungary, Italy, Latvia, Netherlands, Norway, Poland, Portugal, Romania, Russia, Serbia, Slovakia, Slovenia, Spain, Sweden, Switzerland, Turkey, Ukraine, Great Britain. Oceania: Australia, New Zealand [22]. | Leaves turn wrinkled and distorted and manifest linear patterns, chlorotic ringspots, mottling and vein banding [65,74]. |
| Arabis mosaic virus | Family: Secoviridae Genus: Nepovirus [22] | Grapevine (Vitis vinifera); apricot (Prunus armeniaca); sweet cherry (Prunus avium); plum (Prunus domestica); almond (Prunus dulcis); cherry laurel (Prunus laurocerasus); peach (Prunus persica); rhubarb (Rheum rhabarbarum); raspberry (Rubus idaeus); black elderberry (Sambucus nigra); representatives of the genus Gladiolus; celery (Apium graveolens); horseradish (Armoracia rusticana); common beet (Beta vulgaris); strawberry (Fragaria ananassa); common hop (Humulus lupulus); lettuce (Lactuca sativa); olive (Olea europaea) [22,76] | Africa: Egypt, South Africa. America: Canada, Chile, Mexico, Peru, USA. Asia: Kazakhstan, India, Iran, Japan, Lebanon, Syria. Oceania: Australia, New Zealand. Europe: Austria, Belgium, Bulgaria, Croatia, Cyprus, Czech Republic, Denmark, Finland, France, Germany, Greece, Hungary, Ireland, Italy, Latvia, Lithuania, Luxembourg, Moldova, Netherlands, Norway, Poland, Russia, Romania, Serbia, Slovakia, Slovenia, Spain, Sweden, Switzerland, Turkey, Ukraine, Belarus, Great Britain [22,76]. | Chlorotic spots on the first leaves followed by mottling and ringspots. Later on the developing leaves show little to no symptoms [27]. |
| Pest | Mechanism of Damage to the Plant | Infection Symptoms | Reference |
|---|---|---|---|
| Mites | |||
| Phyllocoptes fructiphilus | Mites overwintering on plants and feeding on plant tissues; transfer by insects, wind and with clothes | Not specified | [83] |
| Tetranychus urticae | Not specified | Leaves mottling and drying | [84] |
| Toxic substances injected by insect | Leaves yellowing; reduced photosynthesis; petals darkening and falling | [85] | |
| Not specified | White tiny flecks at the points of puncture by the mite’s mouthparts on the upper surface of the leaf; plants pale yellowing; dull foliage; leaves and buds inlacing with a cobweb | [86] | |
| Mites feeding on the cell content mainly on the lower surface of the lamina causing destruction of the epidermis and underlying cells | Light spots on the upper surface of leaves; leaves turning yellow-brown and drying out | [87] | |
| Female mites wintering under the plant debris and the bark of shrubs; colonizing young leaves in spring; weaving a web and laying eggs while feeding | Leaves yellowing, distortion and drying; buds failing to open. | [88] | |
| Nematodes | |||
| Meloidogyne hapla | Not specified | Leaves yellowing and prematurely falling; small shoots; reduced productivity and quality of flowers (the stem length and the flower size); symptoms of mineral deficiency; roots bearing galls; necrosis, segments dying-off, bark reducing and failing, roots shrinking and cracking | [89] |
| Not specified | The root system distortion; leaf chlorosis; the stem size decreasing | [90] | |
| Sedentary internal parasites cutting tunnels in the plant root and creating permanent feeding sites without leaving them | Giant cells developing at the feeding site; hyperplasia of the cortical and vascular parenchyma; retarded meristematic activity in the root tips | [26] | |
| Xiphinema diversicaudatum | Migratory external parasites feeding outside the root system | Galls caused by the cortical cells’ hyperplasia developing at the feeding site; cells growing in size two–three fold; retarded meristematic activity | [26] |
| Thrips | |||
| Frankliniellaoccidentalis | Not specified | Retarded or stunted growth of leaves and transmission of certain plant viruses (for instance, Tomato spotted wilt virus). | [91] |
| Immature and adult specimens feeding on the plant tissues by means of their piercing-sucking mouthparts; damage caused by females’ saw-like ovipositor used for laying eggs in leaves, petioles, flower bracts and petals | Surface damage followed by necrotic spots; impaired photosynthesis capacity | [92] | |
| Frankliniella tritici | Not specified | Buds turning brownish; petals curling up | [84] |
| Thrips tabaci | Not specified | Impaired decorative value of leaves and flowers; white flecks on leaves and buds; leaves tarnishing, turning from green to various shades of brown and falling; decreased intensity of flowers’ colour and brightness; silvery dots on the petals developing into stripes | [86] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seitadzhieva, S.; Gulevich, A.A.; Yegorova, N.; Nevkrytaya, N.; Abdurashytov, S.; Radchenko, L.; Pashtetskiy, V.; Baranova, E.N. Viral Infection Control in the Essential Oil-Bearing Rose Nursery: Collection Maintenance and Monitoring. Horticulturae 2022, 8, 629. https://doi.org/10.3390/horticulturae8070629
Seitadzhieva S, Gulevich AA, Yegorova N, Nevkrytaya N, Abdurashytov S, Radchenko L, Pashtetskiy V, Baranova EN. Viral Infection Control in the Essential Oil-Bearing Rose Nursery: Collection Maintenance and Monitoring. Horticulturae. 2022; 8(7):629. https://doi.org/10.3390/horticulturae8070629
Chicago/Turabian StyleSeitadzhieva, Sevilia, Alexander A. Gulevich, Natalya Yegorova, Natalya Nevkrytaya, Suleiman Abdurashytov, Lyudmila Radchenko, Vladimir Pashtetskiy, and Ekaterina N. Baranova. 2022. "Viral Infection Control in the Essential Oil-Bearing Rose Nursery: Collection Maintenance and Monitoring" Horticulturae 8, no. 7: 629. https://doi.org/10.3390/horticulturae8070629