
Selection of Suitable Reference Genes for RT-qPCR Gene Expression Analysis in Centipedegrass under Different Abiotic Stress

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Material Culture and Stress Treatment
2.2. RNA Extraction and Reverse Transcription
2.3. Designing and Validating Primers with Specificity
2.4. Real-Time Quantitative PCR
2.5. Data Analysis and Stability Ranking
3. Results
3.1. Identification of the Primer Specificity
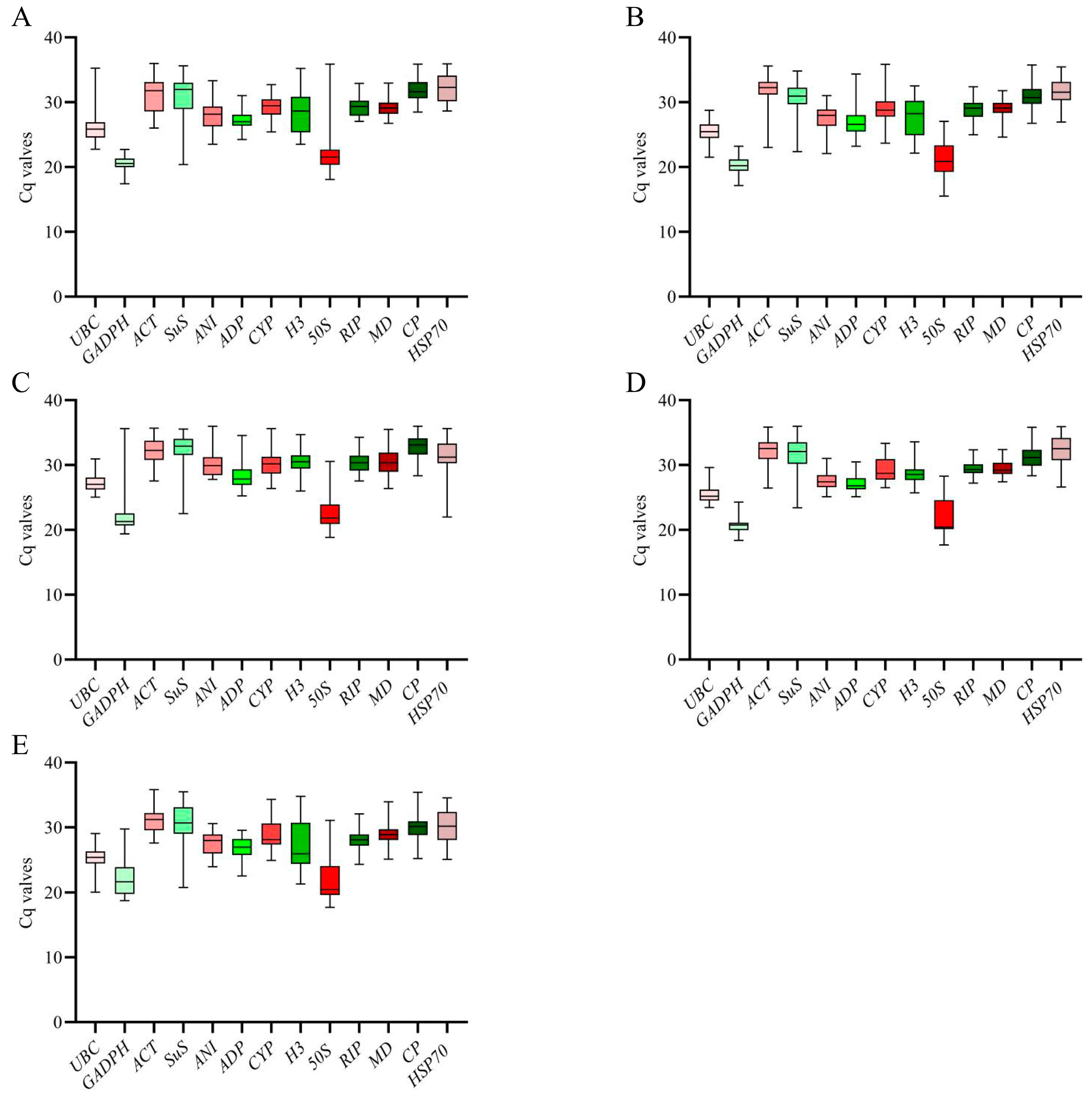
3.2. Analysis of Reference Gene Expression
3.3. Assessment of Expression Stability of Candidate Reference Genes
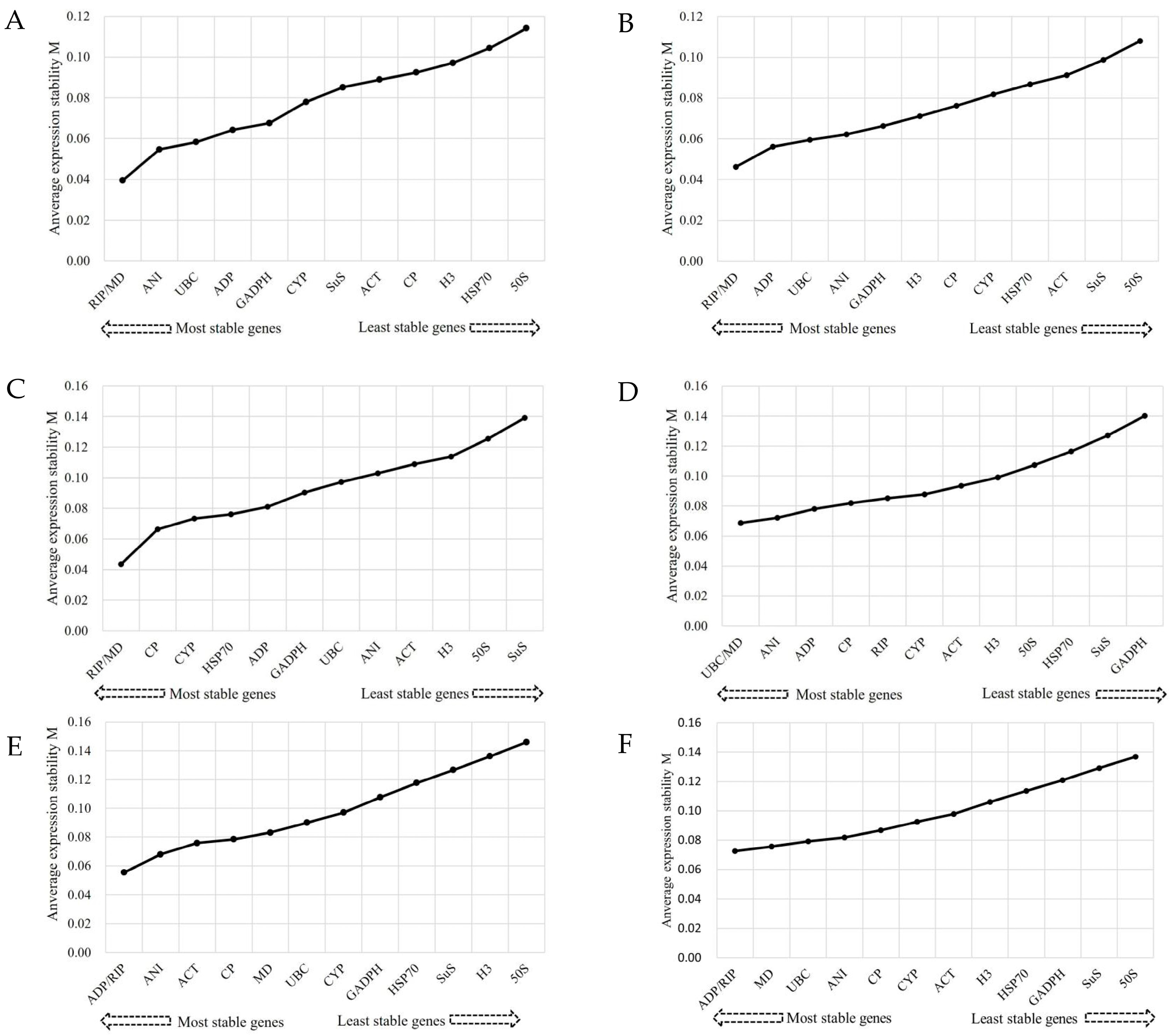
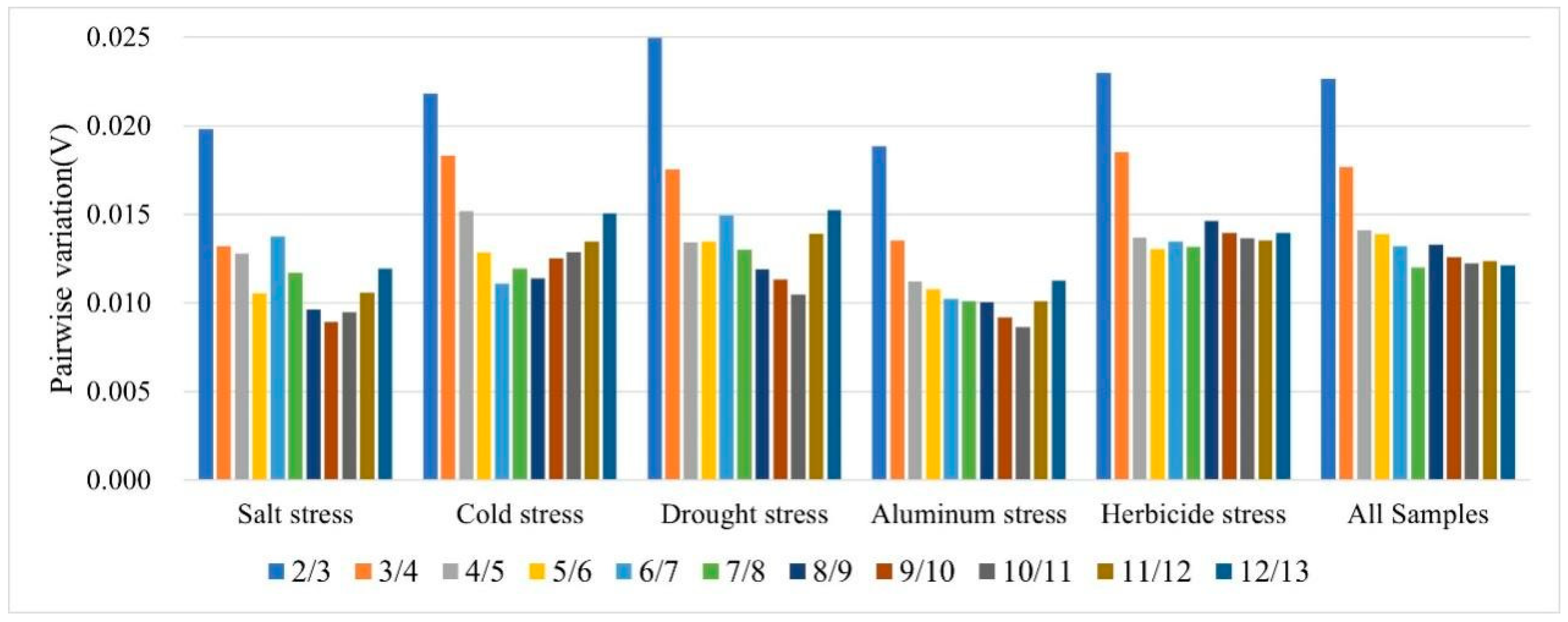
3.3.1. GeNorm Analysis
3.3.2. BestKeeper Analysis
3.3.3. NormFinder Analysis
3.3.4. ReFinder Analysis
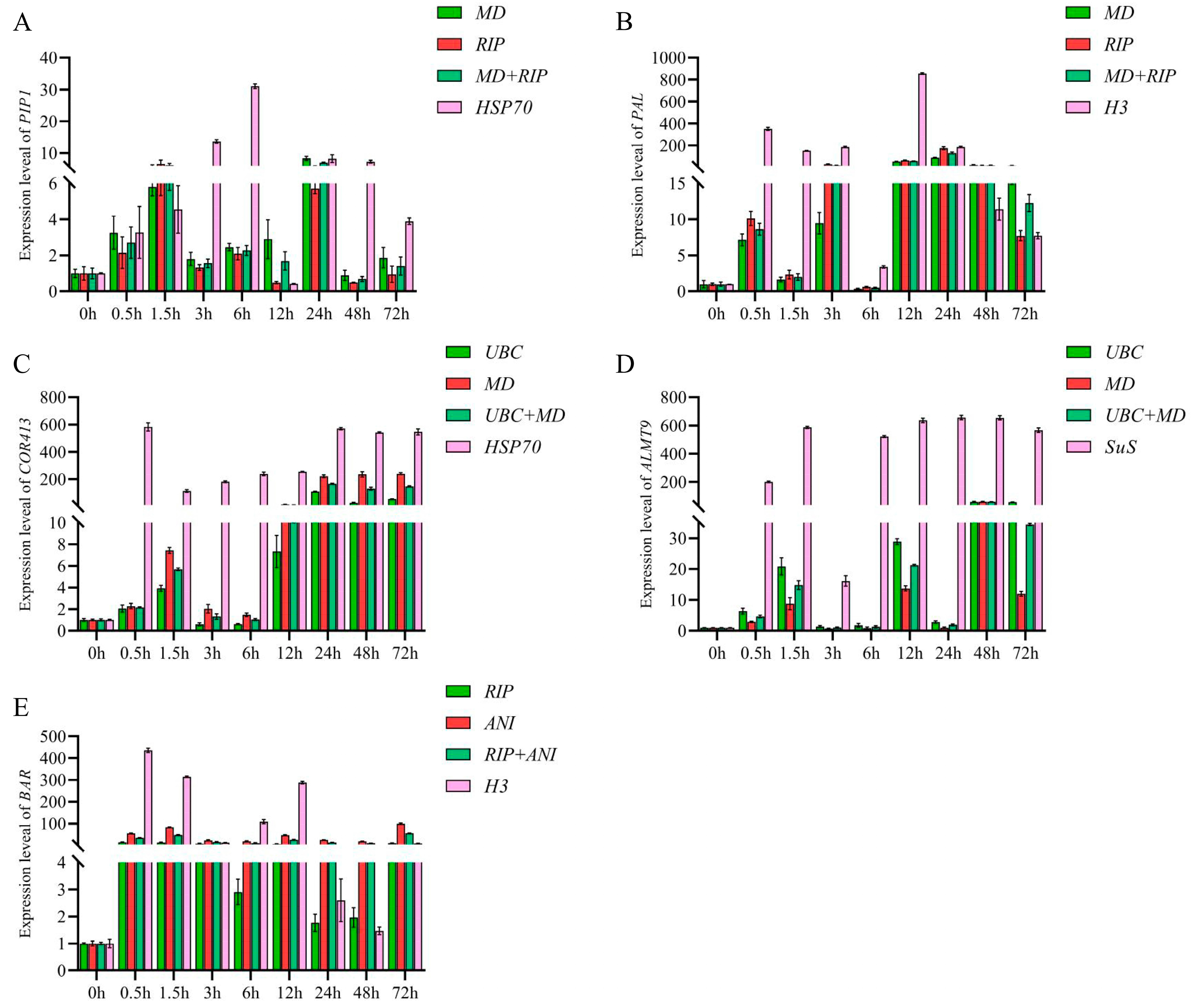
3.3.5. Verification of the Screened Reference Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palakolanu, S.R.; Dumbala, S.R.; Kaliamoorthy, S.; Pooja, B.M.; Vincent, V.; Sharma, K.K. Evaluation of Sorghum [Sorghum bicolor (L.)] Reference genes in various tissues and under abiotic stress conditions for quantitative real-time PCR data normalization. Front. Plant Sci. 2016, 7, 529. [Google Scholar] [CrossRef]
- Deng, L.T.; Wu, Y.L.; Li, J.C.; Ouyang, K.X.; Ding, M.M.; Zhang, J.J.; Li, S.Q.; Lin, M.F.; Chen, H.B.; Hu, X.S.; et al. Screening reliable reference genes for RT-qPCR analysis of gene expression in Moringa oleifera. PLoS ONE 2016, 11, e0159458. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.Y.; Zhang, R.M.; Hu, X.L.; Li, J.; Zhao, P.L.; Guo, F.; Zhao, H.; Wang, P.; Wang, Y.; Ni, D.J.; et al. Reference gene selection for qRT-PCR analysis in the shoots and roots of Camellia sinensis var. sinensis under nutritional stresses. Sci. Hortic. 2023, 320, 112237. [Google Scholar] [CrossRef]
- Lin, Y.; Liu, G.F.; Rao, Y.; Wang, B.; Tian, R.F.; Tan, Y.Y.; Peng, T. Identification and validation of reference genes for qRT-PCR analyses under different experimental conditions in Allium wallichii. J. Plant Physiol. 2023, 281, 153925. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Li, B.; Chang, Y.; Hou, X.; Dai, S. Selection and validation of reference genes for RT-qPCR analysis in Spinacia oleracea under abiotic stress. Biomed Res. Int. 2021, 2021, 4853632. [Google Scholar] [CrossRef]
- Silva, F.; Santos, T.; Figueiredo, M.; Cacefo, V.; Vieira, L.; Ribas, A.F. Validation of reference genes for real-time quantitative PCR in brachiaria grass under salt stress. Plant Gene 2021, 27, 100319. [Google Scholar] [CrossRef]
- Noelia, U.M.; Esteban, M.; Alejandra, C.; Esteban, H.; Daniela, T. Validation of reference genes for quantitative PCR in Johnsongrass (Sorghum halepense L.) under glyphosate stress. Plants 2021, 10, 1555. [Google Scholar] [CrossRef]
- Jain, M.; Nijhawan, A.; Tyagi, A.K.; Khurana, J.P. Validation of housekeeping genes as internal control for studying gene expression in rice by quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2006, 345, 646–651. [Google Scholar] [CrossRef]
- Wang, H.B.; Wang, J.J.; Jiang, J.F.; Chen, S.M.; Guan, Z.Y.; Liao, Y.; Chen, F.D. Reference genes for normalizing transcription in diploid and tetraploid Arabidopsis. Sci. Rep. 2014, 4, 6781. [Google Scholar] [CrossRef]
- Pabuayon, I.M.; Yamamoto, N.; Trinidad, J.L.; Longkumer, T.; Raorane, M.L.; Kohli, A. Reference genes for accurate gene expression analyses across different tissues, developmental stages and genotypes in rice for drought tolerance. Rice 2016, 9, 32. [Google Scholar] [CrossRef]
- Nguyen, D.Q.; Eamens, A.L.; Grof, C.P.L. Reference gene identification for reliable normalisation of quantitative RT-PCR data in Setaria viridis. Plant Methods 2018, 14, 24. [Google Scholar] [CrossRef] [PubMed]
- Niu, K.; Shi, Y.; Ma, H. Selection of candidate reference genes for gene expression analysis in kentucky bluegrass (Poa pratensis L.) under abiotic stress. Front. Plant Sci. 2017, 8, 193. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.Y.; Fu, J.Y.; Li, X.; Peng, L.L.; Yang, L.Q.; Liang, Y.H.; Jiang, M.Y.; Ma, J.; Sun, L.X.; Guo, B.M.; et al. Low-molecular-weight organic acid-mediated tolerance and Pb accumulation in centipedegrass under Pb stress. Ecotoxicol. Environ. Safe 2022, 241, 113755. [Google Scholar] [CrossRef]
- He, Q.Q.; Liu, C.Q.; Li, J.J.; Wang, J.J.; Yao, X.; Zhou, S.J.; Chen, Y.; Wang, H.R. Cloning of EoNLA gene in Eremochloa ophiuroides and the transgenic Arabidopsis phenotypic characterization under various phosphorus levels. J. Nanjing For. Univ. 2022, 46, 134–142. [Google Scholar]
- Li, J.J.; Guo, H.L.; Zong, J.Q.; Chen, J.B.; Liu, J.X. Genetic diversity in centipedegrass [Eremochloa ophiuroides (Munro) Hack.]. Hortic. Res. 2020, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.H.; Bai, S.Q.; Guan, Z.Y.; Chen, X.R.; Xu, G.G. Characteristics and applications of Eremochloa ophiuroides in China. North. Hortic. 2017, 41, 80–86. [Google Scholar]
- Liu, Y.J.; Xiong, Y.; Zhao, J.M.; Bai, S.Q.; Li, D.X.; Chen, L.M.; Fen, J.J.; Li, Y.Z.; Ma, X.; Zhang, J.B. Molecular mechanism of cold tolerance of centipedegrass based on the transcriptome. Int. J. Mol. Sci. 2023, 24, 1265. [Google Scholar] [CrossRef]
- Xu, X.; Wang, H.R.; Zhang, B.; Zhang, L.; Li, X.H.; Hao, D.L.; Chen, J.B.; Zong, J.Q.; Guo, H.L.; Liu, J.X.; et al. Micromorphological and physiological responses of two contrasting centipedegrass (Eremochloa ophiuroides(Munro) Hack.) genotypes in response to aluminum toxicity. J. Soil Sci. Plant Nut. 2023, 23, 2174–2189. [Google Scholar] [CrossRef]
- Islam, M.A.; Hirata, M. Centipedegrass (Eremochloa ophiuroides (Munro) Hack.): Growth behavior and multipurpose usages. Grassl. Sci. 2010, 51, 183–190. [Google Scholar] [CrossRef]
- Liu, Y.M.; Huan, H.F.; Ding, X.P.; Chen, Z.J.; Li, X.Y.; Liu, P.D.; Feng, Z.; Wang, Z.Y.; Bai, C.J.; Liu, G.D. Evaluation of salinity tolerance of 55 centipedegrass ecotypes. Ratacultural Sci. 2017, 34, 2261–2271. [Google Scholar]
- Ian, H. Global perspective of herbicide-resistant weeds. Pest Manag. Sci. 2014, 70, 1306–1315. [Google Scholar] [CrossRef]
- Tang, X.; Zhang, N.; Si, X.J.; Calderón-Urrea, A. Selection and validation of reference genes for RT-qPCR analysis in potato under abiotic stress. Plant Methods 2017, 13, 85. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tan, Z.Q.; Hu, B.Y.; Yang, Z.M.; Xu, B.; Zhuang, L.L.; Huang, B.R. Selection and validation of reference genes for target gene analysis with quantitative RT-PCR in leaves and roots of bermudagrass under four different abiotic stresses. Physiol. Plant. 2015, 155, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.; Chen, J.H.; Tian, Q.Q.; Wang, S.; Xia, X.L.; Yin, W.L. Identification and validation of reference genes for Populus euphratica gene expression analysis during abiotic stresses by quantitative real-time PCR. Physiol. Plant. 2014, 152, 529–545. [Google Scholar] [CrossRef] [PubMed]
- Yvette, M.S.; Eveline, Z.V.; Bas, B.; Victor, P.M.G.R.; Bart, S.; Louis, C.P. A GeNorm algorithm-based selection of reference genes for quantitative real-time PCR in skin biopsies of healthy dogs and dogs with atopic dermatitis. Vet. Immunol. Immunopathol. 2009, 129, 115–118. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, Z.C.; Xu, J.; Ji, A.J.; Luo, H.M.; Song, J.Y.; Sun, C.; Hu, Y.L.; Chen, S.L. Selection and validation of reference genes for normalization of quantitative real-time reverse transcription PCR analysis in Poria cocos (Schw.) Wolf (Fuling). Chin. Med. 2016, 11, 8. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper-Excel-Based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Xie, F.L.; Xiao, P.; Chen, D.L.; Xu, L.; Zhang, B.H. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Gimeno, J.; Eattock, N.; Van, D.A.; Blumwald, E. Selection and validation of reference genes for gene expression analysis in switchgrass (Panicum virgatum) using quantitative real-time RT-PCR. PLoS ONE 2014, 9, e91474. [Google Scholar] [CrossRef]
- Nguyen, D.Q.; Nguyen, N.L.; Nguyen, V.T.; Nguyen, T.H.G.; Nguyen, T.H.; Nguyen, T.K.L.; Nguyen, H.H. Reliable reference genes for accurate gene expression profiling across different tissues and genotypes of rice seedlings (Oryza sativa L.) under salt stress. Russ. J. Plant Physl. 2023, 70, 104. [Google Scholar] [CrossRef]
- Li, G.Y.; Sun, X.H.; Zhu, X.P.; Wu, B.; Hong, H.; Xin, Z.M.; Xin, X.Q.; Peng, J.J.; Jiang, S.S. Selection and validation of reference genes in virus-infected sweet potato Plants. Genes 2023, 14, 1477. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De, P.K.; Pattyn, F.; Poppe, B.; Van, R.N.; De, P.A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 341–3411. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Li, H.; Lu, L.; Ji, Y.L.; Ma, L.L.; Li, S.C. Screening and validation of internal reference genes for quantitative real-time PCR analysis of leaf color mutants in dendrobium officinale. Genes 2023, 14, 1112. [Google Scholar] [CrossRef]
- Ward, D.S.; Jutta, D.W.; Roswitha, W.; Valérie, S.; Hubert, S.; Daniel, N.; Martin, B.; Ralph, B.; Sabine, K.; Linos, V.; et al. Reference gene validation for RT-qPCR, a note on different available software packages. PLoS ONE 2015, 10, e0122515. [Google Scholar] [CrossRef]
- Soltani, N.M.; Amirbakhtiar, N. Evaluating and validating sunflower reference genes for Q-PCR studies under high temperature condition. Iran. J. Biotechnol. 2023, 21, e3357. [Google Scholar] [CrossRef]
- Xie, F.L.; Wang, J.Y.; Zhang, B.H. RefFinder: A web-based tool for comprehensively analyzing and identifying reference genes. Funct. Integr. Genom. 2023, 23, 125. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.F.; Lei, J.; Luo, J.J.; Liu, P.D.; Yu, D.G.; Luo, L.J. Response of phenylpropane ammonia-lyase on biotic and abiotic stress in Stylosanthes. Chin. J. Trop. Crops 2019, 40, 1742–1751. [Google Scholar] [CrossRef]
- Zhang, M.; Shi, H.T.; Li, N.N.; Wei, N.N.; Tian, Y.; Peng, J.F.; Chen, X.C.; Zhang, L.Y.; Zhang, M.X.; Dong, H.S. Aquaporin OsPIP2;2 links the H2O2 signal and a membrane-anchored transcription factor to promote plant defense. Plant Physiol. 2021, 188, 2325–2341. [Google Scholar] [CrossRef]
- Repkina, N.; Ignatenko, A.; Holoptseva, E.; Miszalski, Z.; Kaszycki, P.; Talanova, V. Exogenous Methyl Jasmonate improves cold tolerance with parallel induction of two Cold-Regulated (COR) genes expression in Triticum aestivum L. Plants 2021, 10, 1421. [Google Scholar] [CrossRef]
- Sl-ALMT9 regulates malate accumulation and aluminum tolerance in tomato. Sci. Found. China 2018, 26.
- Ijaz, S.; Vivian, M.; Maqbool, A.; Irfan, M.; Mehnaz, S.; Malik, K.A.; Bashir, A. Stacking of hordeum vulgare vacuolar sodium/proton antiporter and a bar gene in wheat for salt and herbicide tolerance. J. Crop Sci. Biotechnol. 2022, 25, 451–464. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Xia, H.; Liu, X.L.; Lin, Z.Y.; Guo, Y.Q.; Deng, H.H.; Wang, J.; Lin, L.J.; Deng, Q.X.; Lv, X.L.; et al. Identification of suitable reference genes for qRT-PCR normalization in kiwi fruit. Horticulturae 2022, 8, 170. [Google Scholar] [CrossRef]
- Lin, S.K.; Xu, S.C.; Huang, L.Y.; Qiu, F.X.; Zheng, Y.H.; Liu, Q.H.; Ma, S.W.; Wu, B.S.; Wu, J.C. Selection and validation of reference genes for normalization of RT-qPCR analysis in developing or abiotic-stressed tissues of loquat (Eriobotrya japonica). Phyton-Int. J. Exp. Bot. 2023, 92, 1185–1201. [Google Scholar] [CrossRef]
- Feng, K.; Liu, J.X.; Xing, G.M.; Sun, S.; Li, S.; Duan, A.Q.; Wang, F.; Li, M.Y.; Xu, Z.S.; Xiong, A.S. Selection of appropriate reference genes for RT-qPCR analysis under abiotic stress and hormone treatment in celery. PeerJ 2019, 7, e7925. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.; Reddy, R.A.; Ponnuvel, K.M.; Rohela, G.K.; Shabnam, A.A.; Ghosh, M.K.; Mishra, R.K. Selection of suitable reference genes for quantitative real-time PCR gene expression analysis in mulberry (Morus alba L.) under different abiotic stresses. Mol. Biol. Rep. 2019, 46, 1809–1817. [Google Scholar] [CrossRef]
- Deng, Y.; Li, Y.; Sun, H. Selection of reference genes for RT-qPCR normalization in blueberry (Vaccinium corymbosum × angustifolium) under various abiotic stresses. FEBS Open Bio 2020, 10, 1418–1435. [Google Scholar] [CrossRef]
- Tajti, J.; Pál, M.; Janda, T. Validation of reference genes for studying different abiotic stresses in Oat (Avena sativa L.) by RT-qPCR. Plants 2021, 10, 1272. [Google Scholar] [CrossRef]
- Ma, L.; Wu, J.; Qi, W.; Coulter, J.A.; Sun, W. Screening and verification of reference genes for analysis of gene expression in winter rapeseed (Brassica rapa L.) under abiotic stress. PLoS ONE 2020, 15, e236577. [Google Scholar] [CrossRef]
- Kozera, B.; Rapacz, M. Reference genes in real-time PCR. J. Appl. Genet. 2013, 54, 391–406. [Google Scholar] [CrossRef]
- Joseph, J.T.P.; Najya, J.S.; Jasmine, M. Screening internal controls for expression analyses involving numerous treatments by combining statistical methods with reference gene selection tools. Physiol. Mol. Biol. Plants 2018, 25, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.M.; Zhou, M.; Yu, M. Identification and validation of reference genes for RT-qPCR analysis in switchgrass under heavy metal stresses. Genes 2020, 11, 502. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Huang, L.L.; Wang, Y.T.; Li, X.; Feng, X.X.; Li, L.L. Stepwise optimization of the RT-qPCR protocol and the evaluation of housekeeping genes in pears (Pyrus bretschneideri) under various hormone treatments and stresses. Horticulturae 2023, 9, 275. [Google Scholar] [CrossRef]
- Zhang, J.C.; Xie, W.G.; Yu, X.X.; Zhang, Z.Y.; Zhao, Y.Q.; Wang, N.; Wang, Y.R. Selection of suitable reference genes for RT-qPCR gene expression analysis in Siberian wild rye (Elymus sibiricus) under different experimental conditions. Genes 2019, 10, 451. [Google Scholar] [CrossRef]
- Zhang, Y.T.; Zhu, L.J.; Xue, J.Y.; Yang, J.J.; Hu, H.L.; Cui, J.B.; Xu, J. Selection and verification of appropriate reference genes for expression normalization in cryptomeria fortunei under abiotic stress and hormone treatments. Genes 2021, 12, 791. [Google Scholar] [CrossRef]
- Wang, M.; Ren, T.T.; Marowa, P.; Du, H.N.; Xu, Z.C. Identification and selection of reference genes for gene expression analysis by quantitative real-time PCR in Suaeda glauca’s response to salinity. Sci. Rep. 2021, 11, 8569. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.D.; Huang, X.L.; Cong, H.Q.; Qiao, F.; Cheng, Y.J.; Chen, Y.Y. Selection and identification of a reference gene for normalizing real-time PCR in mangos under various stimuli in different tissues. Horticulturae 2022, 8, 882. [Google Scholar] [CrossRef]
- Guo, M.; Zhai, Y.F.; Lu, J.P.; Chai, L.; Chai, W.G.; Gong, Z.H.; Lu, M.H. Characterization of aHsp70-1, a pepper heat-shock protein gene in response to heat stress and some regulation exogenous substances in Capsicum annuum L. Int. J. Mol. Sci. 2014, 15, 19741–19759. [Google Scholar] [CrossRef]
- Benazir, S.; Rochon, D.A. Cucumber necrosis virus recruits cellular heat shock protein 70 homologs at several stages of infection. J. Virol. 2016, 90, 3302–3317. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.P.; Qu, Y.Y.; Fan, R.; Sun, F.L.; Chen, Q.; Shi, J.B.; Zheng, K.; Ni, Z.Y.; Zhang, Y.B.; Chen, Q.J.; et al. Regulating drought tolerance in cotton by the expression of a specific allele of heat shock protein 70. Ind. Crops Prod. 2023, 202, 116820. [Google Scholar] [CrossRef]
- Usman, M.G.; Rafii, M.Y.; Martini, M.Y.; Yusuff, O.A.; Ismail, M.R.; Miah, G. Molecular analysis of Hsp70 mechanisms in plants and their function in response to stress. Biotechnol. Genet. Eng. 2017, 33, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Shibasaka, M.; Horie, T.; Katsuhara, M. Mechanisms activating latent functions of PIP aquaporin water channels via the interaction between PIP1 and PIP2 proteins. Plant Cell Physiol. 2020, 62, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.G.; Chung, O.C.; Bae, T.W.; Sun, H.J.; Song, I.J.; Park, K.W.; Lim, P.O.; Lee, J.; Lee, Y.E.; Song, P.S. Pollen-mediated flow of bar gene in transgenic herbicide-resistant turf grass Zoysia japonica. Plant Biotechnol. Rep. 2021, 15, 241–250. [Google Scholar] [CrossRef]
- Chen, S.; Chen, K.; Ding, Q.Q.; Mou, Y.Y.; Yang, R.; Zhao, M.J.; Ma, B.; Xu, Z.S.; Ma, Y.Z.; Pan, Y.H.; et al. Proteomic Analysis of the function of a novel cold-regulated multispanning transmembrane protein COR413-PM1 in Arabidopsis. Int. J. Mol. Sci 2018, 19, 2572. [Google Scholar] [CrossRef] [PubMed]
- Palaniyandi, K.; Yan, X.X.; Liao, W.; Wu, J.; Chen, F.; Tang, L. Identification and validation of superior reference gene for gene expression normalization via RT-qPCR in staminate and pistillate flowers of Jatropha curcas—A biodiesel plant. PLoS ONE 2017, 12, e0177039. [Google Scholar] [CrossRef]
- Gao, M.M.; Liu, Y.P.; Ma, X.; Shuai, Q.; Gai, J.Y.; Li, Y. Evaluation of reference genes for normalization of gene expression using quantitative RT-PCR under aluminum, cadmium, and heat stresses in soybean. PLoS ONE 2017, 12, e168965. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank | Cold Stress | Salt Stress | Drought Stress | Al Stress | Herbicide Stress | All Samples |
|---|---|---|---|---|---|---|
| 1 | UBC(4.24 ± 1.15) | MD(3.63 ± 1.05) | GADPH(5.08 ± 1.04) | RIP(3.1 ± 0.91) | RIP(3.83 ± 1.07) | MD(4.26 ± 1.25) |
| 2 | RIP(4.3 ± 1.31) | GADPH(5.68 ± 1.15) | MD(3.66 ± 1.07) | GADPH(4.65 ± 0.97) | ADP(4.79 ± 1.29) | ADP(4.79 ± 1.3) |
| 3 | CP(4.09 ± 1.35) | ADP(4.99 ± 1.32) | ADP(4.38 ± 1.19) | MD(3.36 ± 0.99) | MD(4.62 ± 1.34) | RIP(4.52 ± 1.32) |
| 4 | H3(4.72 ± 1.44) | UBC(5.33 ± 1.35) | RIP(4.25 ± 1.25) | UBC(4.09 ± 1.05) | UBC(5.39 ± 1.35) | UBC(5.37 ± 1.39) |
| 5 | ADP(5.12 ± 1.45) | RIP(4.7 ± 1.07) | CYP(5.02 ± 1.46) | ADP(3.93 ± 1.07) | ACT(4.68 ± 1.45) | GADPH(7.4 ± 1.57) |
| 6 | MD(4.95 ± 1.51) | CP(4.39 ± 1.36) | CP(4.79 ± 1.52) | ANI(4.08 ± 1.12) | ANI(5.53 ± 1.52) | CP(5.34 ± 1.67) |
| 7 | ACT(4.74 ± 1.52) | HSP70(5.32 ± 1.68) | UBC(5.9 ± 1.54) | H3(4.32 ± 1.24) | CP(5.52 ± 1.64) | ANI(6.06 ± 1.71) |
| 8 | CYP(5.18 ± 1.57) | CYP(6.04 ± 1.74) | HSP70(8.36 ± 2.62) | CP(4.51 ± 1.41) | GADPH(8.89 ± 1.97) | CYP(5.9 ± 1.73) |
| 9 | ANI(5.8 ± 1.75) | ANI(6.38 ± 1.74) | 50S(8.22 ± 1.79) | CYP(5.39 ± 1.85) | CYP(6.85 ± 1.97) | ACT(5.91 ± 1.87) |
| 10 | HSP70(7.52 ± 2.32) | SuS(6.66 ± 2.03) | ANI(6.88 ± 1.94) | ACT(5.65 ± 1.81) | HSP70(7.36 ± 2.21) | HSP70(6.31 ± 1.99) |
| 11 | 50S(9.44 ± 2.14) | ACT(6.91 ± 2.17) | ACT(6.9 ± 2.16) | HSP70(5.93 ± 1.91) | 50S(11.55 ± 2.51) | 50S(10.29 ± 2.24) |
| 12 | GADPH(10.35 ± 2.34) | 50S(10.95 ± 2.31) | H3(9.05 ± 2.6) | SuS(7.42 ± 2.34) | SuS(8.47 ± 2.58) | H3(8.31 ± 2.38) |
| 13 | SuS(7.75 ± 2.47) | H3(8.85 ± 2.46) | SuS(9.95 ± 3.04) | 50S(11.01 ± 2.4) | H3(11.59 ± 3.14) | SuS(8.35 ± 2.59) |
| Rank | Cold Stress | Salt Stress | Drought Stress | Al Stress | Herbicide Stress | All Samples |
|---|---|---|---|---|---|---|
| 1 | UBC(1.006) | MD(0.478) | MD(0.352) | MD(0.895) | ANI(0.701) | MD(0.941) |
| 2 | MD(1.230) | RIP(0.582) | RIP(0.753) | UBC(0.943) | RIP(0.914) | RIP(0.959) |
| 3 | ANI(1.270) | ANI(0.996) | ADP(0.855) | RIP(0.991) | ADP(1.067) | ADP(1.245) |
| 4 | RIP(1.308) | UBC(1.040) | GADPH(1.190) | ADP(1.080) | ACT(1.103) | UBC(1.323) |
| 5 | CYP(1.427) | GADPH(1.293) | UBC(1.616) | ANI(1.117) | CP(1.268) | ANI(1.339) |
| 6 | ADP(1.481) | CYP(1.353) | CP(1.772) | CP(1.192) | MD(1.298) | CP(1.541) |
| 7 | CP(1.489) | CP(1.551) | ANI(1.779) | GADPH(1.23) | UBC(1.747) | CYP(1.568) |
| 8 | 50S(1.852) | ADP(1.566) | CYP(1.838) | CYP(1.345) | CYP(1.865) | ACT(1.865) |
| 9 | ACT(1.976) | SuS(1.636) | ACT(2.263) | ACT(1.773) | GADPH(2.272) | GADPH(2.304) |
| 10 | H3(2.198) | ACT(1.797) | H3(2.286) | HSP70(1.814) | HSP70(2.701) | 50S(2.416) |
| 11 | SuS(3.178) | 50S(2.149) | 50S(2.998) | H3(1.991) | 50S(2.741) | H3(2.468) |
| 12 | GADPH(3.555) | H3(2.294) | SuS(3.749) | 50S(2.121) | SuS(2.851) | HSP70(2.527) |
| 13 | HSP70(4.490) | HSP70(2.362) | HSP70(5.39) | SuS(2.405) | H3(3.138) | SuS(2.839) |
| Method | Stability (High–Low) | ||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | |
| Cold stress | |||||||||||||
| BestKeeper | UBC | RIP | CP | H3 | ADP | MD | ACT | CYP | ANI | HSP70 | 50S | GADPH | SuS |
| NormFinder | UBC | MD | ANI | RIP | CYP | ADP | CP | 50S | ACT | H3 | SuS | GADPH | HSP70 |
| Genorm | UBC/MD | ANI | ADP | CP | RIP | CYP | ACT | H3 | 50S | HSP70 | SuS | GADPH | |
| RefFinder | UBC | MD | RIP | ANI | ADP | CP | CYP | H3 | ACT | 50S | SuS | GADPH | HSP70 |
| Salt stress | |||||||||||||
| BestKeeper | MD | GADPH | ADP | UBC | RIP | CP | HSP70 | CYP | ANI | SuS | ACT | 50S | H3 |
| NormFinder | MD | RIP | ANI | UBC | GADPH | CYP | CP | ADP | SuS | ACT | 50S | H3 | HSP70 |
| Genorm | RIP/MD | ANI | UBC | ADP | GADPH | CYP | SuS | ACT | CP | H3 | HSP70 | 50S | |
| RefFinder | MD | RIP | UBC | GADPH | ANI | ADP | CYP | CP | SuS | ACT | HSP70 | 50S | H3 |
| Drought stress | |||||||||||||
| BestKeeper | GADPH | MD | ADP | RIP | CYP | CP | UBC | HSP70 | 50S | ANI | ACT | H3 | SuS |
| NormFinder | MD | RIP | ADP | GADPH | UBC | CP | ANI | CYP | ACT | H3 | 50S | SuS | HSP70 |
| Genorm | RIP/MD | CP | CYP | HSP70 | ADP | GADPH | UBC | ANI | ACT | H3 | 50S | SuS | |
| RefFinder | MD | RIP | GADPH | ADP | UBC | CP | CYP | ANI | H3 | ACT | 50S | SuS | HSP70 |
| Method | Stability (High–Low) | ||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | |
| Al stress | |||||||||||||
| BestKeeper | RIP | GADPH | MD | UBC | ADP | ANI | H3 | CP | CYP | ACT | HSP70 | SuS | 50S |
| NormFinder | MD | UBC | RIP | ADP | ANI | CP | GADPH | CYP | ACT | HSP70 | H3 | 50S | SuS |
| Genorm | RIP/MD | ADP | UBC | ANI | GADPH | H3 | CP | CYP | HSP70 | ACT | SuS | 50S | |
| RefFinder | UBC | MD | RIP | GADPH | ADP | ANI | CP | CYP | H3 | ACT | HSP70 | 50S | SuS |
| Herbicide stress | |||||||||||||
| BestKeeper | RIP | ADP | MD | UBC | ACT | ANI | CP | GADPH | CYP | HSP70 | 50S | SuS | H3 |
| NormFinder | ANI | RIP | ADP | ACT | CP | MD | UBC | CYP | GADPH | HSP70 | 50S | SuS | H3 |
| Genorm | ADP/RIP | ANI | ACT | CP | MD | UBC | CYP | GADPH | HSP70 | SuS | H3 | 50S | |
| RefFinder | RIP | ANI | ADP | ACT | MD | UBC | CP | GADPH | CYP | HSP70 | 50S | SuS | H3 |
| All samples | |||||||||||||
| BestKeeper | MD | ADP | RIP | UBC | GADPH | CP | ANI | CYP | ACT | HSP70 | 50S | H3 | SuS |
| NormFinder | MD | RIP | ADP | UBC | ANI | CP | CYP | ACT | GADPH | 50S | H3 | HSP70 | SuS |
| Genorm | ADP/RIP | MD | UBC | ANI | CP | CYP | ACT | H3 | HSP70 | GADPH | SuS | 50S | |
| RefFinder | ANI | RIP | UBC | ADP | CYP | ACT | GADPH | MD | SuS | H3 | 50S | CP | HSP70 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Shu, X.; Su, X.; Xiong, Y.; Xiong, Y.; Chen, M.; Tong, Q.; Ma, X.; Zhang, J.; Zhao, J. Selection of Suitable Reference Genes for RT-qPCR Gene Expression Analysis in Centipedegrass under Different Abiotic Stress. Genes 2023, 14, 1874. https://doi.org/10.3390/genes14101874
Wang X, Shu X, Su X, Xiong Y, Xiong Y, Chen M, Tong Q, Ma X, Zhang J, Zhao J. Selection of Suitable Reference Genes for RT-qPCR Gene Expression Analysis in Centipedegrass under Different Abiotic Stress. Genes. 2023; 14(10):1874. https://doi.org/10.3390/genes14101874
Chicago/Turabian StyleWang, Xiaoyun, Xin Shu, Xiaoli Su, Yanli Xiong, Yi Xiong, Minli Chen, Qi Tong, Xiao Ma, Jianbo Zhang, and Junming Zhao. 2023. "Selection of Suitable Reference Genes for RT-qPCR Gene Expression Analysis in Centipedegrass under Different Abiotic Stress" Genes 14, no. 10: 1874. https://doi.org/10.3390/genes14101874