
Association between IRF6, TP63, GREM1 Gene Polymorphisms and Non-Syndromic Orofacial Cleft Phenotypes in Vietnamese Population: A Case–Control and Family-Based Study
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Study
2.2. Single-Nucleotide Polymorphism Selection and Genotyping
2.3. Statistical Analysis
3. Results
3.1. Baseline Characteristics
3.2. Single-Marker Association Analysis
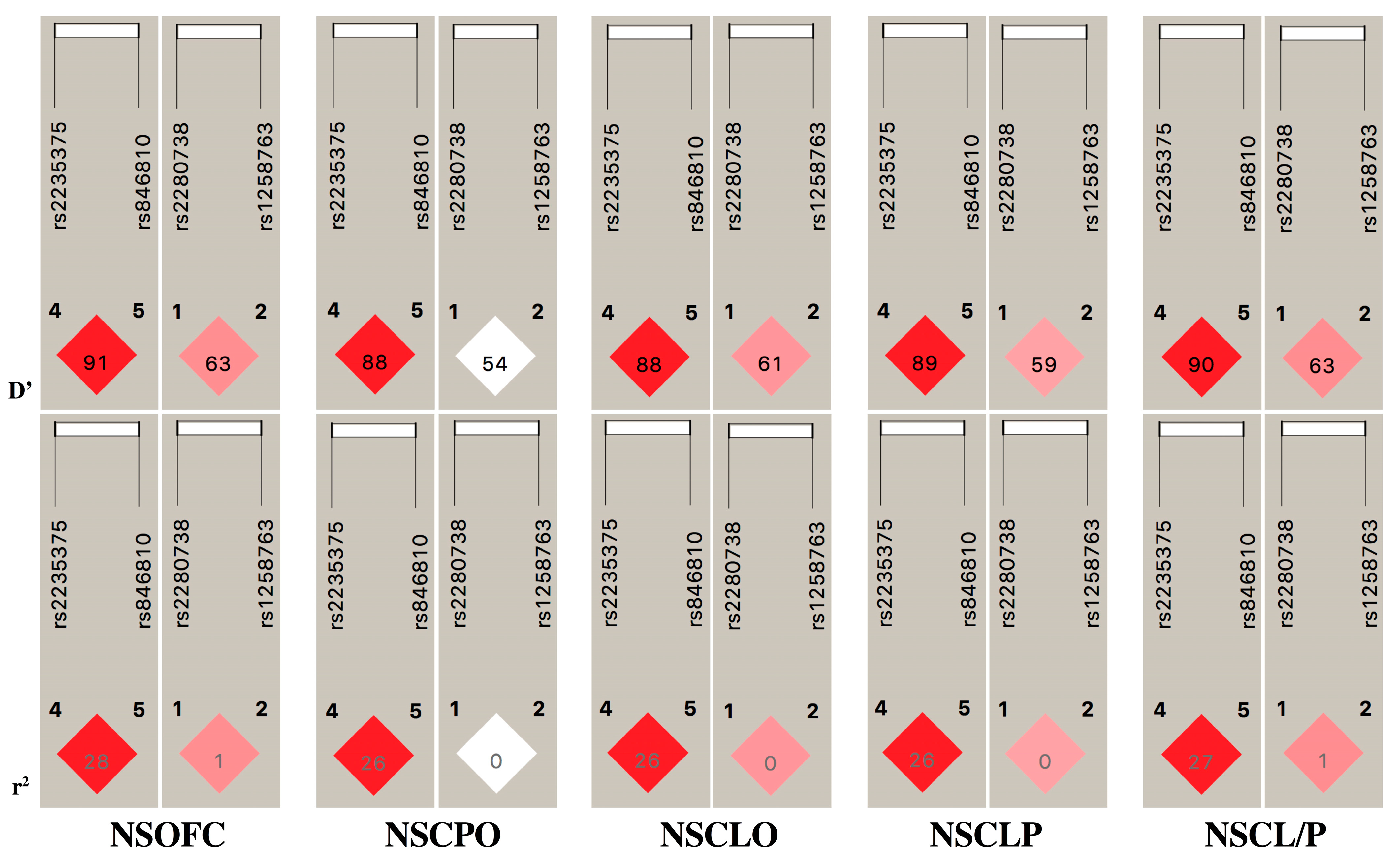
3.3. Case–Control Comparison and Haplotype Analysis
3.4. Family-Based Association Study and Haplotype Analysis
3.5. SNP × SNP Epistasis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dixon, M.J.; Marazita, M.L.; Beaty, T.H.; Murray, J.C. Cleft lip and palate: Understanding genetic and environmental influences. Nat. Rev. Genet. 2011, 12, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Natsume, N.; Kawai, T.; Le, H. In Vietnam, many congenital anomalies are believed to result from the scattering of defoliants, including dioxin. Cleft Palate-Craniofacial J. Off. Publ. Am. Cleft Palate-Craniofacial Assoc. 1998, 35, 183. [Google Scholar] [CrossRef]
- Jugessur, A.; Farlie, P.; Kilpatrick, N. The genetics of isolated orofacial clefts: From genotypes to subphenotypes. Oral Dis. 2009, 15, 437–453. [Google Scholar] [CrossRef] [PubMed]
- Leslie, E.J.; Marazita, M.L. Genetics of cleft lip and cleft palate. Am. J. Med. Genet. Part C Semin. Med. Genet. 2013, 163, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Mangold, E.; Ludwig, K.U.; Birnbaum, S.; Baluardo, C.; Ferrian, M.; Herms, S.; Reutter, H.; de Assis, N.A.; Chawa, T.A.; Mattheisen, M. Genome-wide association study identifies two susceptibility loci for nonsyndromic cleft lip with or without cleft palate. Nat. Genet. 2010, 42, 24–26. [Google Scholar] [CrossRef]
- Ludwig, K.U.; Mangold, E.; Herms, S.; Nowak, S.; Reutter, H.; Paul, A.; Becker, J.; Herberz, R.; AlChawa, T.; Nasser, E. Genome-wide meta-analyses of nonsyndromic cleft lip with or without cleft palate identify six new risk loci. Nat. Genet. 2012, 44, 968–971. [Google Scholar] [CrossRef]
- Beaty, T.; Taub, M.; Scott, A.; Murray, J.; Marazita, M.; Schwender, H.; Parker, M.; Hetmanski, J.; Balakrishnan, P.; Mansilla, M. Confirming genes influencing risk to cleft lip with/without cleft palate in a case–parent trio study. Hum. Genet. 2013, 132, 771–781. [Google Scholar] [CrossRef]
- Kondo, S.; Schutte, B.C.; Richardson, R.J.; Bjork, B.C.; Knight, A.S.; Watanabe, Y.; Howard, E.; Ferreira de Lima, R.L.; Daack-Hirsch, S.; Sander, A. Mutations in IRF6 cause Van der Woude and popliteal pterygium syndromes. Nat. Genet. 2002, 32, 285–289. [Google Scholar] [CrossRef]
- Zucchero, T.M.; Cooper, M.E.; Maher, B.S.; Daack-Hirsch, S.; Nepomuceno, B.; Ribeiro, L.; Caprau, D.; Christensen, K.; Suzuki, Y.; Machida, J. Interferon regulatory factor 6 (IRF6) gene variants and the risk of isolated cleft lip or palate. N. Engl. J. Med. 2004, 351, 769–780. [Google Scholar] [CrossRef]
- Beaty, T.H.; Murray, J.C.; Marazita, M.L.; Munger, R.G.; Ruczinski, I.; Hetmanski, J.B.; Liang, K.Y.; Wu, T.; Murray, T.; Fallin, M.D. A genome-wide association study of cleft lip with and without cleft palate identifies risk variants near MAFB and ABCA4. Nat. Genet. 2010, 42, 525–529. [Google Scholar] [CrossRef]
- Moretti, F.; Marinari, B.; Iacono, N.L.; Botti, E.; Giunta, A.; Spallone, G.; Garaffo, G.; Vernersson-Lindahl, E.; Merlo, G.; Mills, A.A. A regulatory feedback loop involving p63 and IRF6 links the pathogenesis of 2 genetically different human ectodermal dysplasias. J. Clin. Investig. 2010, 120, 1570–1577. [Google Scholar] [CrossRef]
- Yang, A.; Schweitzer, R.; Sun, D.; Kaghad, M.; Walker, N.; Bronson, R.T.; Tabin, C.; Sharpe, A.; Caput, D.; Crum, C. p63 is essential for regenerative proliferation in limb, craniofacial and epithelial development. Nature 1999, 398, 714–718. [Google Scholar] [CrossRef]
- Fakhouri, W.D.; Rahimov, F.; Attanasio, C.; Kouwenhoven, E.N.; Ferreira De Lima, R.L.; Felix, T.M.; Nitschke, L.; Huver, D.; Barrons, J.; Kousa, Y.A. An etiologic regulatory mutation in IRF6 with loss-and gain-of-function effects. Hum. Mol. Genet. 2014, 23, 2711–2720. [Google Scholar] [CrossRef] [PubMed]
- Wu-Chou, Y.-H.; Lu, Y.-C.; Chen, K.-T.P.; Chang, H.-F.; Lin, Y.-T.; Lo, L.-J. Association studies between regulatory regions of IRF6/TP63 genes and nonsyndromic oral clefts. Cleft Palate-Craniofacial J. 2019, 56, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Kurosaka, H.; Iulianella, A.; Williams, T.; Trainor, P.A. Disrupting hedgehog and WNT signaling interactions promotes cleft lip pathogenesis. J. Clin. Investig. 2014, 124, 1660–1671. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, E.; Li, B.; Zewdu, R.; Wells, V.; Hebert, J.M.; Karner, C.; Anderson, M.J.; Williams, T.; Dixon, J.; Dixon, M.J. A conserved Pbx-Wnt-p63-Irf6 regulatory module controls face morphogenesis by promoting epithelial apoptosis. Dev. Cell 2011, 21, 627–641. [Google Scholar] [CrossRef]
- Kimura-Yoshida, C.; Nakano, H.; Okamura, D.; Nakao, K.; Yonemura, S.; Belo, J.A.; Aizawa, S.; Matsui, Y.; Matsuo, I. Canonical Wnt signaling and its antagonist regulate anterior-posterior axis polarization by guiding cell migration in mouse visceral endoderm. Dev. Cell 2005, 9, 639–650. [Google Scholar] [CrossRef]
- Babai, A.; Irving, M. Orofacial Clefts: Genetics of Cleft Lip and Palate. Genes 2023, 14, 1603. [Google Scholar] [CrossRef]
- Jia, S.; Zhou, J.; D’Souza, R.N. Pax9′s dual roles in modulating Wnt signaling during murine palatogenesis. Dev. Dyn. 2020, 249, 1274–1284. [Google Scholar] [CrossRef]
- Nguyen, D.M.; Suzuki, S.; Imura, H.; Niimi, T.; Furukawa, H.; Ta, T.V.; Tong, S.M.; Nguyen, T.T.; Pham, L.N.G.; Tran, D.L. Family based and case–control designs reveal an association of TFAP2A in nonsyndromic cleft lip only among Vietnamese population. Mol. Genet. Genom. Med. 2021, 9, e1754. [Google Scholar] [CrossRef]
- Church, R.H.; Krishnakumar, A.; Urbanek, A.; Geschwindner, S.; Meneely, J.; Bianchi, A.; Basta, B.; Monaghan, S.; Elliot, C.; Strömstedt, M. Gremlin1 preferentially binds to bone morphogenetic protein-2 (BMP-2) and BMP-4 over BMP-7. Biochem. J. 2015, 466, 55–68. [Google Scholar] [CrossRef]
- Bénazet, J.-D.; Bischofberger, M.; Tiecke, E.; Gonçalves, A.; Martin, J.F.; Zuniga, A.; Naef, F.; Zeller, R. A self-regulatory system of interlinked signaling feedback loops controls mouse limb patterning. Science 2009, 323, 1050–1053. [Google Scholar] [CrossRef]
- Ornitz, D.M.; Itoh, N. Fibroblast growth factors. Genome Biol. 2001, 2, 3005. [Google Scholar] [CrossRef]
- Wang, C.; Chang, J.Y.F.; Yang, C.; Huang, Y.; Liu, J.; You, P.; McKeehan, W.L.; Wang, F.; Li, X. Type 1 fibroblast growth factor receptor in cranial neural crest cell-derived mesenchyme is required for palatogenesis. J. Biol. Chem. 2013, 288, 22174–22183. [Google Scholar] [CrossRef] [PubMed]
- Consortium, G.P.; Auton, A.; Brooks, L.; Durbin, R.; Garrison, E.; Kang, H. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Jugessur, A.; Rahimov, F.; Lie, R.T.; Wilcox, A.J.; Gjessing, H.K.; Nilsen, R.M.; Nguyen, T.T.; Murray, J.C. Genetic variants in IRF6 and the risk of facial clefts: Single-marker and haplotype-based analyses in a population-based case-control study of facial clefts in Norway. Genet. Epidemiol. Off. Publ. Int. Genet. Epidemiol. Soc. 2008, 32, 413–424. [Google Scholar] [CrossRef]
- Maili, L.; Letra, A.; Silva, R.; Buchanan, E.P.; Mulliken, J.B.; Greives, M.R.; Teichgraeber, J.F.; Blackwell, S.J.; Ummer, R.; Weber, R. PBX-WNT-P63-IRF6 pathway in nonsyndromic cleft lip and palate. Birth Defects Res. 2020, 112, 234–244. [Google Scholar] [CrossRef]
- Tam, V.; Patel, N.; Turcotte, M.; Bossé, Y.; Paré, G.; Meyre, D. Benefits and limitations of genome-wide association studies. Nat. Rev. Genet. 2019, 20, 467–484. [Google Scholar] [CrossRef]
- Johnson, J.L.; Abecasis, G.R. GAS Power Calculator: Web-based power calculator for genetic association studies. BioRxiv 2017, 164343. [Google Scholar] [CrossRef]
- Hong, E.P.; Park, J.W. Sample size and statistical power calculation in genetic association studies. Genom. Inform. 2012, 10, 117. [Google Scholar] [CrossRef]
- Suzuki, Y.; Jezewski, P.A.; Machida, J.; Watanabe, Y.; Shi, M.; Cooper, M.E.; Viet, L.T.; Tin, N.T.D.; Hai, H.; Natsume, N. In a Vietnamese population, MSX1 variants contribute to cleft lip and palate. Genet. Med. 2004, 6, 117–125. [Google Scholar] [CrossRef]
- Figueiredo, J.C.; Ly, S.; Raimondi, H.; Magee, K.; Baurley, J.W.; Sanchez-Lara, P.A.; Ihenacho, U.; Yao, C.; Edlund, C.K.; van den Berg, D. Genetic risk factors for orofacial clefts in Central Africans and Southeast Asians. Am. J. Med. Genet. Part A 2014, 164, 2572–2580. [Google Scholar] [CrossRef]
- Tran, D.L.; Imura, H.; Mori, A.; Suzuki, S.; Niimi, T.; Ono, M.; Sakuma, C.; Nakahara, S.; Nguyen, T.T.; Pham, P.T. Association of MEOX2 polymorphism with nonsyndromic cleft palate only in a Vietnamese population. Congenit. Anom. 2018, 58, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Ingraham, C.R.; Kinoshita, A.; Kondo, S.; Yang, B.; Sajan, S.; Trout, K.J.; Malik, M.I.; Dunnwald, M.; Goudy, S.L.; Lovett, M. Abnormal skin, limb and craniofacial morphogenesis in mice deficient for interferon regulatory factor 6 (Irf6). Nat. Genet. 2006, 38, 1335–1340. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; McIntosh, I.; Hetmanski, J.B.; Jabs, E.W.; Vander Kolk, C.A.; Wu-Chou, Y.-H.; Chen, P.K.; Chong, S.S.; Yeow, V.; Jee, S.H. Association between IRF6 and nonsyndromic cleft lip with or without cleft palate in four populations. Genet. Med. 2007, 9, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Cheng, L.; Badner, J.A.; Chen, C.; Chen, Q.; Luo, W.; Craig, D.W.; Redman, M.; Gershon, E.S.; Liu, C. Genetic control of individual differences in gene-specific methylation in human brain. Am. J. Hum. Genet. 2010, 86, 411–419. [Google Scholar] [CrossRef]
- Scapoli, L.; Palmieri, A.; Martinelli, M.; Pezzetti, F.; Carinci, P.; Tognon, M.; Carinci, F. Strong evidence of linkage disequilibrium between polymorphisms at the IRF6 locus and nonsyndromic cleft lip with or without cleft palate, in an Italian population. Am. J. Hum. Genet. 2005, 76, 180–183. [Google Scholar] [CrossRef]
- Huang, Y.; Wu, J.; Ma, J.; Beaty, T.; Sull, J.; Zhu, L.; Lu, D.; Wang, Y.; Meng, T.; Shi, B. Association between IRF6 SNPs and oral clefts in West China. J. Dent. Res. 2009, 88, 715–718. [Google Scholar] [CrossRef]
- Suazo, J.; Recabarren, A.S.; Marín, N.R.; Blanco, R. Association between IRF6 variants and nonsyndromic cleft lip with or without cleft palate in Chile. Reprod. Sci. 2020, 27, 1857–1862. [Google Scholar] [CrossRef]
- Velázquez-Aragón, J.A.; Angel, A.G.-d.; Alcántara-Ortigoza, M.A.; Reyna-Fabián, M.E.; Estandia-Ortega, B. Screening of IRF6 Variants in Patients Subjected to Genetic Association Studies for Nonsyndromic Cleft Lip/Palate. Cleft Palate-Craniofacial J. 2021, 58, 1128–1134. [Google Scholar] [CrossRef]
- Ibarra-Arce, A.; García-Álvarez, M.; Cortés-González, D.; de Zarate-Alarcón, G.O.; Flores-Peña, L.; Sánchez-Camacho, S.; Arenas-Díaz, S.; Romero-Valdovinos, M.; Olivo-Díaz, A. IRF6 polymorphisms in Mexican patients with non-syndromic cleft lip. Meta Gene 2015, 4, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Rinne, T.; Brunner, H.G.; van Bokhoven, H. p63-associated disorders. Cell Cycle 2007, 6, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Candi, E.; Rufini, A.; Terrinoni, A.; Dinsdale, D.; Ranalli, M.; Paradisi, A.; De Laurenzi, V.; Spagnoli, L.; Catani, M.; Ramadan, S. Differential roles of p63 isoforms in epidermal development: Selective genetic complementation in p63 null mice. Cell Death Differ. 2006, 13, 1037–1047. [Google Scholar] [CrossRef] [PubMed]
- Minocha, S.; Valloton, D.; Arsenijevic, Y.; Cardinaux, J.-R.; Guidi, R.; Hornung, J.-P.; Lebrand, C. Nkx2. 1 regulates the generation of telencephalic astrocytes during embryonic development. Sci. Rep. 2017, 7, 43093. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Yoon, S.; Lee, S.; Ha, S.A.; Kim, H.K.; Kim, J.W.; Chung, J. Gremlin-1 induces BMP-independent tumor cell proliferation, migration, and invasion. PLoS ONE 2012, 7, e35100. [Google Scholar] [CrossRef]
- Müller, I.I.; Chatterjee, M.; Schneider, M.; Borst, O.; Seizer, P.; Schönberger, T.; Vogel, S.; Müller, K.A.; Geisler, T.; Lang, F. Gremlin-1 inhibits macrophage migration inhibitory factor-dependent monocyte function and survival. Int. J. Cardiol. 2014, 176, 923–929. [Google Scholar] [CrossRef]
- Ludwig, K.U.; Ahmed, S.T.; Böhmer, A.C.; Sangani, N.B.; Varghese, S.; Klamt, J.; Schuenke, H.; Gültepe, P.; Hofmann, A.; Rubini, M. Meta-analysis reveals genome-wide significance at 15q13 for nonsyndromic clefting of both the lip and the palate, and functional analyses implicate GREM1 as a plausible causative gene. PLoS Genet. 2016, 12, e1005914. [Google Scholar] [CrossRef]
- Zuniga, A.; Michos, O.; Spitz, F.; Haramis, A.-P.G.; Panman, L.; Galli, A.; Vintersten, K.; Klasen, C.; Mansfield, W.; Kuc, S. Mouse limb deformity mutations disrupt a global control region within the large regulatory landscape required for Gremlin expression. Genes Dev. 2004, 18, 1553–1564. [Google Scholar] [CrossRef]
- Al Chawa, T.; Ludwig, K.U.; Fier, H.; Pötzsch, B.; Reich, R.H.; Schmidt, G.; Braumann, B.; Daratsianos, N.; Böhmer, A.C.; Schuencke, H. Nonsyndromic cleft lip with or without cleft palate: Increased burden of rare variants within Gremlin-1, a component of the bone morphogenetic protein 4 pathway. Birth Defects Res. Part A Clin. Mol. Teratol. 2014, 100, 493–498. [Google Scholar] [CrossRef]
- Mostowska, A.; Hozyasz, K.K.; Wójcicki, P.; Żukowski, K.; Dąbrowska, A.; Lasota, A.; Zadurska, M.; Radomska, A.; Dunin-Wilczyńska, I.; Jagodziński, P.P. Association between polymorphisms at the GREM1 locus and the risk of nonsyndromic cleft lip with or without cleft palate in the Polish population. Birth Defects Res. Part A Clin. Mol. Teratol. 2015, 103, 847–856. [Google Scholar] [CrossRef]
- Yin, B.; Shi, J.Y.; Lin, Y.S.; Shi, B.; Jia, Z.L. SNPs at TP63 gene was specifically associated with right-side cleft lip in Han Chinese population. Oral Dis. 2021, 27, 559–566. [Google Scholar] [CrossRef]
- Wang, X.; Song, H.; Jiao, X.; Hao, Y.; Zhang, W.; Gao, Y.; Li, Y.; Mi, N.; Yan, J. Association between a single-nucleotide polymorphism in the GREM1 gene and non-syndromic orofacial cleft in the Chinese population. J. Oral Pathol. Med. 2018, 47, 206–210. [Google Scholar] [CrossRef]
- Viena, C.S.; Machado, R.A.; Persuhn, D.C.; Martelli-Júnior, H.; Medrado, A.P.; Coletta, R.D.; Reis, S.R. Understanding the participation of GREM1 polymorphisms in nonsyndromic cleft lip with or without cleft palate in the Brazilian population. Birth Defects Res. 2019, 111, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Boehringer, S.; Van Der Lijn, F.; Liu, F.; Günther, M.; Sinigerova, S.; Nowak, S.; Ludwig, K.U.; Herberz, R.; Klein, S.; Hofman, A. Genetic determination of human facial morphology: Links between cleft-lips and normal variation. Eur. J. Hum. Genet. 2011, 19, 1192–1197. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.G. The role of haplotypes in candidate gene studies. Genet. Epidemiol. Off. Publ. Int. Genet. Epidemiol. Soc. 2004, 27, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.W.; Kaplan, N.L. On the advantage of haplotype analysis in the presence of multiple disease susceptibility alleles. Genet. Epidemiol. Off. Publ. Int. Genet. Epidemiol. Soc. 2002, 23, 221–233. [Google Scholar] [CrossRef]
- Cheng, X.; Du, F.; Long, X.; Huang, J. Genetic Inheritance Models of Non-Syndromic Cleft Lip with or without Palate: From Monogenic to Polygenic. Genes 2023, 14, 1859. [Google Scholar] [CrossRef]
- Bernheim, N.; Georges, M.; Malevez, C.; De Mey, A.; Mansbach, A. Embryology and epidemiology of cleft lip and palate. B ENT 2006, 2, 11–19. [Google Scholar]
- Leslie, E.J.; Carlson, J.C.; Shaffer, J.R.; Butali, A.; Buxó, C.J.; Castilla, E.E.; Christensen, K.; Deleyiannis, F.W.; Leigh Field, L.; Hecht, J.T. Genome-wide meta-analyses of nonsyndromic orofacial clefts identify novel associations between FOXE1 and all orofacial clefts, and TP63 and cleft lip with or without cleft palate. Hum. Genet. 2017, 136, 275–286. [Google Scholar] [CrossRef]
- Nasreddine, G.; El Hajj, J.; Ghassibe-Sabbagh, M. Orofacial clefts embryology, classification, epidemiology, and genetics. Mutat. Res./Rev. Mutat. Res. 2021, 787, 108373. [Google Scholar] [CrossRef]


{kind=link}
| Type of Cleft | N (%) | Sex | Cleft Side | ||
|---|---|---|---|---|---|
| Male | Female | Right Side | Left Side | ||
| NSOFC | 527 (100.0) | 313 (59.4) | 214 (40.6) | NA | NA |
| NSCPO | 101 (19.2) | 58 (57.4) | 43 (42.6) | NA | NA |
| NSCL/P | 426 (80.8) | 255 (58.9) | 171 (41.1) | 113 (26.5) | 313 (65.2) |
| NSCLO | 172 (32.6) | 100 (58.1) | 72 (41.9) | 46 (26.7) | 126 (73.3) |
| NSCLP | 254 (48.2) | 155 (61.0) | 99 (39.0) | 65 (25.6) | 189 (74.4) |
| NSUCL/P | 325 (76.3) | 184 (56.6) | 141 (43.4) | 113 (34.8) | 212 (65.2) |
| NSBCL/P | 101 (23.7) | 71 (70.3) | 30 (29.7) | NA | NA |
| Gene | SNP | Allele | HWEp | Minor Allele Frequency | |||||
|---|---|---|---|---|---|---|---|---|---|
| Control | NSOFC | NSCPO | NSCLO | NSCLP | NSCL/P | ||||
| IRF6 | rs2235375 | C>G | 0.7908 | 0.4374 | 0.4972 | 0.4010 | 0.5174 | 0.5217 | 0.5200 |
| IRF6 | rs846810 | A>G | 0.7906 | 0.2068 | 0.2571 | 0.2079 | 0.2762 | 0.2638 | 0.2688 |
| TP63 | rs9332461 | G>A | 0.2273 | 0.2362 | 0.2410 | 0.2426 | 0.2238 | 0.2520 | 0.2406 |
| GREM1 | rs2280738 | C>G | 0.1106 | 0.2315 | 0.2324 | 0.2129 | 0.2006 | 0.2618 | 0.2371 |
| GREM1 | rs1258763 | C>T | 0.1772 | 0.0721 | 0.0778 | 0.0693 | 0.0930 | 0.0709 | 0.0798 |
| Gene | Case (Control) | PGeno Value | ORallele (95% CI) Pallele Value | ORDom (95% CI) PDom Value | ORRec (95% CI) PRec Value | OROver (95% CI) POver Value | |
|---|---|---|---|---|---|---|---|
| NSOFC | |||||||
| IRF6 | |||||||
| rs2235375 | CC/CG/GG | 139/252/136 (165/263/99) | 0.0159 | 1.27 (1.07–1.51) 0.0060 | 0.79 (0.60–1.02) 0.0771 | 1.50 (1.12–2.02) 0.0062 | 0.92 (0.72–1.17) 0.4979 |
| rs846810 | AA/AG/GG | 298/187/42 (330/176/21) | 0.0113 | 1.33 (1.08–1.63) 0.0062 | 0.78 (0.60–0.99) 0.0446 | 2.09 (1.21–3.58) 0.0064 | 1.10 (0.85–1.41) 0.4758 |
| TP63 | |||||||
| rs9332461 | GG/AG/AA | 297/206/24 (302/201/24) | 0.9497 | 1.03 (0.84–1.25) 0.7983 | 0.96 (0.75–1.23) 0.7558 | 1.00 (0.56–1.78) 1 | 1.04 (0.81–1.33) 0.7518 |
| GREM1 | |||||||
| rs2280738 | CC/CG/GG | 309/191/27 (318/174/35) | 0.3766 | 1.01 (0.82–1.23) 0.9588 | 0.93 (0.73–1.19) 0.5723 | 0.76 (0.45–1.27) 0.295 | 1.15 (0.89–1.49) 0.2711 |
| rs1258763 | CC/CT/TT | 450/72/5 (456/66/5) | 0.8604 | 1.09 (0.78–1.50) 0.6191 | 0.90 (0.64–1.29) 0.5948 | 1.00 (0.29–3.47) 1 | 1.11 (0.77–1.58) 0.5838 |
| NSCPO | |||||||
| IRF6 | |||||||
| rs2235375 | CC/CG/GG | 38/45/18 (165/263/99) | 0.4533 | 0.86 (0.63–1.17) 0.3388 | 1.32 (0.85–2.06) 0.2139 | 0.94 (0.54–1.63) 0.8197 | 0.81 (0.53–1.24) 0.3244 |
| rs846810 | AA/AG/GG | 64/32/5 (330/176/21) | 0.8722 | 1.01 (0.69–1.46) 0.9721 | 1.03 (0.66–1.60) 0.8868 | 1.25 (0.46–3.41) 0.6554 | 0.92 (0.59–1.46) 0.7375 |
| TP63 | |||||||
| rs9332461 | GG/AG/AA | 56/41/4 (302/201/24) | NA | 1.04 (0.73–1.47) 0.8464 | NA | NA | 1.11 (0.72–1.71) 0.6425 |
| GREM1 | |||||||
| rs2280738 | CC/CG/GG | 62/35/4 (318/174/35) | NA | 0.90 (0.62–1.29) 0.5635 | NA | NA | 1.08 (0.69–1.68) 0.7492 |
| rs1258763 | CC/CT/TT | 88/12/1 (456/66/5) | NA | 0.96 (0.53–1.73) 0.8876 | NA | NA | 0.94 (0.49–1.81) 0.8577 |
| NSCLO | |||||||
| IRF6 | |||||||
| rs2235375 | CC/CG/GG | 40/86/46 (165/263/99) | 0.0329 | 1.38 (1.08–1.76) 0.0096 | 0.66 (0.45–0.99) 0.0440 | 1.58 (1.06–2.40) 0.0254 | 1.00 (0.71–1.41) 0.9828 |
| rs846810 | AA/AG/GG | 93/63/16 (330/176/21) | 0.0117 | 1.46 (1.11–1.93) 0.0074 | 0.70 (0.50–0.99) 0.0464 | 2.47 (1.26–4.85) 0.0068 | 1.15 (0.80–1.65) 0.4379 |
| TP63 | |||||||
| rs9332461 | GG/AG/AA | 101/65/6 (302/201/24) | 0.8219 | 0.93 (0.70–1.25) 0.6366 | 1.06 (0.75–1.50) 0.7443 | 0.76 (0.30–1.89) 0.5493 | 0.99 (0.69–1.40) 0.9346 |
| GREM1 | |||||||
| rs2280738 | CC/CG/GG | 109/57/6 (318/174/35) | 0.3022 | 0.83 (0.62–1.13) 0.2323 | 1.14 (0.80–1.62) 0.4791 | 0.51 (0.21–1.23) 0.1265 | 1.01 (0.70–1.45) 0.9763 |
| rs1258763 | CC/CT/TT | 142/28/2 (456/66/5) | NA | 1.32 (0.86–2.03) 0.2071 | NA | NA | 1.35 (0.84–2.19) 0.2100 |
| NSCLP | |||||||
| IRF6 | |||||||
| rs2235375 | CC/CG/GG | 61/121/72 (165/263/99) | 0.0053 | 1.40 (1.13–1.74) 0.0018 | 0.69 (0.49–0.98) 0.0352 | 1.71 (1.21–2.43) 0.0025 | 0.91 (0.68–1.23) 0.5527 |
| rs846810 | AA/AG/GG | 141/92/21 (330/176/21) | 0.0215 | 1.37 (1.07–1.76) 0.0116 | 0.75 (0.55–1.00) 0.0572 | 2.17 (1.16–4.06) 0.0129 | 1.13 (0.83–1.55) 0.4362 |
| TP63 | |||||||
| rs9332461 | GG/AG/AA | 140/100/14 (302/201/24) | 0.7652 | 1.09 (0.85–1.39) 0.4962 | 0.92 (0.68–1.24) 0.5634 | 1.22 (0.62–2.41) 0.5600 | 1.05 (0.77–1.43) 0.7408 |
| GREM1 | |||||||
| rs2280738 | CC/CG/GG | 138/99/17 (318/174/35) | 0.2463 | 1.18 (0.92–1.50) 0.1897 | 0.78 (0.58–1.06) 0.1104 | 1.01 (0.55–1.84) 0.9784 | 1.30 (0.95–1.77) 0.1018 |
| rs1258763 | CC/CT/TT | 220/32/2 (456/66/5) | NA | 0.98 (0.65–1.48) 0.9291 | NA | NA | 1.01 (0.64–1.58) 0.9764 |
| NSCL/P | |||||||
| IRF6 | |||||||
| rs2235375 | CC/CG/GG | 101/207/118 (165/263/99) | 0.0014 | 1.39 (1.16–1.67) 0.0003 | 0.68 (0.51–0.91) 0.0093 | 1.66 (1.22–2.25) 0.0011 | 0.95 (0.73–1.22) 0.6868 |
| rs846810 | AA/AG/GG | 234/155/37 (330/176/21) | 0.0031 | 1.41 (1.14–1.74) 0.0015 | 0.73 (0.56–0.94) 0.0164 | 2.29 (1.32–3.98) 0.0025 | 1.14 (0.87–1.49) 0.3354 |
| TP63 | |||||||
| rs9332461 | GG/AG/AA | 241/165/20 (302/201/24) | 0.9735 | 1.02 (0.83–1.27) 0.8239 | 0.97 (0.75–1.25) 0.8203 | 1.03 (0.56–1.90) 0.9180 | 1.03 (0.79–1.33) 0.8518 |
| GREM1 | |||||||
| rs2280738 | CC/CG/GG | 247/156/23 (318/174/35) | 0.427 | 1.03 (0.83–1.28) 0.7744 | 0.90 (0.70–1.18) 0.4609 | 0.80 (0.47–1.38) 0.4251 | 1.17 (0.90–1.53) 0.2452 |
| rs1258763 | CC/CT/TT | 362/60/4 (456/66/5) | NA | 1.12 (0.79–1.57) 0.5268 | NA | NA | 1.15 (0.79–1.67) 0.4794 |
| Gene/Haplotype | Haplotype Freq. | Case, Control Freq. | PCC Value | T/U | PTDT Value | |
|---|---|---|---|---|---|---|
| NSOFC | ||||||
| IRF6 | rs2235375–rs846810 | |||||
| C-A | 0.522 | 0.497, 0.548 | 0.0183 | 240.0/281.7 | 0.0679 | |
| G-A | 0.246 | 0.246, 0.245 | 0.9553 | 191.2/177.5 | 0.4753 | |
| G-G | 0.221 | 0.251, 0.192 | 0.0012 | 198.5/164.6 | 0.0747 | |
| GREM1 | rs2280738–rs1258763 | |||||
| C-C | 0.699 | 0.695, 0.703 | 0.6844 | 217.5/206.8 | 0.6026 | |
| G-C | 0.226 | 0.227, 0.224 | 0.8941 | 185.7/169.4 | 0.3855 | |
| C-T | 0.069 | 0.072, 0.065 | 0.5151 | 63.4/85.3 | 0.0725 | |
| NSCPO | ||||||
| IRF6 | rs2235375–rs846810 | |||||
| C-A | 0.555 | 0.596, 0.547 | 0.2017 | 59.2/42.9 | 0.1077 | |
| G-A | 0.238 | 0.196, 0.246 | 0.1277 | 29.2/31.4 | 0.7719 | |
| G-G | 0.193 | 0.205, 0.191 | 0.6580 | 31.0/45.0 | 0.1083 | |
| GREM1 | rs2280738–rs1258763 | |||||
| C-C | 0.707 | 0.724, 0.704 | 0.5566 | 37.9/34.3 | 0.6635 | |
| G-C | 0.221 | 0.206, 0.224 | 0.5777 | 32.4/27.1 | 0.4963 | |
| C-T | 0.064 | 0.063, 0.065 | 0.9189 | 10.2/18.0 | 0.1410 | |
| NSCLO | ||||||
| IRF6 | rs2235375–rs846810 | |||||
| C-A | 0.529 | 0.473, 0.547 | 0.0171 | 70.0/104.9 | 0.0082 | |
| G-A | 0.247 | 0.251, 0.246 | 0.8635 | 67.6/61.1 | 0.5671 | |
| G-G | 0.210 | 0.267, 0.191 | 0.0028 | 74.4/46.9 | 0.0125 | |
| GREM1 | rs2280738–rs1258763 | |||||
| C-C | 0.705 | 0.711, 0.704 | 0.7839 | 73.1/64.1 | 0.4433 | |
| G-C | 0.217 | 0.196, 0.224 | 0.2627 | 54.0/54.9 | 0.9327 | |
| C-T | 0.071 | 0.088, 0.065 | 0.1455 | 26.1/33.1 | 0.3642 | |
| NSCLP | ||||||
| IRF6 | rs2235375–rs846810 | |||||
| C-A | 0.523 | 0.471, 0.547 | 0.0049 | 110.8/133.9 | 0.1399 | |
| G-A | 0.252 | 0.265, 0.246 | 0.4189 | 94.4/84.9 | 0.4749 | |
| G-G | 0.213 | 0.257, 0.191 | 0.0031 | 93.1/72.6 | 0.1124 | |
| GREM1 | rs2280738–rs1258763 | |||||
| C-C | 0.694 | 0.674, 0.704 | 0.2278 | 106.5/108.4 | 0.8983 | |
| G-C | 0.234 | 0.256, 0.224 | 0.1718 | 99.4/87.4 | 0.3822 | |
| C-T | 0.065 | 0.065, 0.065 | 0.9826 | 27.1/34.2 | 0.3627 | |
| NSCL/P | ||||||
| IRF6 | rs2235375–rs846810 | |||||
| C-A | 0.514 | 0.472, 0.548 | 0.0011 | 180.8/238.8 | 0.0046 | |
| G-A | 0.251 | 0.259, 0.246 | 0.5112 | 162.0/146.0 | 0.3612 | |
| G-G | 0.223 | 0.261, 0.192 | 0.0003 | 167.5/119.5 | 0.0046 | |
| GREM1 | rs2280738–rs1258763 | |||||
| C-C | 0.697 | 0.689, 0.703 | 0.4861 | 179.6/172.4 | 0.7008 | |
| G-C | 0.228 | 0.231, 0.224 | 0.7154 | 153.3/142.4 | 0.5239 | |
| C-T | 0.069 | 0.074, 0.065 | 0.4333 | 53.2/67.4 | 0.1958 | |
| Gene | SNP | A1/A2 | TDT | POO | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Paternal | Maternal | PPOO | ||||||||
| T/U | OR (95% CI) | PTDT | T/U | PPat | T/U | PMat | ||||
| NSOFC | ||||||||||
| IRF6 | rs2235375 | G/C | 290/243 | 1.19 (1.01–1.42) | 0.0418 | 138.5/120.5 | 0.2634 | 151.5/122.5 | 0.0798 | 0.6738 |
| rs846810 | G/A | 201/173 | 1.16 (0.95–1.42) | 0.1477 | 94/89 | 0.7117 | 107/84 | 0.0961 | 0.3670 | |
| TP63 | rs9332461 | A/G | 197/197 | 1.00 (0.82–1.22) | 1.0000 | 100/98 | 0.8870 | 97/99 | 0.8864 | 0.8403 |
| GREM1 | rs2280738 | G/C | 185/174 | 1.06 (0.86–1.31) | 0.5615 | 95/87 | 0.5532 | 90/87 | 0.8216 | 0.7980 |
| rs1258763 | T/C | 66/93 | 0.71 (0.52–0.97) | 0.0323 | 33/56 | 0.0148 | 33/37 | 0.6326 | 0.2020 | |
| NSCPO | ||||||||||
| IRF6 | rs2235375 | G/C | 42/59 | 0.71 (0.48–1.06) | 0.0907 | 16/29 | 0.0526 | 26/30 | 0.5930 | 0.2717 |
| rs846810 | G/A | 31/45 | 0.69 (0.44–1.09) | 0.1083 | 12.5/24.5 | 0.0485 | 18.5/20.5 | 0.7488 | 0.2279 | |
| TP63 | rs9332461 | A/G | 39/39 | 1.00 (0.64–1.56) | 1.0000 | 17/21 | 0.5164 | 22/18 | 0.5271 | 0.3657 |
| GREM1 | rs2280738 | G/C | 30/26 | 1.15 (0.68–1.95) | 0.5930 | 14.5/12.5 | 0.7003 | 15.5/13.5 | 0.7103 | 0.9847 |
| rs1258763 | T/C | 11/20 | 0.55 (0.26–1.15) | 0.1060 | 6/11 | 0.2253 | 5/9 | 0.2850 | 0.9806 | |
| NSCLO | ||||||||||
| IRF6 | rs2235375 | G/C | 106/72 | 1.47 (1.09–1.99) | 0.0108 | 55.5/33.5 | 0.0197 | 50.5/38.5 | 0.2034 | 0.4454 |
| rs846810 | G/A | 76/48 | 1.58 (1.10–2.27) | 0.0119 | 40.5/22.5 | 0.0233 | 35.5/25.5 | 0.2004 | 0.4868 | |
| TP63 | rs9332461 | A/G | 63/59 | 1.07 (0.75–1.52) | 0.7172 | 34/31 | 0.7098 | 29/28 | 0.8946 | 0.8747 |
| GREM1 | rs2280738 | G/C | 54/56 | 0.96 (0.66–1.40) | 0.8488 | 30.5/30.5 | 1.0000 | 23.5/25.5 | 0.7751 | 0.8315 |
| rs1258763 | T/C | 26/34 | 0.76 (0.46–1.27) | 0.3017 | 12.5/21.5 | 0.1227 | 13.5/12.5 | 0.8445 | 0.2424 | |
| NSCLP | ||||||||||
| IRF6 | rs2235375 | G/C | 142/112 | 1.27 (0.99–1.62) | 0.0598 | 67/58 | 0.4208 | 75/54 | 0.0645 | 0.4665 |
| rs846810 | G/A | 94/80 | 1.18 (0.87–1.58) | 0.2885 | 41/42 | 0.9126 | 53/38 | 0.1159 | 0.2430 | |
| TP63 | rs9332461 | A/G | 95/99 | 0.96 (0.72–1.27) | 0.7740 | 49/46 | 0.7582 | 46/53 | 0.4817 | 0.4764 |
| GREM1 | rs2280738 | G/C | 101/92 | 1.10 (0.83–1.46) | 0.5171 | 50/44 | 0.5360 | 51/48 | 0.7630 | 0.8157 |
| rs1258763 | T/C | 29/39 | 0.74 (0.46–1.20) | 0.2253 | 14.5/23.5 | 0.1443 | 14.5/15.5 | 0.8551 | 0.4005 | |
| NSCL/P | ||||||||||
| IRF6 | rs2235375 | G/C | 248/184 | 1.34 (1.11–1.63) | 0.0020 | 122.5/91.5 | 0.0341 | 125.5/92.5 | 0.0254 | 0.9454 |
| rs846810 | G/A | 170/128 | 1.33 (1.06–1.67) | 0.0150 | 81.5/64.5 | 0.1594 | 88.5/63.5 | 0.0426 | 0.6755 | |
| TP63 | rs9332461 | A/G | 158/158 | 1.00 (0.80–1.25) | 1.0000 | 83/77 | 0.6353 | 75/81 | 0.6310 | 0.4997 |
| GREM1 | rs2280738 | G/C | 155/148 | 1.05 (0.84–1.31) | 0.6876 | 80.5/74.5 | 0.6299 | 74.5/73.5 | 0.9345 | 0.7809 |
| rs1258763 | T/C | 55/73 | 0.75 (0.53–1.07) | 0.1116 | 27/45 | 0.0339 | 28/28 | 1.0000 | 0.1576 | |
| Gene 1 | SNP1 | Gene 2 | SNP2 | PNSOFC | PNSCPO | PNSCLO | PNSCLP | PNSCL/P |
|---|---|---|---|---|---|---|---|---|
| IRF6 | rs2235375 | TP63 | rs9332461 | 0.1871 | 0.9353 | 0.3621 | 0.1295 | 0.1200 |
| rs846810 | rs9332461 | 0.6215 | 0.7205 | 0.6412 | 0.5306 | 0.4804 | ||
| rs2235375 | GREM1 | rs2280738 | 0.7350 | 0.3522 | 0.2438 | 0.4572 | 0.9533 | |
| rs2235375 | rs1258763 | 0.6445 | 0.6755 | 0.5768 | 0.9947 | 0.7338 | ||
| rs846810 | rs2280738 | 0.5479 | 0.3734 | 0.4334 | 0.3005 | 0.7436 | ||
| rs846810 | rs1258763 | 0.1696 | 0.0851 | 0.1996 | 0.5862 | 0.2867 | ||
| TP63 | rs9332461 | GREM1 | rs2280738 | 0.7330 | 0.4430 | 0.3672 | 0.6451 | 0.4836 |
| rs9332461 | rs1258763 | 0.7343 | 0.9310 | 0.6662 | 0.8212 | 0.7264 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pham, L.N.G.; Niimi, T.; Suzuki, S.; Nguyen, M.D.; Nguyen, L.C.H.; Nguyen, T.D.; Hoang, K.A.; Nguyen, D.M.; Sakuma, C.; Hayakawa, T.; et al. Association between IRF6, TP63, GREM1 Gene Polymorphisms and Non-Syndromic Orofacial Cleft Phenotypes in Vietnamese Population: A Case–Control and Family-Based Study. Genes 2023, 14, 1995. https://doi.org/10.3390/genes14111995
Pham LNG, Niimi T, Suzuki S, Nguyen MD, Nguyen LCH, Nguyen TD, Hoang KA, Nguyen DM, Sakuma C, Hayakawa T, et al. Association between IRF6, TP63, GREM1 Gene Polymorphisms and Non-Syndromic Orofacial Cleft Phenotypes in Vietnamese Population: A Case–Control and Family-Based Study. Genes. 2023; 14(11):1995. https://doi.org/10.3390/genes14111995
Chicago/Turabian StylePham, Loc Nguyen Gia, Teruyuki Niimi, Satoshi Suzuki, Minh Duc Nguyen, Linh Cao Hoai Nguyen, Tuan Duc Nguyen, Kien Ai Hoang, Duc Minh Nguyen, Chisato Sakuma, Toko Hayakawa, and et al. 2023. "Association between IRF6, TP63, GREM1 Gene Polymorphisms and Non-Syndromic Orofacial Cleft Phenotypes in Vietnamese Population: A Case–Control and Family-Based Study" Genes 14, no. 11: 1995. https://doi.org/10.3390/genes14111995